
Cell Retention as a Viable Strategy for PHA Production from Diluted VFAs with Bacillus megaterium


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Media for Cultivation
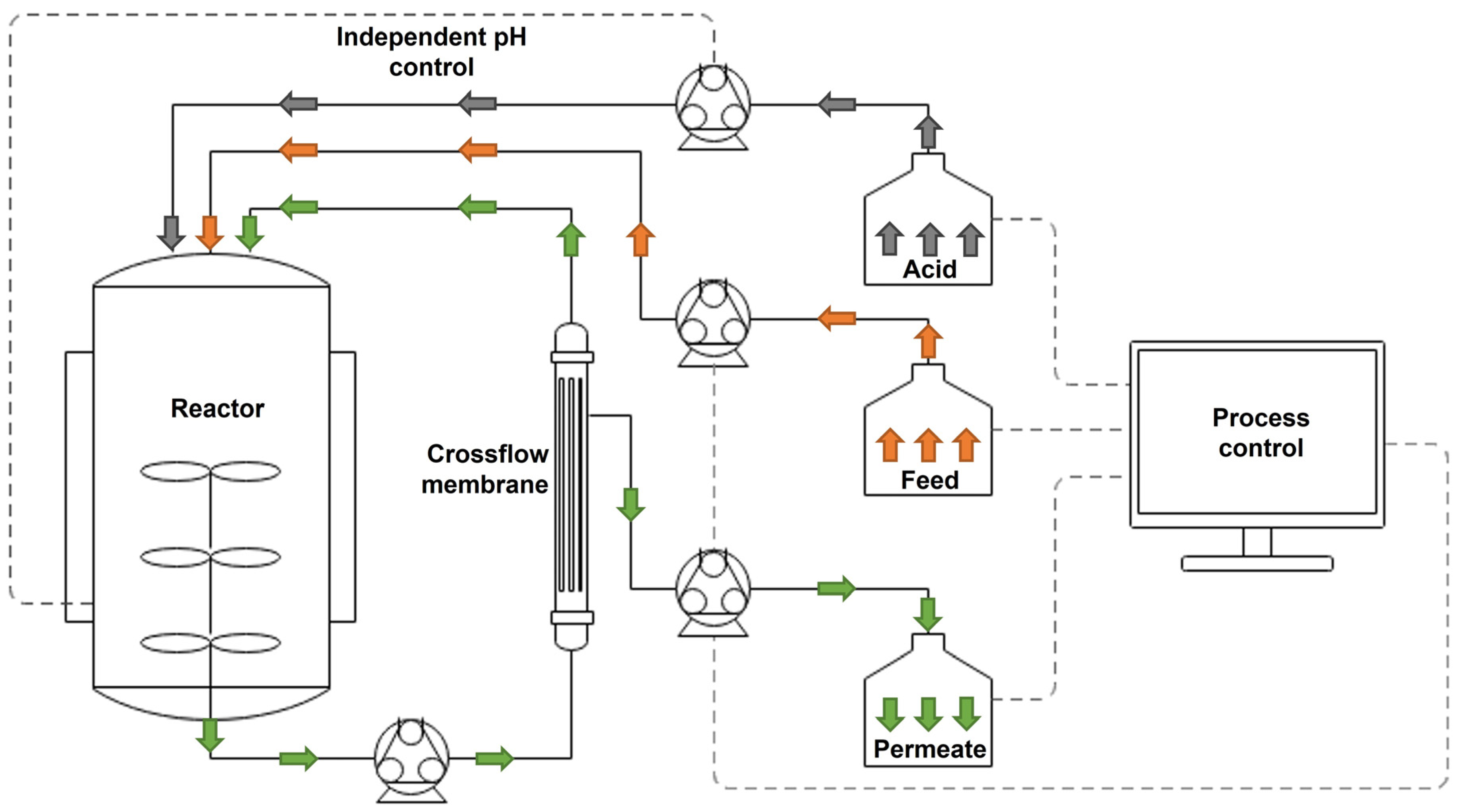
2.2. Fermentation Setup
2.3. Analytical Methods
2.4. Polymer Extraction and Characterization
3. Results and Discussion
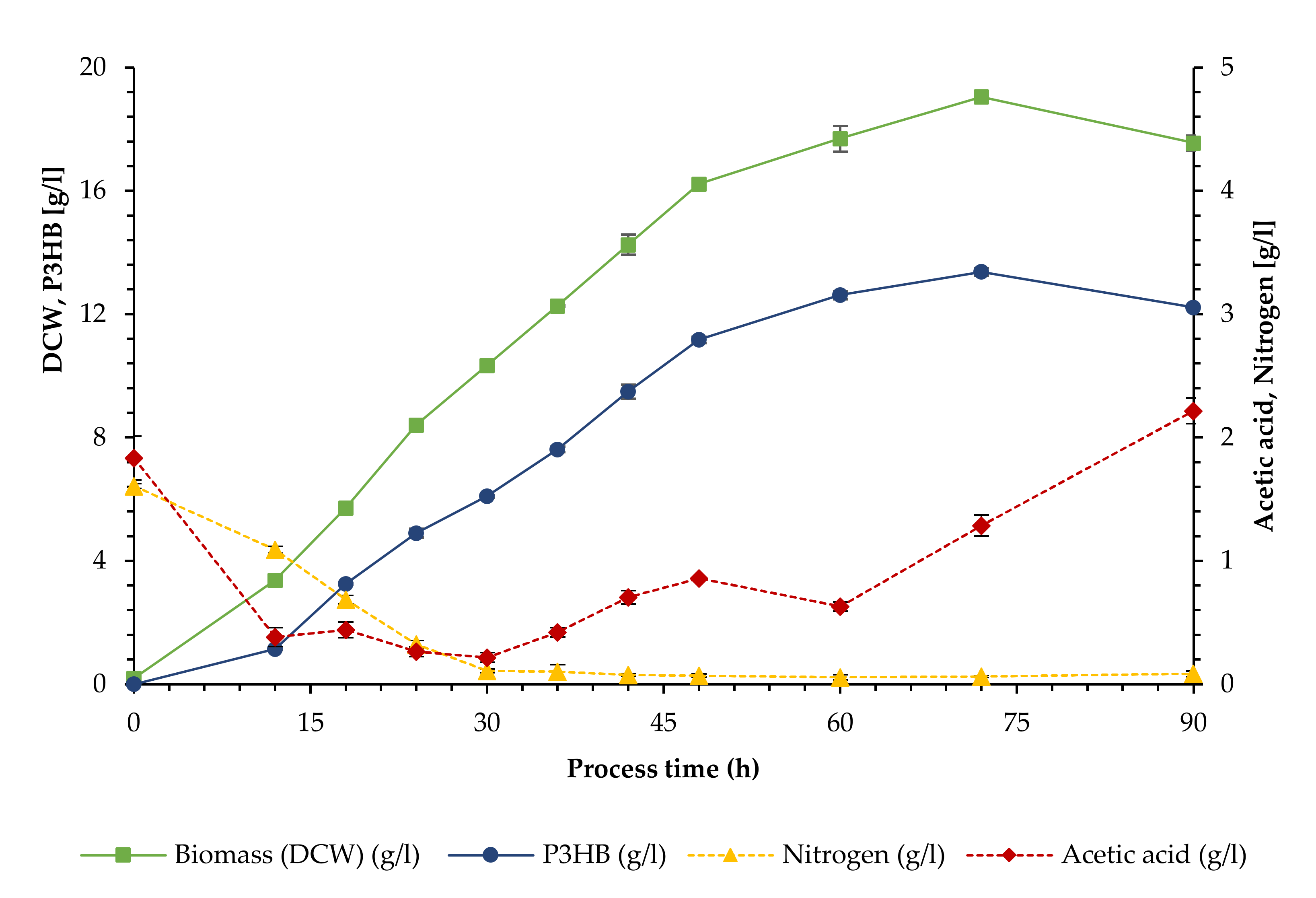
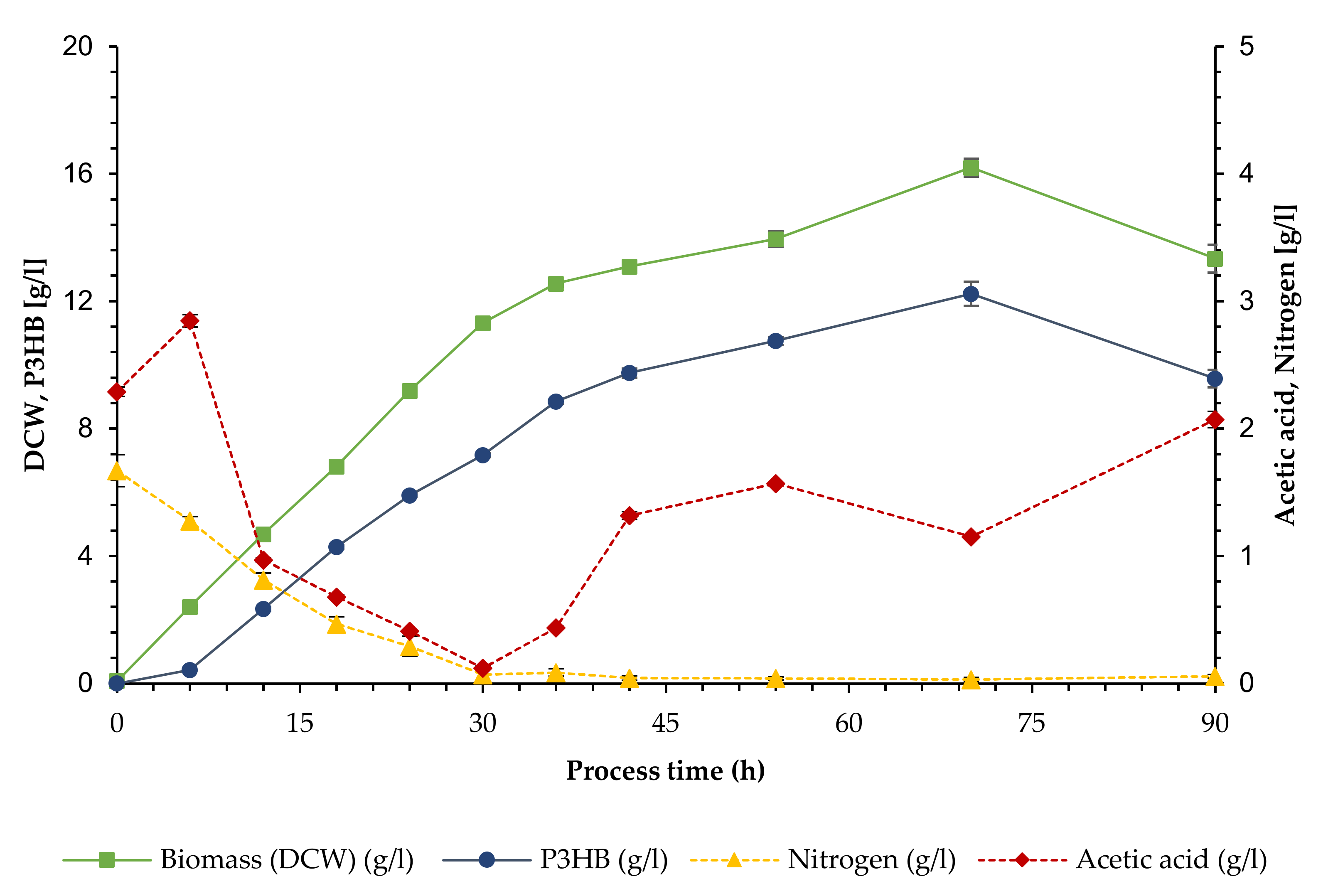
3.1. Growth and Polymer Production
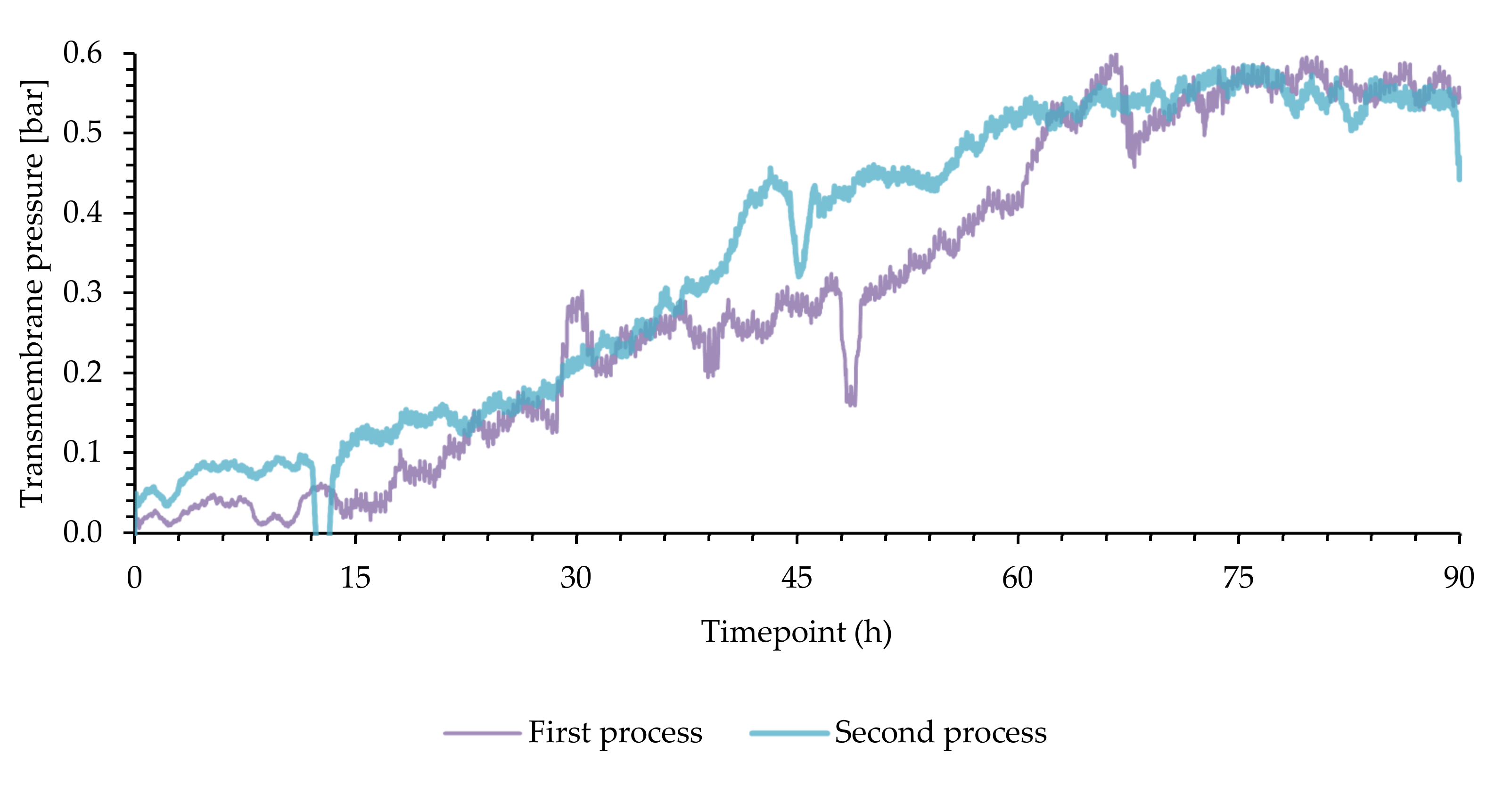
3.2. Membrane Performance
3.3. Polymer Characterization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bugnicourt, E.; Cinelli, P.; Lazzeri, A.; Alvarez, V. Polyhydroxyalkanoate (PHA): Review of Synthesis, Characteristics, Processing and Potential Applications in Packaging. Express Polym. Lett. 2014, 8, 791–808. [Google Scholar] [CrossRef] [Green Version]
- Yadav, B.; Pandey, A.; Kumar, L.R.; Tyagi, R.D. Bioconversion of Waste (Water)/Residues to Bioplastics–A Circular Bioeconomy Approach. Bioresour. Technol. 2020, 298, 122584. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, K.; Dumont, M.J.; Del Rio, L.F.; Orsat, V. Producing PHAs in the Bioeconomy–Towards a Sustainable Bioplastic. Sustain. Prod. Consum. 2017, 9, 58–70. [Google Scholar] [CrossRef]
- Obruca, S.; Sedlacek, P.; Koller, M.; Kucera, D.; Pernicova, I. Involvement of Polyhydroxyalkanoates in Stress Resistance of Microbial Cells: Biotechnological Consequences and Applications. Biotechnol. Adv. 2018, 36, 856–870. [Google Scholar] [CrossRef] [PubMed]
- Philip, S.; Keshavarz, T.; Roy, I. Polyhydroxyalkanoates: Biodegradable Polymers with a Range of Applications. J. Chem. Technol. Biotechnol. 2007, 82, 233–247. [Google Scholar] [CrossRef]
- Taguchi, S.; Doi, Y. Evolution of Polyhydroxyalkanoate (PHA) Production System by “Enzyme Evolution”: Successful Case Studies of Directed Evolution. Macromol. Biosci. 2004, 4, 145–156. [Google Scholar] [CrossRef]
- Licciardello, G.; Catara, A.F.; Catara, V. Production of Polyhydroxyalkanoates and Extracellular Products Using Pseudomonas corrugata and P. mediterranea: A Review. Bioengineering 2019, 6, 105. [Google Scholar] [CrossRef] [Green Version]
- Albuquerque, P.B.S.; Malafaia, C.B. Perspectives on the Production, Structural Characteristics and Potential Applications of Bioplastics Derived from Polyhydroxyalkanoates. Int. J. Biol. Macromol. 2018, 107, 615–625. [Google Scholar] [CrossRef]
- Koller, M.; Maršálek, L.; de Sousa Dias, M.M.; Braunegg, G. Producing Microbial Polyhydroxyalkanoate (PHA) Biopolyesters in a Sustainable Manner. N. Biotechnol. 2017, 37, 24–38. [Google Scholar] [CrossRef]
- Koller, M.; Bona, R.; Hermann, C.; Horvat, P.; Martinz, J.; Neto, J.; Pereira, L.; Varila, P.; Braunegg, G. Biotechnological Production of Poly(3-Hydroxybutyrate) with Wautersia eutropha by Application of Green Grass Juice and Silage Juice as Additional Complex Substrates. Biocatal. Biotransformation 2005, 23, 329–337. [Google Scholar] [CrossRef]
- Braunegg, G.; Bona, R.; Koller, M. Sustainable Polymer Production. Polym. Plast. Technol. Eng. 2004, 43, 1779–1793. [Google Scholar] [CrossRef]
- Koller, M.; Hesse, P.; Bona, R.; Kutschera, C.; Atlić, A.; Braunegg, G. Potential of Various Archae- and Eubacterial Strains as Industrial Polyhydroxyalkanoate Producers from Whey. Macromol. Biosci. 2007, 7, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Haas, C.; Steinwandter, V.; De Apodaca, E.D.; Madurga, B.M.; Smerilli, M.; Dietrich, T.; Neureiter, M. Production of PHB from Chicory Roots–Comparison of Three Cupriavidus necator Strains. Chem. Biochem. Eng. Q. 2015, 29, 99–112. [Google Scholar] [CrossRef]
- Schmid, M.T.; Song, H.; Raschbauer, M.; Emerstorfer, F.; Omann, M.; Stelzer, F.; Neureiter, M. Utilization of Desugarized Sugar Beet Molasses for the Production of Poly(3-Hydroxybutyrate) by Halophilic Bacillus megaterium uyuni S29. Process Biochem. 2019, 86, 9–15. [Google Scholar] [CrossRef]
- Werker, A.; Bengtsson, S.; Korving, L.; Hjort, M.; Anterrieu, S.; Alexandersson, T.; Johansson, P.; Karlsson, A.; Karabegovic, L.; Magnusson, P.; et al. Consistent Production of High Quality PHA Using Activated Sludge Harvested from Full Scale Municipal Wastewater Treatment-PHARIO. Water Sci. Technol. 2018, 78, 2256–2269. [Google Scholar] [CrossRef]
- Tamis, J.; Mulders, M.; Dijkman, H.; Rozendal, R.; van Loosdrecht, M.C.M.; Kleerebezem, R. Pilot-Scale Polyhydroxyalkanoate Production from Paper Mill Wastewater: Process Characteristics and Identification of Bottlenecks for Full-Scale Implementation. J. Environ. Eng. 2018, 144, 04018107. [Google Scholar] [CrossRef]
- Cerrone, F.; Choudhari, S.K.; Davis, R.; Cysneiros, D.; O’Flaherty, V.; Duane, G.; Casey, E.; Guzik, M.W.; Kenny, S.T.; Babu, R.P.; et al. Medium Chain Length Polyhydroxyalkanoate (mcl-PHA) Production from Volatile Fatty Acids Derived from the Anaerobic Digestion of Grass. Appl. Microbiol. Biotechnol. 2014, 98, 611–620. [Google Scholar] [CrossRef]
- Huschner, F.; Grousseau, E.; Brigham, C.J.; Plassmeier, J.; Popovic, M.; Rha, C.; Sinskey, A.J. Development of a Feeding Strategy for High Cell and PHA Density Fed-Batch Fermentation of Ralstonia eutropha H16 from Organic Acids and Their Salts. Process Biochem. 2015, 50, 165–172. [Google Scholar] [CrossRef]
- Follonier, S.; Riesen, R.; Zinn, M. Pilot-Scale Production of Functionalized Mcl-PHA from Grape Pomace Supplemented with Fatty Acids. Chem. Biochem. Eng. Q. 2015, 29, 113–121. [Google Scholar] [CrossRef]
- Chalima, A.; De Castro, L.F.; Burgstaller, L.; Sampaio, P.; Carolas, A.L.; Gildemyn, S.; Velghe, F.; Ferreira, B.S.; Pais, C.; Neureiter, M.; et al. Waste-Derived Volatile Fatty Acids as Carbon Source for Added-Value Fermentation Approaches. FEMS Microbiol. Lett. 2021, 368, fnab054. [Google Scholar] [CrossRef]
- Lee, W.S.; Chua, A.S.M.; Yeoh, H.K.; Ngoh, G.C. A Review of the Production and Applications of Waste-Derived Volatile Fatty Acids. Chem. Eng. J. 2014, 235, 83–99. [Google Scholar] [CrossRef]
- Strazzera, G.; Battista, F.; Garcia, N.H.; Frison, N.; Bolzonella, D. Volatile Fatty Acids Production from Food Wastes for Biorefinery Platforms: A Review. J. Environ. Manage. 2018, 226, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Koutinas, A.A.; Vlysidis, A.; Pleissner, D.; Kopsahelis, N.; Lopez Garcia, I.; Kookos, I.K.; Papanikolaou, S.; Kwan, T.H.; Lin, C.S.K. Valorization of Industrial Waste and By-Product Streams via Fermentation for the Production of Chemicals and Biopolymers. Chem. Soc. Rev. 2014, 43, 2587–2627. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, S.; Hallquist, J.; Werker, A.; Welander, T. Acidogenic Fermentation of Industrial Wastewaters: Effects of Chemostat Retention Time and PH on Volatile Fatty Acids Production. Biochem. Eng. J. 2008, 40, 492–499. [Google Scholar] [CrossRef]
- Choi, S.; Song, C.W.; Shin, J.H.; Lee, S.Y. Biorefineries for the Production of Top Building Block Chemicals and Their Derivatives. Metab. Eng. 2015, 28, 223–239. [Google Scholar] [CrossRef]
- Atasoy, M.; Owusu-Agyeman, I.; Plaza, E.; Cetecioglu, Z. Bio-Based Volatile Fatty Acid Production and Recovery from Waste Streams: Current Status and Future Challenges. Bioresour. Technol. 2018, 268, 773–786. [Google Scholar] [CrossRef] [PubMed]
- Moretto, G.; Valentino, F.; Pavan, P.; Majone, M.; Bolzonella, D. Optimization of Urban Waste Fermentation for Volatile Fatty Acids Production. Waste Manag. 2019, 92, 21–29. [Google Scholar] [CrossRef]
- Ahn, W.S.; Park, S.J.; Lee, S.Y. Production of Poly(3-Hydroxybutyrate) from Whey by Cell Recycle Fed-Batch Culture of Recombinant Escherichia coli. Biotechnol. Lett. 2001, 23, 235–240. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Saha, J.; Haldar, S.; Bhowmic, A.; Mukhopadhyay, U.K.; Mukherjee, J. Production of Poly-3-(Hydroxybutyrate-Co-Hydroxyvalerate) by Haloferax mediterranei Using Rice-Based Ethanol Stillage with Simultaneous Recovery and Re-Use of Medium Salts. Extremophiles 2014, 18, 463–470. [Google Scholar] [CrossRef]
- Obruca, S.; Benesova, P.; Marsalek, L.; Marova, I. Use of Lignocellulosic Materials for PHA Production. Chem. Biochem. Eng. Q. 2015, 29, 135–144. [Google Scholar] [CrossRef]
- Koller, M. A Review on Established and Emerging Fermentation Schemes for Microbial Production of Polyhydroxyalkanoate (PHA) Biopolyesters. Fermentation 2018, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Haas, C.; El-Najjar, T.; Virgolini, N.; Smerilli, M.; Neureiter, M. High Cell-Density Production of Poly(3-Hydroxybutyrate) in a Membrane Bioreactor. N. Biotechnol. 2017, 37, 117–122. [Google Scholar] [CrossRef]
- Ienczak, J.L.; Schmidt, M.; Quines, L.K.; Zanfonato, K.; da Cruz Pradella, J.G.; Schmidell, W.; de Aragao, G.M.F. Poly(3-Hydroxybutyrate) Production in Repeated Fed-Batch with Cell Recycle Using a Medium with Low Carbon Source Concentration. Appl. Biochem. Biotechnol. 2016, 178, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Du, G.; Yu, J. Green Technology for Conversion of Food Scraps to Biodegradable Thermoplastic Polyhydroxyalkanoates. Environ. Sci. Technol. 2002, 36, 5511–5516. [Google Scholar] [CrossRef] [PubMed]
- Zeb, B.S.; Mahmood, Q.; Ping, Z.; Lin, Q.; Lu, H.F.; Tingting, C.; Abbas, G. Assessment of Toxicity of Volatile Fatty Acids to Photobacterium phosphoreum. Microbiol. Russian Fed. 2014, 83, 510–515. [Google Scholar] [CrossRef]
- Albuquerque, M.G.E.; Martino, V.; Pollet, E.; Avérous, L.; Reis, M.A.M. Mixed Culture Polyhydroxyalkanoate (PHA) Production from Volatile Fatty Acid (VFA)-Rich Streams: Effect of Substrate Composition and Feeding Regime on PHA Productivity, Composition and Properties. J. Biotechnol. 2011, 151, 66–76. [Google Scholar] [CrossRef]
- Pradhan, S.; Dikshit, P.K.; Moholkar, V.S. Production, Ultrasonic Extraction, and Characterization of Poly(3-Hydroxybutyrate) (PHB) Using Bacillus megaterium and Cupriavidus necator. Polym. Adv. Technol. 2018, 29, 2392–2400. [Google Scholar] [CrossRef]
- Täubel, M.; Kämpfer, P.; Buczolits, S.; Lubitz, W.; Busse, H.J. Bacillus barbaricus sp. nov., Isolated from an Experimental Wall Painting. Int. J. Syst. Evol. Microbiol. 2003, 53, 725–730. [Google Scholar] [CrossRef]
- Karr, D.B.; Waters, J.K.; Emerich, D.W. Analysis of Poly-β-Hydroxybutyrate in Rhizobium japonicum Bacteroids by Ion-Exclusion High-Pressure Liquid Chromatography and UV Detection. Appl. Environ. Microbiol. 1983, 46, 1339–1344. [Google Scholar] [CrossRef] [Green Version]
- Mannina, G.; Presti, D.; Montiel-Jarillo, G.; Carrera, J.; Suárez-Ojeda, M.E. Recovery of Polyhydroxyalkanoates (PHAs) from Wastewater: A Review. Bioresour. Technol. 2020, 297, 122478. [Google Scholar] [CrossRef]
- Kourmentza, C.; Plácido, J.; Venetsaneas, N.; Burniol-Figols, A.; Varrone, C.; Gavala, H.N.; Reis, M.A.M. Recent Advances and Challenges towards Sustainable Polyhydroxyalkanoate (PHA) Production. Bioengineering 2017, 4, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinn, M.; Witholt, B.; Egli, T. Dual Nutrient Limited Growth: Models, Experimental Observations, and Applications. J. Biotechnol. 2004, 113, 263–279. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, L.; Wen, Q.; Guo, Z. Efficient Polyhydroxyalkanoate (PHA) Accumulation by a New Continuous Feeding Mode in Three-Stage Mixed Microbial Culture (MMC) PHA Production Process. J. Biotechnol. 2015, 209, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Atlić, A.; Koller, M.; Scherzer, D.; Kutschera, C.; Grillo-Fernandes, E.; Horvat, P.; Chiellini, E.; Braunegg, G. Continuous Production of Poly([R]-3-Hydroxybutyrate) by Cupriavidus necator in a Multistage Bioreactor Cascade. Appl. Microbiol. Biotechnol. 2011, 91, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Du, G.; Chen, J.; Yu, J.; Lun, S. Continuous Production of Poly-3-Hydroxybutyrate by Ralstonia eutropha in a Two-Stage Culture System. J. Biotechnol. 2001, 88, 59–65. [Google Scholar] [CrossRef]
- Yue, H.; Ling, C.; Yang, T.; Chen, X.; Chen, Y.; Deng, H.; Wu, Q.; Chen, J.; Chen, G.Q. A Seawater-Based Open and Continuous Process for Polyhydroxyalkanoates Production by Recombinant Halomonas campaniensis LS21 Grown in Mixed Substrates. Biotechnol. Biofuels 2014, 7, 108. [Google Scholar] [CrossRef] [Green Version]
- Schmid, M.T.; Sykacek, E.; O’Connor, K.; Omann, M.; Mundigler, N.; Neureiter, M. Pilot Scale Production and Evaluation of Mechanical and Thermal Properties of P(3HB) from Bacillus megaterium Cultivated on Desugarized Sugar Beet Molasses. J. Appl. Polym. Sci. 2022, 139, 51503. [Google Scholar] [CrossRef]
- Rodríguez-Contreras, A.; Koller, M.; Miranda-de Sousa Dias, M.; Calafell-Monfort, M.; Braunegg, G.; Marqués-Calvo, M.S. High Production of Poly(3-Hydroxybutyrate) from a Wild Bacillus megaterium Bolivian Strain. J. Appl. Microbiol. 2013, 114, 1378–1387. [Google Scholar] [CrossRef]
- Meixner, K.; Kovalcik, A.; Sykacek, E.; Gruber-Brunhumer, M.; Zeilinger, W.; Markl, K.; Haas, C.; Fritz, I.; Mundigler, N.; Stelzer, F.; et al. Cyanobacteria Biorefinery–Production of Poly(3-Hydroxybutyrate) with Synechocystis salina and Utilisation of Residual Biomass. J. Biotechnol. 2018, 265, 46–53. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Origin | Mw [kg/mol] | PDI | Tm (°C) | Tg (°C) | Td (°C) |
|---|---|---|---|---|---|
| This study | 56.83 | 1.29 | 154.30 | −23.27 | 184.50 |
| This study | 59.65 | 1.43 | 151.84 | −24.02 | 194.97 |
| B.megaterium S29 [47] | 350 | 2.76 | 178 | 1.8 | 238.7 |
| B.megaterium S29 [48] | 600–125 | nd | 161 | −11 | nd |
| B.megaterium S29 [48] | 600–125 | nd | 136.8 | −16 | nd |
| Mirel TM F1005 Mirel [49] | 71.7 | 2.1 | 166.7 | nd | 283.3 |
| Mirel TM F1006 Mirel [49] | 71.4 | 2.2 | 165.7 | nd | 275.5 |
| Enmat Y1000 [49] | 77.6 | 2.6 | 168.9 | nd | 272.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kacanski, M.; Pucher, L.; Peral, C.; Dietrich, T.; Neureiter, M. Cell Retention as a Viable Strategy for PHA Production from Diluted VFAs with Bacillus megaterium. Bioengineering 2022, 9, 122. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering9030122
Kacanski M, Pucher L, Peral C, Dietrich T, Neureiter M. Cell Retention as a Viable Strategy for PHA Production from Diluted VFAs with Bacillus megaterium. Bioengineering. 2022; 9(3):122. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering9030122
Chicago/Turabian StyleKacanski, Milos, Lukas Pucher, Carlota Peral, Thomas Dietrich, and Markus Neureiter. 2022. "Cell Retention as a Viable Strategy for PHA Production from Diluted VFAs with Bacillus megaterium" Bioengineering 9, no. 3: 122. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering9030122