Impact of Dual Cell Co-culture and Cell-conditioned Media on Yield and Function of a Human Olfactory Cell Line for Regenerative Medicine

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Primary Cell Isolation and Culture
2.2. Cell Line Generation and Culture
2.3. Feeder Layer Generation
2.4. Collection of Conditioned Media
2.5. Culture of PA5 Cells
2.6. In Vitro Co-culture with NG108-15 Neurons
2.7. Immunocytochemistry (ICC)
2.8. Circularity Analysis
2.9. Neuronal Growth Analysis
2.10. Statistical Analysis and Data Accessibility
3. Results and Discussion
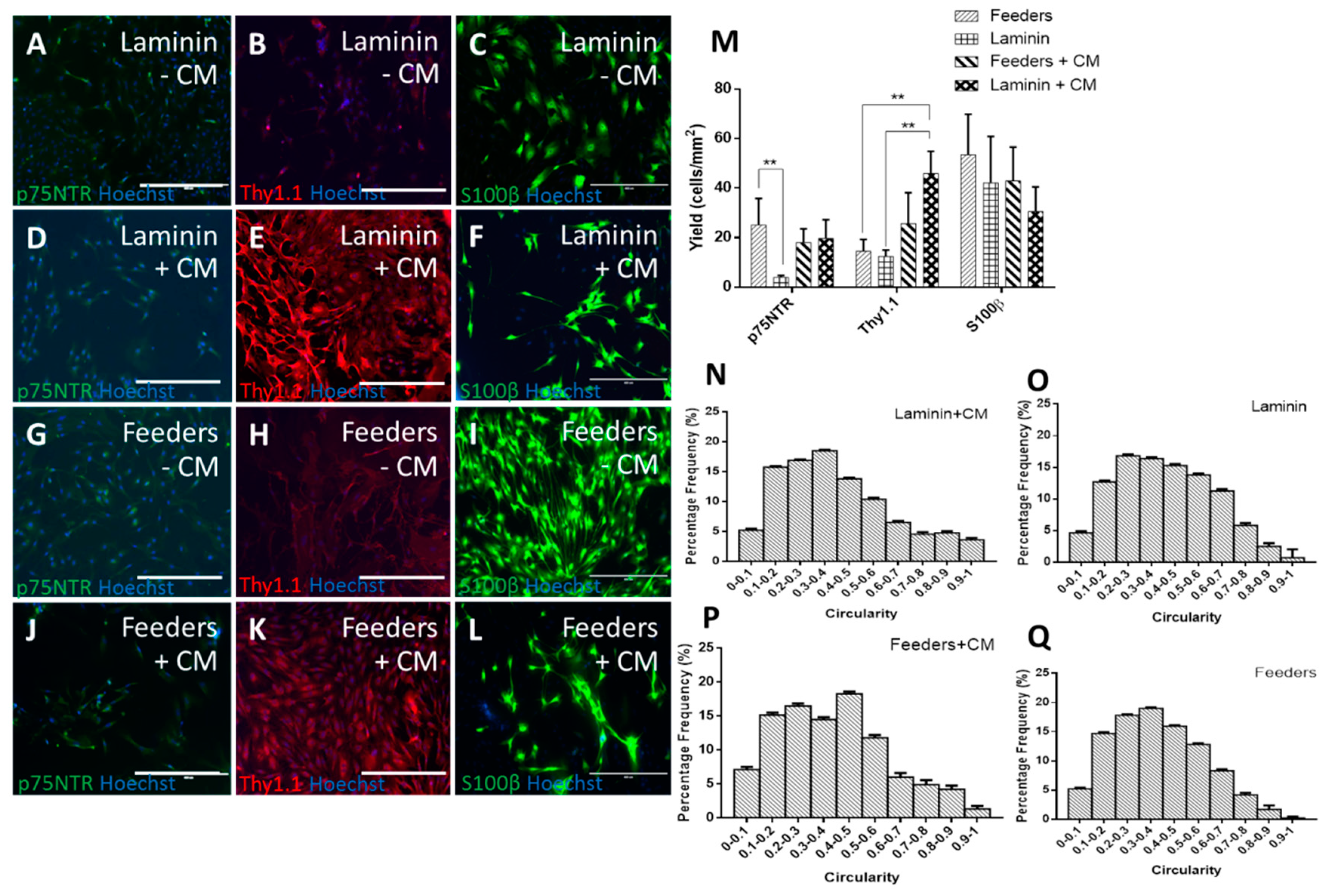
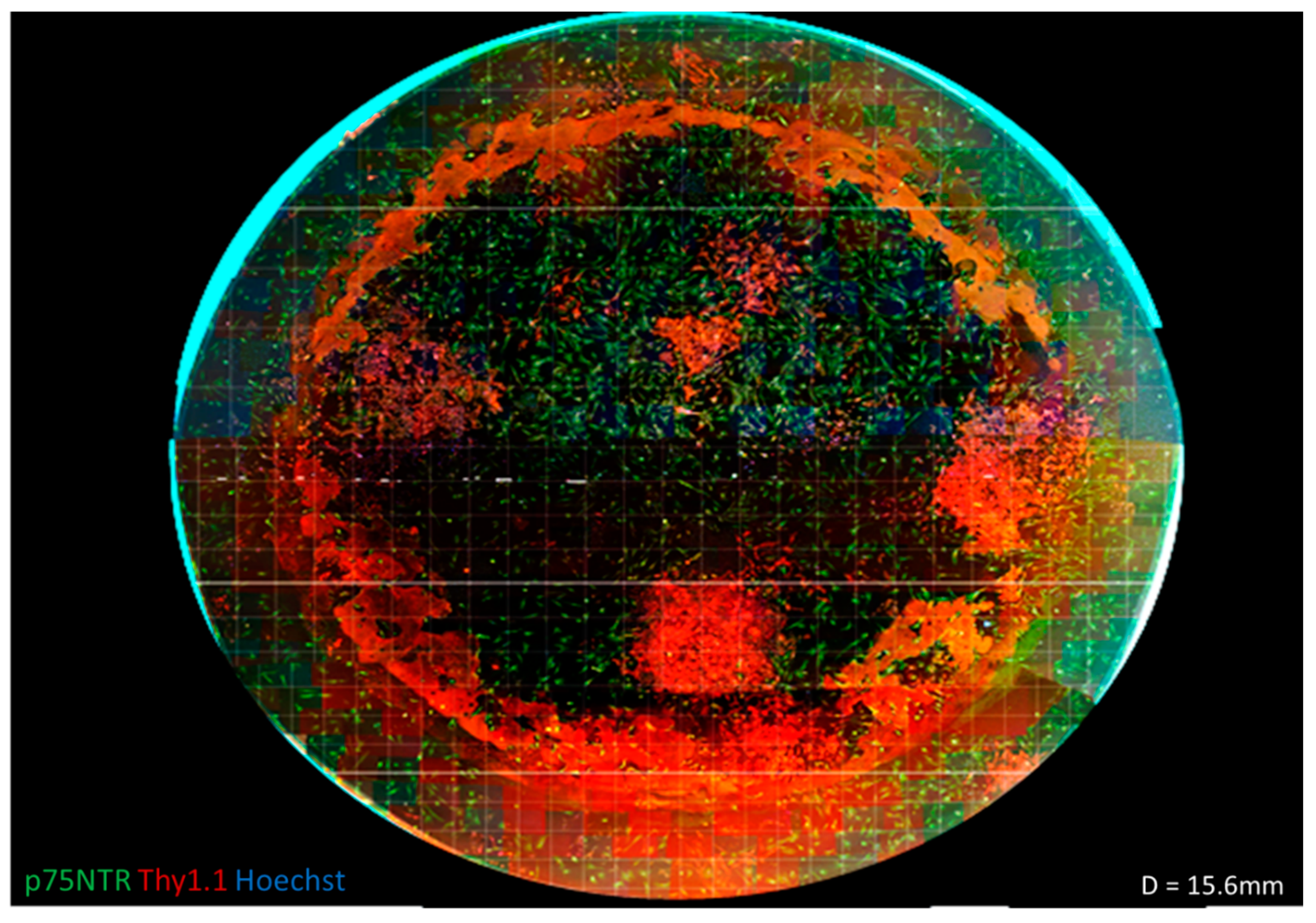
3.1. Human Feeders Encourage an Increase in p75NTR and Spindle-Shaped Cells in Rat OECs
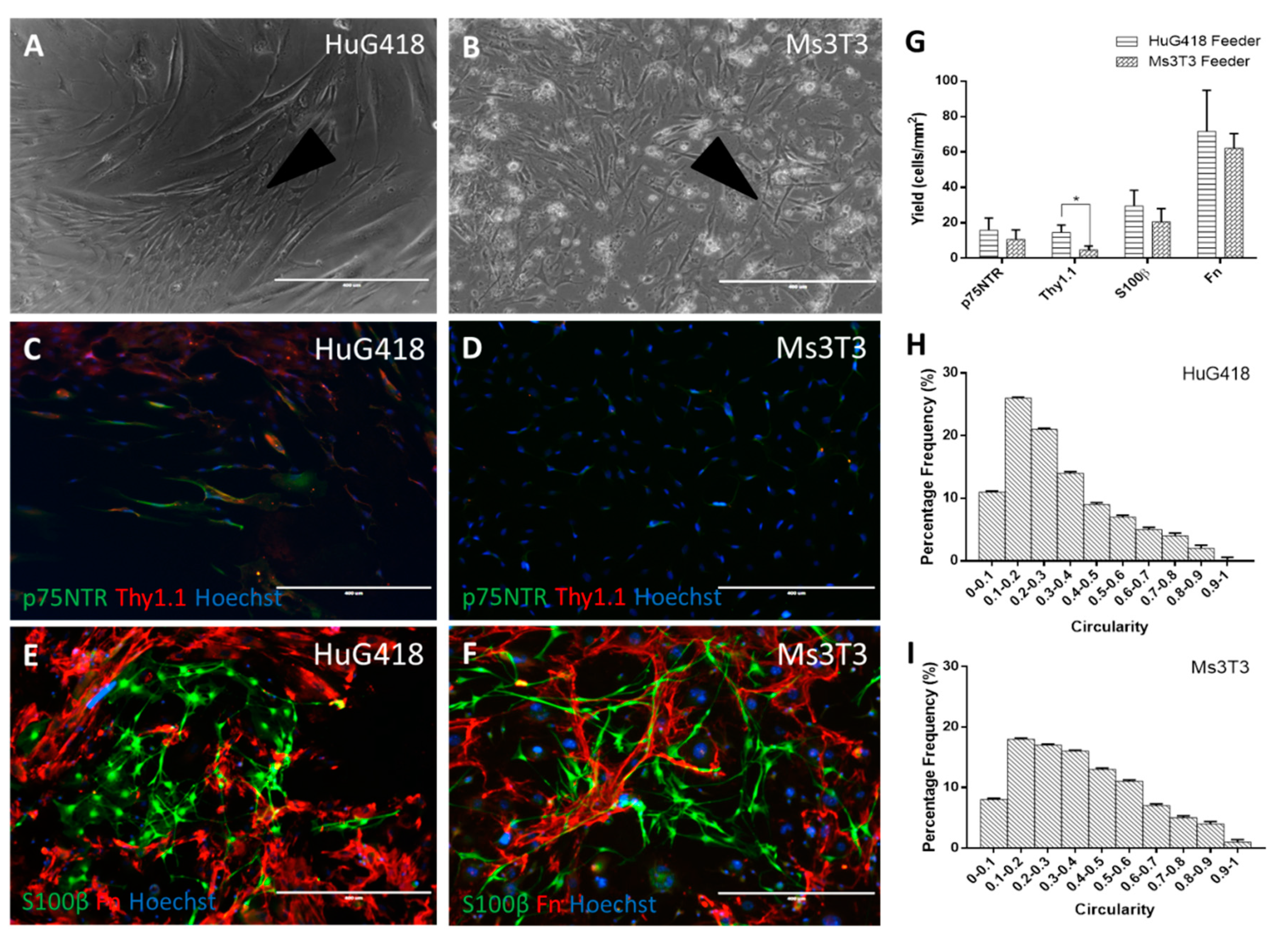
3.2. Comparison of Different Feeder Layers
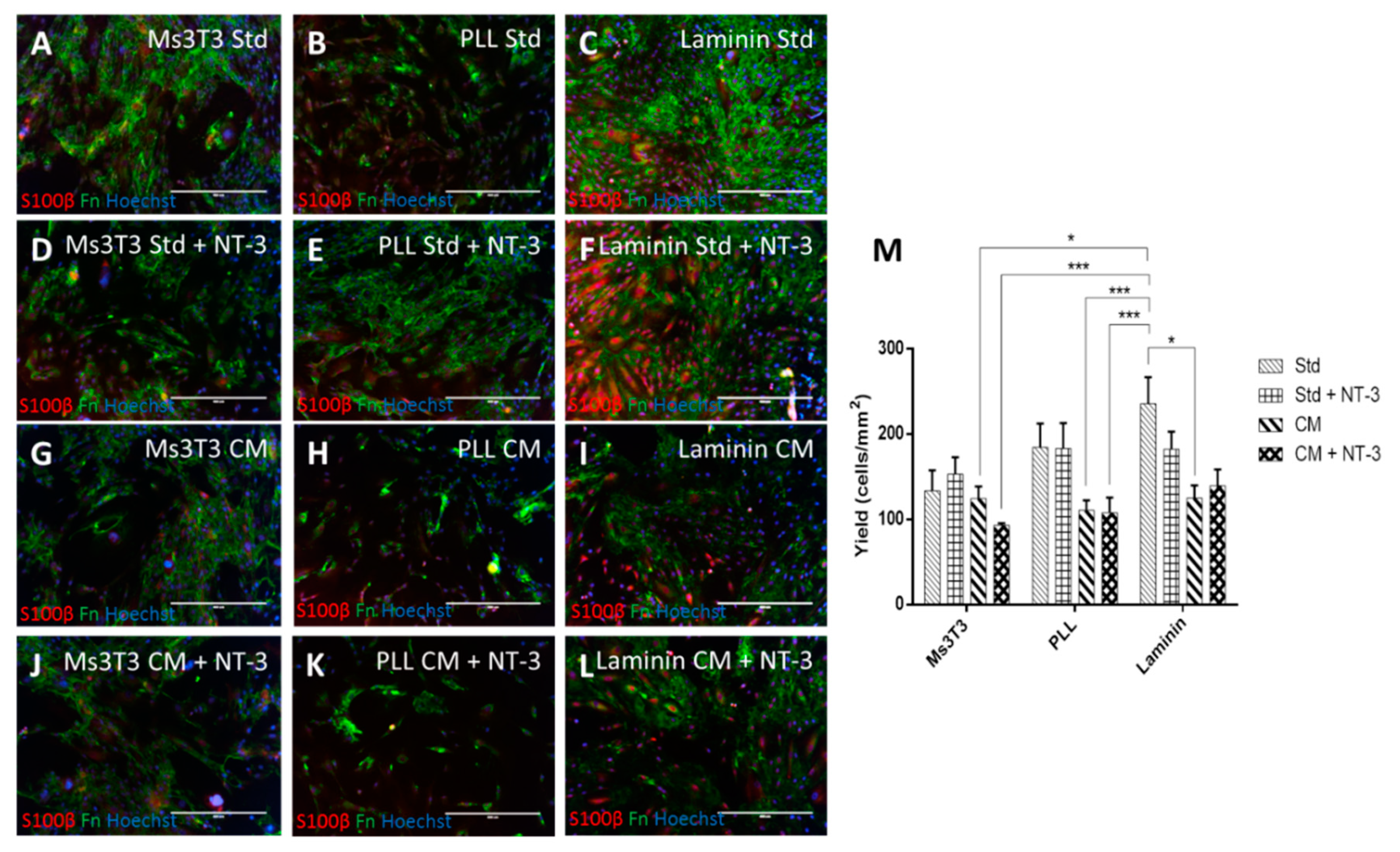
3.3. Human OECs Behave Differently to Rat OECs in Culture
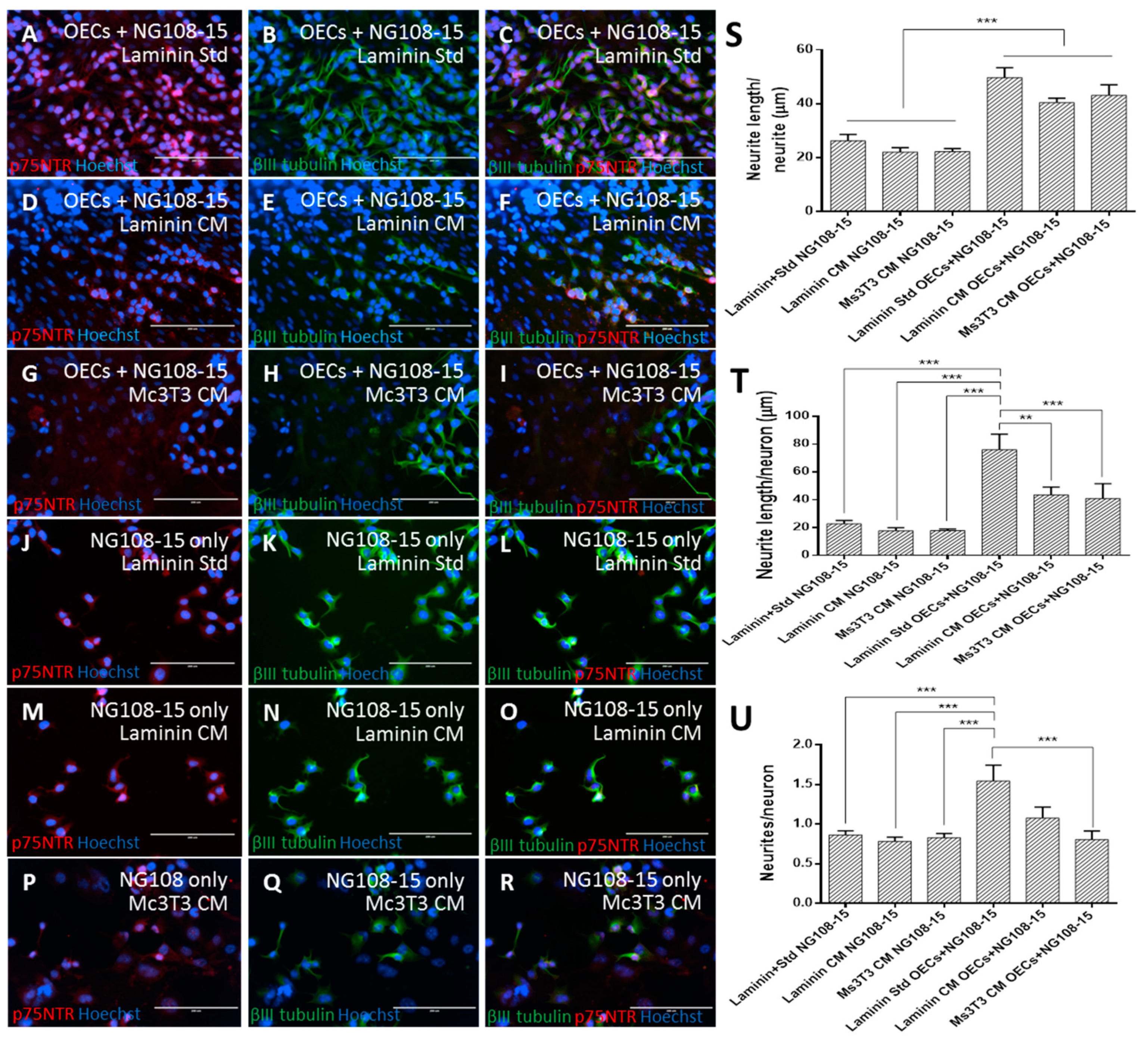
3.4. Human OEC Co-culture with Neurons
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. International Perspectives on Spinal Cord Injury; WHO: Geneva, Switzerland, 2013; p. 16. [Google Scholar]
- National Spinal Cord Injury Center; National Spinal Cord Injury Statistical Center. Fact Sheet; National Spinal Cord Injury Center; National Spinal Cord Injury Statistical Center: Glasgow, UK, 2006. [Google Scholar]
- Willerth, S.M.; Sakiyama-Elbert, S.E. Cell therapy for spinal cord regeneration. Adv. Drug Deliv.Rev. 2008, 60, 263–276. [Google Scholar] [CrossRef] [Green Version]
- Gwak, Y.S.; Kang, J.; Unabia, G.C.; Hulsebosch, C.E. Spatial and temporal activation of spinal glial cells: Role of gliopathy in central neuropathic pain following spinal cord injury in rats. Exp. Neurol. 2012, 234, 362–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fawcett, J.W.; Asher, R.A. The glial scar and central nervous system repair. Brain Res. Bull. 1999, 49, 377–391. [Google Scholar] [CrossRef]
- Ronaghi, M.; Erceg, S.; Moreno-Manzano, V.; Stojkovic, M. Challenges of stem cell therapy for spinal cord injury: Human embryonic stem cells, endogenous neural stem cells, or induced pluripotent stem cells? Stem Cells 2010, 28, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Kigerl, K.A.; de Rivero Vaccari, J.P.; Dietrich, W.D.; Popovich, P.G.; Keane, R.W. Pattern recognition receptors and central nervous system repair. Exp. Neurol. 2014, 258, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; He, B.R.; Hao, D.J. Biological Roles of Olfactory Ensheathing Cells in Facilitating Neural Regeneration: A Systematic Review. Mol. Neurobiol. 2015, 51, 168–179. [Google Scholar] [CrossRef]
- Fraher, J.P. The transitional zone and CNS regeneration. J. Anat. 2000, 196, 137–158. [Google Scholar] [CrossRef]
- Ramón-Cueto, A.; Cordero, M.I.; Santos-Benito, F.F.; Avila, J. Functional Recovery of Paraplegic Rats and Motor Axon Regeneration in Their Spinal Cords by Olfactory Ensheathing Glia. Neuron 2000, 25, 425–435. [Google Scholar] [CrossRef] [Green Version]
- Kawaja, M.D.; Boyd, J.G.; Smithson, L.J.; Jahed, A.; Doucette, R. Technical strategies to isolate olfactory ensheathing cells for intraspinal implantation. J. Neurotrauma 2009, 26, 155–177. [Google Scholar] [CrossRef]
- Thuret, S.; Moon, L.D.F.; Gage, F.H. Therapeutic interventions after spinal cord injury. Nat. Rev. Neurosci. 2006, 7, 628–643. [Google Scholar] [CrossRef]
- Orbay, H.; Little, C.J.; Lankford, L.; Olson, C.A.; Sahar, D.E. The Key Components of Schwann Cell-like Differentiation Medium and their Effects on Gene Expression Pattern of Adipose-Derived Stem Cells. Ann. Plast. Surg. 2015, 74, 584–588. [Google Scholar] [CrossRef] [Green Version]
- Tong, L.; Ji, L.; Wang, Z.; Tong, X.; Zhang, L.; Sun, X. Differentiation of neural stem cells into Schwann-like cells in vitro. Biochem. Biophys. Res. Commun. 2010, 401, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Niapour, A.; Karamali, F.; Karbalaie, K.; Kiani, A.; Mardani, M.; Nasr-Esfahani, M.H.; Baharvand, H. Novel method to obtain highly enriched cultures of adult rat Schwann cells. Biotechnol. Lett. 2010, 32, 781–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zeng, J.; Chen, Y.; Wang, X.; Yao, G.; Wang, W.; Qi, W.; Kong, K. Multiple roles of the p75 neurotrophin receptor in the nervous system. J. Int. Med. Res. 2009, 37, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.R.; Kachramanoglou, C.; Li, D.Q.; Andrews, P.; Choi, D. Anatomy and Cellular Constituents of the Human Olfactory Mucosa: A Review. J. Neurol. Surg. Part B 2014, 75, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Chong, Z.Z. S100B raises the alert in subarachnoid hemorrhage. Rev. Neurosci. 2016, 27, 745–759. [Google Scholar] [CrossRef]
- Wu, X.; Bolger, W.E.; Anders, J.J. Fibroblasts isolated from human middle turbinate mucosa cause neural progenitor cells to differentiate into glial lineage cells. PLoS ONE 2013, 8, e76926. [Google Scholar] [CrossRef]
- Ebel, C.; Brandes, G.; Radtke, C.; Rohn, K.; Wewetzer, K. Clonal In Vitro Analysis of Neurotrophin Receptor p75-Immunofluorescent Cells Reveals Phenotypic Plasticity of Primary Rat Olfactory Ensheathing Cells. Neurochem. Res. 2013, 38, 1078–1087. [Google Scholar] [CrossRef]
- Ramon-Cueto, A.; Perez, J.; Nieto-Sampedro, M. In vitro enfolding of olfactory neurites by p75 NGF receptor positive ensheathing cells from adult rat olfactory bulb. Eur. J. Neurosci. 1993, 5, 1172–1180. [Google Scholar] [CrossRef]
- Rath, N.; Balain, B. Spinal cord injury-The role of surgical treatment for neurological improvement. J. Clin. Orthop. Trauma 2017, 8, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Raisman, G.; Carlstedt, T.; Choi, D.; Li, Y. Clinical prospects for transplantation of OECs in the repair of brachial and lumbosacral plexus injuries: Opening a door. Exp. Neurol. 2011, 229, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.R.N.; Druschel, C.; Curt, A.; Fehlings, M.G. Traumatic spinal cord injury. Nat. Rev. Dis. Primers 2017, 3, 17018. [Google Scholar] [CrossRef] [PubMed]
- Bramlett, H.M.; Dietrich, W.D. Progressive damage after brain and spinal cord injury: Pathomechanisms and treatment strategies. In Progress in Brain Research; John, T.W., Andrew, I.R.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 161, pp. 125–141. [Google Scholar]
- Santiago-Toledo, G.; Georgiou, M.; dos Reis, J.; Roberton, V.H.; Valinhas, A.; Wood, R.C.; Phillips, J.B.; Mason, C.; Li, D.; Li, Y.; et al. Generation of c-MycERTAM-transduced human late-adherent olfactory mucosa cells for potential regenerative applications. Sci. Rep. 2019, 9, 13190. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, M.; Reis, J.N.D.; Wood, R.; Esteban, P.P.; Roberton, V.; Mason, C.; Li, D.; Li, Y.; Choi, D.; Wall, I. Bioprocessing strategies to enhance the challenging isolation of neuro-regenerative cells from olfactory mucosa. Sci. Rep. 2018, 8, 14440. [Google Scholar] [CrossRef]
- Stationery Office. Animals (Scientific Procedures) Act 1986: Guidance on the Operation of the Animals (Scientific Procedures) Act 1986; Stationery Office: London, UK, 2000. [Google Scholar]
- Pollock, K.; Stroemer, P.; Patel, S.; Stevanato, L.; Hope, A.; Miljan, E.; Dong, Z.; Hodges, H.; Price, J.; Sinden, J.D. A conditionally immortal clonal stem cell line from human cortical neuroepithelium for the treatment of ischemic stroke. Exp. Neurol. 2006, 199, 143–155. [Google Scholar] [CrossRef]
- Meijering, E.; Jacob, M.; Sarria, J.C.; Steiner, P.; Hirling, H.; Unser, M. Design and validation of a tool for neurite tracing and analysis in fluorescence microscopy images. Cytom. Part A 2004, 58, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Wojtowicz, A.M.; Oliveira, S.; Carlson, M.W.; Zawadzka, A.; Rousseau, C.F.; Baksh, D. The importance of both fibroblasts and keratinocytes in a bilayered living cellular construct used in wound healing. Wound Rep. Regen. 2014, 22, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Hayat, S.; Thomas, A.; Afshar, F.; Sonigra, R.; Wigley, C.B. Manipulation of olfactory ensheathing cell signaling mechanisms: Effects on their support for neurite regrowth from adult CNS neurons in coculture. Glia 2003, 44, 232–241. [Google Scholar] [CrossRef]
- Nash, H.H.; Borke, R.C.; Anders, J.J. New method of purification for establishing primary cultures of ensheathing cells from the adult olfactory bulb. Glia 2001, 34, 81–87. [Google Scholar] [CrossRef]
- Nash, H.H.; Borke, R.C.; Anders, J.J. Ensheathing cells and methylprednisolone promote axonal regeneration and functional recovery in the lesioned adult rat spinal cord. J. Neurosci. 2002, 22, 7111–7120. [Google Scholar] [CrossRef] [Green Version]
- Sonigra, R.J.; Brighton, P.C.; Jacoby, J.; Hall, S.; Wigley, C.B. Adult rat olfactory nerve ensheathing cells are effective promoters of adult central nervous system neurite outgrowth in coculture. Glia 1999, 25, 256–269. [Google Scholar] [CrossRef]
- Ulrich, R.; Imbschweiler, I.; Kalkuhl, A.; Lehmbecker, A.; Ziege, S.; Kegler, K.; Becker, K.; Deschl, U.; Wewetzer, K.; Baumgartner, W. Transcriptional profiling predicts overwhelming homology of Schwann cells, olfactory ensheathing cells, and Schwann cell-like glia. Glia 2014, 62, 1559–1581. [Google Scholar] [CrossRef] [PubMed]
- Barbacid, M. The Trk family of neurotrophin receptors. J. Neurobiol. 1994, 25, 1386–1403. [Google Scholar] [CrossRef] [PubMed]
- Stephens, P.; Genever, P.G.; Wood, E.J.; Raxworthy, M.J. Integrin receptor involvement in actin cable formation in an in vitro model of events associated with wound contraction. Int. J. Biochem. Cell Biol. 1997, 29, 121–128. [Google Scholar] [CrossRef]
- Li, Y.; Sauve, Y.; Li, D.; Lund, R.D.; Raisman, G. Transplanted olfactory ensheathing cells promote regeneration of cut adult rat optic nerve axons. J. Neurosci. 2003, 23, 7783–7788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakatos, A.; Smith, P.M.; Barnett, S.C.; Franklin, R.J.M. Meningeal cells enhance limited CNS remyelination by transplanted olfactory ensheathing cells. Brain 2003, 126, 598–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumenthal, J.; Cohen-Matsliah, S.I.; Levenberg, S. Olfactory Bulb-Derived Cells Seeded on 3D Scaffolds Exhibit Neurotrophic Factor Expression and Pro-Angiogenic Properties. Tissue Eng. Part A 2013, 19, 2284–2291. [Google Scholar] [CrossRef]
- Kachramanoglou, C.; Law, S.; Andrews, P.; Li, D.; Choi, D. Culture of Olfactory Ensheathing Cells for Central Nerve Repair: The Limitations and Potential of Endoscopic Olfactory Mucosal Biopsy. Neurosurgery 2013, 72, 170–178. [Google Scholar] [CrossRef]
- Keyvan-Fouladi, N.; Raisman, G.; Li, Y. Functional repair of the corticospinal tract by delayed transplantation of olfactory ensheathing cells in adult rats. J. Neurosci. 2003, 23, 9428–9434. [Google Scholar] [CrossRef] [Green Version]
- Tabakow, P.; Raisman, G.; Fortuna, W.; Czyz, M.; Huber, J.; Li, D.; Szewczyk, P.; Okurowski, S.; Miedzybrodzki, R.; Czapiga, B.; et al. Functional Regeneration of Supraspinal Connections in a Patient With Transected Spinal Cord Following Transplantation of Bulbar Olfactory Ensheathing Cells With Peripheral Nerve Bridging. Cell Trans. 2014, 23, 1631–1655. [Google Scholar] [CrossRef] [Green Version]
- Teng, X.; Nagata, I.; Li, H.-P.; Kimura-Kuroda, J.; Sango, K.; Kawamura, K.; Raisman, G.; Kawano, H. Regeneration of nigrostriatal dopaminergic axons after transplantation of olfactory ensheathing cells and fibroblasts prevents fibrotic scar formation at the lesion site. J. Neurosci. Res. 2008, 86, 3140–3150. [Google Scholar] [CrossRef] [PubMed]
- Raisman, G.; Li, Y. Repair of neural pathways by olfactory ensheathing cells. Nat. Rev. Neurosci. 2007, 8, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Jani, H.R.; Raisman, G. Ensheathing cell cultures from the olfactory bulb and mucosa. Glia 2004, 47, 130–137. [Google Scholar] [CrossRef]
- Wewetzer, K.; Radtke, C.; Kocsis, J.; Baumgartner, W. Species-specific control of cellular proliferation and the impact of large animal models for the use of olfactory ensheathing cells and Schwann cells in spinal cord repair. Exp. Neurol. 2011, 229, 80–87. [Google Scholar] [CrossRef]
- Li, Y.; Decherchi, P.; Raisman, G. Transplantation of olfactory ensheathing cells into spinal cord lesions restores breathing and climbing. J. Neurosci. 2003, 23, 727–731. [Google Scholar] [CrossRef]
- Hahn, C.-G.; Han, L.-Y.; Rawson, N.E.; Mirza, N.; Borgmann-Winter, K.; Lenox, R.H.; Arnold, S.E. In vivo and in vitro neurogenesis in human olfactory epithelium. J. Comp. Neurol. 2005, 483, 154–163. [Google Scholar] [CrossRef]
- Krudewig, C.; Deschl, U.; Wewetzer, K. Purification and in vitro characterization of adult canine olfactory ensheathing cells. Cell Tissue Res. 2006, 326, 687–696. [Google Scholar] [CrossRef]
- Omar, M.; Hansmann, F.; Kreutzer, R.; Kreutzer, M.; Brandes, G.; Wewetzer, K. Cell Type- and Isotype-Specific Expression and Regulation of β-Tubulins in Primary Olfactory Ensheathing Cells and Schwann Cells In Vitro. Neurochem. Res. 2013, 38, 981–988. [Google Scholar] [CrossRef]
- Rubio, M.-P.; Munoz-Quiles, C.; Ramon-Cueto, A. Adult olfactory bulbs from primates provide reliable ensheathing glia for cell therapy. Glia 2008, 56, 539–551. [Google Scholar] [CrossRef]
- Bianco, J.I.; Perry, C.; Harkin, D.G.; Mackay-Sim, A.; Feron, F. Neurotrophin 3 promotes purification and proliferation of olfactory ensheathing cells from human nose. Glia 2004, 45, 111–123. [Google Scholar] [CrossRef]
- Lozano, A.M.; Schmidt, M.; Roach, A. A convenient in vitro assay for the inhibition of neurite outgrowth by adult mammalian CNS myelin using immortalized neuronal cells. J. Neurosci. Methods 1995, 63, 23–28. [Google Scholar] [CrossRef]
- Encinas, M.; Iglesias, M.; Liu, Y.; Wang, H.; Muhaisen, A.; Cena, V.; Gallego, C.; Comella, J.X. Sequential treatment of SH-SY5Y cells with retinoic acid and brain-derived neurotrophic factor gives rise to fully differentiated, neurotrophic factor-dependent, human neuron-like cells. J. Neurochem. 2000, 75, 991–1003. [Google Scholar] [CrossRef]
- Lehmann, M.; Fournier, A.; Selles-Navarro, I.; Dergham, P.; Sebok, A.; Leclerc, N.; Tigyi, G.; McKerracher, L. Inactivation of Rho signaling pathway promotes CNS axon regeneration. J. Neurosci. 1999, 19, 7537–7547. [Google Scholar] [CrossRef]
- Fournier, A.E.; Gould, G.C.; Liu, B.P.; Strittmatter, S.M. Truncated soluble Nogo receptor binds Nogo-66 and blocks inhibition of axon growth by myelin. J. Neurosci. 2002, 22, 8876–8883. [Google Scholar] [CrossRef]
- Simpson, P.B.; Bacha, J.I.; Palfreyman, E.L.; Woollacott, A.J.; McKernan, R.M.; Kerby, J. Retinoic acid-evoked differentiation of neuroblastoma cells predominates over growth factor stimulation: An automated image capture and quantitation approach to neuritogenesis. Anal. Biochem. 2001, 298, 163–169. [Google Scholar] [CrossRef]
- Lai, P.-L.; Naidu, M.; Sabaratnam, V.; Kah Hui, W.; David, P.; Kuppusamy, U.R.; Abdullah, N.; Abd Malek, S.N. Neurotrophic Properties of the Lion’s Mane Medicinal Mushroom, Hericium erinaceus (Higher Basidiomycetes) from Malaysia. Int. J. Med. Mushrooms 2013, 15, 539–554. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.-J.; Trachtenberg, J.T.; Thompson, W.J. Schwann cells induce and guide sprouting and reinnervation of neuromuscular junctions. Trends Neurosci. 1996, 19, 280–285. [Google Scholar] [CrossRef]
- Menager, C.; Arimura, N.; Fukata, Y.; Kaibuchi, K. PIP3 is involved in neuronal polarization and axon formation. J. Neurochem. 2004, 89, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-L.; Yu, W.-M.; Strickland, S. Peripheral regeneration. Annu. Rev. Neurosci. 2007, 30, 209–233. [Google Scholar] [CrossRef]
- Kuhn, T.B.; Schmidt, M.F.; Kater, S.B. Laminin and Fibronectin Guideposts signal sustained but opposite effects to passing growth cones. Neuron 1995, 14, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Liesi, P.; Dahl, D.; Vaheri, A. Neurons cultured from developing rat brain attach and spread preferentially to laminin. J. Neurosci. Res. 1984, 11, 241–251. [Google Scholar] [CrossRef] [PubMed]
- di Summa, P.G.; Kalbermatten, D.F.; Raffoul, W.; Terenghi, G.; Kingham, P.J. Extracellular Matrix Molecules Enhance the Neurotrophic Effect of Schwann Cell-Like Differentiated Adipose-Derived Stem Cells and Increase Cell Survival Under Stress Conditions. Tissue Eng. Part A 2013, 19, 368–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagg, T.; Oudega, M. Degenerative and spontaneous regenerative processes after spinal cord injury. J. Neurotrauma 2006, 23, 264–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franssen, E.H.P.; de Bree, F.M.; Verhaagen, J. Olfactory ensheathing glia: Their contribution to primary olfactory nervous system regeneration and their regenerative potential following transplantation into the injured spinal cord. Brain Res. Rev. 2007, 56, 236–258. [Google Scholar] [CrossRef]
- Pizzi, M.A.; Crowe, M.J. Transplantation of fibroblasts that overexpress matrix metalloproteinase-3 into the site of spinal cord injury in rats. J. Neurotrauma 2006, 23, 1750–1765. [Google Scholar] [CrossRef]
- Kisselbach, L.; Merges, M.; Bossie, A.; Boyd, A. CD90 Expression on human primary cells and elimination of contaminating fibroblasts from cell cultures. Cytotechnology 2009, 59, 31–44. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wood, R.; Durali, P.; Wall, I. Impact of Dual Cell Co-culture and Cell-conditioned Media on Yield and Function of a Human Olfactory Cell Line for Regenerative Medicine. Bioengineering 2020, 7, 37. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering7020037
Wood R, Durali P, Wall I. Impact of Dual Cell Co-culture and Cell-conditioned Media on Yield and Function of a Human Olfactory Cell Line for Regenerative Medicine. Bioengineering. 2020; 7(2):37. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering7020037
Chicago/Turabian StyleWood, Rachael, Pelin Durali, and Ivan Wall. 2020. "Impact of Dual Cell Co-culture and Cell-conditioned Media on Yield and Function of a Human Olfactory Cell Line for Regenerative Medicine" Bioengineering 7, no. 2: 37. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering7020037