
Antidiabetic Actions of Ethanol Extract of Camellia sinensis Leaf Ameliorates Insulin Secretion, Inhibits the DPP-IV Enzyme, Improves Glucose Tolerance, and Increases Active GLP-1 (7–36) Levels in High-Fat-Diet-Fed Rats

 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Extraction
2.2. In Vitro Studies on Insulin Release
2.3. Membrane Potential and Intracellular Calcium Ions Concentration
2.4. Assay for Cellular Glucose Uptake
2.5. Insulin Glycation
2.6. In Vitro Dipeptidyl Peptidase-IV Enzyme Activity
2.7. In Vitro Digestion of Starch
2.8. In Vitro Glucose Diffusion
2.9. Animals
2.10. Acute Oral Glucose Tolerance Test
2.11. In Vivo Dipeptidyl Peptidase-IV Enzyme Activity
2.12. Feeding Test
2.13. Metabolic Studies
2.14. Gut Motility
2.15. Statistical Analysis
2.16. Phytochemical Screening
3. Results
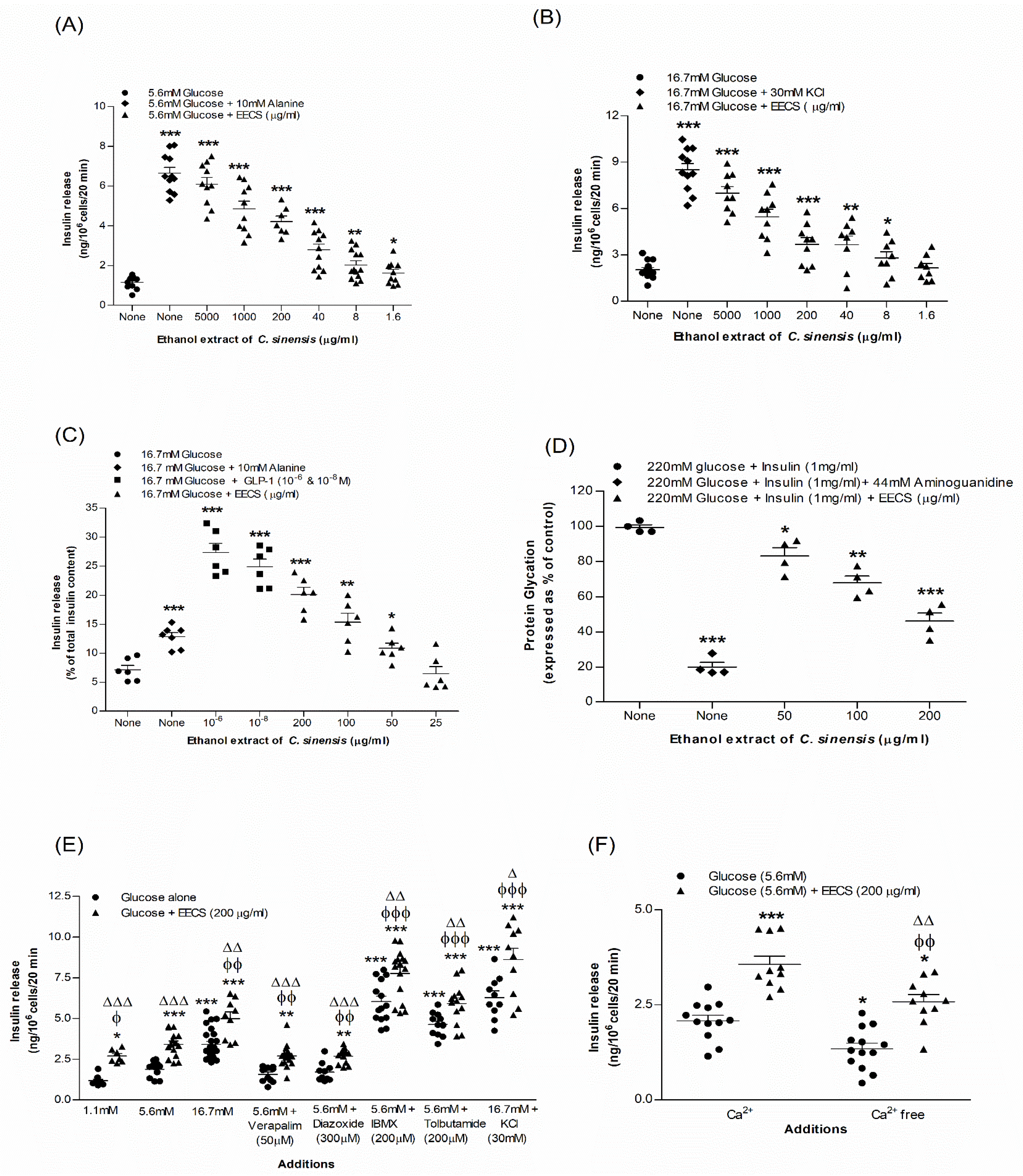
3.1. EECS and Insulin Release from BRIN BD11 Cells
3.2. EECS and Insulin Release from Isolated Mouse Islets
3.3. EECS and Known Modulators/Inhibitors of Insulin Release
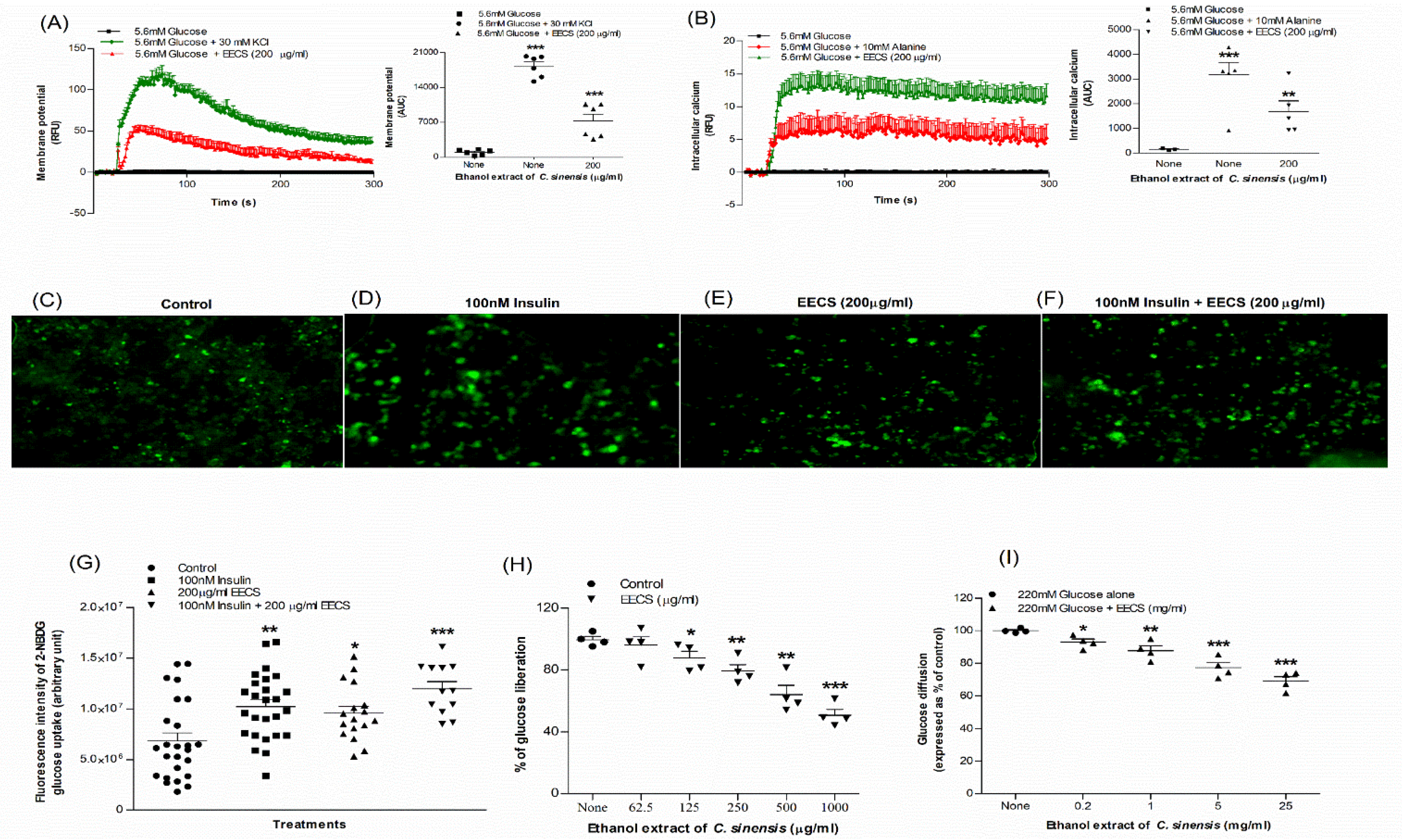
3.4. EECS and Cell Membrane Depolarization and [Ca2+]i Concentration
3.5. EECS and Insulin Glycation
3.6. EECS and Glucose Uptake
3.7. EECS and Starch Digestion
3.8. EECS and In Vitro Glucose Diffusion
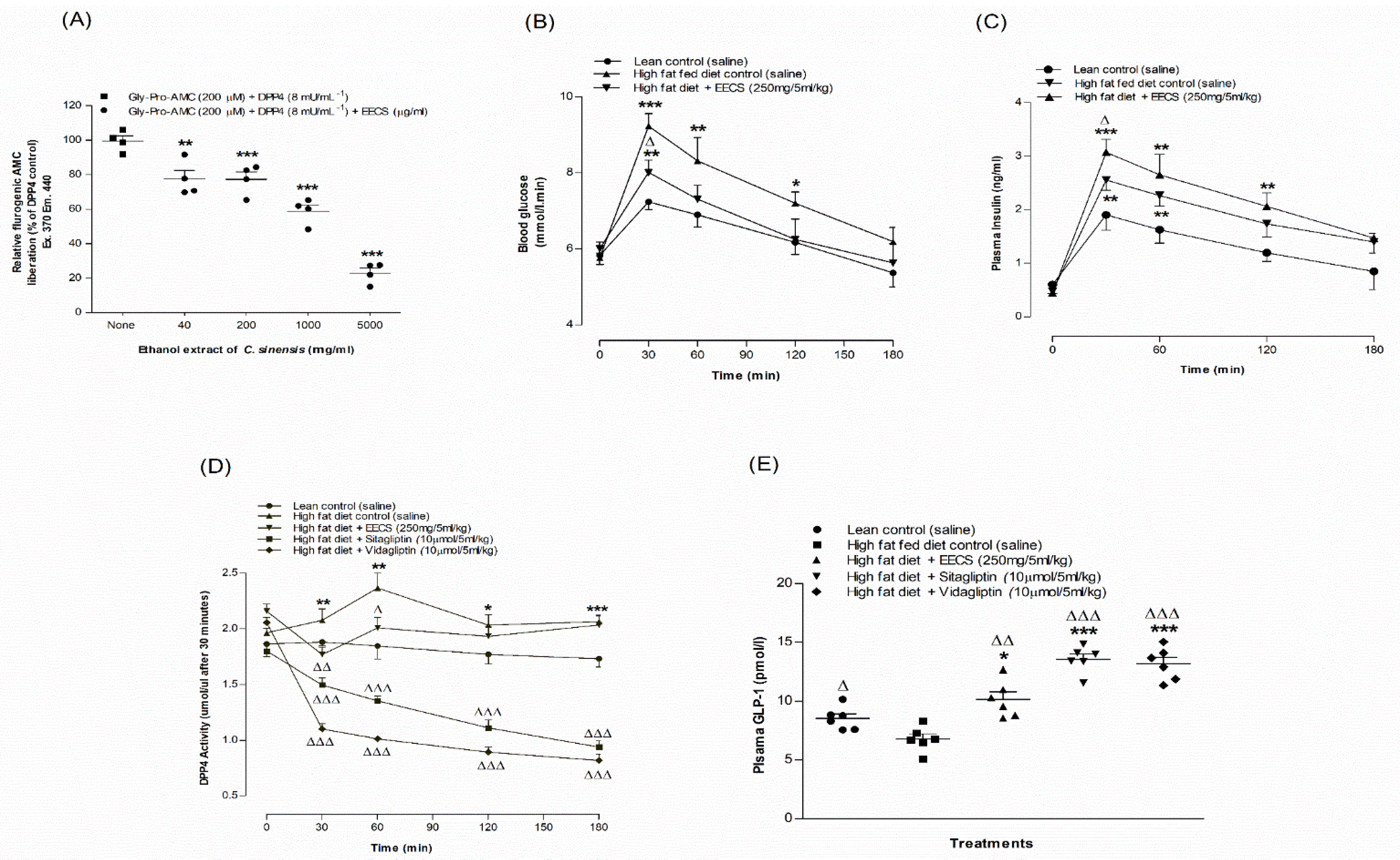
3.9. EECS and In Vitro Dipeptidyl Peptidase-IV Enzyme Activity
3.10. EECS and Oral Glucose Tolerance and Plasma Insulin Levels
3.11. EECS and Plasma DPP-IV Enzyme Activity and Active GLP-1 (7–36) Levels
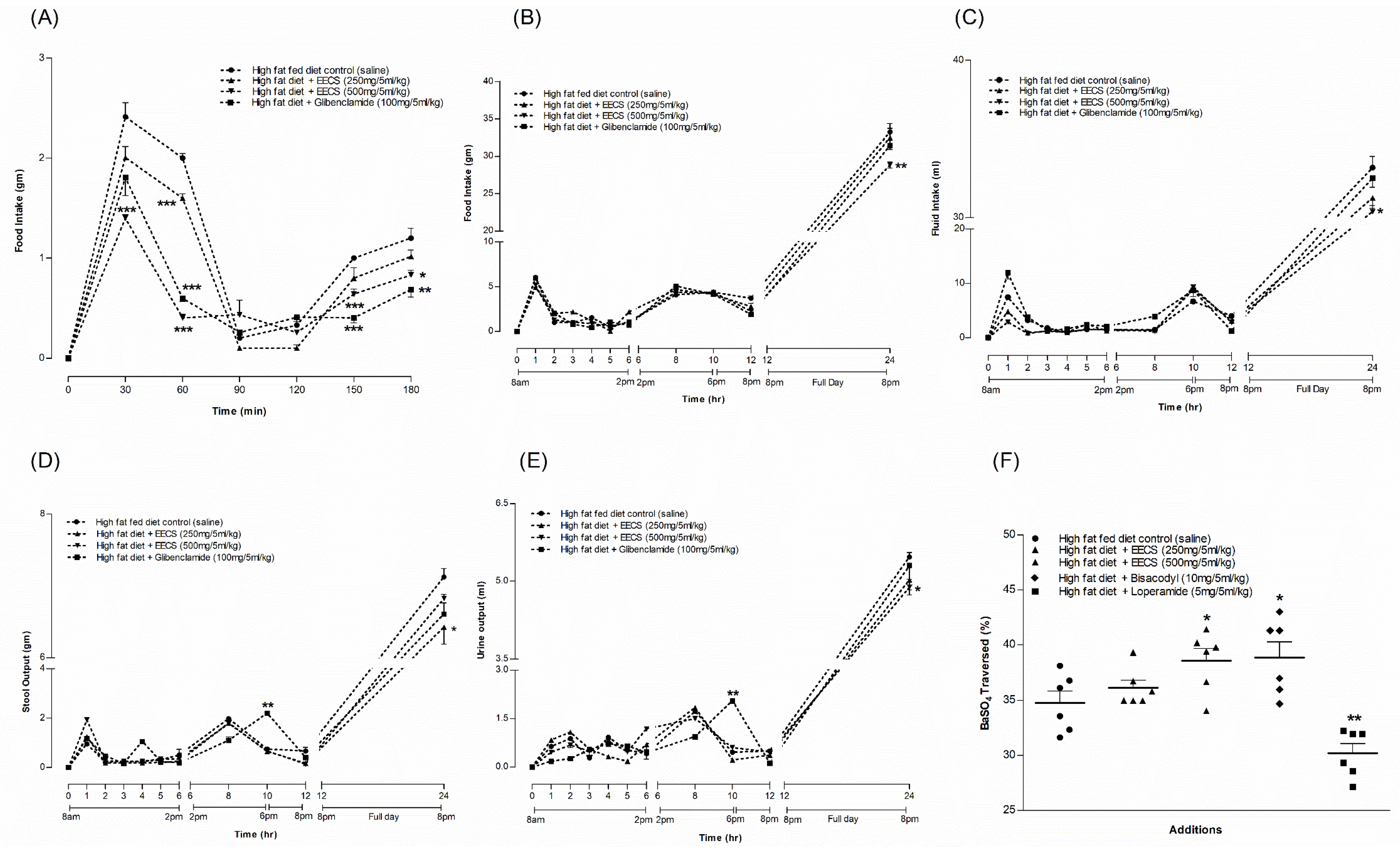
3.12. EECS and Feeding Test
3.13. EECS and Metabolic Parameters
3.14. EECS and Gastrointestinal Motility
3.15. EECS and Phytochemical Screening
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Modak, M.; Dixit, P.; Londhe, J.; Ghaskadbi, S.; Devasagayam, T.P.A. Indian Herbs and Herbal Drugs Used for the Treatment of Diabetes. J. Clin. Biochem. Nutr. 2007, 40, 163–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scartezzini, P.; Speroni, E. Review on Some Plants of Indian Traditional Medicine with Antioxidant Activity. J. Ethnopharmacol. 2000, 71, 23–43. [Google Scholar] [CrossRef]
- Singh, N.; Kesherwani, R.; Tiwari, A.K.; Patel, D.K. A Review on Diabetes Mellitus. J. Pharm. Innov. 2016, 5, 36–40. [Google Scholar]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, Regional and Country-Level Diabetes Prevalence Estimates for 2021 and Projections for 2045. Diabetes Res. Clin. Pract. 2021, 183, 109119. [Google Scholar] [CrossRef]
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes. Diabetes Care 2014, 38 (Suppl. 1), S8–S16. [Google Scholar] [CrossRef] [Green Version]
- Ansari, P.; Flatt, P.R.; Harriott, P.; Abdel-Wahab, Y.H. Anti-hyperglycaemic and insulin-releasing effects of Camellia sinensis leaves and isolation and characterization of active compounds. Brit. J. Nutr. 2020, 126, 1149–1163. [Google Scholar] [CrossRef]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms Linking Obesity to Insulin Resistance and Type 2 Diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef]
- Piché, M.-E.; Tchernof, A.; Després, J.-P. Obesity Phenotypes, Diabetes, and cardiovascular diseases. Circ. Res. 2020, 126, 1477–1500. [Google Scholar] [CrossRef]
- Kovesdy, C.P.; Furth, S.L.; Zoccali, C. Obesity and Kidney Disease. Can. J. Kidney Health Dis. 2017, 4, 205435811769866. [Google Scholar] [CrossRef] [Green Version]
- Ansari, P.; Azam, S.; Hannan, J.M.A.; Flatt, P.R.; Abdel Wahab, Y.H.A. Anti-Hyperglycaemic Activity of H. Rosa-Sinensis Leaves Is Partly Mediated by Inhibition of Carbohydrate Digestion and Absorption, and Enhancement of Insulin Secretion. J. Ethnopharmacol. 2020, 253, 112647. [Google Scholar] [CrossRef]
- Vella, A. Mechanism of Action of DPP-4 Inhibitors—New Insights. J. Clin. Endocrinol. Metab. 2012, 97, 2626–2628. [Google Scholar] [CrossRef]
- Flatt, P.R. Dipeptidyl Peptidase IV (DPP IV) and Related Molecules in Type 2 Diabetes. Front. Biosci. 2008, 13, 3648. [Google Scholar] [CrossRef] [Green Version]
- Seino, Y.; Fukushima, M.; Yabe, D. GIP and GLP-1, the Two Incretin Hormones: Similarities and Differences. J. Diabetes Investig. 2010, 1, 8–23. [Google Scholar] [CrossRef] [Green Version]
- Hannan, J.M.A.; Ansari, P.; Azam, S.; Flatt, P.R.; Abdel Wahab, Y.H.A. Effects of Spirulina platensis on Insulin Secretion, Dipeptidyl Peptidase IV Activity and Both Carbohydrate Digestion and Absorption Indicate Potential as an Adjunctive Therapy for Diabetes. Brit. J. Nutr. 2020, 124, 1021–1034. [Google Scholar] [CrossRef]
- Saeed, M.; Naveed, M.; Arif, M.; Kakar, M.U.; Manzoor, R.; Abd El-Hack, M.E.; Alagawany, M.; Tiwari, R.; Khandia, R.; Munjal, A.; et al. Green Tea (Camellia sinensis) and L -Theanine: Medicinal Values and Beneficial Applications in Humans—A Comprehensive Review. Biomed. Pharmacother. 2017, 95, 1260–1275. [Google Scholar] [CrossRef]
- Bahorun, T.; Luximon-Ramma, A.; Neergheen-Bhujun, V.S.; Gunness, T.K.; Googoolye, K.; Auger, C.; Crozier, A.; Aruoma, O.I. The Effect of Black Tea on Risk Factors of Cardiovascular Disease in a Normal Population. Prev. Med. 2012, 54, S98–S102. [Google Scholar] [CrossRef]
- Hakim, I.A.; Alsaif, M.A.; Alduwaihy, M.; Al-Rubeaan, K.; Al-Nuaim, A.R.; Al-Attas, O.S. Tea Consumption and the Prevalence of Coronary Heart Disease in Saudi Adults: Results from a Saudi National Study. Prev. Med. 2003, 36, 64–70. [Google Scholar] [CrossRef]
- Fu, Q.-Y.; Li, Q.-S.; Lin, X.-M.; Qiao, R.-Y.; Yang, R.; Li, X.-M.; Dong, Z.-B.; Xiang, L.-P.; Zheng, X.-Q.; Lu, J.-L.; et al. Antidiabetic Effects of Tea. Molecules 2017, 22, 849. [Google Scholar] [CrossRef] [Green Version]
- Ekayanti, M.; Sauriasari, R.; Elya, B. Dipeptidyl Peptidase IV Inhibitory Activity of Fraction from White Tea Ethanolic Extract (Camellia sinensis (L.) Kuntze) Ex Vivo. Pharmacogn. J. 2017, 10, 190–193. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, Y.; Watanabe, M.; Morikawa-Ichinose, T.; Fujino, K.; Yamamoto, M.; Nishioka, S.; Inoue, C.; Ogawa, F.; Yonekura, M.; Nakasone, A.; et al. Metabolic Profiling for Evaluating the Dipeptidyl Peptidase-IV Inhibitory Potency of Diverse Green Tea Cultivars and Determining Bioactivity-Related Ingredients and Combinations. J. Agric. Food Chem. 2022, 70, 6455–6466. [Google Scholar] [CrossRef]
- Nag, A.; Dhull, N.; Gupta, A. Evaluation of Tea (Camellia sinensis L.) Phytochemicals as Multi-Disease Modulators, a Multidimensional in Silico Strategy with the Combinations of Network Pharmacology, Pharmacophore Analysis, Statistics and Molecular Docking. Mol. Divers. 2022, 26, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Barreira, S.; Moutinho, C.; Silva, A.M.N.; Neves, J.; Seo, E.-J.; Hegazy, M.-E.F.; Efferth, T.; Gomes, L.R. Phytochemical Characterization and Biological Activities of Green Tea (Camellia sinensis) Produced in the Azores, Portugal. Phytomed. Plus 2021, 1, 100001. [Google Scholar] [CrossRef]
- Fan, J.; Johnson, M.H.; Lila, M.A.; Yousef, G.; de Mejia, E.G. Berry and Citrus Phenolic Compounds Inhibit Dipeptidyl Peptidase IV: Implications in Diabetes Management. Evid. Based Complement. Altern. Med. 2013, 2013, 479505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aboulwafa, M.M.; Youssef, F.S.; Gad, H.A.; Altyar, A.E.; Al-Azizi, M.M.; Ashour, M.L. A Comprehensive Insight on the Health Benefits and Phytoconstituents of Camellia sinensis and Recent Approaches for Its Quality Control. Antioxidants 2019, 8, 455. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.-K.; Lin, S.-R.; Chang, C.-H.; Tsai, M.-J.; Lee, D.-N.; Weng, C.-F. Natural Phenolic Compounds Potentiate Hypoglycemia via Inhibition of Dipeptidyl Peptidase IV. Sci. Rep. 2019, 9, 15585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, P.; Hannon-Fletcher, M.P.; Flatt, P.R.; Abdel-Wahab, Y.H. Effects of 22 Traditional Anti-Diabetic Medicinal Plants on DPP-IV Enzyme Activity and Glucose Homeostasis in High-Fat Fed Obese Diabetic Rats. Biosci. Rep. 2021, 41, BSR20203824. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, Z.; Zhao, X.; Xie, H.; Du, L.; Gao, H.; Xie, C. Mechanisms of Kaempferol in the Treatment of Diabetes: A Comprehensive and Latest Review. Front. Endocrinol. 2022, 13, 990299. [Google Scholar] [CrossRef]
- Rakhmat, I.I.; Yuslianti, E.R.; Koswara, T. Flavonoid-Rutin Effect to Blood Glucose Level and Pancreas Regeneration in Diabetic Rats. In Proceedings of the 2th Annual Scientific Meeting, Medical Faculty, Universitas Jenderal Achmad Yani, International Symposium on “Emergency Preparedness and Disaster Response during COVID 19 Pandemic” (ASMC 2021), Virtual, 10–12 May 2021; pp. 64–66. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, S.-T.; Yin, Y.-C.; Xing, S.; Li, W.-N.; Fu, X.-Q. Hypoglycemic Effect and Mechanism of Isoquercitrin as an Inhibitor of Dipeptidyl Peptidase-4 in Type 2 Diabetic Mice. RSC Adv. 2018, 8, 14967–14974. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Vaghari-Tabari, M.; Malakoti, F.; Moein, S.; Qujeq, D.; Yousefi, B.; Asemi, Z. Quercetin: An Effective Polyphenol in Alleviating Diabetes and Diabetic Complications. Crit. Rev. Food Sci. Nutr. 2022, 1–24. [Google Scholar] [CrossRef]
- Abdulkhaleq, L.A.; Assi, M.A.; Noor, M.H.M.; Abdullah, R.; Saad, M.Z.; Taufiq-Yap, Y.H. Therapeutic Uses of Epicatechin in Diabetes and Cancer. Vet. World 2017, 10, 869–872. [Google Scholar] [CrossRef]
- Elbandrawy, M.M.; Sweef, O.; Elgamal, D.; Mohamed, T.M.; EhabTousson; Elgharabawy, R.M. Ellagic Acid Regulates Hyperglycemic State through Modulation of Pancreatic IL-6 and TNF- α Immunoexpression. Saudi J. Biol. Sci. 2022, 29, 3871–3880. [Google Scholar] [CrossRef]
- Wolfram, S.; Raederstorff, D.; Preller, M.; Wang, Y.; Teixeira, S.R.; Riegger, C.; Weber, P. Epigallocatechin Gallate Supplementation Alleviates Diabetes in Rodents. J. Nutr. 2006, 136, 2512–2518. [Google Scholar] [CrossRef] [Green Version]
- Ansari, P.; Azam, S.; Seidel, V.; Abdel-Wahab, Y.H.A. In vitro and in vivo antihyperglycemic activity of the ethanol extract of Heritiera fomes bark and characterization of pharmacologically active phytomolecules. J. Pharm. Pharmacol. 2022, 3, 415–425. [Google Scholar] [CrossRef]
- Ansari, P.; Flatt, P.R.; Harriott, P.; Abdel-Wahab, Y.H.A. Insulin secretory and antidiabetic actions of Heritiera fomes bark together with isolation of active phytomolecules. PLoS ONE 2022, 17, e0264632. [Google Scholar] [CrossRef]
- McClenaghan, N.H.; Barnett, C.R.; O’Harte, F.P.M.; Flatt, P.R. Mechanisms of Amino Acid-Induced Insulin Secretion from the Glucose-Responsive BRIN-BD11 Pancreatic B-Cell Line. J. Endocrinol. 1996, 151, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Ojo, O.O.; Srinivasan, D.K.; Owolabi, B.O.; Vasu, S.; Conlon, J.M.; Flatt, P.R.; Abdel-Wahab, Y.H.A. Esculentin-2CHa-Related Peptides Modulate Islet Cell Function and Improve Glucose Tolerance in Mice with Diet-Induced Obesity and Insulin Resistance. PLoS ONE 2015, 10, e0141549. [Google Scholar] [CrossRef]
- Flatt, P.R.; Bailey, C.J. Abnormal plasma glucose and insulin responses in heterozygous lean (ob/+) mice. Diabetologia 1981, 20, 573–577. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Wahab, Y.H.A.; Marenah, L.; Flatt, P.R.; Conlon, J.M. Insulin releasing properties of the Temporin family of antimicrobial peptides. Protein Pept. Lett. 2007, 14, 702–707. [Google Scholar] [CrossRef]
- Hannan, J.M.A.; Ali, L.; Khaleque, J.; Akhter, M.; Flatt, P.R.; Abdel-Wahab, Y.H.A. Antihyperglycemic activity of Asparagus racemosus roots is partly mediated by inhibition of carbohydrate digestion and absorption, and enhancement of cellular insulin action. Brit. J. Nutri. 2012, 107, 1316–1323. [Google Scholar] [CrossRef] [Green Version]
- Ansari, P.; Flatt, P.R.; Harriott, P.; Abdel-Wahab, Y.H. Insulinotropic and antidiabetic properties of Eucalyptus citriodora leaves and isolation of bioactive phytomolecules. J. Pharm. Pharmacol. 2021, 73, 1049–1061. [Google Scholar] [CrossRef]
- O’Harte, F.P.M.; Højrup, P.; Barnett, C.R.; Flatt, P.R. Identification of the Site of Glycation of Human Insulin. Peptides 1996, 17, 1323–1330. [Google Scholar] [CrossRef]
- Duffy, N.A.; Green, B.D.; Irwin, N.; Gault, V.A.; McKillop, A.M.; O’Harte, F.P.M.; Flatt, P.R. Effects of Antidiabetic Drugs on Dipeptidyl Peptidase IV Activity: Nateglinide Is an Inhibitor of DPP IV and Augments the Antidiabetic Activity of Glucagon-like Peptide-1. Eur. J. Pharmacol. 2007, 568, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Thomson, H.; Ojo, O.; Flatt, P.; AbdelWahab, Y. Antidiabetic actions of aqueous bark extract of Swertia chirayita on insulin secretion, cellular glucose uptake and protein glycation. J. Exp. Integrat. Med. 2014, 4, 268. [Google Scholar] [CrossRef]
- Gallagher, A.M.; Flatt, P.R.; Duffy, G.; Abdel-Wahab, Y.H.A. The effects of traditional antidiabetic plants on in vitro glucose diffusion. Nutri. Res. 2003, 23, 413–424. [Google Scholar] [CrossRef]
- Ansari, P.; Flatt, P.R.; Harriott, P.; Abdel-Wahab, Y.H.A. Evaluation of the Antidiabetic and Insulin Releasing Effects of A. squamosa, Including Isolation and Characterization of Active Phytochemicals. Plants 2020, 9, 1348. [Google Scholar] [CrossRef]
- Hannan, J.M.A.; Ansari, P.; Haque, A.; Sanju, A.; Huzaifa, A.; Rahman, A.; Ghosh, A.; Azam, S. Nigella sativa Stimulates Insulin Secretion from Isolated Rat Islets and Inhibits the Digestion and Absorption of (CH2O)N in the Gut. Biosci. Rep. 2019, 39, BSR20190723. [Google Scholar] [CrossRef] [Green Version]
- Ansari, P.; Badhan, S.S.; Azam, S.; Sultana, N.; Anwar, S.; Mohamed Abdurahman, M.S.; Hannan, J.M.A. Evaluation of Antinociceptive and Anti-Inflammatory Properties of Methanolic Crude Extract of Lophopetalum javanicum (Bark). J. Basic Clin. Physiol. Pharmacol. 2016, 27, 379–385. [Google Scholar] [CrossRef]
- Tabish, S.A. Is Diabetes Becoming the Biggest Epidemic of the Twenty-First Century? Int. J. Health Sci. 2007, 1, V–VIII. [Google Scholar]
- Grover, J.K.; Yadav, S.; Vats, V. Medicinal Plants of India with Anti-Diabetic Potential. J. Ethnopharmacol. 2002, 81, 81–100. [Google Scholar] [CrossRef]
- Salehi, B.; Ata, A.; Anil Kumar, N.V.; Sharopov, F.; Ramirez-Alarcon, K.; Ruiz-Ortega, A.; Abdulmajid Ayatollahi, S.; Valere Tsouh Fokou, P.; Kobarfard, F.; Amiruddin Zakaria, Z. Antidiabetic Potential of Medicinal Plants and Their Active Components. Biomolecules 2019, 9, 551. [Google Scholar] [CrossRef] [Green Version]
- Patel, D.; Prasad, S.; Kumar, R.; Hemalatha, S. An Overview on Antidiabetic Medicinal Plants Having Insulin Mimetic Property. Asian Pac. J. Trop. Biomed. 2012, 2, 320–330. [Google Scholar] [CrossRef]
- Dietary Fibre and Incidence of Type 2 Diabetes in Eight European Countries: The EPIC-InterAct Study and a Meta-Analysis of Prospective Studies. Diabetologia 2015, 58, 1394–1408. [CrossRef] [Green Version]
- Chopade, V.V.; Phatak, A.A.; Upaganlawar, A.B.; Tankar, A.A. Green Tea (Camellia sinensis): Chemistry, Traditional, Medicinal Uses and Its Pharmacological Activities- a Review. Pharmacogn. Rev. 2008, 2, 157–162. [Google Scholar]
- Haidari, F.; Omidian, K.; Rafiei, H.; Zarei, M.; Mohamad Shahi, M. Green Tea (Camellia sinensis) Supplementation to Diabetic Rats Improves Serum and Hepatic Oxidative Stress Markers. Iran. J. Pharm. Res. 2013, 12, 109–114. [Google Scholar]
- Islam, M.S. Effects of the Aqueous Extract of White Tea (Camellia sinensis) in a Streptozotocin-Induced Diabetes Model of Rats. Phytomedicine 2011, 19, 25–31. [Google Scholar] [CrossRef]
- Ferreira, M.C.L.; Lima, L.N.; Cota, L.H.T.; Costa, M.B.; Orsi, P.M.E.; Espíndola, R.P.; Albanez, A.V.; Rosa, B.B.; Carvalho, M.G.S.; Garcia, J.A.D. Effect of Camellia sinensis Teas on Left Ventricular Hypertrophy and Insulin Resistance in Dyslipidemic Mice. Braz. J. Med. Biol. 2020, 53, e9303. [Google Scholar] [CrossRef]
- Sola, D.; Rossi, L.; Schianca, G.P.C.; Maffioli, P.; Bigliocca, M.; Mella, R.; Corlianò, F.; Fra, G.P.; Bartoli, E.; Derosa, G. Sulfonylureas and Their Use in Clinical Practice. Arch. Med. Sci. 2015, 4, 840–848. [Google Scholar] [CrossRef] [Green Version]
- Lambadiari, V.; Triantafyllou, K.; Dimitriadis, G.D. Insulin Action in Muscle and Adipose Tissue in Type 2 Diabetes: The Significance of Blood Flow. World J. Diabetes 2015, 6, 626. [Google Scholar] [CrossRef] [Green Version]
- Ashcroft, F.M.; Rorsman, P. KATP Channels and Islet Hormone Secretion: New Insights and Controversies. Nat. Rev. Endocrinol. 2013, 9, 660–669. [Google Scholar] [CrossRef] [Green Version]
- Park, J.E.; Han, J.S. A Portulaca oleracea L. extract promotes insulin secretion via a K+ATP channel dependent pathway in INS-1 pancreatic β-cells. Nutr. Res. Pract. 2018, 12, 183–190. [Google Scholar] [CrossRef]
- Billington, C.K.; Ojo, O.O.; Penn, R.B.; Ito, S. CAMP Regulation of Airway Smooth Muscle Function. Pulm. Pharmacol. Ther. 2013, 26, 112–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeFronzo, R.A.; Tripathy, D. Skeletal Muscle Insulin Resistance Is the Primary Defect in Type 2 Diabetes. Diabetes Care 2009, 32, S157–S163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.-T.; Song, Z.; Zhang, W.-C.; Jiao, B.; Yu, Z.-B. Impaired Translocation of GLUT4 Results in Insulin Resistance of Atrophic Soleus Muscle. Biomed Res. Int. 2015, 2015, 291987. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.-K.; Gao, J.; Zhu, D.-N. Kaempferol and Quercetin Isolated from Euonymus Alatus Improve Glucose Uptake of 3T3-L1 Cells without Adipogenesis Activity. Life Sci. 2008, 82, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Sayem, A.; Arya, A.; Karimian, H.; Krishnasamy, N.; Ashok Hasamnis, A.; Hossain, C. Action of Phytochemicals on Insulin Signaling Pathways Accelerating Glucose Transporter (GLUT4) Protein Translocation. Molecules 2018, 23, 258. [Google Scholar] [CrossRef] [Green Version]
- Gonbad, R.A.; Afzan, A.; Karimi, E.; Sinniah, U.R.; Swamy, M.K. Phytoconstituents and Antioxidant Properties among Commercial Tea (Camellia sinensis L.) Clones of Iran. Electron. J. Biotechnol. 2015, 18, 433–438. [Google Scholar] [CrossRef] [Green Version]
- Sukito, A.; Tachibana, S. Isolation of Hyperoside and Isoquercitrin from Camellia sasanqua as Antioxidant Agents. Pak. J. Biol. Sci. 2014, 17, 999–1006. [Google Scholar] [CrossRef]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced Glycation End Products and Diabetic Complications. Korean J. Physiol. Pharmacol. 2014, 18, 1. [Google Scholar] [CrossRef] [Green Version]
- Setyawan, E.I.; Setyowati, E.P.; Rohman, A.; Nugroho, A.K. Simultaneous Determination of Epigallocatechin gallate, Catechin, and Caffeine from Green Tea Leaves (Camellia sinensis L.) Extract by RP-HPLC. Res. J. Pharm. Technol. 2020, 13, 1489. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, G.; Hu, X.; Pan, J.; Liao, Y.; Ding, H. Inhibitory Effect of Epicatechin Gallate on Protein Glycation. Int. Food Res. J. 2019, 122, 230–240. [Google Scholar] [CrossRef]
- Jang, D.; Kim, J.; Kim, J.; Yoo, J.; Kim, Y.; Kim, J. Effects of Compounds Isolated from the Fruits ofRumex Japonicuson the Protein Glycation. Chem. Biodivers. 2008, 5, 2718–2723. [Google Scholar] [CrossRef]
- Dias, D.T.M.; Palermo, K.R.; Motta, B.P.; Kaga, A.K.; Lima, T.F.O.; Brunetti, I.L.; Baviera, A.M. Rutin Inhibits the in Vitroformation of Advanced Glycation Products and Protein Oxidation More Efficiently than Quercetin. Rev. Cienc. Farm. Basica Apl. 2021, 42, e718. [Google Scholar] [CrossRef]
- Falla, N.M.; Demasi, S.; Caser, M.; Scariot, V. Phytochemical Profile and Antioxidant Properties of Italian Green Tea, a New High Quality Niche Product. Horticulturae 2021, 7, 91. [Google Scholar] [CrossRef]
- Gromova, L.V.; Fetissov, S.O.; Gruzdkov, A.A. Mechanisms of Glucose Absorption in the Small Intestine in Health and Metabolic Diseases and Their Role in Appetite Regulation. Nutrients 2021, 13, 2474. [Google Scholar] [CrossRef]
- Takahama, U.; Hirota, S. Interactions of Flavonoids with α-Amylase and Starch Slowing down Its Digestion. Food Funct. 2018, 9, 677–687. [Google Scholar] [CrossRef]
- Wickramasinghe, A.S.D.; Kalansuriya, P.; Attanayake, A.P. Herbal Medicines Targeting the Improved β-Cell Functions and β-Cell Regeneration for the Management of Diabetes Mellitus. Evid. Based Complementary Altern. Med. 2021, 2021, 2920530. [Google Scholar] [CrossRef]
- Al-Goblan, A.S.; Al-Alfi, M.A.; Khan, M.Z. Mechanism linking diabetes mellitus and obesity. Diabetes Metab. Syndr. Obes. Targets Ther. 2014, 7, 587. [Google Scholar] [CrossRef] [Green Version]
- McKillop, A.M.; Duffy, N.A.; Lindsay, J.R.; Green, B.D.; Patterson, S.; O’Harte, F.P.M.; Bell, P.M.; Flatt, P.R. Insulinotropic Actions of Nateglinide in Type 2 Diabetic Patients and Effects on Dipeptidyl Peptidase-IV Activity and Glucose-Dependent Insulinotropic Polypeptide Degradation. Eur. J. Endocrinol. 2009, 161, 877–885. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Zhang, Q.; Zhang, C.; Yang, W.; Liu, H.; Lv, Z.; Liu, J.; Jiao, Z. Inhibition of Dipeptidyl Peptidase-4 by Flavonoids: Structure–Activity Relationship, Kinetics and Interaction Mechanism. Front. Nutr. 2022, 9, 892426. [Google Scholar] [CrossRef]
- Golic, M.; Kräker, K.; Fischer, C.; Alenina, N.; Haase, N.; Herse, F.; Schütte, T.; Henrich, W.; Müller, D.N.; Busjahn, A.; et al. Continuous Blood Glucose Monitoring Reveals Enormous Circadian Variations in Pregnant Diabetic Rats. Front. Endocrinol. 2018, 9, 271. [Google Scholar] [CrossRef]
- AL-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and Their Anti-Diabetic Effects: Cellular Mechanisms and Effects to Improve Blood Sugar Levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Zhang, M.; Wang, D.; Yu, F.; Zhang, N.; Song, C.; Granato, D. Analytical Strategy Coupled to Chemometrics to Differentiate Camellia sinensis Tea Types Based on Phenolic Composition, Alkaloids, and Amino Acids. J. Food Sci. 2020, 85, 3253–3263. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ho, C.; Zhou, J.; Santos, J.S.; Armstrong, L.; Granato, D. Chemistry and Biological Activities of Processed Camellia sinensis Teas: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1474–1495. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, M.; Zohari, F.; Sadeghi, H. Antioxidant and Protective Effects of Major Flavonoids from Teucrium Poliumon β-Cell Destruction in a Model of Streptozotocin-Induced Diabetes. Planta Medica 2009, 75, 1418–1420. [Google Scholar] [CrossRef] [PubMed]
- Babujanarthanam, R.; Kavitha, P.; Pandian, M.R. Quercitrin, a Bioflavonoid Improves Glucose Homeostasis in Streptozotocin-Induced Diabetic Tissues by Altering Glycolytic and Gluconeogenic Enzymes. Fundam. Clin. Pharmacol. 2009, 24, 357–364. [Google Scholar] [CrossRef]
- Dubey, S.; Ganeshpurkar, A.; Ganeshpurkar, A.; Bansal, D.; Dubey, N. Glycolytic Enzyme Inhibitory and Antiglycation Potential of Rutin. Future J. Pharm. Sci. 2017, 3, 158–162. [Google Scholar] [CrossRef]
- Muhammad, I.; Rahman, N.; Gul-E-Nayab; Nishan, U.; Shah, M. Antidiabetic Activities of Alkaloids Isolated from Medicinal Plants. Braz. J. Pharm. Sci. 2021, 57. [Google Scholar] [CrossRef]
- Li, Y.; Park, J.; Wu, Y.; Cui, J.; Jia, N.; Xi, M.; Wen, A. Identification of AMPK Activator from Twelve Pure Compounds Isolated from Aralia taibaiensis: Implication in Antihyperglycemic and Hypolipidemic Activities. Korean J. Physiol. Pharmacol. 2017, 21, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Muthusamy, V.S.; Anand, S.; Sangeetha, K.N.; Sujatha, S.; Arun, B.; Lakshmi, B.S. Tannins Present in Cichorium Intybus Enhance Glucose Uptake and Inhibit Adipogenesis in 3T3-L1 Adipocytes through PTP1B Inhibition. Chem. Biol. Interact. 2008, 174, 69–78. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Test | Observation |
|---|---|
| Alkaloids | + |
| Tannins | + |
| Saponins | + |
| Steroids | - |
| Glycoside | - |
| Flavonoids | + |
| Reducing Sugar | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ansari, P.; Hannan, J.M.A.; Choudhury, S.T.; Islam, S.S.; Talukder, A.; Seidel, V.; Abdel-Wahab, Y.H.A. Antidiabetic Actions of Ethanol Extract of Camellia sinensis Leaf Ameliorates Insulin Secretion, Inhibits the DPP-IV Enzyme, Improves Glucose Tolerance, and Increases Active GLP-1 (7–36) Levels in High-Fat-Diet-Fed Rats. Medicines 2022, 9, 56. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines9110056
Ansari P, Hannan JMA, Choudhury ST, Islam SS, Talukder A, Seidel V, Abdel-Wahab YHA. Antidiabetic Actions of Ethanol Extract of Camellia sinensis Leaf Ameliorates Insulin Secretion, Inhibits the DPP-IV Enzyme, Improves Glucose Tolerance, and Increases Active GLP-1 (7–36) Levels in High-Fat-Diet-Fed Rats. Medicines. 2022; 9(11):56. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines9110056
Chicago/Turabian StyleAnsari, Prawej, J. M. A. Hannan, Samara T. Choudhury, Sara S. Islam, Abdullah Talukder, Veronique Seidel, and Yasser H. A. Abdel-Wahab. 2022. "Antidiabetic Actions of Ethanol Extract of Camellia sinensis Leaf Ameliorates Insulin Secretion, Inhibits the DPP-IV Enzyme, Improves Glucose Tolerance, and Increases Active GLP-1 (7–36) Levels in High-Fat-Diet-Fed Rats" Medicines 9, no. 11: 56. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines9110056