
Synthetic Progestins in Waste and Surface Waters: Concentrations, Impacts and Ecological Risk



1
Laboratory of Histology and Embryology, Department of Microscopy, Institute of Biomedical Sciences Abel Salazar (ICBAS), University of Porto (U.Porto), 4050-313 Porto, Portugal
2
Histomorphology, Physiopathology, and Applied Toxicology Team, Interdisciplinary Centre of Marine and Environmental Research (CIIMAR), University of Porto (U.Porto), 4450-208 Porto, Portugal
*
Author to whom correspondence should be addressed.
Toxics 2022, 10(4), 163; https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040163
Submission received: 28 December 2021
/
Revised: 22 March 2022
/
Accepted: 23 March 2022
/
Published: 29 March 2022
(This article belongs to the Special Issue Occurrence, Effects and Ecological Risk Assessment of Emerging Contaminants in Rivers)
Abstract
:Synthetic progestins (PGs) are a large family of hormones used in continuously growing amounts in human and animal contraception and medicinal therapies. Because wastewater treatment plants (WWTPs) are unable to eradicate PGs after excretion, they are discharged into aquatic systems, where they can also be regenerated from conjugated PG metabolites. This review summarises the concentrations of 12 PGs in waters from 2015 to 2021. The selected PGs were considered of particular interest due to their wide use, activity, and hormonal derivation (from testosterone, progesterone, and spirolactone). We concluded that PGs had been analysed in WWTPs influents and effluents and, to a lesser extent, in other matrices, including surface waters, where their concentrations range from ng/L to a few µg/L. Because of their high affinity for cell hormone receptors, PGs are endocrine disruptor compounds that may alter the reproductive fitness and development of biota. This review focused on their biological effects in fish, which are the most used aquatic model organisms to qualify the impacts of PGs, highlighting the risks that environmental concentrations pose to their health, fecundity, and fertility. It is concluded that PGs research should be expanded because of the still limited data on their environmental concentrations and effects.
Keywords:
drospirenone; EDCs; estranes; gestagens; gonanes; norpregnanes; pregnanes; risk assessment
1. Introduction
Due to water’s vital importance for life, its availability, quality, and governance have been the subject of intense concern, conflicting interests, and heated debate involving communities, industries, governments, and the media [1]. Nonetheless, past actions and the uncontrolled spread of human activities continue to impact water quality and, more broadly, the vast global aquatic ecosystems [2]. One contemporaneous problem widely recognised as serious for mankind is water pollution, including the increase of the concentrations of compounds defined as micropollutants [3,4].
Water micropollutants are currently mostly anthropogenic in origin and include natural and synthetic compounds that enter the aquatic compartment at concentrations ranging from ng/L to µg/L [5]. Among these contaminants are endocrine disruptor compounds (EDCs). Many of them are active ingredients in hormonal medicines, such as synthetic progestins (also called gestagens, progestogens, or progestins), being of particular concern because they are massively used and designed to act in extremely low dosages in specific cellular receptors [6,7].
In humans, progestins (PGs) are used instead of progesterone in endocrine therapy due to the rapid metabolisation of the latter hormone [8]. These substances are used not only as contraceptives, as PGs can inhibit ovulation and the proliferation of the endometrium, but also to treat and prevent endometrial hyperplasia and carcinoma [9,10], to control dysfunctional uterine bleeding [11], and even to stimulate the appetite of cancer patients [12]. In veterinary medicine and zootechny, these compounds are also used in therapies of cows and mares (viz. in disorders of the reproductive system) and for estrus synchronisation and preparation of donor and receptor animals in cases of embryo transfer [13].
Presently, estimating global PGs usage is challenging due to a lack of data on the issue as well as differences in the types of active pharmaceutical chemicals commonly used by each nation [6]. However, recent data point to this issue as an emerging problem due to the increasing worldwide use of PGs, with the “Progesterone Market” predicting a 13.1% increase over the next five years (https://www.mordorintelligence.com/industry-reports/progesterone-market, accessed on 27 December 2021).
Thus, the widespread use of PGs around the globe and their potential to disrupt non-target organisms in aquatic environments has been considered a hot topic that deserves investigation and timely synthesis reviews [6,7]. This review intends to (a) summarise the properties and the levels of 12 highly prominent synthetic PGs in aquatic environments, covering from 2015 to 2021—i.e., considering the period after the last elaborate reviews on this subject [6,7]—in Section 2 and Section 3; (b) look over the biologic impacts of PGs on fish in Section 4; (c) consider their ability to promote biologic effects similar to those referred in humans in Section 5; and (d) provide hazard coefficients with the objective of prevising possible risks for the aquatic ecosystems exposed to PGs in Section 6.
2. Classification and Properties of the Most Prominent PGs in Aquatic Environments
PGs are typically classified considering their structural derivation and “generation” (Table 1). The latter broadly indicates when PGs were introduced to the market. Thus, to understand the effects of PGs, the most relevant classification system is to group them by structure based on the steroid molecule from which they were created; i.e., testosterone, progesterone, and spironolactone [14].
Most of the older PGs were designed during the 1960–1970s and have antigonadotrophic effects [15]. The testosterone derivatives, the “gonanes and estranes”, also referred to as levonorgestrel (LNG) and norethisterone (NTD) families [16], have variate activities (Table 1). The gonanes, such as gestodene (GES), norgestrel (NET), and more specifically, its active stereoisomer levonorgestrel (LNG), have high androgenic effects [17]. In contrast, etonogestrel (ENG), which is the biologically active metabolite of desogestrel, is an agonist of the progesterone receptor (PR), showing low androgenic activity and simultaneous glucocorticoid effects [18].
The estranes, NTD and norethisterone acetate (NTDA), have medium androgenic activity [17]. Dienogest (DIE), classified as a fourth generation progestin, is highly specific for the PR [19] and has no androgenic activity [20]. DIE is usually known as a hybrid progestin, as it has the chemical structure of 19-nortestosterone derivatives but shows antiandrogenic activity characteristics, which are typical of progesterone derivatives [20].
The progesterone derivatives, such as those closely related to 19-norprogesterone, which includes nomegestrol acetate (NOMAC), are called “pure” progestational molecules as they bind almost exclusively to the PR and do not interfere with another steroid receptor [19].
In contrast, those PGs derived from 17-hydroxyprogesterone exhibit varying activities. Thus, medroxyprogesterone acetate (MPA) and its metabolite medroxyprogesterone (MEP) has slight androgenic action and exerts glucocorticoid activity when given at high doses [21]. Megestrol acetate (MGA) has 50% fewer glucocorticoid effects than MPA [15]. These PGs also act in specific areas of the hypothalamus as antiandrogenic molecules [22]. This action control male sexual behaviour and urine marking—typical of several animals [22]. Moreover, while designed as a PR agonist, MPA has a high binding affinity for glucocorticoid receptors [23,24].
Usually, the most recent PGs derived from progesterone are progestational PGs without androgenic, estrogenic, or glucocorticoid activity. These PGs were conceived to mimic the benefits of progesterone without the undesirable effects of older PGs, such as acne, a decrease in high-density lipoprotein cholesterol (HDL-C), or bloating and water retention [15].
Drospirenone (DSP) is an aldosterone antagonist derived from spironolactone. The primary effect of the latter PG is its anti-mineralocorticoid activity, which causes decreased salt and water retention, leading to lower blood pressure and absence of androgenic effects [25]. Additionally, DSP exhibits partial antiandrogenic activity [26]—a property that may counter the adverse impact of androgens on hair growth, lipid fluctuation patterns, and insulin, and the possible influence of body composition in postmenopausal women [26]. Further details about PGs’ cellular targets and biological activities in humans can be found in the literature [27,28,29,30].
3. Waste and Surface Waters Concentrations of Synthetic Progestins
PGs are considered emerging micropollutants in aquatic ecosystems, where they are usually present in concentrations in the order of ng/L. However, accurately knowing their concentrations in waters is crucial since such tiny amounts are potentially harmful to (at least) fish [6,7]. Likely because analysing PGs requires trace analytical methods for their extraction and quantification, the number of studies concerning the environmental levels of these compounds is still scarce and, in a majority, focused on the concentrations of these hormones in influents and effluents from wastewater treatment plants (WWTPs).
In addition, the surveyed areas are still limited in space (Figure 1). From 2015 to 2021, most publications were performed in Europe (48%) and North America (24%). In Asia (19%), South America (5%), and Australia (5%), there are fewer details about the levels of synthetic PGs, and in Africa, as far as we notice, there are no data on this subject (Figure 1 and Table 2).
Besides, there are also differences concerning the types of PGs analysed. For example, in Europe, the most prevalent PGs in Switzerland [31] were DIE and MPA, whilst in the Czech Republic, it was MGA [32], and in Germany [33], it was DIE. In Asia, a recent study showed LNG, DSP, and dydrogesterone as the most frequently detected PGs in China [34].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 2.
Concentrations of synthetic progestins in waste and surface waters. Average (Av); not detected (ND); not evaluated (n.e.); quantification method (QM); surface waters (Sw); WWTP influent (WWTPi); WWTP effluents (WWTPe).
Table 2.
Concentrations of synthetic progestins in waste and surface waters. Average (Av); not detected (ND); not evaluated (n.e.); quantification method (QM); surface waters (Sw); WWTP influent (WWTPi); WWTP effluents (WWTPe).
| Testosterone derivatives (Gonanes) | PGs | QM | Sw (ng/L) | WWTPi (ng/L) | WWTPe (ng/L) | Local (Country) | References |
| GES | (1) | 0.2 | 3 | 1 | Basel and canton Zürich WWTPs (Switzerland). | [31] | |
| (2) | <0.05 | <0.38–7.7 | <0.29–0.71 | Blanice River and WWTPs (Czech Republic). | [32] | ||
| (2) | <0.64 | <0.41–7.0 | <0.19–<3.5 | Several WWTPs (Czech and Slovak Republics) | [35] | ||
| (1) | <0.3 | n.e. | <1.0 | Several WWTPs and rivers (Germany). | [33] | ||
| (3) | <0.2 | <3.0 | <1.0 | Jona River and WWTPs (Switzerland). | [36] | ||
| (4) | <21.5 | <21.5 | <21.5 | Five WWTPs (Portugal). | [37] | ||
| LNG | (1) | <2.5–117 | 493–811 | 32–39 | Langat River Basin (Malaysia). | [38] | |
| (5) | <2.5 | n.e. | <2.5 | Southeast Queensland (Australia). | [39] | ||
| (6) | 0.85–3.40 | n.e. | n.e. | Lake Balaton (Hungry). | [40] | ||
| (7) | <15 | n.e. | <15 | Two WWTPs in Quebec (Canada). | [41] | ||
| (2) | <0.08 | <0.26–<2.1 | <0.22–<0.83 | Blanice River and WWTPs (Czech Republic). | [32] | ||
| (2) | <0.09 | <0.07–<1.2 | <0.03–<0.32 | Several WWTPs (Czech and Slovak Republics). | [35] | ||
| (1) | <0.05–<0.7 | n.e. | <0.3–<1.0 | Several WWTPs and rivers (Germany). | [33] | ||
| (1) | ND | ND–38.4 | ND–20.1 | Several WWTPs, Quebec (Canada). | [42] | ||
| (8) | <2.5 | <5–299 ± 17 | <3.0 | Québec and Ontario (Canada). | [43] | ||
| (4) | n.e. | 2.81 | 1.37 | 21 WWTPs (China). | [34] | ||
| (4) | n.e. | n.e. | <1.0 | Several WWTPs effluents (Germany). | [44] | ||
| NET | (4) | n.e. | n.e. | <2.0 | Gran Canaria (Spain) | [45] | |
| (4) | n.e. | 11.2 | 1.92 | 21 WWTPs (China). | [34] | ||
| ENG | (2) | <0.07 | <0.28–<1.4 | <0.21–<0.89 | Blanice River and WWTPs (Czech Republic). | [32] | |
| (2) | <0.09 | <0.25–<1.2 | <0.18–<0.94 | Several WWTPs (Czech and Slovak Republics). | [35] | ||
| (1) | <0.3 | n.e. | <0.5 | Several WWTPs and rivers (Germany). | [33] | ||
| (4) | n.e. | n.e. | <1.2 | Several WWTPs effluents (Germany). | [44] | ||
| Testosterone derivatives (Estranes) | NTD | (1) | <2.5–230 | 1048–1137 | 218–265 | Langat River Basin (Malaysia). | [38] |
| (4) | n.e. | n.e. | <2.0 | Gran Canaria (Spain). | [45] | ||
| (9) | ND–5.20 | 1.02–94.7 Av. = 25.7 | ND–1.68 Av. = 1.25 | Four WWTPs, Shanghai (China). | [46] | ||
| (5) | <0.21–3.1 | n.e. | n.e. | Freshwater aquaculture (China). | [47] | ||
| (1) | <0.3 | <3 | <0.6 | Basel and canton Zürich WWTPs (Switzerland). | [31] | ||
| (7) | <11 | n.e. | <11 | Two WWTPs in Quebec (Canada). | [41] | ||
| (2) | <0.04 | <0.02–<0.17 | <0.03–0.85 | Blanice River and WWTPs (Czech Republic). | [32] | ||
| (2) | <0.01 | <0.02–<0.91 | <0.02–<4.1 | Several WWTPs (Czech and Slovak Republics). | [35] | ||
| (1) | n.e. | n.e. | <0.40 | Pharmaceutical manufacturing facility discharges (USA). | [48] | ||
| (3) | <0.3 | <3 | <0.6 | Jona River and several WWTPs (Switzerland). | [36] | ||
| (1) | <0.1–<0.3 | n.e. | <1.0 | Several WWTPs and rivers (Germany). | [33] | ||
| (8) | 1.7 ± 0.05–2.7 ± 0.17 | <4.8 | 2 ± 0.2–132 ± 2.2 | Québec and Ontario (Canada). | [43] | ||
| (10) | <2.3 | <2.3 | <2.3 | Basque Country (Spain). | [49] | ||
| (1) | ND | ND–78.8 | ND–31.8 | Several WWTPs, Quebec (Canada). | [42] | ||
| (4) | n.e. | 4.02 | 0.20 | 21 WWTPs (China). | [34] | ||
| (4) | n.e. | n.e. | <1.0 | Several WWTPs effluents (Germany). | [44] | ||
| NTDA | (4) | n.e. | 10.5 | 0.24 | 21 WWTPs (China). | [34] | |
| (1) | <0.3 | n.e. | <0.5 | Several WWTPs and rivers (Germany). | [33] | ||
| (4) | n.e. | n.e. | <1.0 | Several WWTPs (Germany). | [44] | ||
| DIE | (1) | <0.3 | <0.8 | <0.3 | Basel and canton Zürich WWTPs (Switzerland). | [31] | |
| (3) | <0.3 | <0.8 | <0.3 | Jona River and several WWTPs (Switzerland). | [36] | ||
| (2) | <0.09 | 1.9–11.0 | <0.05–1.0 | Blanice River and WWTPs (Czech Republic). | [32] | ||
| (2) | <0.04 | 1.3–12 | <0.04–<4.0 | Several WWTPs (Czech and Slovak Republics). | [35] | ||
| (1) | <0.02–2.3 | n.e. | 1.3–4.4 | Several WWTPs and rivers (Germany). | [33] | ||
| (4) | n.e. | n.e. | 0.3–3.7 | Several WWTPs effluents (Germany). | [44] | ||
Progesterone derivatives | NOMAC | (2) | <0.07 | <0.08–3.6 | <0.03–0.26 | Blanice River and WWTPs (Czech Republic). | [32] |
| MEP | (5) | <0.07–1.3 | n.e. | n.e. | Freshwater aquaculture (China). | [47] | |
| (1) | <0.6 | <6 | <3 | Basel and canton Zürich WWTPs (Switzerland). | [31] | ||
| (3) | <0.6 | <6 | <3 | Jona River and several WWTPs (Switzerland). | [36] | ||
| (2) | <0.06 | <0.02–<0.13 | <0.03–0.23 | Blanice River and WWTPs (Czech Republic). | [32] | ||
| (1) | ND | ND–5.7 | ND–2.9 | Several WWTPs, Quebec (Canada). | [42] | ||
| (2) | <0.04 | <0.01–<0.53 | <0.01–0.95 | Several WWTPs (Czech and Slovak Republics). | [35] | ||
| (1) | <0.05 | n.e. | <0.08 | Several WWTPs and rivers (Germany). | [33] | ||
| MPA | (5) | <0.21–0.31 | n.e. | n.e. | Freshwater aquaculture (China). | [47] | |
| (1) | <0.1 | <0.8 | <0.2 | Basel and canton Zürich WWTPs (Switzerland). | [31] | ||
| (2) | <0.1 | <0.15–4.4 | <0.09–0.58 | Blanice River and WWTPs (Czech Republic). | [32] | ||
| (2) | <0.01 | <0.04–8.1 | <0.04–0.38 | Several WWTPs (Czech and Slovak Republics). | [35] | ||
| (3) | <0.1 | <0.8–5.3 | <0.2 | Jona River and several WWTPs (Switzerland). | [36] | ||
| (1) | <0.05–0.1 | n.e. | <0.08–<0.3 | Several WWTPs and rivers (Germany). | [33] | ||
| (4) | n.e. | 3.09 | 0.23 | 21 WWTPs (China). | [34] | ||
| (4) | n.e. | n.e. | <0.6 | Several WWTPs effluents (Germany). | [44] | ||
| MGA | (4) | n.e. | n.e. | <60 | Gran Canaria (Spain). | [45] | |
| (1) | <0.1 | <1 | <0.6 | Basel and canton Zürich WWTPs (Switzerland). | [31] | ||
| (2) | <0.01 | 0.52–13.0 | 0.13–1.0 | Several WWTPs (Czech and Slovak Republics). | [35] | ||
| (1) | <0.05–<0.2 | n.e. | <0.06–<0.3 | Several WWTPs and rivers (Germany). | [33] | ||
| (2) | <0.07 | <0.03–<6.3 | <0.06–0.4 | Blanice River and WWTPs (Czech Republic). | [32] | ||
| (7) | <6–<20 | n.e. | n.e. | Water bodies in Santa Maria (Brazil). | [50] | ||
| (4) | n.e. | 0.84 | 0.29 | 21 WWTPs (China). | [34] | ||
| Spironolactone derivative | DSP | (6) | 0.26–4.30 | n.e. | n.e. | Lake Balaton (Hungry). | [40] |
| (1) | <0.3 | <4 | <1 | Basel and canton Zürich WWTPs (Switzerland). | [31] | ||
| (2) | <0.85 | 0.64–0.77 | <0.18–<0.62 | Blanice River and WWTPs (Czech Republic). | [32] | ||
| (2) | <0.04 | 0.34–6.7 | <0.07–<0.29 | Several WWTPs (Czech and Slovak Republics). | [35] | ||
| (3) | <0.3 | <4 | <1 | Jona River and several WWTPs (Switzerland). | [36] | ||
| (1) | <0.3 | n.e. | <0.05 | Several WWTPs and rivers (Germany). | [33] | ||
| (4) | n.e. | 0.69 | 0.39 | 21 WWTPs (China). | [34] | ||
| (4) | n.e. | n.e. | <0.8 | Several WWTPs effluents (Germany). | [44] |
(1) Liquid chromatography with tandem mass spectrometry detection (LC-MS/MS); (2) liquid chromatography tandem atmospheric pressure chemical ionization/atmospheric pressure photoionization with hybrid quadrupole/orbital trap mass spectrometry operated in high-resolution product scan mode (LC-APCI/APPI-HRPS); (3) high-performance liquid chromatography coupled to a triple quadrupole mass spectrometry (HPLC-MS/MS); (4) ultra-performance liquid chromatography coupled with tandem mass detection (UPLC-MS/MS); (5) gas chromatography with tandem mass spectrometry detection (GC-MS/MS); (6) high-performance liquid chromatography–mass spectrometry (HPLC-MS); (7) liquid chromatography–mass spectrometry (LC-MS); (8) triple quadrupole-linear ion trap mass spectrometer using the sMRM (scheduled multiple reaction monitoring) mode (TripleQuad-LIT-MS); (9) rapid resolution liquid chromatography/tandem mass spectrometry (RRLC-MS/MS); (10) laser diode thermal desorption–tandem mass spectrometry (LDTD–MS/MS).
Here, concerning the 12 PGs in Table 1, the most investigated (%) were NTD (20%) and LNG (14%). There are still less data concerning MPA, DSP (10%), MEP, MGA (9%), GES, DIE (8%), ENG (5%), NTDA (4%), NET (3%), and NOMAC (1%) (Table 2).
Therefore, in an accessible and organised way, this paper compiles the existing data in the bibliography relative to the concentrations of 12 PGs from 2015 to 2021, using the “Web of Science Core Collection” and “PubMed” as primary databases. Thus, Table 2 presents data on the concentrations of these hormones in surface waters and wastewater treatment plants (WWTPs) worldwide, considering their influents and effluents.
Data in Table 2 were gathered from investigations conducted in various geographic locations, with varying PG inputs, and analysed according to well-established analytical techniques, despite the varying detection and quantification levels and accuracies. It is important to stress that some of the surveyed areas in Asia [38,46,47] are densely populated, which may explain the high amounts of PGs measured in surface waters. Therefore, the disparities between studies from distinct regions are not surprising, corresponding to a wide range of concentrations even when including the three compartments of surface waters, WWTP influents, and WWTP effluents.
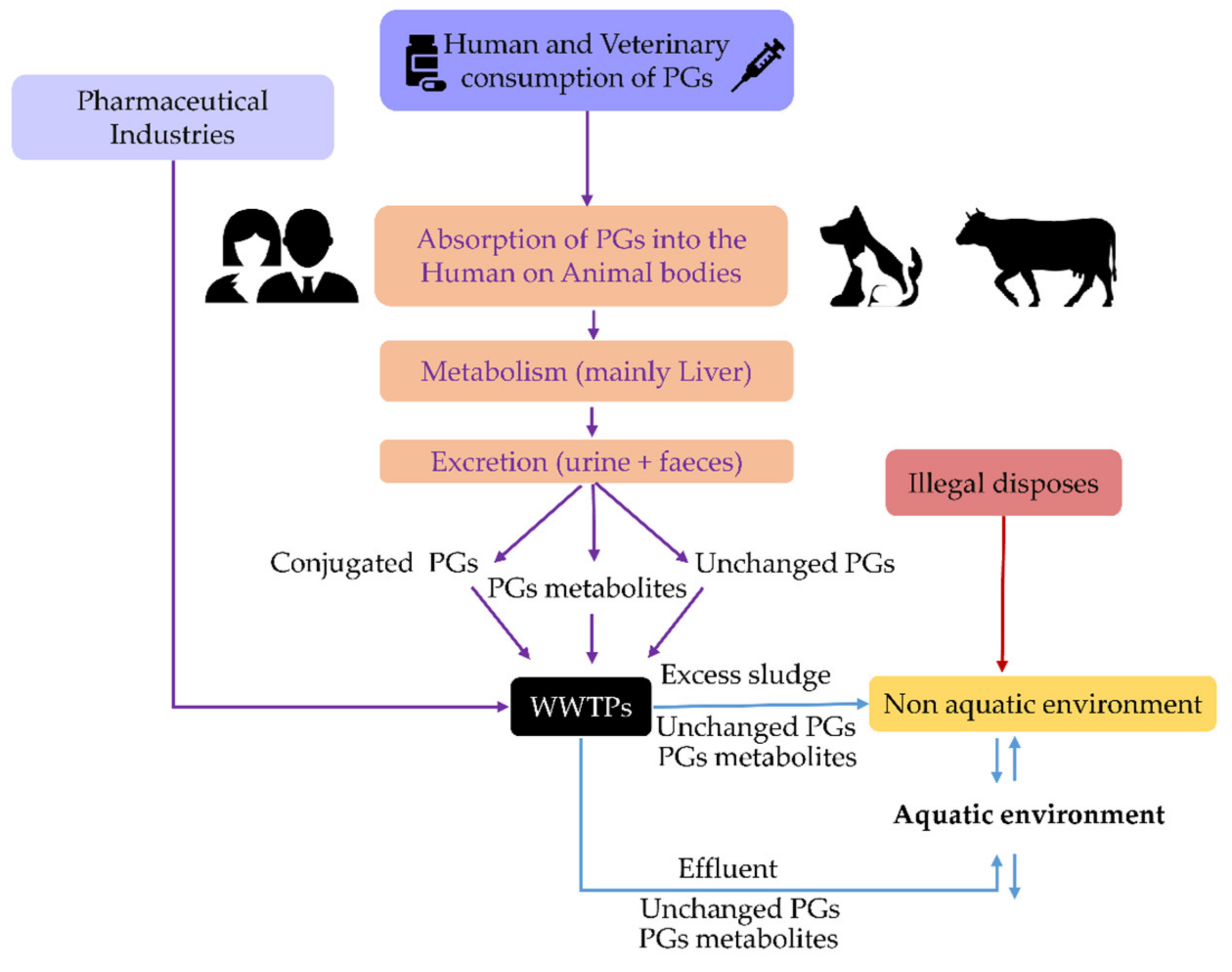
Despite the differences mentioned, Figure 2 shows that synthetic PGs are still present in surface waters in amounts comparable to those observed in WWTP effluents, which is concerning given that dilution is predicted in surface waters. A similar observation was also noticed in previous studies [6,7]. As a result, one can infer that WWTPs do not effectively remove these compounds and/or that some of them can be regenerated in the aquatic environment by deconjugation phenomena (Figure 3).
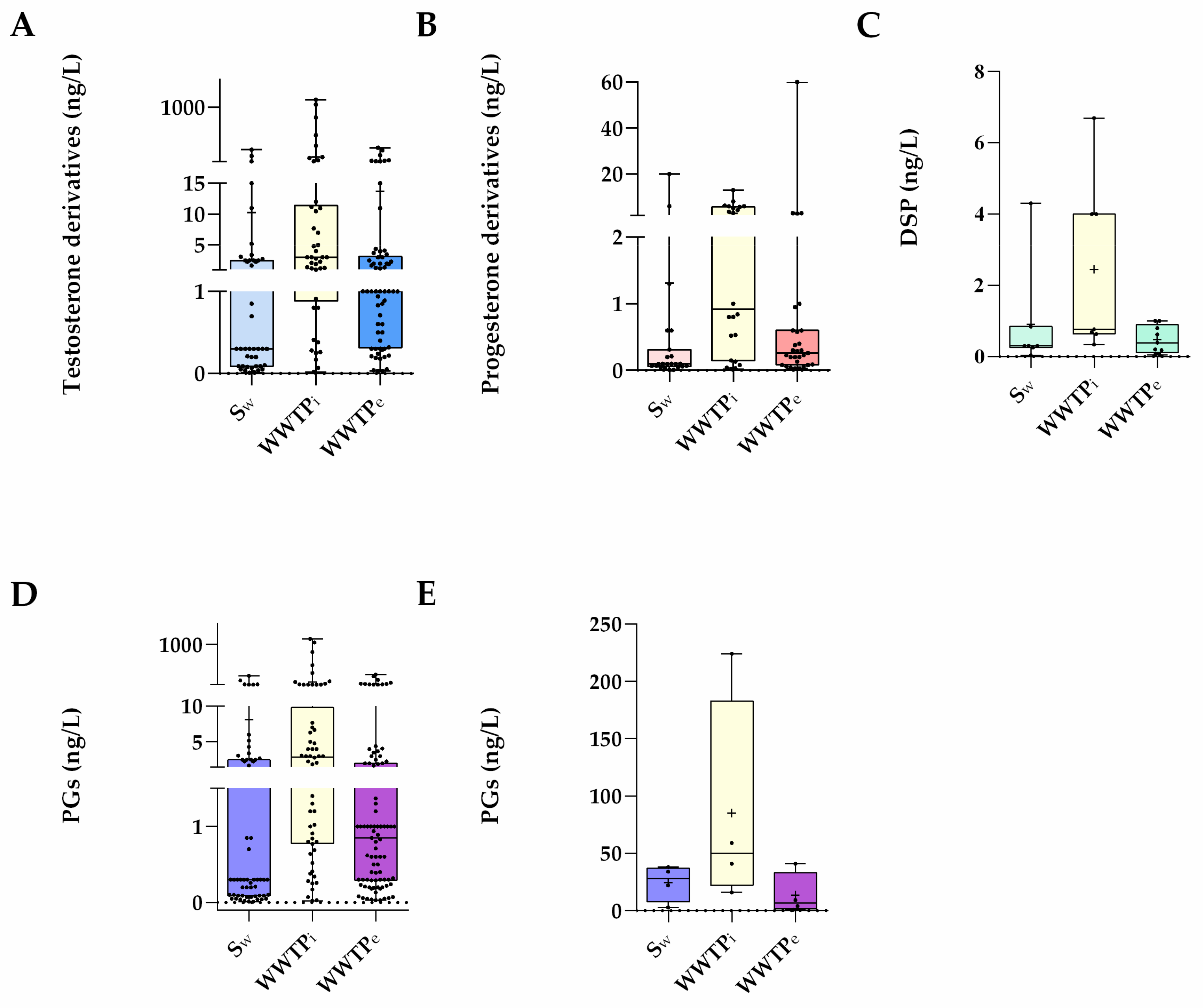
In particular, Figure 2A shows that PGs derived from testosterone, besides being evaluated in a higher number of studies, were also the hormones with higher concentrations (up to ≅1 µg/L) in the aquatic environments, where their global load reaches ≅97.0% of all PGs considered in Table 2 vs. 2.49% and 0.57% for progesterone and spironolactone derivatives.
Table 2 reveals that in surface waters, the concentrations of PGs derived from testosterone were typically higher for LNG (<0.05–117 ng/L) and NTD (<0.01–230 ng/L) than those for GES (<0.05–21.5 ng/L), DIE (<0.02–2.3 ng/L), and NTDA (<0.3 ng/L) ≅ ENG (<0.07–<0.3 ng/L). Data concerning NET in surface waters were not available.
In WWTP influents, the concentrations of LNG (<0.07–811 ng/L ) and NTD (<0.02–1137 ng/L) were consistently higher than those of GES (<0.38–<21.5 ng/L), DIE (<0.8–12.0 ng/L) ≅ NET (11.2 ng/L) ≅ NTDA (10.5 ng/L), and ENG (<0.28–<1.4 ng/L).
In WWTP effluents, the highest concentrations were measured for LNG (<0.03–39 ng/L), NTD (<0.03–265 ng/L), followed by GES (<0.19–<21.5 ng/L), DIE (<0.04–4.4 ng/L), NET (<2.0–1.92 ng/L), NTDA (0.24–<1.0 ng/L) ≅ ENG (<0.21–<1.2 ng/L).
Progesterone-derived PGs more commonly exist in surface waters in concentrations ca. 17-fold lower (Figure 2B) than those reported above for the testosterone derivatives. Such PGs showed similar concentrations to those of the natural hormone progesterone, which ranged from ND to 13.67 ng/L [38,40] in surface waters and from <0.04 ng/L to 24.8 ng/L [32,42] in WWTP influents. In WWTPs effluents, the levels of those PGs were lower than those of progesterone (ND to 110 ng/L) [32,43]. Despite this, progesterone-derived PGs concentrations in surface waters are comparable to those in WWTP effluents, much as testosterone-derived PGs. Moreover, the two most prevalent progesterone-derived PGs, MGA and MEP, were found in identical amounts in all three aquatic compartments.
Regarding surface waters, the concentrations of MGA (<0.01–<20.0 ng/L) seem higher than those of MEP (<0.04–1.3 ng/L), MPA (<0.01–0.31 ng/L) and NOMAC (<0.07 ng/L).
In WWTP influents, the concentrations of MGA (<0.03–13.0 ng/L) reach higher levels, despite overlapping to some extent with those of MPA (<0.04–8.1 ng/L) and MEP (<0.02–5.7 ng/L), with NOMAC (<0.08–3.6 ng/L) levels being suggestively lower. The occurrence of higher amounts of MEP and MPA, despite being punctual, in surface waters than in WWTP influents is very worrying, stressing the need for more studies concerning this subject.
In WWTP effluents, the highest levels were measured for MGA (<0.06–<60 ng/L), followed by MEP (<0.01–2.9 ng/L), MPA (<0.08–0.58 ng/L), and NOMAC (<0.03–0.26 ng/L).
Finally, considering the spironolactone derivative DSP, it is observed that its concentrations in surface waters (<0.04–4.3 ng/L) were higher than those in WWTP effluents (<0.05–<1.0 ng/L) but lower than those from WWTP influents (0.34–6.7 ng/L) (Figure 2C).
Altogether, this review found (Figure 2D) that the current number of studies on PGs in waters sharply increased when compared with those reported in previous reviews [7] (Figure 2E). In addition, the more recent environmental concentrations of these compounds have risen compared to data published before 2015. Additionally, there are studies on DSP that were not available before [7].
As shown in Table 2, there are parent PGs in surface waters whose origin is unknown. Therefore, it is not established if deconjugation occurs in the aquatic environments and/or if there is a lack of efficient removal of these compounds by WWTPs (Figure 3). As such, we found it helpful to determine PGs removal efficiency in WWTPs. For this purpose, when this information was not available in the bibliography, we used concentrations of PGs in WWTPs influents and effluents reported in Table 2 and inserted them in Equation (1):
Globally, the removal efficiency values of PGs in WWTPs are, on average, 73% (Table 3), which is considered a standard removal percentage for steroids in WWTPs [52,53]. However, at some locations (Table 3, values in bold), the presence of parent compounds was higher in WWTP effluents than in their influents; e.g., GES, LNG, ENG, NTD, DIE and MGA in Czech and Slovak Republics’ WWTPs [32,35]. These negative removal efficiency rates have been explained by the deconjugation of metabolised steroid hormones, including synthetic PGs in WWTPs, which become regenerated free parent steroids (Figure 3) by biodegradation, hydrolysis, and even photolysis [51,52,53,54].
Another important aspect shown in Figure 3 is that, beyond the hypothesised regeneration of parent PGs, active metabolites of these pharmaceuticals also arrive in the aquatic environment. Some of these metabolites are still awaiting their identification and answers about their activity [55]. However, others have already been identified. An example of this is shown by the metabolisation of NTDA, which originates ethinylestradiol (EE2), a potent estrogen known to produce endocrine disorders in concentrations as low as a few ng/L [44,56]. Additionally, through side-chain cleavage, the PGs closely related to progesterone by metabolisation can produce potent androgens; e.g., 4-adrostene-3,17-dione, and 5α-dihydrotestosterone [57].
Table 4 shows the most recent advances concerning the human metabolisation of the 12 PGs referred to herein. Thus, further studies involving the parent and the active metabolites of these molecules should be considered in future monitoring programs once it is already established that the latter can also induce health disorders in aquatic organisms [57].
4. Biological Effects of PGs in Aquatic Organisms, Particularly Fishes
In invertebrates, progesterone also plays a central role in reproduction [73,74]. An ancient origin of progesterone and its receptor is well shown in a study using the micro invertebrate Brachionus manjavacas (Rotifera) [73]. This work undoubtedly exemplifies that progesterone and its receptor exhibit conservation of function over a broad range of animals across phylogenies, presenting further evidence about the ancient origin of hormonal steroid regulation and suggesting that the endocrine regulation of mammalian reproduction may be derived from primitive regulatory pathways [73].
In amphibians, natural PGs are also involved in numerous biological activities, which includes gonadal development/differentiation, germinal vesicle breakdown (GVBD), and adequate homeostasis of the hypothalamic–pituitary–gonadal axis (HPG), among others [75].
Compared with tetrapods, the plasma levels of progesterone in fish are usually low, as the hormone is essentially an intermediate in the steroidogenic pathway of these organisms and other natural PGs predominate, such as 17α-hydroxyprogesterone (17-OHP), 17,20β-dihydroxypregn-4-en-3-one (17,20β-P) and 17,20β,21-trihydroxypregn-4-en-3-one (17,20β-S) [76]. These and other PGs are involved in reproductive functions such as follicular steroidogenesis, spermatogenesis, pheromone synthesis, in the homeostasis of the immune and cardiovascular systems, and even as neuroprotectors [76,77,78,79,80,81].
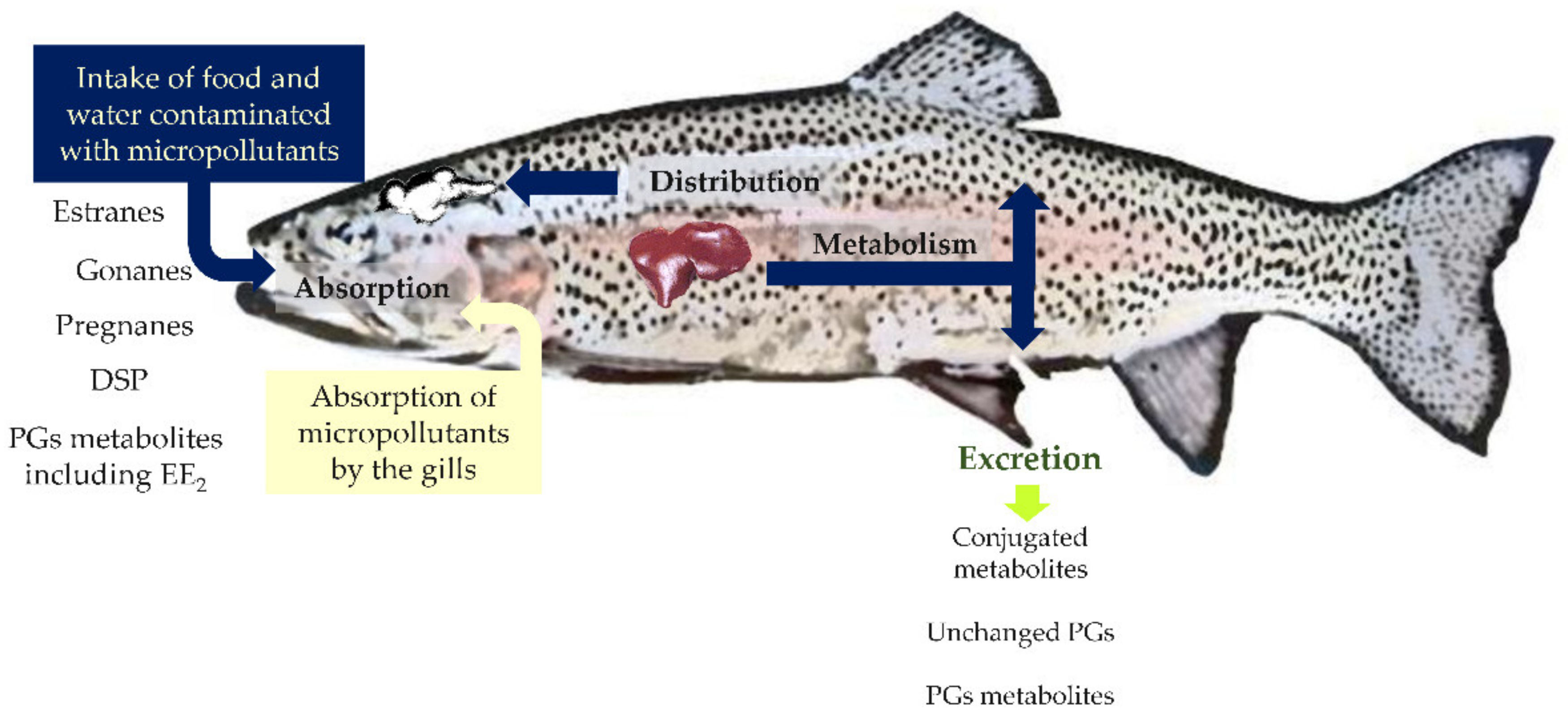
These observations demonstrate that PGs are essential steroid hormones of aquatic organisms [76]. Thus, it is not surprising that once in the water, synthetic PGs, when uptaken by animals via food, water ingestion, or direct contact with gills, interact with cell PGs receptors of these organisms, disrupting their normal physiological status (Figure 4).
Therefore, and according to the National Institute of Environmental Health Sciences (NIEHS), PGs are EDCs, as this classification embraces natural or man-made compounds that can mimic or interfere with the function of hormones in an organism, producing a variety of adverse effects on, e.g., reproductive, neurological and immune systems [82].
It was previously stressed that concentrations as low as 0.8–1.0 ng/L of PGs induce endocrine disturbances in fish [6], which falls within the range observed in waters referred to in Table 2. This fact makes PGs one of the most critical pharmaceutical compounds of concern after EE2, which can be a metabolisation product of NTDA [33]. The latter estrogen is already included in the EU watchlist of substances with environmental interest [83].
Once the deleterious role of PGs in fish [6], amphibians (Xenopus laevis and X. tropicalenses) [84,85], and even mussels (Dreissena polymorpha) [86] has been recognised, as they interfere with the normal function of vital organs and produce sufficiently important alterations that impact survival, growth, and reproduction [6,75,87,88], more studies involving fish as test organisms have been published. Therefore, we elaborated Table 5 to summarise the advances on the effects of PGs in fish as published from 2015 to 2021.
5. Bioconcentration Factors and Predicted Effect Concentrations of PGs in Fish Plasma
Another critical aspect of PGs in fish and other aquatic organisms is that after absorption, the substances can be bioconcentrated, bioaccumulated, or both (Figure 4).
Bioconcentration factors (BCFs) describe the readiness of chemicals to concentrate in organisms when they are present in the environment. These are determined by the ratio between the concentration of a specific chemical inside biological tissues and its levels in the surrounding environment [131]. In vivo experiments led to the obtention of the BCFs for three PGs: the LNG 17–53 [132], NET 2.6–40.8 [133,134], and MPA 4.3–37.8 [135].
Nonetheless, when in vivo assays are not available, it is possible to use “log Kow—based models” to assess the BCFs [6]. In fact, BCF values have already been used to predict critical environmental concentrations of 500 pharmaceuticals [6]. Thus, when plasma concentrations of a specific PG and BCFs (either measured or predicted) are known, the environmental concentration of progestins in the surrounding water for fish can be back-calculated.
So, using this method, the BCFFP = bioconcentration factor in fish plasma can be calculated by applying Equation (2) [136,137,138]:
In addition, the “Predicted Effect Concentration” (PECw) of a particular chemical can be taken from the bibliography or from in vivo experiments [6,139,140,141]. For instance, when exposing rainbow trout (Oncorhynchus mykiss) to 1 ng/mL of LNG, a maximum of 12 ng/mL of this compound existed in plasma, which is a concentration that exceeds five times the human therapeutic dosage of 2.4 ng/mL [136]. In this case, LNG is likely to produce effects in fish in line with those seen in humans. Alternatively, PECw can be calculated mathematically (3). This approach is based on the concept that when the concentration in the plasma of a fish (CFP) reaches the “therapeutic dose” observed in humans, similar effects, at least to some extent, can be expected in fish. This perspective is grounded on the fact that many receptors and enzyme systems are conserved across mammalian and non-mammalian species, making mechanism of action extrapolations possible for a particular compound, considering its environmental concentrations [6,137].
Here, the values of BCFFP and PECw for the surveyed PGs were calculated as shown above and used to assess possible deleterious effects of those chemicals in fish (Table 6) beyond their possible bioaccumulation in these organisms [6].
When comparing the BCFFP determined in vivo with those reported in Table 6, it is observed that the current data are consistent with the results obtained for LNG and NET using the channel catfish (Ictalurus punctatus), fathead minnow (P. promelas) [133,134], and roach (R. rutilus) [132]. Specifically, in vivo, the BCFs for the latter two PGs ranged from 2.6 to 40.8, similar to the mathematically estimated BCFFP of 46 (Table 6). However, this mathematical approach is not always comparable with the in vivo assays, as shown by the data obtained for MPA in carp (C. carpio) [135]. The previous studies revealed that BCFs for MPA range from 4.3 to 37.8, whereas those determined in Table 6 point to 128.
Thus, regardless of the utility of the theoretical assessments, studies exposing different fish species to the current PGs are needed to avoid inaccurate conclusions that may derive from those kinds of estimates.
Taking this in mind, but still relying on the data in Table 6, it is probable that both LNG and NET exist in surface waters and WWTPs influents in amounts that can induce fish endocrine disruption, as suggested both the theoretically calculated and the in vivo data for these two PGs. However, further studies involving in vivo assays are required to prove indubitably this hypothesis.
Ultimately, when humans eat PGs-contaminated fish, they are unwittingly exposed to these chemicals and thus, at least in theory, human health could be impacted by these contaminants. However, as far as we noticed, there are no published data about this issue.
6. Evaluation of Risk Quotients (RQs) for PGs
Because several PGs showed potential to induce adverse effects in fish, it was considered opportune to investigate their impact by examining their risk quotients (RQs) in the aquatic environment. The parameter RQ is known to realistically estimate the potential ecological risk; i.e., the probability of an expected effect or potential danger caused by an environmental concentration of a pollutant.
The calculus of this quotient involves the ratio between the “Measured Environmental Concentrations (MECs)” and the “Predicted No Effect Concentrations (PNECs)” of a certain compound (4) [141,142]:
The MECs in this study were the average environmental concentrations provided in Table 2 for surface waters, and the ranking criteria used were RQ > 1.0 for high ecological risk, 0.1 < RQ < 1.0 for medium risk, and RQ < 0.1 for low risk [143]. The use of maximum concentrations and ranking criteria could estimate extreme worst-case scenarios.
The PNECs reflect the relative toxicity of each molecule for fish, and when they are not published, they can be derived by following the standard scientific assessment procedures defined in the EU Guidance for REACH implementation [144]. In those situations, the PNECs can be calculated considering several endpoint values found in the literature or using the “Species Sensitivity Distribution” (SSD) method, divided by their respective “Assessment factors” (AF), as proposed in Table 7 [145].
The preferred endpoints for screening-level risk assessments are values of chronic toxicity tests, represented by the “No Observed Effect Concentration” (NOEC). Whenever NOECs are not accessible, it is also adequate to use the “Lowest Observed Effect Concentration” (LOEC), the median “Effective Concentration” (EC50), or the Lethal Concentration (LC50) taken from acute toxicity tests.
Although the PNECs of GES, ENG and NTDA were calculated considering acute toxicity tests, those for NOR, NOMAC, and MPA were based on data from chronic toxicity tests in fish. Thus, some of the present conclusions will be valid for chronic while others for acute exposures.
The calculation of the RQs values (Table 8) reveals that the categories of PGs posing a higher ecologic risk for fish are those structurally related to testosterone. All the analysed PGs of this category, except for DIE and NET, show extremely high RQ values. This observation can be related to the extensive usage of these compounds and the higher number of published data considering these compounds vs. the other PGs (i.e., most of the studies involved the measurement of LNG ≅ 40 %, followed by NTD ≅ 18 % and DSP ≅ 16%). It is stressed that both LNG and NET have been referred to in the last section as having the ability to attain or even surpass in fish plasma the therapeutical dosages used in humans (Table 6).
In opposition, almost all PGs structurally related to progesterone and DSP showed RQs < 1, suggesting that these EDCs are less problematic than those referred to above. However, this hypothesis needs further investigation as the number of studies involving these compounds is lower than those for gonanes and estranes.
A final word of caution is due because the estimated risks are for single progestins. To precisely apprehend the global impact of these EDCs on aquatic organisms, research using mixtures and, therefore, better replicating environmental conditions are required. In this regard, there are still very few studies addressing the effects of mixtures of PGs in aquatic animals, covering only a few PGs and a couple of fish species [121,123]. As such, it is premature and would be incorrect to take a mixture toxicology approach.
7. Final Remarks
Progestins are confirmed EDCs for aquatic organisms—in particular, for fish living in polluted environments. However, when compared to other pollutants in the same category, such as EE2, it can be concluded that PGs are still understudied in terms of their functional effects on aquatic organisms at different trophic levels.
Except for NET, which has not been examined, all other synthetic PGs studied are present in surface waters, and all occur in WWTP influents and effluents. The latter two matrices have been the primary focus of environmental monitoring. As a result, there is a need for other aquatic matrices (e.g., lakes, estuaries, seashores, subterranean waters) to be investigated. In parallel, there is a need to widen the monitoring to more geographic areas, as the majority of studies have been conducted in Europe, Canada, and China.
In our view, the state of art already calls for regulation on the concentration limits for the discharge of PGs in WWTP effluents. Considering the precautionary principle, the pertinence of that possibility should be taken into account in future WFD updates.
Furthermore, because most PG metabolites remain biologically active, their proper identification should be evaluated, and their hazardous impact should be included in future investigations. To fully appreciate the influence of these chemicals in aquatic systems, it is also necessary to examine the biological consequences of complex combinations of parent PGs and their active metabolites.
Author Contributions
Conceptualisation, M.J.R. and E.R.; methodology, M.J.R.; formal analysis, M.J.R. and E.R.; writing—original draft preparation, M.J.R.; writing—review and editing, E.R.; visualisation, M.J.R.; funding acquisition, M.J.R. and E.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was a result of the participation in the project ATLANTIDA—Platform for the monitoring of the North Atlantic Ocean and tools for the sustainable exploitation of the marine resources (NORTE-01-0145-FEDER-000040), supported by Norte Portugal Regional Operational Programme (NORTE 2020), under the PORTUGAL 2020 Partnership Agreement, through the European Regional Development Fund (ERDF). The authors’ efforts were additionally supported by the Strategic Funding UID/Multi/04423/2019 through national funds provided by FCT and ERDF to CIIMAR/CIMAR, in the framework of the program PT2020.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wilson, N.J.; Harris, L.M.; Nelson, J.; Shah, S.H. Re-theorizing politics in water governance. Water 2019, 11, 1470. [Google Scholar] [CrossRef] [Green Version]
- Cosgrove, W.J.; Loucks, D.P. Water management: Current and future challenges and research directions. Water Resour. Res. 2015, 51, 4823–4839. [Google Scholar] [CrossRef] [Green Version]
- EU Water Initiative (EUWI). Water Scarcity Management in the Context of WFD. 2006, SCG Agenda Point 8b (WGB/15160506/25d). Available online: https://ec.europa.eu/environment/water/quantity/pdf/comm_droughts/8a_1.pdf (accessed on 27 December 2021).
- EU. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 establishing a framework for Community action in the field of water policy. Off. J. Eur. Communities 2000, L327, 0001–0073. [Google Scholar]
- Arnold, K.E.; Brown, A.R.; Ankley, G.T.; Sumpter, J.P. Medicating the environment: Assessing risks of pharmaceuticals to wildlife and ecosystems. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130569. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Johnson, A.C.; Trubiroha, A.; Tumová, J.; Ihara, M.; Grabic, R.; Kloas, W.; Tanaka, H.; Kroupová, H.K. The Challenge presented by progestins in ecotoxicological research: A critical review. Environ. Sci. Technol. 2015, 49, 2625–2638. [Google Scholar] [CrossRef]
- Fent, K. Progestins as endocrine disrupters in aquatic ecosystems: Concentrations, effects and risk assessment. Environ. Int. 2015, 84, 115–130. [Google Scholar] [CrossRef]
- Stanczyk, F.Z.; Hapgood, J.P.; Winer, S.; Mishell, D.R., Jr. Progestogens used in postmenopausal hormone therapy: Differences in their pharmacological properties, intracellular actions, and clinical effects. Endocr. Rev. 2013, 34, 171–208. [Google Scholar] [CrossRef] [Green Version]
- Gambrell, R.D., Jr.; Bagnell, C.A.; Greenblatt, R.B. Role of estrogens and progesterone in the etiology and prevention of endometrial cancer: Review. Am. J. Obstet. Gynecol. 1983, 146, 696–707. [Google Scholar] [CrossRef]
- Lobo, R.A. The role of progestins in hormone replacement therapy. Am. J. Obstet. Gynecol. 1992, 166, 1997–2004. [Google Scholar] [CrossRef]
- Chuong, C.J.; Brenner, P.F. Management of abnormal uterine bleeding. Am. J. Obstet. Gynecol. 1996, 175, 787–792. [Google Scholar] [CrossRef]
- Kornek, G.V.; Schenk, T.; Ludwig, H.; Hejna, M.; Scheithauer, W. Placebo-controlled trial of medroxy—Progesterone acetate in gastrointestinal malignancies and cachexia. Onc. Res. Treat. 1996, 19, 164–168. [Google Scholar] [CrossRef]
- EU. The European agency for the evaluation of medicinal products. Eval. Med. Insp. 1999, 33, 969–978. [Google Scholar]
- Apgar, B.S.; Greenberg, G. Using progestins in clinical practice. Am. Fam. Physician 2000, 62, 1839–1846. [Google Scholar]
- Sitruk-Ware, R. New progestagens for contraceptive use. Hum. Reprod. Update 2006, 12, 169–178. [Google Scholar] [CrossRef]
- Edgren, R.A.; Stanczyk, F.Z. Nomenclature of the gonane progestins. Contraception 1999, 60, 313. [Google Scholar] [CrossRef]
- Mathur, R.; Levin, O.; Azziz, R. Use of ethinylestradiol/drospirenone combination in patients with the polycystic ovary syndrome. Ther. Clin. Risk Manag. 2008, 4, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Hohmann, H.; Creinin, M.D. The contraceptive implant. Clin. Obstet. Gynecol. 2007, 50, 907–917. [Google Scholar] [CrossRef]
- Sitruk-Ware, R. Pharmacological profile of progestins. Maturitas 2004, 47, 277–283. [Google Scholar] [CrossRef]
- Katsuki, Y.; Sasagawa, S.; Takano, Y.; Shibutani, Y.; Aoki, D.; Udagawa, Y.; Nozawa, S. Animal studies on the endocrinological profile of dienogest, a novel synthetic steroid. Drugs Exp. Clin. Res. 1997, 23, 45–62. [Google Scholar]
- Bullock, L.P.; Bardin, C.W. Androgenic, synandrogenic, and antiandrogenic actions of progestins. Ann. N. Y. Acad. Sci. 1977, 286, 321–330. [Google Scholar] [CrossRef]
- Seksel, K. Chapter 7—Behavior-modifying drugs. In Small Animal Clinical Pharmacology, 2nd ed.; Maddison, J.E., Page, S.W., Church, D.B., Eds.; W.B. Saunders: Edinburgh, UK, 2008; pp. 126–147. [Google Scholar]
- Moore, N.L.; Hanson, A.R.; Ebrahimie, E.; Hickey, T.E.; Tilley, W.D. Anti-proliferative transcriptional effects of medroxyprogesterone acetate in estrogen receptor positive breast cancer cells are predominantly mediated by the progesterone receptor. J. Steroid Biochem. Mol. Biol. 2020, 199, 105548. [Google Scholar] [CrossRef]
- Thomas, C.P.; Liu, K.Z.; Vats, H.S. Medroxyprogesterone acetate binds the glucocorticoid receptor to stimulate α-ENaC and sgk1 expression in renal collecting duct epithelia. Am. J. Physiol. Renal. Physiol. 2006, 290, F306–F312. [Google Scholar] [CrossRef] [Green Version]
- Murphy, S.J.; Littleton-Kearney, M.T.; McCullough, L.D.; Hurn, P.D. Sex, hormones and the endothelium. In Advances in Molecular and Cell Biology; Elsevier: Amsterdam, The Netherlands, 2004; Volume 34, pp. 71–84. [Google Scholar]
- Sitruk-Ware, R. Pharmacology of different progestogens: The special case of drospirenone. Climacteric 2005, 8, 4–12. [Google Scholar] [CrossRef]
- Schindler, A.E.; Campagnoli, C.; Druckmann, R.; Huber, J.; Pasqualini, J.R.; Schweppe, K.W.; Thijssen, J.H.H. Reprint of Classification and pharmacology of progestins. Maturitas 2008, 61, 171–180. [Google Scholar] [CrossRef]
- Fraser, I.S. Non-contraceptive health benefits of intrauterine hormonal systems. Contraception 2010, 82, 396–403. [Google Scholar] [CrossRef]
- Regidor, P.-A. The clinical relevance of progestogens in hormonal contraception: Present status and future developments. Oncotarget 2018, 9, 34628–34638. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.H.; Kaunitz, A.M.; Hickey, M. Chapter 18—Hormonal Contraception. In Williams Textbook of Endocrinology, 13th ed.; Melmed, S., Polonsky, K.S., Larsen, P.R., Kronenberg, H.M., Eds.; Elsevier: Philadelphia, PA, USA, 2016; pp. 664–693. [Google Scholar]
- Zhang, K.; Zhao, Y.; Fent, K. Occurrence and ecotoxicological effects of free, conjugated, and halogenated steroids including 17α-hydroxypregnanolone and pregnanediol in Swiss. Environ. Sci. Technol. 2017, 51, 6498–6506. [Google Scholar] [CrossRef]
- Golovko, O.; Šauer, P.; Fedorova, G.; Kroupová, H.K.; Grabic, R. Determination of progestogens in surface and waste water using SPE extraction and LC-APCI/APPI-HRPS. Sci. Total Environ. 2018, 621, 1066–1073. [Google Scholar] [CrossRef] [PubMed]
- Weizel, A.; Schlüsener, M.P.; Dierkes, G.; Ternes, T.A. Occurrence of glucocorticoids, mineralocorticoids, and progestogens in various treated wastewater, rivers, and streams. Environ. Sci. Technol. 2018, 52, 5296–5307. [Google Scholar] [CrossRef]
- Yu, Q.; Geng, J.; Zong, X.; Zhang, Y.; Xu, K.; Hu, H.; Deng, Y.; Zhao, F.; Ren, H. Occurrence and removal of progestagens in municipal wastewater treatment plants from different regions in China. Sci. Total Environ. 2019, 668, 1191–1199. [Google Scholar] [CrossRef]
- Šauer, P.; Stará, A.; Golovko, O.; Valentová, O.; Bořík, A.; Grabic, R.; Kroupová, H.K. Two synthetic progestins and natural progesterone are responsible for most of the progestagenic activities in municipal wastewater treatment plant effluents in the Czech and Slovak republics. Water Res. 2018, 137, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Fent, K. Determination of two progestin metabolites (17α-hydroxypregnanolone and pregnanediol) and different classes of steroids (androgens, estrogens, corticosteroids, progestins) in rivers and wastewaters by high-performance liquid chromatography-tandem mass spectrometry (HPLC-MS/MS). Sci. Total Environ. 2018, 610–611, 1164–1172. [Google Scholar] [CrossRef]
- Silva, S.; Cardoso, V.V.; Duarte, L.; Carneiro, R.N.; Almeida, C.M.M. Characterization of five Portuguese wastewater treatment plants: Removal efficiency of pharmaceutical active compounds through conventional treatment processes and environmental risk. Appl. Sci. 2021, 11, 7388. [Google Scholar] [CrossRef]
- Tan, E.S.S.; Ho, Y.B.; Zakaria, M.P.; Latif, P.A.; Saari, N. Simultaneous extraction and determination of pharmaceuticals and personal care products (PPCPs) in river water and sewage by solid-phase extraction and liquid chromatography-tandem mass spectrometry. Int. J. Environ. Anal. Chem. 2015, 95, 816–832. [Google Scholar] [CrossRef]
- King, O.C.; van de Merwe, J.P.; McDonald, J.A.; Leusch, F.D.L. Concentrations of levonorgestrel and ethinylestradiol in wastewater effluents: Is the progestin also cause for concern? Environ. Toxicol. Chem. 2016, 35, 1378–1385. [Google Scholar] [CrossRef]
- Avar, P.; Maasz, G.; Takacs, P.; Lovas, S.; Zrinyi, Z.; Svigruha, R.; Takatsy, A.; Toth, L.G.; Pirger, Z. HPLC-MS/MS analysis of steroid hormones in environmental water samples. Drug Test. Anal. 2016, 8, 123–127. [Google Scholar] [CrossRef]
- Comtois-Marotte, S.; Chappuis, T.; Vo Duy, S.; Gilbert, N.; Lajeunesse, A.; Taktek, S.; Desrosiers, M.; Veilleux, É.; Sauvé, S. Analysis of emerging contaminants in water and solid samples using high resolution mass spectrometry with a Q Exactive orbital ion trap and estrogenic activity with YES-assay. Chemosphere 2017, 166, 400–411. [Google Scholar] [CrossRef]
- Yarahmadi, H.; Duy, S.V.; Hachad, M.; Dorner, S.; Sauvé, S.; Prévost, M. Seasonal variations of steroid hormones released by wastewater treatment plants to river water and sediments: Distribution between particulate and dissolved phases. Sci. Total Environ. 2018, 635, 144–155. [Google Scholar] [CrossRef]
- Goeury, K.; Vo Duy, S.; Munoz, G.; Prévost, M.; Sauvé, S. Analysis of Environmental Protection Agency priority endocrine disruptor hormones and bisphenol A in tap, surface and wastewater by online concentration liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2019, 1591, 87–98. [Google Scholar] [CrossRef]
- Weizel, A.; Schlüsener, M.P.; Dierkes, G.; Wick, A.; Ternes, T.A. Fate and behavior of progestogens in activated sludge treatment: Kinetics and transformation products. Water Res. 2021, 188, 116515. [Google Scholar] [CrossRef]
- Guedes-Alonso, R.; Ciofi, L.; Sosa-Ferrera, Z.; Santana-Rodríguez, J.J.; Bubba, M.d.; Kabir, A.; Furton, K.G. Determination of androgens and progestogens in environmental and biological samples using fabric phase sorptive extraction coupled to ultra-high performance liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2016, 1437, 116–126. [Google Scholar] [CrossRef]
- Xu, G.; Ma, S.; Tang, L.; Sun, R.; Xiang, J.; Xu, B.; Bao, Y.; Wu, M. Occurrence, fate, and risk assessment of selected endocrine disrupting chemicals in wastewater treatment plants and receiving river of Shanghai, China. Environ. Sci. Pollut. Res. Int. 2016, 23, 25442–25450. [Google Scholar] [CrossRef]
- Liu, S.; Xu, X.R.; Qi, Z.H.; Chen, H.; Hao, Q.W.; Hu, Y.X.; Zhao, J.L.; Ying, G.G. Steroid bioaccumulation profiles in typical freshwater aquaculture environments of South China and their human health risks via fish consumption. Environ. Pollut. 2017, 228, 72–81. [Google Scholar] [CrossRef]
- Scott, T.-M.; Phillips, P.J.; Kolpin, D.W.; Colella, K.M.; Furlong, E.T.; Foreman, W.T.; Gray, J.L. Pharmaceutical manufacturing facility discharges can substantially increase the pharmaceutical load to US wastewaters. Sci. Total Environ. 2018, 636, 69–79. [Google Scholar] [CrossRef]
- Miossec, C.; Lanceleur, L.; Monperrus, M. Multi-residue analysis of 44 pharmaceutical compounds in environmental water samples by solid-phase extraction coupled to liquid chromatography-tandem mass spectrometry. J. Sep. Sci. 2019, 42, 1853–1866. [Google Scholar] [CrossRef]
- Pivetta, G.G.; Gastaldini, M.D.C. Presence of emerging contaminants in urban water bodies in southern Brazil. J. Water Health 2019, 17, 329–337. [Google Scholar] [CrossRef]
- Besse, J.-P.; Garric, J. Progestagens for human use, exposure and hazard assessment for the aquatic environment. Environ. Pollut. 2009, 157, 3485–3494. [Google Scholar] [CrossRef] [Green Version]
- Kasambala, H.R.; Rwiza, M.J.; Mdegela, R.H. Levels and distribution of progesterone in receiving waters and wastewaters of a growing urban area. Water Sci. Technol. 2019, 80, 1107–1117. [Google Scholar] [CrossRef]
- Gomes, R.L.; Scrimshaw, M.D.; Lester, J.N. Fate of conjugated natural and synthetic steroid estrogens in crude sewage and activated sludge batch studies. Environ. Sci. Technol. 2009, 43, 3612–3618. [Google Scholar] [CrossRef] [Green Version]
- Blair, B.; Nikolaus, A.; Hedman, C.; Klaper, R.; Grundl, T. Evaluating the degradation, sorption, and negative mass balances of pharmaceuticals and personal care products during wastewater treatment. Chemosphere 2015, 134, 395–401. [Google Scholar] [CrossRef]
- Ojoghoro, J.O.; Scrimshaw, M.D.; Sumpter, J.P. Steroid hormones in the aquatic environment. Sci. Total Environ. 2021, 792, 148306. [Google Scholar] [CrossRef]
- Mills, L.J.; Chichester, C. Review of evidence: Are endocrine-disrupting chemicals in the aquatic environment impacting fish populations? Sci. Total Environ. 2005, 343, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Geng, J.; Huo, H.; Xu, K.; Huang, H.; Hu, H.; Ren, H. Bioaugmentated activated sludge degradation of progesterone: Kinetics and mechanism. Chem. Eng. J. 2018, 352, 214–224. [Google Scholar] [CrossRef]
- Ward, S.; Back, D.J. Metabolism of gestodene in human liver cytosol and microsomes in vitro. J. Steroid Biochem. Mol. Biol. 1993, 46, 235–243. [Google Scholar] [CrossRef]
- National Library of Medicine. Available online: https://pubchem.ncbi.nlm.nih.gov/source/hsdb/3594 (accessed on 27 December 2021).
- Kook, K.; Gabelnick, H.; Duncan, G. Pharmacokinetics of levonorgestrel 0.75 mg tablets. Contraception 2002, 66, 73–76. [Google Scholar] [CrossRef]
- DrugBank. Available online: https://www.drugbank.ca/drugs/DB00367 (accessed on 27 December 2021).
- Liu, S.; Ying, G.-G.; Liu, Y.-S.; Peng, F.-Q.; He, L.-Y. Degradation of norgestrel by bacteria from activated sludge: Comparison to progesterone. Environ. Sci. Technol. 2013, 47, 10266–10276. [Google Scholar] [CrossRef] [PubMed]
- DrugBank. Available online: https://www.drugbank.ca/drugs/DB00294 (accessed on 27 December 2021).
- Stanczyk, F.Z.; Roy, S. Metabolism of levonorgestrel, norethindrone, and structurally related contraceptive steroids. Contraception 1990, 42, 67–96. [Google Scholar] [CrossRef]
- Bick, A.J.; Louw-du Toit, R.; Skosana, S.B.; Africander, D.; Hapgood, J.P. Pharmacokinetics, metabolism and serum concentrations of progestins used in contraception. Pharmacol. Therapeut. 2021, 222, 107789. [Google Scholar] [CrossRef]
- Walker, C.J.; Cowan, D.A.; James, V.H.; Lau, J.C.; Kicman, A.T. Doping in sport: 3. Metabolic conversion of oral norethisterone to urinary 19-norandrosterone. Steroids 2009, 74, 341–349. [Google Scholar] [CrossRef]
- DrugBank. Available online: https://www.drugbank.ca/drugs/DB09123 (accessed on 27 December 2021).
- DrugBank. Available online: https://www.drugbank.ca/drugs/DB13981 (accessed on 27 December 2021).
- DrugBank. Available online: https://www.drugbank.ca/drugs/DB00603 (accessed on 27 December 2021).
- National Library of Medicine. Compound Summary for CID 6279, Medroxyprogesterone Acetate. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Medroxyprogesterone-acetate (accessed on 27 December 2021).
- National Library of Medicine. Compound Summary for CID 11683, Megestrol Acetate. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Megestrol-acetate (accessed on 27 December 2021).
- DrugBank. Available online: https://www.drugbank.ca/drugs/DB01395 (accessed on 27 December 2021).
- Stout, E.P.; La Clair, J.J.; Snell, T.W.; Shearer, T.L.; Kubanek, J. Conservation of progesterone hormone function in invertebrate reproduction. Proc. Natl. Acad. Sci. USA 2010, 107, 11859–11864. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Chung-Davidson, Y.-W.; Jia, L.; Li, W. Genomic sequence analyses of classical and non-classical lamprey progesterone receptor genes and the inference of homologous gene evolution in metazoans. BMC Evol. Biol. 2019, 19, 136. [Google Scholar] [CrossRef] [PubMed]
- Ziková, A.; Lorenz, C.; Hoffmann, F.; Kleiner, W.; Lutz, I.; Stöck, M.; Kloas, W. Endocrine disruption by environmental gestagens in amphibians—A short review supported by new in vitro data using gonads of Xenopus laevis. Chemosphere 2017, 181, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, M. Follicular Steroidogenesis in Fish, 1st ed.; CRC Press: Boca Raton, FL, USA, 2008; p. 632. [Google Scholar]
- Stacey, N.E.; Sorensen, P.W. Hormonally Derived Sex Pheromones in Fish, 1st ed.; CRC Press: Boca Raton, FL, USA, 2008; p. 632. [Google Scholar]
- Pankhurst, N.W. Gonadal Steroids: Functions and Patterns of Change, 1st ed.; CRC Press: Boca Raton, FL, USA, 2008; p. 632. [Google Scholar]
- Melcangi, R.C.; Giatti, S.; Calabrese, D.; Pesaresi, M.; Cermenati, G.; Mitro, N.; Viviani, B.; Garcia-Segura, L.M.; Caruso, D. Levels and actions of progesterone and its metabolites in the nervous system during physiological and pathological conditions. Prog. Neurobiol. 2014, 113, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Kime, D.E. A strategy for assessing the effects of xenobiotics on fish reproduction. Sci. Total Environ. 1999, 225, 3–11. [Google Scholar] [CrossRef]
- Nagahama, Y.; Yamashita, M. Regulation of oocyte maturation in fish. Dev. Growth Differ. 2008, 50, S195–S219. [Google Scholar] [CrossRef]
- NIEHS. Endocrine Disruptors. National Institute of Environmental Health Sciences. U.S. Department of Health and Human Sciences. Available online: www.niehs.nih.gov/ (accessed on 27 December 2021).
- EU. (EU) 2018/840 of 5 June 2018 establishing a watch list of substances for Union-wide monitoring in the field of water policy pursuant to Directive 2008/105/E.C. of the European Parliament and of the Council and repealing Commission Implementing Decision (EU) 2015/495. Off. J. Eur. Union 2018, 141, 9–12. [Google Scholar]
- Hoffmann, F.; Kloas, W. The synthetic progestogen, levonorgestrel, but not natural progesterone, affects male mate calling behavior of Xenopus laevis. Gen. Comp. Endocrinol. 2012, 176, 385–390. [Google Scholar] [CrossRef]
- Lorenz, C.; Contardo-Jara, V.; Pflugmacher, S.; Wiegand, C.; Nutzmann, G.; Lutz, I.; Kloas, W. The synthetic gestagen levonorgestrel impairs metamorphosis in Xenopus laevis by disruption of the thyroid system. Toxicol. Sci. 2011, 123, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Contardo-Jara, V.; Lorenz, C.; Pflugmacher, S.; Nützmann, G.; Kloas, W.; Wiegand, C. Molecular effects and bioaccumulation of levonorgestrel in the non-target organism Dreissena polymorpha. Environ. Pollut. 2011, 159, 38–44. [Google Scholar] [CrossRef]
- Orlando, E.F.; Ellestad, L.E. Sources, concentrations, and exposure effects of environmental gestagens on fish and other aquatic wildlife, with an emphasis on reproduction. Gen. Comp. Endocrinol. 2014, 203, 241–249. [Google Scholar] [CrossRef]
- Chaves-Pozo, E.; Garcia-Ayala, A.; Cabas, I. Effects of Sex Steroids on Fish Leukocytes. Biology 2018, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Frankel, T.E.; Meyer, M.T.; Kolpin, D.W.; Gillis, A.B.; Alvarez, D.A.; Orlando, E.F. Exposure to the contraceptive progestin, gestodene, alters reproductive behavior, arrests egg deposition, and masculinises development in the fathead minnow (Pimephales promelas). Environ. Sci. Technol. 2016, 50, 5991–5999. [Google Scholar] [CrossRef]
- Liang, Y.Q.; Huang, G.Y.; Lin, Z.; Li, J.; Yang, J.W.; Zhong, L.Y.; Ying, G.G. Reproductive effects of synthetic progestin norgestrel in zebrafish (Danio rerio). Chemosphere 2018, 190, 17–24. [Google Scholar] [CrossRef]
- Šauer, P.; Tumová, J.; Steinbach, C.; Golovko, O.; Komen, H.; Maillot-Maréchal, E.; Máchová, J.; Grabic, R.; Aït-Aïssa, S.; Kocour Kroupová, H. Chronic simultaneous exposure of common carp (Cyprinus carpio) from embryonic to juvenile stage to drospirenone and gestodene at low ng/L level caused intersex. Ecotoxicol. Environ. Saf. 2020, 188, 109912. [Google Scholar] [CrossRef]
- Chen, S.; Lin, C.; Tan, J.; Wang, Y.; Wang, X.; Wang, X.; Liu, L.; Li, J.; Hou, L.; Liu, J.; et al. Reproductive potential of mosquitofish is reduced by the masculinising effect of a synthetic progesterone, gestodene: Evidence from morphology, courtship behaviour, ovary histology, sex hormones and gene expressions. Sci. Total Environ. 2021, 769, 144570. [Google Scholar] [CrossRef]
- Hua, J.; Han, J.; Guo, Y.; Zhou, B. The progestin levonorgestrel affects sex differentiation in zebrafish at environmentally relevant concentrations. Aquat. Toxicol. 2015, 166, 1–9. [Google Scholar] [CrossRef]
- Overturf, M.D.; Huggett, D.B. Responses to various exposure durations of levonorgestrel during early-life stages of fathead minnows (Pimephales promelas). Aquat. Toxicol. 2015, 161, 33–40. [Google Scholar] [CrossRef]
- Runnalls, T.J.; Beresford, N.; Kugathas, S.; Margiotta-Casaluci, L.; Scholze, M.; Scott, A.P.; Sumpter, J.P. From single chemicals to mixtures—Reproductive effects of levonorgestrel and ethinylestradiol on the fathead minnow. Aquat. Toxicol. 2015, 169, 152–167. [Google Scholar] [CrossRef] [Green Version]
- Svensson, J.; Mustafa, A.; Fick, J.; Schmitz, M.; Brunström, B. Developmental exposure to progestins causes male bias and precocious puberty in zebrafish (Danio rerio). Aquat. Toxicol. 2016, 177, 316–323. [Google Scholar] [CrossRef]
- Cardoso, P.G.; Rodrigues, D.; Madureira, T.V.; Oliveira, N.; Rocha, M.J.; Rocha, E. Warming modulates the effects of the endocrine disruptor progestin levonorgestrel on the zebrafish fitness, ovary maturation kinetics and reproduction success. Environ. Pollut. 2017, 229, 300–311. [Google Scholar] [CrossRef]
- Frankel, T.E.; Meyer, M.T.; Orlando, E.F. Aqueous exposure to the progestin, levonorgestrel, alters anal fin development and reproductive behavior in the eastern mosquitofish (Gambusia holbrooki). Gen. Comp. Endocrinol. 2016, 234, 161–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankel, T.; Yonkos, L.; Frankel, J. Exposure effects of levonorgestrel on oogenesis in the fathead minnow (Pimephales promelas). Environ. Toxicol. Chem. 2017, 36, 3299–3304. [Google Scholar] [CrossRef] [PubMed]
- Maasz, G.; Zrinyi, Z.; Takacs, P.; Lovas, S.; Fodor, I.; Kiss, T.; Pirger, Z. Complex molecular changes induced by chronic progestogens exposure in roach, Rutilus rutilus. Ecotox. Environ. Saf. 2017, 139, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankel, T.; Yonkos, L.; Ampy, F.; Frankel, J. Exposure to levonorgestrel increases nest acquisition success and decreases sperm motility in the male fathead minnow (Pimephales promelas). Environ. Toxicol. Chem. 2018, 37, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, P.G.; Rodrigues, D.; Madureira, T.V.; Rocha, M.J.; Rocha, E. Histopathological evaluation of combined impacts of the synthetic progestin levonorgestrel and temperature on the female zebrafish maturation using a semi-quantitative grading analysis—is it enough? B Environ. Contam. Tox. 2018, 101, 417–422. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, K.; Fent, K. Regulation of zebrafish (Danio rerio) locomotor behavior and circadian rhythm network by environmental steroid hormones. Environ. Pollut. 2018, 232, 422–429. [Google Scholar] [CrossRef]
- Cardoso, P.G.; Resende-de-Oliveira, R.; Rocha, E. Combined effects of increased temperature and levonorgestrel exposure on zebrafish female liver, using stereology and immunohistochemistry against catalase, CYP1A, HSP90 and vitellogenin. Environ. Pollut. 2019, 252, 1059–1067. [Google Scholar] [CrossRef]
- De Courten, B.M.; Forbes, J.P.; Roark, H.K.; Burns, N.P.; Major, K.M.; White, J.W.; Li, J.; Mehinto, A.C.; Connon, R.E.; Brander, S.M. Multigenerational and transgenerational effects of environmentally relevant concentrations of endocrine disruptors in an estuarine fish model. Environ. Sci. Technol. 2020, 54, 13849–13860. [Google Scholar] [CrossRef]
- Teigeler, M.; Schaudien, D.; Böhmer, W.; Länge, R.; Schäfers, C. Effects of the gestagen levonorgestrel in a life cycle test with zebrafish (Danio rerio). Environ. Toxicol. Chem. 2021, 41, 580–591. [Google Scholar] [CrossRef]
- Pandelides, Z.; Ussery, E.J.; Overturf, M.D.; Guchardi, J.; Holdway, D.A. Inhibition of swim bladder inflation in Japanese medaka (Oryzias latipes) embryos following exposure to select pharmaceuticals alone and in combination. Aquat. Toxicol. 2021, 234, 105796. [Google Scholar] [CrossRef]
- Liang, Y.Q.; Huang, G.Y.; Zhao, J.L.; Shi, W.J.; Hu, L.X.; Tian, F.; Liu, S.S.; Jiang, Y.X.; Ying, G.G. Transcriptional alterations induced by binary mixtures of ethinylestradiol and norgestrel during the early development of zebrafish (Danio rerio). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2017, 195, 60–67. [Google Scholar] [CrossRef]
- Hou, L.P.; Chen, H.; Tian, C.E.; Liang, Y.; Wu, R.R.; Zhang, X.M.; Fang, X.W.; Zhang, C.P.; Hu, J.J.; Song, L.Y.; et al. Alterations of secondary sex characteristics, reproductive histology and behaviors by norgestrel in the western mosquitofish (Gambusia affinis). Aquat. Toxicol. 2018, 198, 224–230. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.; Chen, S.; Liu, J.; Guo, J.; Chen, Z.; Zhu, Q.; Zhang, W.; Xu, G.; Liang, Y.; Wu, R.; et al. Transcriptomic and physiological changes in western mosquitofish (Gambusia affinis) after exposure to norgestrel. Ecotoxicol. Environ. Saf. 2019, 171, 579–586. [Google Scholar] [CrossRef]
- Steinbach, C.; Cisar, P.; Sauer, P.; Klicnarova, J.; Schmidt-Posthaus, H.; Golovko, O.; Kroupova, H.K. Synthetic progestin etonogestrel negatively affects mating behavior and reproduction in Endler’s guppies (Poecilia wingei). Sci. Total Environ. 2019, 663, 206–215. [Google Scholar] [CrossRef]
- Petersen, L.H.; Hala, D.; Carty, D.; Cantu, M.; Martinović, D.; Huggett, D.B. Effects of progesterone and norethindrone on female fathead minnow (Pimephales promelas) steroidogenesis. Environ. Toxicol. Chem. 2015, 34, 379–390. [Google Scholar] [CrossRef]
- Bridges, K.N.; Magnuson, J.T.; Curran, T.E.; Barker, A.; Roberts, A.P.; Venables, B.J. Alterations to the vision-associated transcriptome of zebrafish (Danio rerio) following developmental norethindrone exposure. Environ. Toxicol. Pharmacol. 2019, 69, 137–142. [Google Scholar] [CrossRef]
- Hou, L.; Chen, S.; Shi, W.; Chen, H.; Liang, Y.; Wang, X.; Tan, J.; Wang, Y.; Deng, X.; Zhan, M.; et al. Norethindrone alters mating behaviors, ovary histology, hormone production and transcriptional expression of steroidogenic genes in zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2020, 195, 110496. [Google Scholar] [CrossRef]
- Liang, Y.Q.; Jing, Z.; Pan, C.G.; Lin, Z.; Zhen, Z.; Hou, L.; Dong, Z. The progestin norethindrone alters growth, reproductive histology and gene expression in zebrafish (Danio rerio). Chemosphere 2020, 242, 125285. [Google Scholar] [CrossRef]
- Liang, Y.Q.; Xu, W.; Liang, X.; Jing, Z.; Pan, C.G.; Tian, F. The synthetic progestin norethindrone causes thyroid endocrine disruption in adult zebrafish. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 236, 108819. [Google Scholar] [CrossRef]
- Luo, Y.; Chen, H.; Li, D.; Zhan, M.; Hou, L.; Dong, W.; Luo, Y.; Xie, L. The effects of norethindrone on the ontogeny of gene expression along the hypothalamic-pituitary-adrenal and hypothalamic-pituitary-gonadal axes in zebrafish (Danio rerio). Sci. Total Environ. 2020, 747, 141554. [Google Scholar] [CrossRef]
- Vaillant, C.; Gueguen, M.-M.; Feat, J.; Charlier, T.D.; Coumailleau, P.; Kah, O.; Brion, F.; Pellegrini, E. Neurodevelopmental effects of natural and synthetic ligands of estrogen and progesterone receptors in zebrafish eleutheroembryos. Gen. Comp. Endocrinol. 2020, 288, 113345. [Google Scholar] [CrossRef]
- Wang, X.; Tan, Z.; Chen, S.; Gui, L.; Li, X.; Ke, D.; Hou, L.; Leung, J.Y.S. Norethindrone causes cellular and hepatic injury in zebrafish by compromising the metabolic processes associated with antioxidant defence: Insights from metabolomics. Chemosphere 2021, 275, 130049. [Google Scholar] [CrossRef]
- Brown, S.; Fraga, L.R.; Cameron, G.; Erskine, L.; Vargesson, N. The primodos components norethisterone acetate and ethinyl estradiol induce developmental abnormalities in zebrafish embryos. Sci. Rep. 2018, 8, 2917. [Google Scholar] [CrossRef] [Green Version]
- Schmid, S.; Willi, R.A.; Salgueiro-González, N.; Fent, K. Effects of new generation progestins, including as mixtures and in combination with other classes of steroid hormones, on zebrafish early life stages. Sci. Total Environ. 2020, 709, 136262. [Google Scholar] [CrossRef]
- Macikova, P.; Groh, K.J.; Ammann, A.A.; Schirmer, K.; Suter, M.J. Endocrine disrupting compounds affecting corticosteroid signaling pathways in Czech and Swiss waters: Potential impact on fish. Environ. Sci. Technol. 2014, 48, 12902–12911. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Castiglioni, S.; Fent, K. Synthetic progestins medroxyprogesterone acetate and dydrogesterone and their binary mixtures adversely affect reproduction and lead to histological and transcriptional alterations in zebrafish (Danio rerio). Environ. Sci. Technol. 2015, 49, 4636–4645. [Google Scholar] [CrossRef]
- Shi, W.-J.; Ma, D.-D.; Jiang, Y.-X.; Xie, L.; Zhang, J.-N.; Huang, G.-Y.; Chen, H.-X.; Hou, L.-P.; Liu, Y.-S.; Ying, G.-G. Medroxyprogesterone acetate affects sex differentiation and spermatogenesis in zebrafish. Aquat. Toxicol. 2019, 212, 70–76. [Google Scholar] [CrossRef]
- Shi, W.J.; Huang, G.Y.; Jiang, Y.X.; Ma, D.D.; Chen, H.X.; Huang, M.Z.; Hou, L.P.; Xie, L.; Ying, G.G. Medroxyprogesterone acetate affects eye growth and the transcription of associated genes in zebrafish. Ecotoxicol. Environ. Saf. 2020, 193, 110371. [Google Scholar] [CrossRef]
- Han, J.; Wang, Q.; Wang, X.; Li, Y.; Wen, S.; Liu, S.; Ying, G.; Guo, Y.; Zhou, B. The synthetic progestin megestrol acetate adversely affects zebrafish reproduction. Aquat. Toxicol. 2014, 150, 66–72. [Google Scholar] [CrossRef]
- Hua, J.; Han, J.; Wang, X.; Guo, Y.; Zhou, B. The binary mixtures of megestrol acetate and 17α-ethynylestradiol adversely affect zebrafish reproduction. Environ. Pollut. 2016, 213, 776–784. [Google Scholar] [CrossRef]
- Hua, J.; Han, J.; Guo, Y.; Zhou, B. Endocrine disruption in Chinese rare minnow (Gobiocypris rarus) after long-term exposure to low environmental concentrations of progestin megestrol acetate. Ecotox. Environ. Saf. 2018, 163, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Blanco, M.; Fernandes, D.; Medina, P.; Blázquez, M.; Porte, C. Drospirenone intake alters plasmatic steroid levels and cyp17a1 expression in gonads of juvenile sea bass. Environ. Pollut. 2016, 213, 541–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossier, N.M.; Chew, G.; Zhang, K.; Riva, F.; Fent, K. Activity of binary mixtures of drospirenone with progesterone and 17α-ethinylestradiol in vitro and in vivo. Aquat. Toxicol. 2016, 174, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Vračko, M. Chapter 10—Mathematical (Structural) Descriptors in QSAR: Applications in Drug Design and Environmental Toxicology. In Advances in Mathematical Chemistry and Applications; Basak, S.C., Restrepo, G., Villaveces, J.L., Eds.; Bentham Science Publishers: Sharjah, United Arab Emirates, 2015; pp. 222–250. [Google Scholar]
- Kroupova, H.K.; Trubiroha, A.; Lorenz, C.; Contardo-Jara, V.; Lutz, I.; Grabic, R.; Kocour, M.; Kloas, W. The progestin levonorgestrel disrupts gonadotropin expression and sex steroid levels in pubertal roach (Rutilus rutilus). Aquat. Toxicol. 2014, 154, 154–162. [Google Scholar] [CrossRef]
- Nallani, G.C.; Paulos, P.M.; Venables, B.J.; Edziyie, R.E.; Constantine, L.A.; Huggett, D.B. Tissue-specific uptake and bioconcentration of the oral contraceptive norethindrone in two freshwater fishes. Arch. Environ. Con. Tox. 2012, 62, 306–313. [Google Scholar] [CrossRef]
- Gomez, C.F.; Constantine, L.; Huggett, D.B. The influence of gill and liver metabolism on the predicted bioconcentration of three pharmaceuticals in fish. Chemosphere 2010, 81, 1189–1195. [Google Scholar] [CrossRef]
- Steele, W.B.t.; Garcia, S.N.; Huggett, D.B.; Venables, B.J.; Barnes, S.E., 3rd; La Point, T.W. Tissue-specific bioconcentration of the synthetic steroid hormone medroxyprogesterone acetate in the common carp (Cyprinus carpio). Environ. Toxicol. Pharmacol. 2013, 36, 1120–1126. [Google Scholar] [CrossRef]
- Fick, J.; Lindberg, R.H.; Parkkonen, J.; Arvidsson, B.; Tysklind, M.; Larsson, D.G.J. Therapeutic levels of levonorgestrel detected in blood plasma of fish: Results from screening rainbow trout exposed to treated sewage effluents. Environ. Sci. Technol. 2010, 44, 2661–2666. [Google Scholar] [CrossRef]
- Huggett, D.B.; Cook, J.C.; Ericson, J.F.; Williams, R.T. A theoretical model for utilising mammalian pharmacology and safety data to prioritise potential impacts of human pharmaceuticals to fish. Hum. Ecol. Risk Assess. 2003, 9, 1789–1799. [Google Scholar] [CrossRef]
- Fitzsimmons, P.N.; Fernandez, J.D.; Hoffman, A.D.; Butterworth, B.C.; Nichols, J.W. Branchial elimination of superhydrophobic organic compounds by rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2001, 55, 23–34. [Google Scholar] [CrossRef]
- Shi, W.-J.; Zhao, J.-L.; Jiang, Y.-X.; Huang, G.-Y.; Liu, Y.-S.; Zhang, J.-N.; Ying, G.-G. Transcriptional and histological alterations in gonad of adult zebrafish after exposure to the synthetic progestin norgestrel. Environ. Toxicol. Chem. 2017, 36, 3267–3276. [Google Scholar] [CrossRef]
- Chang, H.; Wan, Y.; Wu, S.; Fan, Z.; Hu, J. Occurrence of androgens and progestogens in wastewater treatment plants and receiving river waters: Comparison to estrogens. Water Res. 2011, 42, 732–740. [Google Scholar] [CrossRef]
- Sanderson, H.; Johnson, D.J.; Wilson, C.J.; Brain, R.A.; Solomon, K.R. Probabilistic hazard assessment of environmentally occurring pharmaceuticals toxicity to fish, daphnids and algae by ECOSAR screening. Toxicol. Lett. 2003, 144, 383–395. [Google Scholar] [CrossRef]
- Von der Ohe, P.C.; Dulio, V.; Slobodnik, J.; De Deckere, E.; Kühne, R.; Ebert, R.-U.; Ginebreda, A.; De Cooman, W.; Schüürmann, G.; Brack, W. A new risk assessment approach for the prioritisation of 500 classical and emerging organic microcontaminants as potential river basin specific pollutants under the European Water Framework Directive. Sci. Total Environ. 2011, 409, 2064–2077. [Google Scholar] [CrossRef]
- Leung, H.W.; Minh, T.B.; Murphy, M.B.; Lam, J.C.W.; So, M.K.; Martin, M.; Lam, P.K.S.; Richardson, B.J. Distribution, fate and risk assessment of antibiotics in sewage treatment plants in Hong Kong, South China. Environ. Int. 2012, 42, 1–9. [Google Scholar] [CrossRef]
- ECHA. Guidance on Information Requirements and Chemical Safety Assessment: Characterisation of Dose [Concentration]-Response for Environment; European Chemicals Agency: Helsinki, Finland, 2008; Chapter R.10. [Google Scholar]
- Pickup, J.A.; Dewaele, J.; Furmanski, N.L.; Kowalczyk, A.; Luijkx, G.C.A.; Mathieu, S.; Stelter, N. A risk-based, product-level approach for assuring aquatic environmental safety of cleaning products in the context of sustainability: The Environmental Safety Check (ESC) scheme of the AISE Charter for Sustainable Cleaning. Integr. Environ. Assess. 2017, 13, 127–138. [Google Scholar] [CrossRef]
- Cano-Nicolau, J.; Garoche, C.; Hinfray, N.; Pellegrini, E.; Boujrad, N.; Pakdel, F.; Kah, O.; Brion, F. Several synthetic progestins disrupt the glial cell specific-brain aromatase expression in developing zebrafish. Toxicol. Appl. Pharm. 2016, 305, 12–21. [Google Scholar] [CrossRef]
- Liang, Y.-Q.; Huang, G.-Y.; Liu, S.-S.; Zhao, J.-L.; Yang, Y.-Y.; Chen, X.-W.; Tian, F.; Jiang, Y.-X.; Ying, G.-G. Long-term exposure to environmentally relevant concentrations of progesterone and norgestrel affects sex differentiation in zebrafish (Danio rerio). Aquat. Toxicol. 2015, 160, 172–179. [Google Scholar] [CrossRef]
- Bayer. Short-Term Reproduction Test with Dienogest (ZK 37659) on the Fathead Minnow (Pimephales promelas); Bayer: Leverkusen, Germany, 2009. [Google Scholar]
- FASS. Estradiol & Nomegestrolacetat. Available online: https://www.fass.se/LIF/product?-1.-documentTabPanel-tabs-panel-article~tools~bottom-articletools-printbiglink&userType=2&nplId=20100226000021&docType=78 (accessed on 27 December 2021).
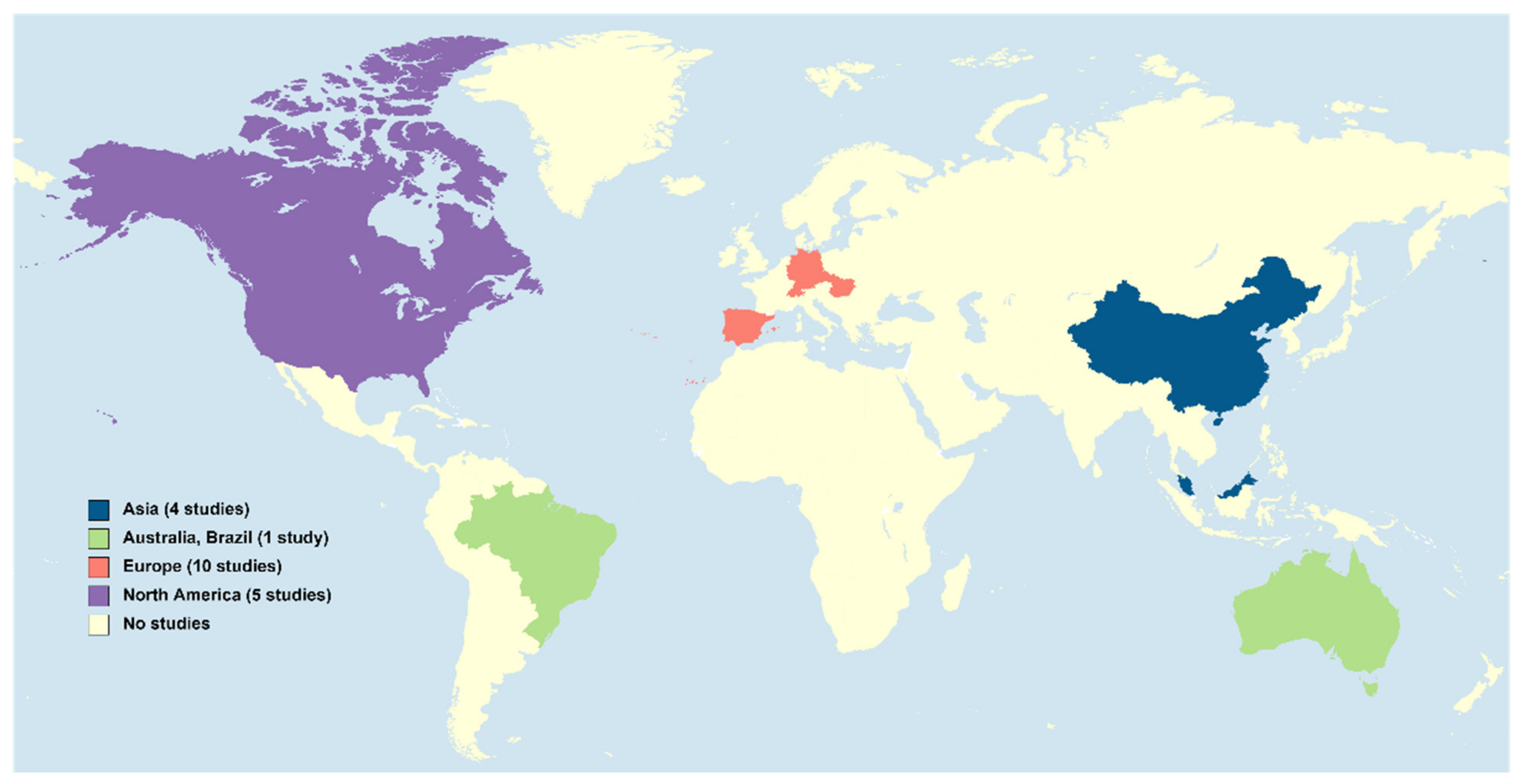
- Zucchi, S.; Castiglioni, S.; Fent, K. Progestins and antiprogestins affect gene expression in early development in zebrafish (Danio rerio) at environmental concentrations. Environ. Sci. Technol. 2012, 46, 5183–5192. [Google Scholar] [CrossRef]
Figure 1.
Locations in which studies on the levels of the synthetic PGs considered in this article were conducted in the aquatic environment from 2015 to 2021 (map generated from https://mapchart.net/world.html, accessed on 27 December 2021).
Figure 1.
Locations in which studies on the levels of the synthetic PGs considered in this article were conducted in the aquatic environment from 2015 to 2021 (map generated from https://mapchart.net/world.html, accessed on 27 December 2021).

Figure 2.
Data are expressed in boxplots with the minimum, median, maximum, average (+), and interquartile range Q1–Q3. Dots represent average individual values measured in surface waters (Sw), WWTP influent (WWTPi) and WWTP effluents (WWTPe) around the world concerning PGs derivates from (A) Testosterone (n = 42 Sw, n = 42 WWTPi, and n = 62 WWTPe), (B) Progesterone (n = 23 Sw, n = 22 WWTPi, and n = 29 WWTPe), (C) Spirolactone (n = 7 Sw, n = 7 WWTPi, and n = 9 WWTPe), (D) all PGs as a whole (n = 72 Sw, n = 71 WWTPi and n = 100 WWTPe), (E) all PGs referred in a previous review (n = 4) [7].
Figure 2.
Data are expressed in boxplots with the minimum, median, maximum, average (+), and interquartile range Q1–Q3. Dots represent average individual values measured in surface waters (Sw), WWTP influent (WWTPi) and WWTP effluents (WWTPe) around the world concerning PGs derivates from (A) Testosterone (n = 42 Sw, n = 42 WWTPi, and n = 62 WWTPe), (B) Progesterone (n = 23 Sw, n = 22 WWTPi, and n = 29 WWTPe), (C) Spirolactone (n = 7 Sw, n = 7 WWTPi, and n = 9 WWTPe), (D) all PGs as a whole (n = 72 Sw, n = 71 WWTPi and n = 100 WWTPe), (E) all PGs referred in a previous review (n = 4) [7].

Figure 3.
Sources and pathways for the occurrence of progestins in the environment. The distributions of PGs were based on Besse and Garric (2009) [51].
Figure 3.
Sources and pathways for the occurrence of progestins in the environment. The distributions of PGs were based on Besse and Garric (2009) [51].

Figure 4.
Routes of entry, circulation, major places of action, and the fate of progestins such as pregnanes, estranes, gonanes, spironolactone derivatives, and other EDCs.
Figure 4.
Routes of entry, circulation, major places of action, and the fate of progestins such as pregnanes, estranes, gonanes, spironolactone derivatives, and other EDCs.

Table 1.
Pharmacological groups of the selected progestins referred to in this article considering their structural derivation, generation, and androgenic effects in humans: (+++) highly androgenic; (++) medium androgenic; (+) low androgenic; (-) no androgenic effects.
Table 1.
Pharmacological groups of the selected progestins referred to in this article considering their structural derivation, generation, and androgenic effects in humans: (+++) highly androgenic; (++) medium androgenic; (+) low androgenic; (-) no androgenic effects.
| Testosterone derivatives | Hormone | Family | Common Name (Acronym) & CAS | Structure & Molecular Formula | Generation & Activity |
 19-Nortestosterone | Gonanes (C17) or LNG family | Gestodene (GES) 60282-87-4 |  C21H28O2 | 3rd Generation 1986 (+++) | |
| Levonorgestrel (LNG) 797-63-7 |  C21H28O2 | 2nd Generation 1966 (+++) | |||
| Norgestrel (NET) 6533-00-2 |  C21H28O2 | 2nd Generation 1966 (+++) | |||
| Etonogestrel (ENG) 54048-10-1 |  C22H28O2 | 3rd Generation 1998 (+) | |||
Estranes (C18) or NTD family | Norethisterone (NTD) 68-22-4 |  C20H26O2 | 1st Generation 1951 (++) | ||
| Norethisterone acetate (NTDA) 51-98-9 |  C22H28O3 | 1st Generation 1951 (++) | |||
| Dienogest (DIE) 65928-58-7 |  C22H28O2 | 4th Generation 1978 (-) | |||
Progesterone derivatives |  19-Norprogesterone | Norpregnanes (C20) | Nomegestrol acetate (NOMAC) 58652-20-3 |  C23H30O4 | 4th Generation 1986 (-) |
 17α-Hydroxyprogesterone | Pregnanes (C21) | Medroxyprogesterone (MEP) 520-85-4 |  C22H32O3 | 1st Generation 1957 (+) | |
| Medroxyprogesterone acetate (MPA) 71-58-9 |  C24H34O4 | 1st Generation 1957 (+) | |||
| Megestrol acetate (MGA) 595-33-5 |  C22H30O4 | 1st Generation 1963 (-) | |||
| Spironolactone derivative |  Spironolactone | Drospirenone (DSP) 67392-87-4 |  C24H30O3 | 4th Generation 1976 (-) | |
Table 3.
Worldwide WWTPs removal efficiency of PGs. In bold are shown the situations when the treated effluent contains higher amounts of a certain PG than its influent (these values were set apart from the global average percentage of removal, % RAv.). Quantification method (QM); WWTP influents (WWTPi); WWTP effluents (WWTPe); % Removal (% R).
Table 3.
Worldwide WWTPs removal efficiency of PGs. In bold are shown the situations when the treated effluent contains higher amounts of a certain PG than its influent (these values were set apart from the global average percentage of removal, % RAv.). Quantification method (QM); WWTP influents (WWTPi); WWTP effluents (WWTPe); % Removal (% R).
| Testosterone derivatives (Gonanes) | PGs | QM | WWTPi (ng/L) | WWTPe (ng/L) | % R | % RAv. | Local | References |
| GES | (1) | <3.0 | <1.0 | 66.7 | 82.9 | WWTPs (Switzerland). | [31] | |
| (2) | 6.6 | 0.5 | 92.8 | WWTPs (Czech Republic). | [32] | |||
| (2) | 4.3 | 0.77 | 95.5 | WWTPs (Czech and Slovak Republics). | [35] | |||
| (3) | 3.0 | 1.0 | 66.7 | WWTPs (Switzerland). | [36] | |||
| (2) | <0.38 | <0.49 | −28.9 | −186 | WWTPs (Czech Republic). | [32] | ||
| (2) | <0.79 | <3.5 | −343 | WWTPs (Czech and Slovak Republics). | [35] | |||
| LNG | (1) | 652 | 35.5 | 94.4 | 62.4 | WWTPs (Malaysia). | [38] | |
| (2) | <1.8 | 0.5 | 72.4 | WWTPs (Czech Republic). | [32] | |||
| (2) | 0.43 | 0.20 | 54.3 | WWTPs (Czech and Slovak Republics). | [35] | |||
| (1) | 58.6 | 26.0 | 53.7 | WWTPs (Canada). | [42] | |||
| (4) | Data from bibliography | 37.0 | WWTPs (China). | [34] | ||||
| (2) | <0.26 | <0.53 | −103.8 | −104 | WWTPs (Czech Republic). | [32] | ||
| NET | (4) | Data from bibliography | 96.0 | 96.0 | WWTPs (China). | [34] | ||
| ENG | (2) | 1.3 | 0.6 | 58.7 | 42.9 | WWTPs (Czech Republic). | [32] | |
| (2) | 0.52 | 0.38 | 27.1 | WWTPs (Czech and Slovak Republics). | [35] | |||
| NTD | (1) | 1093 | 242 | 78.0 | 75.7 | WWTPs (Malaysia). | [38] | |
| (6) | Data from bibliography | 98 | WWTPs (China). | [46] | ||||
| (1) | <3.0 | <0.6 | 80.0 | WWTPs (Switzerland). | [31] | |||
| (2) | 0.14 | 0.08 | 39 | WWTPs (Czech and Slovak Republics). | [35] | |||
| (3) | <3.0 | <0.6 | 80.0 | WWTPs (Switzerland). | [36] | |||
| (1) | 4.8 | 2.0 | 58.3 | WWTPs (Canada). | [42] | |||
| (5) | 78.8 | 31.0 | 59.6 | WWTPs (Canada). | [43] | |||
| (4) | Data from bibliography | > 90 | WWTPs (China). | [34] | ||||
| (2) | 0.1 | 0.4 | −225 | −600 | WWTPs (Czech Republic). | [32] | ||
| (2) | 0.2 | 2.15 | −975 | WWTPs (Czech and Slovak Republics). | [35] | |||
| NTDA | (4) | Data from bibliography | >90.0 | >90.0 | WWTPs (China). | [34] | ||
| DIE | (1) | <0.8 | <0.3 | 62.5 | 83.0 | WWTPs (Switzerland). | [31] | |
| (3) | <0.8 | <0.3 | 62.5 | WWTPs (Switzerland). | [36] | |||
| (2) | 6.5 | 0.1 | 95.9 | WWTPs (Czech Republic). | [32] | |||
| (2) | 6.4 | 0.30 | 95.3 | WWTPs (Czech and Slovak Republics). | [35] | |||
| (2) | 3.9 | 4 | −2.6 | −2.6 | WWTPs (Czech and Slovak Republics). | [35] | ||
| Progesterone derivatives | NOMAC | (2) | 1.3 | 0.1 | 72.0 | 72.0 | WWTPs (Czech Republic). | [32] |
| MEP | (1) | 6 | 3 | 50.0 | 52.6 | WWTPs (Switzerland). | [31] | |
| (3) | 6 | 3 | 50.0 | WWTPs (Switzerland). | [36] | |||
| (2) | 0.1 | 0.04 | 46.8 | WWTPs (Czech Republic). | [32] | |||
| (2) | 0.2 | 0.05 | 72.9 | WWTPs (Czech and Slovak Republics). | [35] | |||
| (2) | <0.02 | 0.23 | −1050 | −725 | WWTPs (Czech Republic). | [32] | ||
| (2) | 0.19 | 0.95 | −400 | WWTPs (Czech and Slovak Republics). | [35] | |||
| MPA | (1) | Data from bibliography | 93.0 | 76.4 | WWTPs (Switzerland). | [31] | ||
| (2) | 2.4 | 0.3 | 71.0 | WWTPs (Czech Republic). | [32] | |||
| (2) | 2.2 | 0.22 | 90.1 | WWTPs (Czech and Slovak Republics). | [35] | |||
| (3) | 3.1 | 0.2 | 85.6 | WWTPs (Switzerland). | [36] | |||
| (4) | Data from bibliography | 24.0 | WWTPs (China). | [34] | ||||
| MGA | (1) | Data from bibliography | 99.6 | 78.2 | WWTPs (Switzerland). | [31] | ||
| (2) | 6.4 | 0.3 | 95.3 | WWTPs (Czech and Slovak Republics). | [35] | |||
| (3) | <0.03 | 0.4 | 93.7 | WWTPs (Czech Republic). | [32] | |||
| (4) | Data from bibliography | 24.0 | WWTPs (China). | [34] | ||||
| Spironolactone derivative | DSP | (1) | <4.0 | <1.0 | 75.0 | 61.5 | WWTPs (Switzerland). | [31] |
| (2) | 0.7 | 0.4 | 49.0 | WWTPs (Czech Republic). | [32] | |||
| (2) | 3.5 | 0.1 | 88.2 | WWTPs (Czech and Slovak Republics). | [35] | |||
| (3) | <4.0 | <1.0 | 75.0 | WWTPs (Switzerland). | [36] | |||
| (4) | Data from bibliography | 42.0 | WWTPs (China). | [34] | ||||
Note: Average values for WWTPi and WWTPe > 0 were calculated for Refs. [32,35,36,38,42]. Values of <0, corresponding to the last references are shown in bold. (1) Liquid chromatography with tandem mass spectrometry detection (LC-MS/MS); (2) liquid chromatography tandem atmospheric pressure chemical ionization/atmospheric pressure photoionization with hybrid quadrupole/orbital trap mass spectrometry operated in high-resolution product scan mode (LC-APCI/APPI-HRPS); (3) high-performance liquid chromatography coupled to triple quadrupole mass spectrometry (HPLC-MS/MS); (4) ultra-performance liquid chromatography coupled with tandem mass detection (UPLC-MS/MS); (5) triple quadrupole-linear ion trap mass spectrometer using the sMRM (scheduled multiple reaction monitoring) mode (TripleQuad-LIT-MS); (6) rapid resolution liquid chromatography/tandem mass spectrometry (RRLC-MS/MS).
Table 4.
Main metabolisation organ and enzymes, elimination routes, and the number of active metabolites for the PGs referred to in this study. Data not available (n.a.).
Table 4.
Main metabolisation organ and enzymes, elimination routes, and the number of active metabolites for the PGs referred to in this study. Data not available (n.a.).
| Testosterone derivatives (Gonanes) | PGs | Main Metabolization(s) Route(s) | Elimination Route(s) | Active Metabolites | References |
| GES | Liver. Metabolisation occurs by CYP3A4 via partial or total reduction of the A-ring. | Urine and faeces at a ratio of about 6:4. | n.a. | [58,59] | |
| LNG | Liver. Metabolisation by CYP3A4 and CYP3A5. | Urine (45%). Faeces (32%). | In sludge, LNG metabolites generate four active molecules. | [60,61] | |
| NET | Liver. NET is converted to LNG. Then, it follows the same metabolisation paths of LNG. | Urine (46%). Faeces (32%). | In sludge, NET metabolites generate four active molecules. | [61,62] | |
| ENG | Liver Metabolization by CYP3A4. | n.a. | n.a. | [63] | |
| Testosterone derivatives Estranes) | NTD | Liver. Metabolisation occurs via partial or total reduction of the A-ring and oxidation by CYP3A4 and, to a much lesser extent, CYP2C19, CYP1A2, and CYP2A6. | Urine (50%). Faeces (20–40%). | The most known and active biologic metabolite is EE2. | [44,64,65,66] |
| NTDA | Liver. NTDA is converted to NTD. Then, it follows the same metabolisation paths of NTD. | Urine (50%). Faeces (20–40%). | The most known and active biologic metabolite is EE2. | [44,64,65,66] | |
| DIE | Liver. Metabolisation by P450 enzymes. | Urine and faeces at a ratio of about 3:1. | The metabolites are all inactive. | [67] | |
| Progesterone derivatives | NOMAC | n.a. | n.a. | n.a. | [68] |
| MEP | Liver. Metabolisation occurs via partial or total reduction of the A-ring. May happen side-chain reduction, loss of the acetyl group, hydroxylation in the 2-, 6-, and 21-positions or a combination of these positions. | Urine. | More than ten active metabolites. | [69] | |
| MPA | Liver. MPA is converted to MEP. Then, it follows the same metabolisation paths of MEP. | Urine. | More than ten active metabolites. | [44,69,70] | |
| MGA | Liver. | Urine. Respiratory excretion. Fat storage. | n.a. | [71] | |
| Spironolactone derivative | DSP | Liver. Metabolisation occurs by the opening of the lactone ring, known as M11, followed by the action of CYP3A4. | Urine (38–47%). Faeces (17–20%). | n.a. | [72] |
Table 5.
Data retrieved from “Web of Science Core Collection”, covering years from 2015 to 2021, concerning the effects of the synthetic PGs in fish. No data were available (n.a.) for NOMAC.
Table 5.
Data retrieved from “Web of Science Core Collection”, covering years from 2015 to 2021, concerning the effects of the synthetic PGs in fish. No data were available (n.a.) for NOMAC.
| Testosterone derivatives (Gonanes) | PGs | Structural or Functional Impact on Fish | References |
| GES | Induction of masculinisation in fathead minnow (Pimephales promelas). | [89] | |
| Reproductive disorders in zebrafish (Danio rerio). | [90] | ||
| Induction of intersex in common carp (Cyprinus carpio). | [91] | ||
| Masculinisation, potential reproduction reduction in mosquitofish (Gambusia affinis). | [92] | ||
| LNG | Interfere with sex differentiation in zebrafish (D. rerio). | [93] | |
| Decrease of larval growth and expression of 20β-HSD and CYP19A1, FSH and 3β-HSD in fathead minnows (P. promelas). | [94] | ||
| Inhibition of egg production in fathead minnows (P. promelas). | [95] | ||
| Induction of precocious puberty in zebrafish (D. rerio). | [96] | ||
| Alteration of fitness, ovary maturation kinetics and reproduction success in zebrafish (D. rerio). | [97] | ||
| Change of the anal fin development and reproductive behaviour in mosquitofish (Gambusia holbrooki). | [98] | ||
| Modification of oogenesis in fathead minnow (P. promelas). | [99] | ||
| Induction of metabolic disorders in roach (Rutilus rutilus). | [100] | ||
| Rise of nest acquisition success and loss of sperm motility in fathead minnow (P. promelas). | [101] | ||
| Decrease of mature oocytes in zebrafish (D. rerio). | [97,102] | ||
| Alteration of circadian gene regulation in zebrafish (D. rerio). | [103] | ||
| Alteration of liver function in zebrafish (D. rerio). | [104] | ||
| Transgenerational effects in inland silverside (Menidia beryllina). | [105] | ||
| Decrease of post-hatch survival in zebrafish (D. rerio). | [106] | ||
| Inhibition of swim bladder inflation in Japanese medaka (Oryzias latipes) embryos. | [107] | ||
| NET | Transcriptional alterations in early development in zebrafish (D. rerio). | [108] | |
| Alteration of secondary sex characteristics, reproductive histology, and behaviours in mosquitofish (G. affinis). | [109] | ||
| Transcriptomic and physiological changes in adult mosquitofish (G. affinis). | [110] | ||
| ENG | Change of mating behaviour and reproduction in Endler’s guppies (Poecilia wingei). | [111] | |
| Testosterone derivatives (Estranes) | NTD | Alteration of steroidogenesis in female fathead minnow (P. promelas). | [112] |
| Alteration of sex differentiation in zebrafish (D. rerio). | [93] | ||
| Alteration of circadian gene regulation in zebrafish (D. rerio). | [103] | ||
| Alter the development of visual function in zebrafish (D. rerio). | [113] | ||
| Induction of masculinisation and hepatopathological disorders in female mosquitofish (G. affinis). | [110] | ||
| Alteration of mating behaviours, ovary histology and hormone production in zebrafish (D. rerio). | [114] | ||
| Alters growth, reproductive histology, and gene expression in zebrafish (D. rerio). | [115] | ||
| Thyroid endocrine disruption in zebrafish (D. rerio). | [116] | ||
| Interfere with the HPG and the hypothalamic-pituitary-adrenal (HPA) axis in zebrafish (D. rerio). | [117] | ||
| Neurodevelopmental effects in zebrafish (D. rerio). | [118] | ||
| Hepatic injury in zebrafish (D. rerio). | [119] | ||
| NTDA | Induction of developmental abnormalities in zebrafish (D. rerio). | [120] | |
| DIE | Minor transcriptional alterations in zebrafish (D. rerio) early life stages. | [121] | |
| Progesterone derivatives | NOMAC | n.a. | n.a. |
| MEP | Potential endocrine disruptor in fish. | [122] | |
| MPA | Reproductive disorders (gonadal histology) in zebrafish (D. rerio). | [123] | |
| Affects sex differentiation and spermatogenesis in zebrafish (D. rerio). | [124] | ||
| Affects eye growth in zebrafish (D. rerio). | [125] | ||
| MGA | Reproductive disorders of zebrafish (D. rerio). | [126] | |
| Alters ovary histology of zebrafish (D. rerio). | [127] | ||
| Endocrine disruption in Chinese rare minnow (Gobiocypris rarus). | [128] | ||
| Spironolactone derivative | DSP | Alter plasma steroid levels and CYP17A1 expression in gonads of juvenile sea bass (Dicentrarchus labrax). | [129] |
| Ethinylestradiol antagonist in zebrafish (D. rerio) embryos. | [130] | ||
| Metabolic disorders in roach (R. rutilus). | [100] | ||
| Together with GES induces intersex of common carp (C. carpio). | [91] |
Table 6.
Bioconcentration factor in fish plasma (BCFFP) and concentration in the plasma of a fish (CFP), which correspond to the human plasma therapeutical levels, and predicted effect concentration (PECw) values. Data in bold are above PECw, considering the average between the minimum and the maximal levels measured in surface waters (Sw), WWTP influents (WWTPi) and effluents (WWTPe) presented in Table 2.
Table 6.
Bioconcentration factor in fish plasma (BCFFP) and concentration in the plasma of a fish (CFP), which correspond to the human plasma therapeutical levels, and predicted effect concentration (PECw) values. Data in bold are above PECw, considering the average between the minimum and the maximal levels measured in surface waters (Sw), WWTP influents (WWTPi) and effluents (WWTPe) presented in Table 2.
| PGs | Log Kow | BCFFP | CFP (ng/mL) | PECw (ng/L) | Sw (ng/L) | WWTPi (ng/L) | WWTPe (ng/L) |
|---|---|---|---|---|---|---|---|
| aGES | 3.26 | 32 | 1.0 | 31 | 10.8 | 10.9 | 10.8 |
| aLNG | 3.48 | 46 | 2.4 | 52 | 58.5 | 405 | 19.5 |
| bNET | 3.48 | 46 | - | 6.7 | - | 11.2 | 2.0 |
| aENG | 3.16 | 27 | 0.8 | 29 | 0.2 | 0.8 | 0.7 |
| aNTD | 2.97 | 19 | 9.8 | 516 | 115 | 568 | 132 |
| aNTDA | 3.99 | 108 | 9.8 | 91 | 0.3 | 10.5 | 0.62 |
| aDIE | 2.34 | 7 | 85.2 | 12,171 | 1.2 | 6.4 | 2.2 |
| aNOMAC | 3.55 | 52 | 7.2 | 138 | 0.1 | 1.8 | 0.1 |
| aMEP | 3.50 | 47 | 1 | 21 | 0.7 | 3.0 | 1.5 |
| aMPA | 4.09 | 128 | 1 | 8 | 0.2 | 4.1 | 0.3 |
| cMGA | 3.20 | 29 | - | - | 10.0 | 6.5 | 30.0 |
| aDSP | 4.02 | 113 | 30.8 | 273 | 2.2 | 3.5 | 0.5 |
Table 7.
Assessment factors used for PNECs derivation [145].
Table 7.
Assessment factors used for PNECs derivation [145].
| Available Data | Assessment Factor (AF) |
|---|---|
| One short-term E(L)C50 from each of the three trophic levels (fish, crustaceans, or algae). | 1000 |
| One long-term NOEC assay (either fish, crustaceans, or algae). | 100 |
| Two long-term NOEC assays considering species from two trophic levels (fish and/or crustaceans and/or algae). | 50 |
| Three long-term NOEC assays considering species from three trophic levels (fish, crustaceans and algae). | 10 |
| Species Sensitivity Distribution (SSD) method | 5–1 |
| Field data or model ecosystems. | Evaluated on a case-by-case basis. |
Table 8.
Risk quotients (RQs) for 9 of the 12 PGs referred to in this study using the considering the average between the minimum and the maximal levels found in surface waters from 2015 to 2021. RQ values were not calculated for NTDA due to the absence of MEC and for NOMAC and MEP due to the lack of endpoint values for fish.
Table 8.
Risk quotients (RQs) for 9 of the 12 PGs referred to in this study using the considering the average between the minimum and the maximal levels found in surface waters from 2015 to 2021. RQ values were not calculated for NTDA due to the absence of MEC and for NOMAC and MEP due to the lack of endpoint values for fish.
| PGs | Endpoint Value (ng/L) Fish | PNEC (ng/L) | MEC (ng/L) | RQs | Risk | References |
|---|---|---|---|---|---|---|
| GES | EC50 = 10; AF = 1000 | 0.01 | 10.8 | 1078 | High | [89] |
| LNG | NOEC = 0.42; AF = 50 | 0.01 | 59 | 6967 | High | [43,95] |
| NET | LOEC = 6.0; AF = 1000 | 0.01 | - | - | [90] | |
| ENG | EC50 = 12,654; AF = 1000 | 12.7 | 0.2 | 0 | Low | [146] |
| NTD | NOEC = 4; AF = 50 | 0.08 | 115 | 1438 | High | [43,147] |
| NTDA | NOEC = 816; AF = 1000 | 0.8 | 0.3 | 0 | Low | [43,147] |
| DIE | NOEC = 44; AF = 1000 | 0.04 | 1.2 | 36 | High | [148] |
| NOMAC | NOEC = 1300; AF = 10 | 130 | 0.1 | 0 | Low | [149] |
| MEP | - | - | 0.7 | - | - | - |
| MPA | NOEC = 342; AF = 50 | 6.8 | 0.2 | 0 | Low | [43,123] |
| MGA | NOEC = 33; AF = 50 | 0.7 | 10.0 | 15 | High | [43,126] |
| DSP | NOEC = 100; AF = 50 | 2.0 | 2.2 | 1.1 | High | [43,150] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rocha, M.J.; Rocha, E. Synthetic Progestins in Waste and Surface Waters: Concentrations, Impacts and Ecological Risk. Toxics 2022, 10, 163. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040163
AMA Style
Rocha MJ, Rocha E. Synthetic Progestins in Waste and Surface Waters: Concentrations, Impacts and Ecological Risk. Toxics. 2022; 10(4):163. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040163
Chicago/Turabian StyleRocha, Maria João, and Eduardo Rocha. 2022. "Synthetic Progestins in Waste and Surface Waters: Concentrations, Impacts and Ecological Risk" Toxics 10, no. 4: 163. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040163
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.