1. Introduction
The disruptive potential of cold plasma for the safety and quality enhancement of a wide range of food products, with an emerging focus on the earlier stages of food production chains including agricultural pre-treatments for seeds, water or soil, is under increased investigation. However, attention is required on the effects on food components integrity, potential for knock-on toxicity and the safety of this emerging technology for application to different food commodities. Research performed in the context of biomedical applications of plasma has demonstrated the cytotoxic properties of plasma-treated solutions (both water and more complex media, containing carbohydrates, lipids and/or proteins) in mammalian cell models [
1,
2]. The cytotoxic and mutagenic effects of plasma-treated, protein-based solutions and produce-based solutions have also been reported [
3,
4,
5]. For instance, water, phosphate-buffered saline and phosphate buffer solution exposed to plasma for 10 min resulted in cell death in more than 50% of cells [
1]. Foetal bovine serum treated with plasma for 5 or 10 min caused not only strong growth inhibition of HeLa cells, but also exhibited increased mutagenic potential [
3].
Chemical reactions between food components and plasma reactive species are responsible for food surface modifications and functionality changes, however, knowledge of the direct interactions between plasma-generated ROS/RNS and components, such as carbohydrates, complex lipids, and proteins, is limited, especially regarding the impact of potentially formed toxic products [
6,
7]. There is a need to evaluate the safety of a wide range of ACP-treated food matrices in order to demonstrate that their consumption is non-hazardous and for regulatory considerations.
The objective of this study was to evaluate the safety profile of ACP-treated cereal grain models using the well-established mammalian cell model CHO-K1. The cytotoxicity of two types of plasma-exposed wheat models was determined, including a liquid model (wheat model medium (WMM)) and a solid model (wheat grain extract, WGE-prepared using wheat grains exposed to plasma). Additionally, the safety of using cold plasma for grains processing and ingestion, i.e., the residual toxicity of plasma treatment, was assessed using an insect feeding trial using the invertebrate model Tribolium castaneum. The beetles were fed with flour produced from plasma-treated grains.
2. Materials and Methods
2.1. Wheat Grains
Organic wheat (origin: United Kingdom) grains were purchased from a local supermarket.
2.2. Preparation of Wheat Model Medium (WMM)
Wheat grains (10 g) were milled using a Nutribullet blender (model NB-101B) for 10 s to obtain fine and homogeneous flour. Wheat model medium (WMM) was prepared as for the method described by Charalampopoulos et al. [
8] with minor modifications. The resulting wheat flour (50 g) was mixed with 450 mL of tap water and centrifuged at 6000
g for 30 min at room temperature. The supernatant was sterilized at 121 °C for 45 min. The procedures of centrifugation and sterilization were repeated four times to minimise the presence of sediments caused by sterilization in the final media. Prepared wheat model media were stored at 4 °C before use. Three independent ACP treatment experiments were conducted.
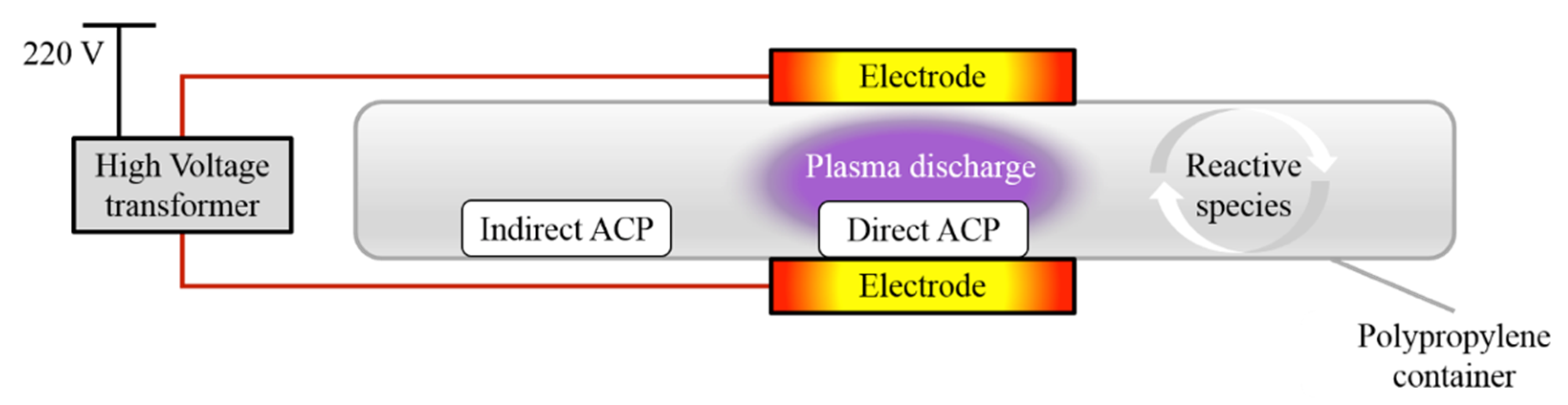
2.3. ACP System Set Up
The ACP system used in this study was a dielectric barrier discharge system with a maximum high voltage output of 120 kV at 50 Hz. The system has been previously described by [
9] and a schematic diagram is presented in
Figure 1. The distance between the two aluminium electrodes was equal to the height of the polypropylene container (20 mm), used as a sample holder and a dielectric barrier. For direct treatment, the distance between the top electrode and the sample was approximately 15 mm. For indirect treatment, the distance between the samples and centre of the electrodes varied from 120 to 160 mm owing to the sample distribution in the petri dish.
The plasma working gas was atmospheric air.
2.4. Plasma Treatment
Samples of wheat grains (10 g, evenly distributed single layer of grains, with thickness equal to the thickness of single grain ~2–3 mm) or WMM (10 mL, evenly distributed) in petri dish were exposed to 80 kV
RMS ACP for 5 or 20 min with both direct and indirect plasma treatments conducted simultaneously. For direct exposure, samples were placed between the electrodes, i.e., within the plasma discharge, while, for indirect plasma treatment, samples were placed in the corner of the container, i.e., outside plasma discharge (
Figure 1). Before treatment, each container was sealed with a high-barrier polypropylene bag (catalog number B2630; Cryovac, Dublin, Ireland). After treatment, the samples were stored at 15 °C for 24 h. Three independent ACP treatment experiments were conducted.
2.5. Preparation of WGE
The ACP-treated wheat grains (10 g) were milled using a Nutribullet blender for 10 s to obtain fine and homogeneous flour. This flour (2.2 g) was mixed with tap water (20 mL) (proportions as per WMM preparation procedure,
Section 2.2.), shaken for 60 min at 200 rpm and centrifuged at 6000×
g for 30 min. The supernatant, i.e., wheat grain extract (WGE), was filter-sterilised and used for cytotoxicity and further analyses.
Untreated and ACP-treated wheat model medium (WMM) and wheat grain extract (WGE) prepared using either untreated or ACP-treated wheat grains, were used for safety evaluations.
2.6. Cytotoxicity Assay
The cytotoxicity assay was performed according to the protocol by Boehm et al. [
11] using Chinese hamster ovary (CHO-K1) cell line obtained from culture stocks of TU Dublin or University College Dublin. Cytotoxicity of the treated medium was measured by assessing adherent cell mass using crystal violet assay. Cells were cultured in Dulbecco’s modified Eagle’s medium/Ham’s F12 Nutrient Mixture (DMEM/F12) supplemented with 2 mM L-glutamine and 10% foetal bovine serum (FBS), supplemented with 2%, 5% and 10% of either WMM or WGE (
v/
v). CHO-K1 cells were grown at 37 °C and 5% CO
2 in a humidified incubator. The growth of cells seeded at 2.5 × 10
4 cells/mL after 3 days of culture was assessed by crystal violet staining according to the protocol described by Tsoukou et al. [
1], with minor modifications. Briefly, the culture supernatant was removed and attached cells were fixed with methanol (70%) for 1 min and followed by staining with crystal violet solution (0.2%) for 10 min. Excess stain was rinsed off, plates were air-dried, and the crystal violet bound to the adherent cells was re-solubilized using acetic acid (10%). The absorbance was measured at 560 nm on a spectrophotometric microplate reader (BioTek, Winooski, VT, USA). Cell growth was expressed as a percentage of control cells grown in medium supplemented with untreated WMM or WGE. To assess the effect of untreated WMM or WGE on CHO-K1 cells, cell growth was also compared to cells grown in medium supplemented with sterile, untreated PBS. As stated above, three independent ACP treatment experiments were conducted, and for each condition the sample absorbance was read in triplicate (three individual wells of the 96-well plate), resulting in total number of nine per condition to be evaluated (
n = 9).
2.7. Chemical Properties of WMM and WGE
Hydrogen peroxide (H
2O
2) concentration measurement was based on the oxidation of potassium iodide (KI; Sigma Aldrich, Arklow, Ireland) to iodine and spectrophotometric measurement at 390 nm [
3]. Briefly, 50 μL of phosphate buffer and 100 μL of 1 M KI solution were added to 50 μL of sample. After 30 min incubation, absorbance was read on a spectrophotometric plate reader at 390 nm. Each sample (of three independent experiments) absorbance was read in triplicate (three individual wells), resulting in total number of nine per each condition to be evaluated (
n = 9).
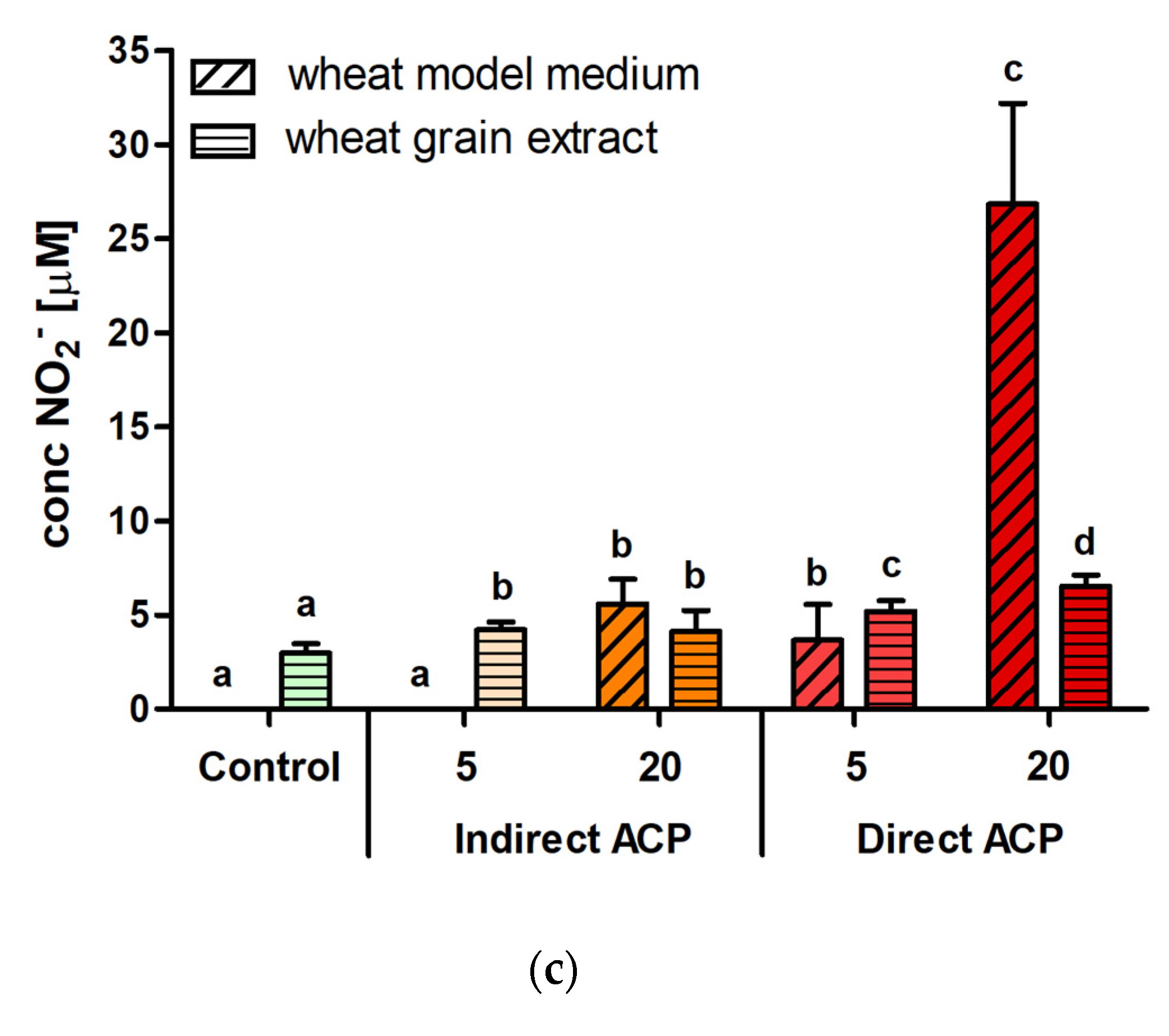
The concentration of nitrite (NO
2−) was determined using Griess reagent (Sigma Aldrich, Ireland) according to the procedure described by Lu et al. [
12]. For 100 μL of sample, 100 μL of Griess reagent was added, and absorbance was read at 548 nm after 30 min incubation. Each sample (of three independent experiments) absorbance was read in triplicate (three individual wells) resulting in total number of nine per each condition to be evaluated (
n = 9).
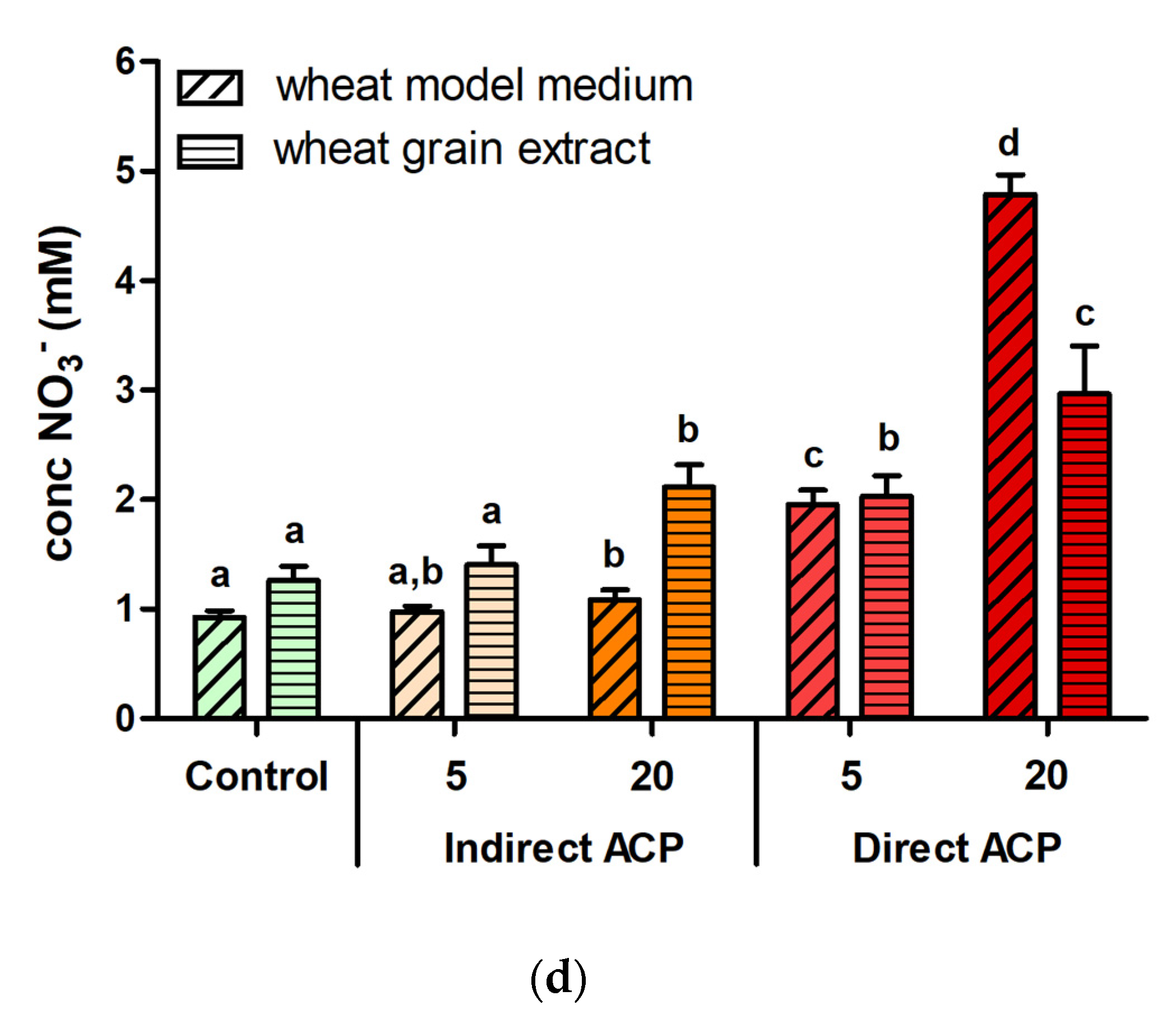
The concentration of nitrate (NO3−) was determined photometrically by 2,6-dimethyl phenol (DMP) using the Spectroquant nitrate assay kit (Merck, Germany). Samples were pre-treated with sulfamic acid to eliminate nitrite interference. Each sample’s (of three independent experiments) absorbance was read in duplicate (two individual wells), resulting in a total number of six per each condition to be evaluated (n = 6).
A standard curve of known H2O2 ((Perhydrol®) for analysis EMSURE® ISO), NO2− (sodium nitrite, Sigma Aldrich, Arklow, Ireland) and NO3− (sodium nitrate, Sigma Aldrich, Arklow, Ireland) concentrations was included on each assay plate and used to convert absorbance into the concentration values. Each sample’s (of three independent experiments) absorbance was read in duplicate (two individual wells), resulting in total number of six per each condition to be evaluated (n = 6).
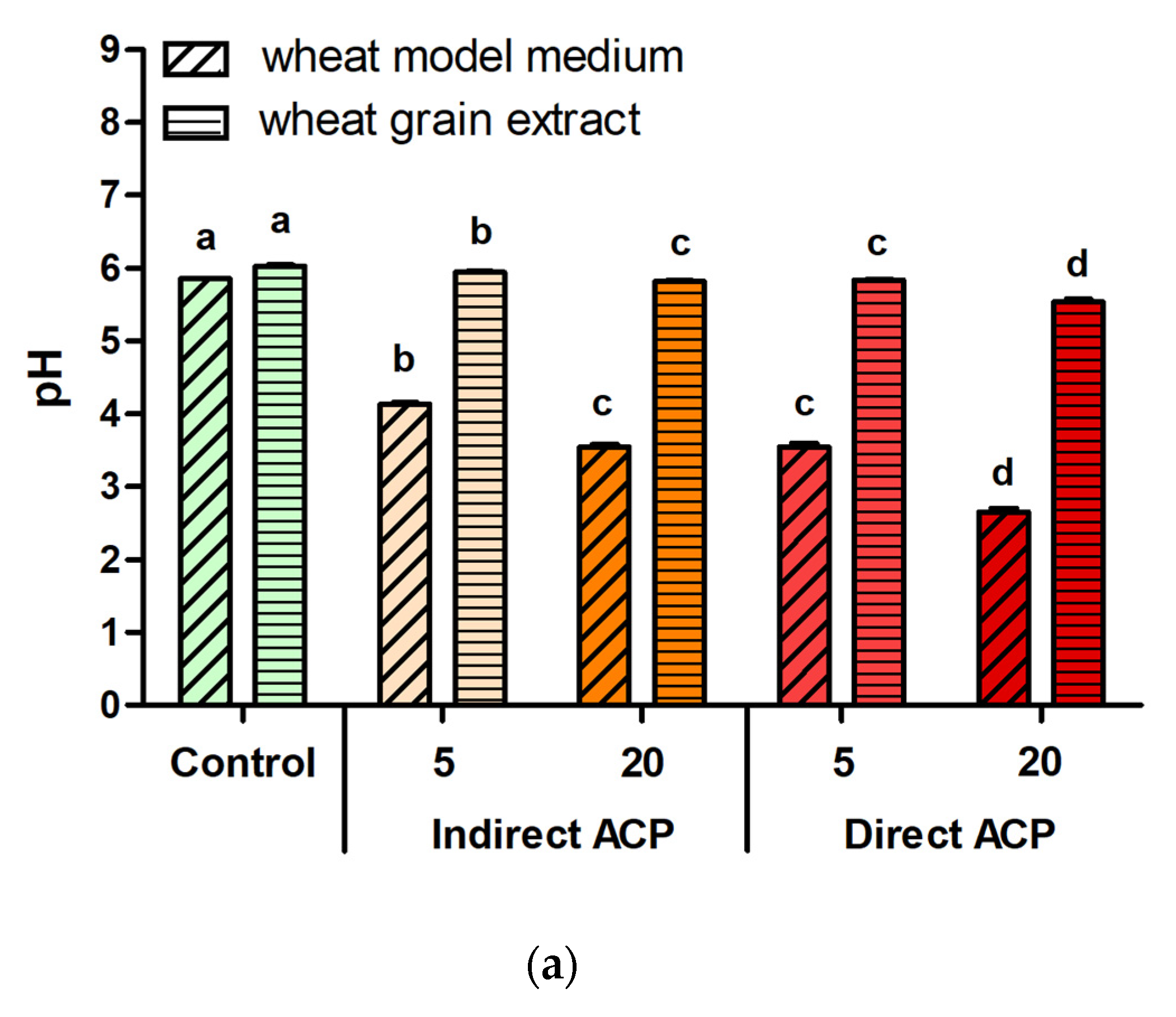
The pH was determined by potentiometric measurement of the suspensions at 25 °C using an Orion pH meter (model 420A). As stated above, three independent ACP treatment experiments were conducted and, for each condition, the sample’s pH was measured once (n = 3).
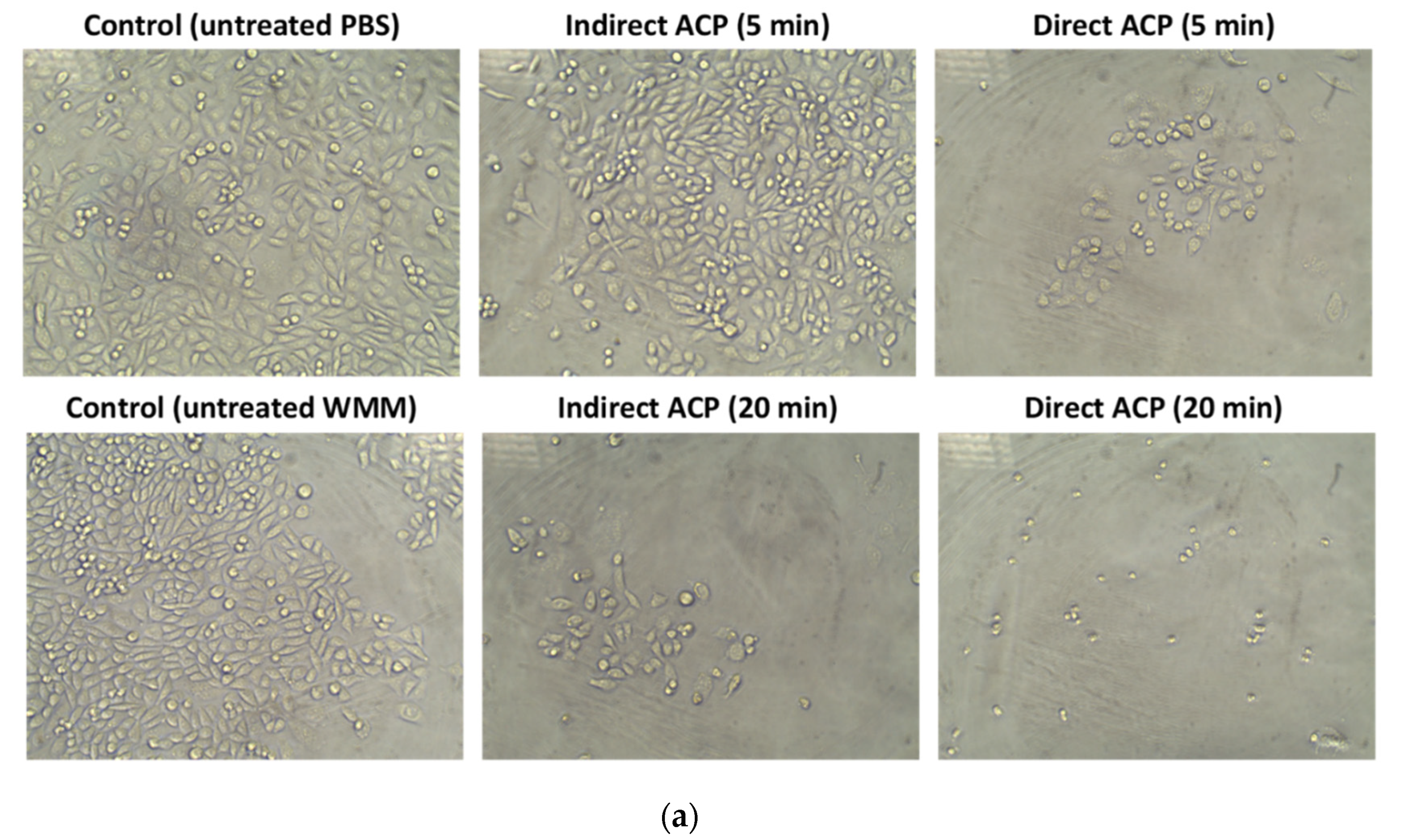
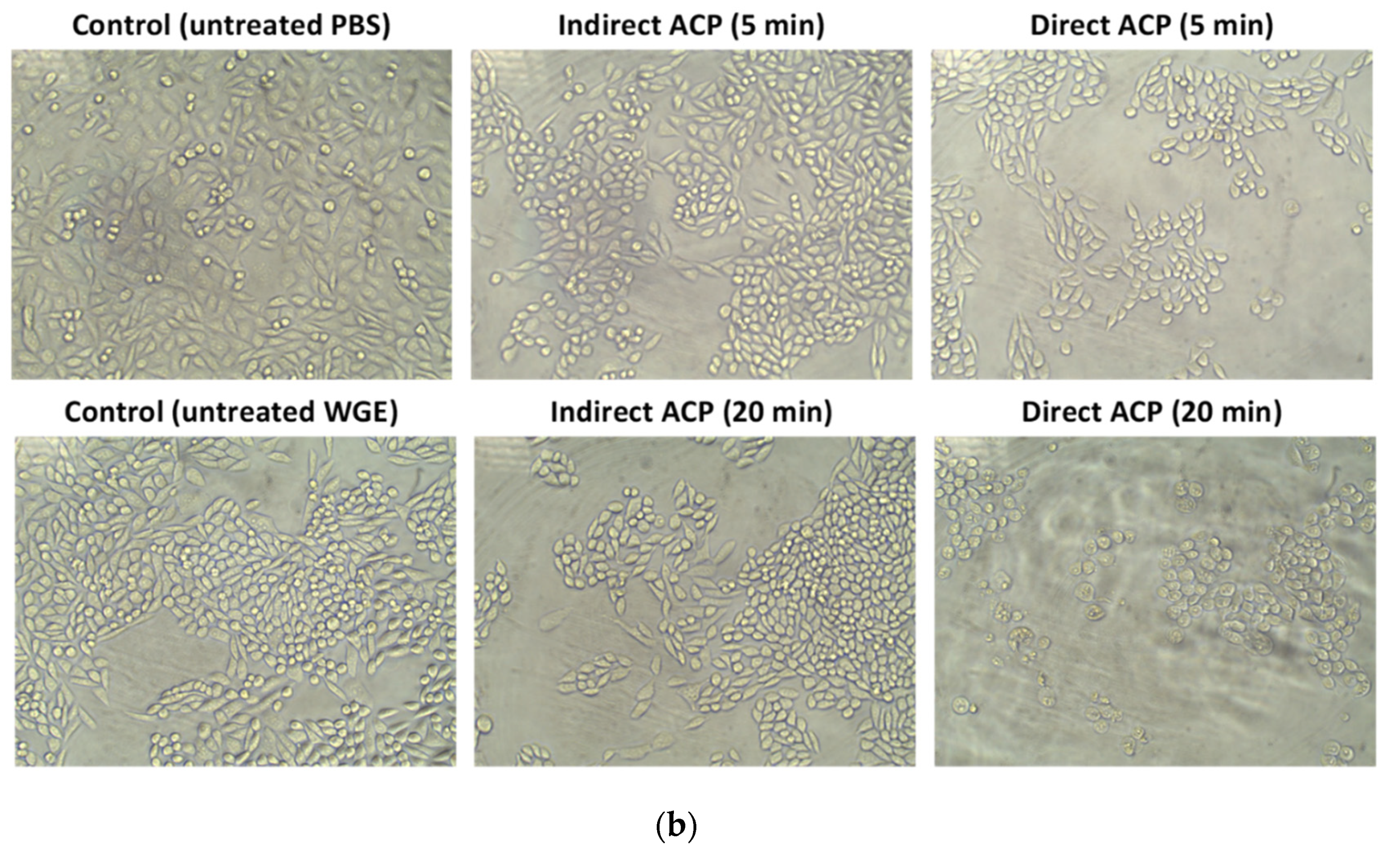
2.8. Optical Microscopy
Microscopic analysis of cells was performed using inverted light microscope (Optika, Pontarenica, Italy, Tucsen Photonics, Co., Ltd.) and a total magnification of 200 X.
2.9. Insect Feeding Trial
Culture rearing conditions: Tribolium castaneum obtained from Blades-Biological (Kent, UK) was reared in plastic containers on 95% organic wheat flour supplemented with 5% brewer’s yeast mixture at 34 °C in the dark. Whole wheat grains (10 g) exposed to direct and indirect plasma treatment for 20 min and post-treatment storage for 24 h at 15 °C were used to prepare flour as food for insects. Three independent ACP treatment experiments were conducted. Control flour feed was produced from grains, which were untreated and stored for 24 h at 15 °C. Immediately after the treatment and post-treatment storage, each sample was ground for 10 s, five times to obtain fine particles using a Nutribullet blender. The resultant flour was sieved to produce particle size of <800 µm, which was then used as the rearing medium for insects. Each sample was divided in two equal aliquots of ~4.5 g resulting in six samples to be evaluated per each condition (n = 6). A 4.5 g aliquot of each sample (either ACP-treated or untreated control) was used to feed T. castaneum-second instar larvae (2–3 days old or 7 days from oviposition, 25 larvae per sample). The samples were incubated at 34 °C in the dark until the emergence of adults in all samples was observed (5 weeks from the time of ACP treatment and inoculation). The mortality of insects was calculated as the number of dead insects/total number of insects for each trial × 100. The weight of adult insects (measured in a pool of 20 insects) for each experimental group was recorded. Each experiment was performed in duplicate and repeated three times (n = 6).
The magnitude of lipid peroxidation, a non-enzymatic oxidative stress marker, was determined by measuring concentration of malondialdehyde (MDA, a by-product of lipid peroxidation, which is thiobarbituric acid reactive substance) according to the procedure described by Wang et al. [
13] with minor modifications. Briefly, twenty living adult insects were homogenized by crushing the beetles with 1 mL of ice-cold phosphate-buffered saline (PBS). The homogenates were centrifuged twice for 10 min at 10,000 rpm. To 0.2 mL of supernatant 0.2 mL of 30% tri-chloroacetic acid (Sigma-Aldrich, Arklow, Ireland) was added. The mixture was vortexed and centrifuged at 10,000 rpm for 10 min at 4 °C. After centrifugation, 0.3 mL of supernatant was mixed with 0.3 mL of 0.8% thiobarbituric acid (Sigma-Aldrich, Arklow, Ireland). The resulting suspension was incubated at 98 °C for 60 min, cooled on ice for 5 min and centrifuged at 4000
g for 1 min. The concentration of MDA in test solutions was determined by measuring absorbance in duplicate (two individual wells per sample) at 532 nm, and comparing to a standard curve of known MDA (Sigma-Aldrich, Arklow, Ireland) concentrations. The MDA level was expressed as μmol per g of weight of insects (
n = 6).
2.10. Statistical Analysis
Statistical analysis was performed using IBM SPSS statistics 21 Software (SPSS Inc., Chicago, IL, USA). Significant differences between samples were determined using one-way analysis of variance (ANOVA), and Fisher’s Least Significant Difference (LSD) at the 0.05 level.
4. Discussion
Assessing the potential for induced modifications or toxicity of ACP-treated food matrices is essential to ensure safe process development and general application in the food industry.
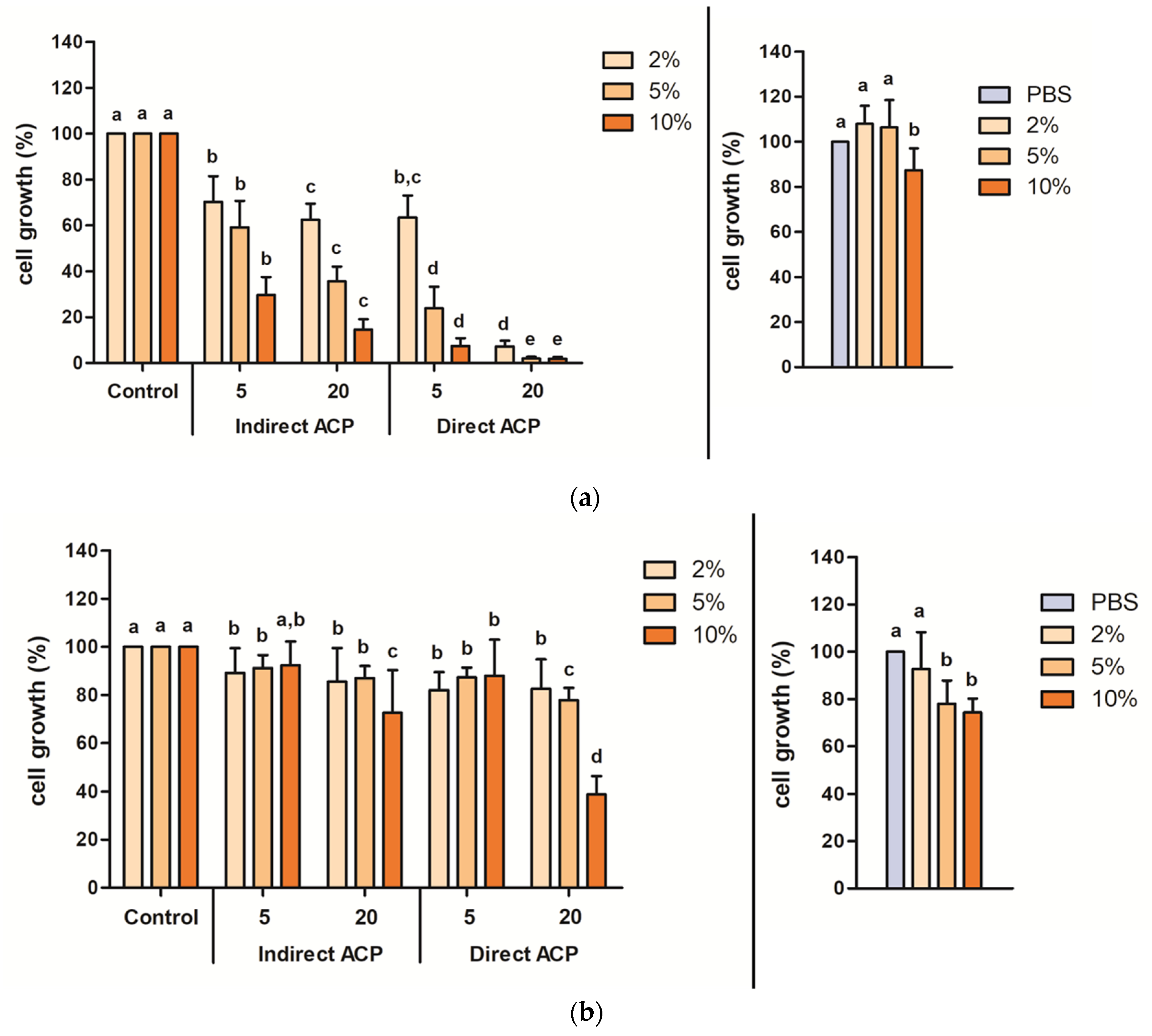
For untreated cereal models, supplementation with the highest tested concentration, i.e., 10% (
v/
v), of WMM and WGE resulted in a significant decrease in CHO-K1 cell growth by 12.6% for WMM and 25.7% for WGE. In a safety evaluation of a fresh produce model—a lettuce broth—the cell growth of CHO-K1 cells was unaffected when supplementing the cell culture media with up to 10%
v/
v of undiluted, untreated lettuce broth, showing that, unlike the grain samples, the lettuce broth itself did not adversely affect the cells [
5]. The growth reduction caused by untreated WMM and WGE could be the result of certain grain components having growth inhibitory effects or interfering with the cells’ adhesion to the culture plate surface. The negative effect of WMM and WGE on the cells’ growth could also suggest that the approach used in this study may overestimate the toxicity for certain food models. Allegra et al. [
15] stated that cell culture systems may overestimate the toxicity of some chemicals, especially low-toxicity chemicals, and suggested the use of insect models, such as
Galleria mellonella (wax moth) as a potential alternative, which, in their study was found to be a reliable predictor for low toxicity chemicals.
The cytotoxicity and changes in chemistry of the ACP-treated samples were hugely dependent on the sample type (liquid vs. solid) and the effects were more pronounced in samples treated in a liquid form. Visually observable changes were found immediately after ACP treatment in WMM samples (change in colour from yellowish to transparent), whereas no change in the colour was noted in WGE (
Figure S1). ACP-treated WMM (treated as a liquid) had a much stronger toxic effect on cells than WGE (treated as solid wheat grains). WMM and WGE could differ in the composition and probably also in the concentration of major components such as protein and carbohydrates due to the difference in the media preparation where thermal and centrifugation steps were not included for the preparation of WGE. The toxic effect of WMM and WGE was affected by both supplementation levels in the cell culture media and ACP treatment parameters, exhibiting the highest decrease in cells’ growth at 10% supplementation with samples subjected to 20-min direct plasma exposure for both WMM (decrease by 98.2%) and WGE (decrease by 61.2%). Using the same DBD–ACP setup, Heslin et al. [
5] evaluated the safety of ACP-treated lettuce broth using in vitro and in vivo toxicity models-CHO-K1 cells and
Galleria mellonella larvae, respectively. A discrepancy between the results for mammalian cell model and larvae were reported. After 1 min of ACP treatment, the cell growth of CHO-K1 cells in the presence of 10%
v/
v plasma-treated lettuce broth decreased to 74% compared to the untreated control. After the initial decrease, cell growth remained stable at 68% compared to the untreated control when supplemented with lettuce broth ACP-treated for up to 10 min. Although low cytotoxic effects were detected in vitro, a strong response of the
Galleria larvae (in vivo model) to injection with plasma-treated lettuce broth was observed for 5-minute treated broth, with less than 10% larvae survival. These results indicate the importance of assessing ACP-treated products in multiple toxicity models and, ultimately, in the context of ingestion systems. The higher toxicity in vitro of cereal-based models used in this study as compared to lettuce broth could be attributed to the different medium complexity and composition [
16] with potential for both carbohydrate, lipid and protein fractions of the wheat flours to undergo structural changes as a result of degradation or oxidation [
17,
18]. ACP-treated WMM and WGE resulted in cytotoxic effects on mammalian cells. Plasma treatment of aqueous solutions generates reactive species in the liquid, including hydrogen peroxide, nitrates, and nitrites, which may react to form further cell toxic compounds, such as peroxynitrite/peroxynitrous acid [
11]. Hydrogen peroxide (H
2O
2) can induce DNA damage, cause cell cycle arrest and trigger apoptosis. Its toxicity is determined largely by the cellular anti-oxidant status and ability to detoxify H
2O
2 through catalase [
11]. Although there was a plasma treatment time-dependent increase in H
2O
2 in lettuce broth, after 10-minute treatment it was still less than 80 µM, potentially due to high concentrations of antioxidants such as vitamins and polyphenols in the lettuce [
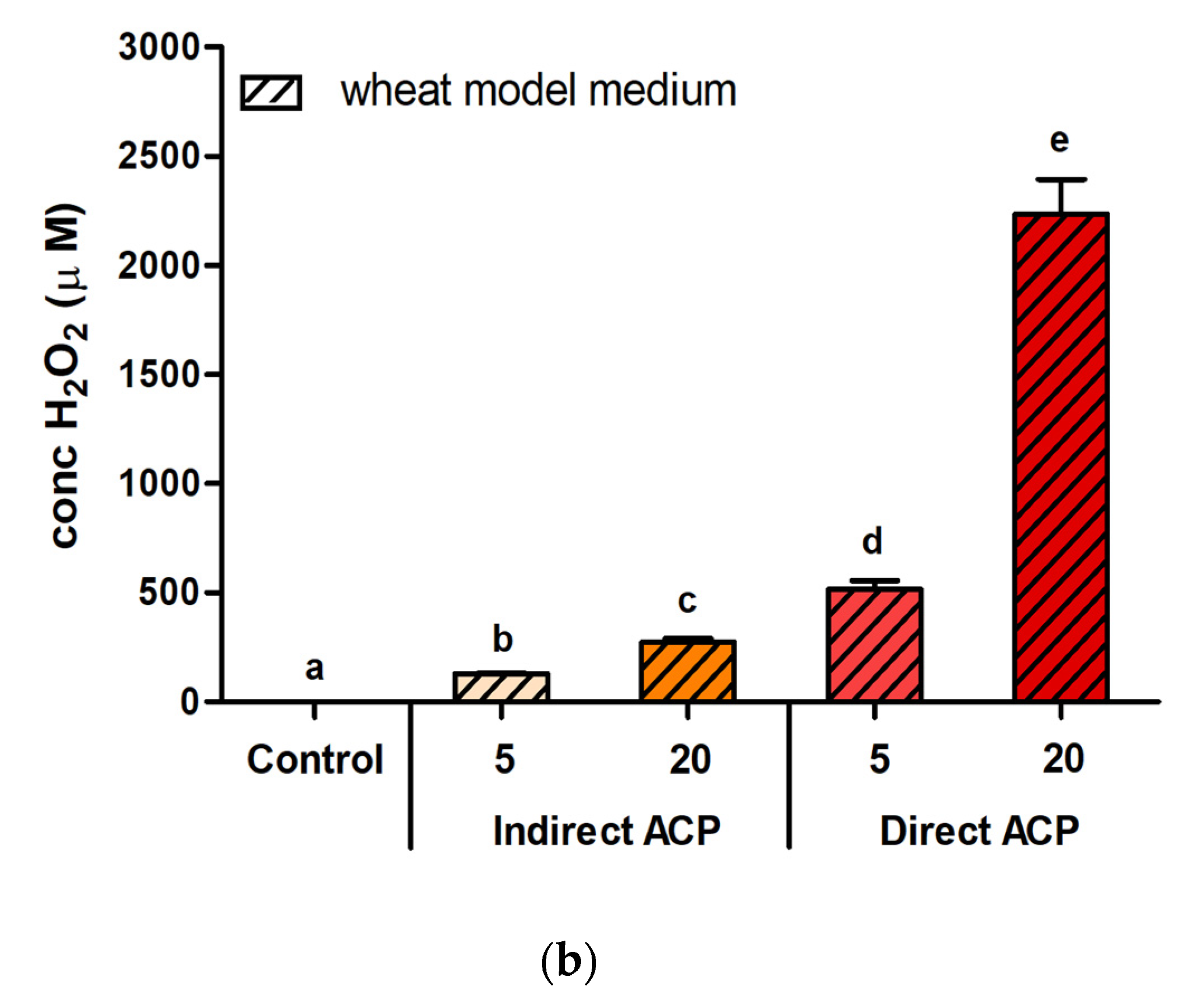
5], whereas the values of H
2O
2 in WMM reached levels approximately 25× higher (2234.0 µM after 20-min direct treatment). Peroxide levels in treated PBS (buffer without organic molecules) were determined to be in the range of 200–400 μM with a treatment duration of 5–10 min, with nitrite and nitrate concentrations around 50 µM and 500 µM, respectively [
1,
3].
Hydrogen peroxide, however, was not the only cytotoxic factor of the tested cereal models. There was no H
2O
2 detected in ACP-treated WGE samples, yet they also showed cytotoxic effects, which could be possibly attributed to other changes observed in chemical composition post-plasma treatment, i.e., decrease in pH and increase in nitrites and nitrates. However, as pointed out by Boehm et al. [
11], the addition of plasma-activated water (PAW) up to concentrations of a total of 80% (
v/
v) did not result in an acidification of DMEM/F12 medium, which is buffered both with HEPES and bicarbonate and contains 10% FBS, and therefore the pH-related effects of PAW and other plasma-treated solutions when diluted in DMEM can be discounted. While medium components such as pyruvate or BSA may also act as scavengers of H
2O
2, this would further reduce the concentrations of H
2O
2 cells were exposed to and support a role for other cytotoxic factors. The final concentrations of nitrite and nitrate resulting from supplementation with WMM did not exceed 5 µM and 0.5 mM, respectively, while up to 1.2 mM nitrite or nitrate were well-tolerated in the same cell model [
11]. In addition to these effects, significant increases in the concentrations of malondialdehyde (MDA) were also observed for ACP-treated wheat grains [
18]. MDA, a product of lipid peroxidation, is known for its cytotoxic properties. MDA is able to form adducts with proteins that lead to their inactivation [
19], as well as react with DNA to form adducts with deoxyguanosine and deoxyadenosine [
20].
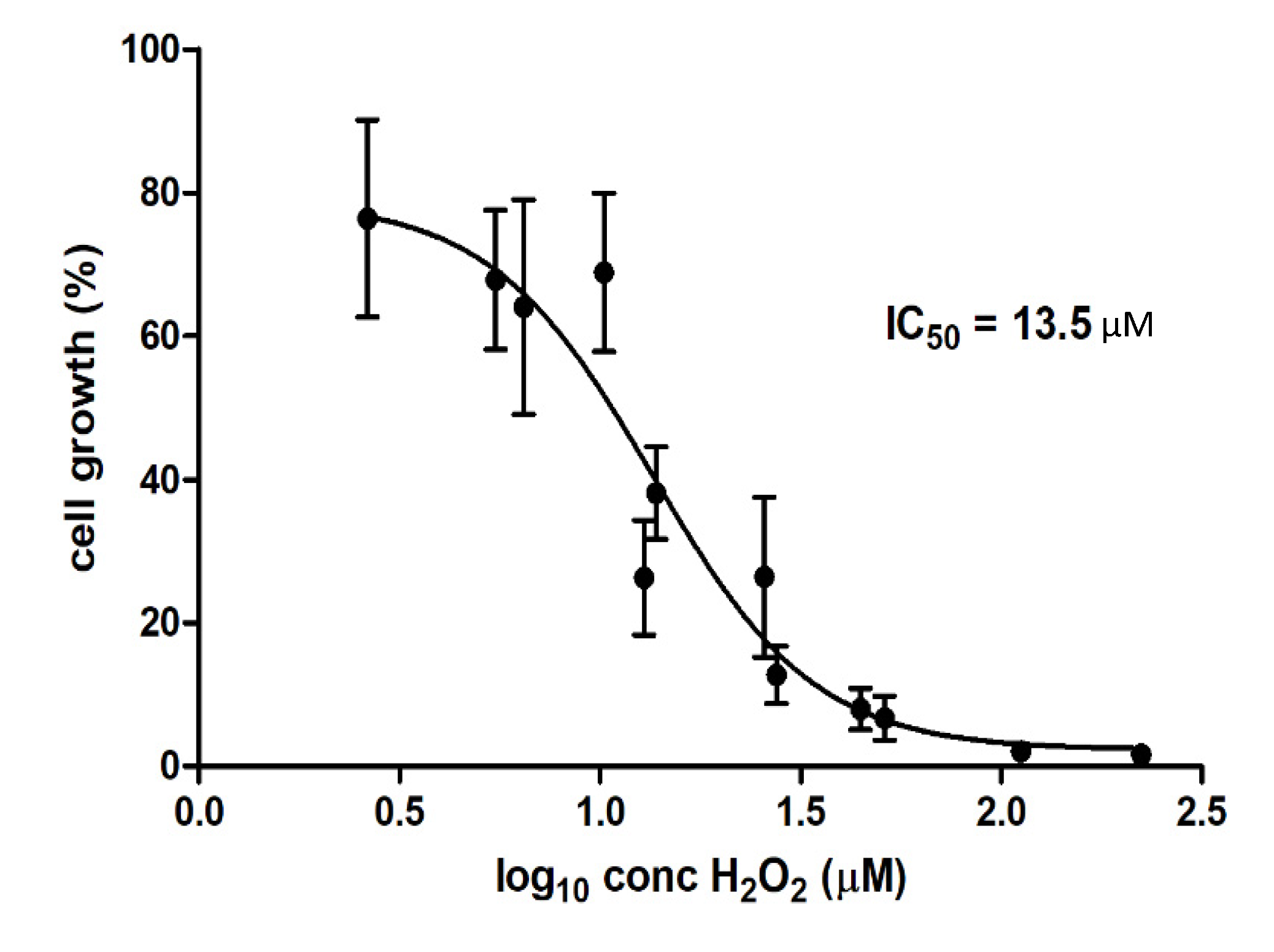
In this study, the IC50 of 13.5 µM for H
2O
2 was obtained for WMM samples (H
2O
2 was not present in WGE). Boehm et al. [
11] reported that the IC50 values of H
2O
2 for CHO-K1 cells for PAW were much lower than those obtained through supplementation of pure H
2O
2 into the cell medium, obtaining IC50 of 105 µM and 225 µM for PAW and pure H
2O
2, respectively. IC50 obtained for WMM was therefore almost 10-fold lower than for PAW and more than 10-fold lower than for pure H
2O
2, confirming that although cytotoxic properties correlate with H
2O
2 concentrations in plasma-treated solutions, other factors contributing to plasma-induced cytotoxicity are involved, and may act synergistically with H
2O
2.
Beetles are the largest insect order, with their representative,
Tribolium castaneum Herbst (Coleoptera: Tenebrionidae) the red flour beetle, emerging as a new potential candidate for a biological model organism and bio-indicator in environmental and biomedical studies due to the similarity of their signalling pathways, basic immune mechanisms, energy metabolism and structural components with mammals [
21].
T. castaneum is also a global pest of stored food products including wheat grains [
22], therefore it is widely employed in studies testing novel insecticides of physical, chemical and biological origin [
23,
24,
25]. This study evaluated the residual toxicity of plasma treatment as a potential method for biological control of stored grains using
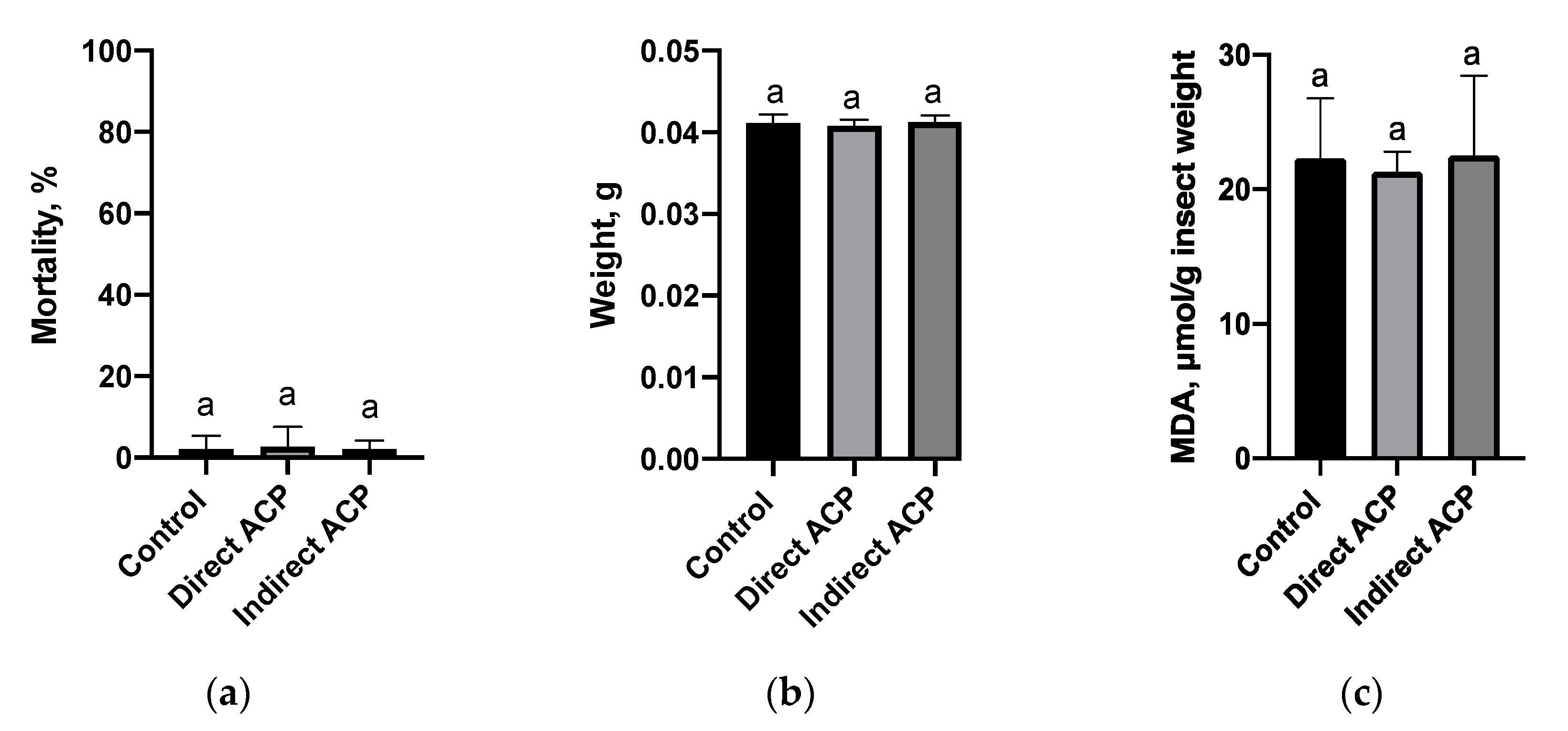
T. castaneum as a model organism. In contrast to the in vitro cytotoxicity assay, flour produced from plasma-treated grains did not affect the viability and weight of insects regardless of the mode of plasma exposure. This can be due to the lower sensitivity of
Tribolium as a model to detect observed chemical fluctuations in flour. It is known that in insects under the influence of environmental and physiological stressors, an enhanced production of ROS can occur. In the cell membrane, polyunsaturated fatty acids are the primary target for ROS stimulating the process of lipid peroxidation and MDA, as one of the secondary products of lipid peroxidation, is used to evaluate the extent of oxidative stress in insects [
26,
27]. Lipid peroxidation level, measured in insects fed on flour generated from directly treated grains, was slightly lower than in the control population or remained unchanged, as noted in insects fed on the flour of indirectly treated grains, suggesting that antioxidant processes were sufficient, or an absence of oxidative stress, and thus grain toxicity to insects after plasma exposure. It is also possible that any lipid peroxidation would occur during the first hours, which is later repaired over the 5 weeks of incubation. A slight reduction in the MDA content could also be associated with antioxidant responses and the change in the level of proteins or peptides known to decrease oxidative stress in insects. For example, caffeine supplementation of food markedly increased total thiol and non-protein thiol glutathione levels in cockroaches, which was associated with significant reduction of lipid peroxidation [
28]. In contrast, Holmstrup et al. [
29] observed a decrease in lipid peroxidation in
T. castanaeum, which was related to oxidative stress, indicating the general depression of insects’ metabolism. Although insects’ feeding, as a part of the preliminary “warning” study, indicated no negative impacts, further investigations are warranted to examine the immediate effect of plasma-treated components on MDA content and to study the molecular and physiological responses, enzymes and non-enzymatic systems in this and other model organisms to understand if ACP treatment mediates toxic effects to ensure safe application of this technology across agricultural and food sectors.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









