
Microbial and Chemical Dynamics during Marula Wine Fermentation

Department of Biochemistry, Microbiology and Biotechnology, University of Limpopo, Private Bag X1106, Sovenga 0727, South Africa
*
Author to whom correspondence should be addressed.
Beverages 2022, 8(3), 50; https://0-doi-org.brum.beds.ac.uk/10.3390/beverages8030050
Submission received: 30 March 2022
/
Revised: 5 May 2022
/
Accepted: 7 May 2022
/
Published: 22 August 2022
(This article belongs to the Special Issue New Insights on Traditional Fermented Beverages)
Abstract
:Marula wine is traditionally produced through a spontaneous fermentation process and has a huge economic potential in Africa. The current study investigated the contributing microbiota and the metabolites produced during the wine fermentation process. Microbial communities were analyzed by selective cultivation and identified by biotyping and rDNA sequencing. Sugars and volatile compounds were determined with the high performance liquid chromatography and gas chromatography, respectively. Different Lactobacillus spp. were present throughout the fermentation process but dominated the earlier stages of fermentation, together with non-Saccharomyces yeasts, whereas Saccharomyces cerevisiae and acetic acid bacteria dominated the latter stages. Sucrose, glucose and fructose were detected during the early stages, while ethanol and butanol were present during the latter stages of fermentation. Interestingly, acetic acid and formic acid were detected in relatively high amounts at the latter stages of fermentation. Lactobacillus spp. and S. cerevisiae were identified as the primary contributing microbiota, and Acetobacter aceti and Acetobacter pasteuriannus were associated with the off taste and spoilage of the marula wine.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Sclerocarya birrea subsp. caffra (Marula) is a deciduous tree, which belongs to the family known as Anacardiaceae. The tree is distributed throughout the frost-free areas of Africa [1] and survives well in sandy loam and clay soils in areas receiving annual rainfall of 200–1370 mm [2]. The marula tree is amongst the most utilized fruit-bearing trees on the African continent [3]. The female marula tree bears pale yellow plum-sized fruits, which are rich in minerals and carbohydrates and are either eaten fresh or processed into beverages [4]. The marula fruit has a unique flavor, and it is rich in nutrients and vitamins. The nutrient and chemical compositions of marula fruits differ widely depending upon genetic and environmental factors [4]. The marula fruit contains high levels of minerals such as potassium, magnesium and calcium [5]. The high mineral and vitamin C contents have dramatically increased the interest in marula fruit utilization for industrial purposes [6]. Additionally, the fruit has high levels of antioxidant activity compared to most other edible fruits, and a positive correlation has been reported between the antioxidant activity and polyphenol and the vitamin C content in marula fruit juice [7].
In many local communities where the marula tree grows, the fruit is frequently processed into non-alcoholic and alcoholic beverages for social and commercial purposes. The alcoholic beverage is so far the most dominant product from marula fruits in rural African communities where the marula trees are abundant. Marula wine forms an integral part of the livelihoods as well as the social and cultural activities of many communities in the Limpopo province of South Africa. Simatende et al. [8] reported that 74% of households in the Limpopo province produce between 138 L and 311 L of marula fruit wine each season.
The marula fruit can vary between 10.4 and 16.0° Brix in sugar content, while the pulp (i.e., total soluble solids of puree and juices) varies from 7.5 to 15.5° Brix depending on the climatic conditions through the seasons [9]. Fermenting marula juice can yield a beverage with an alcohol content as high as 5% (w/v) depending on the period of fermentation [10] and the sugar content of the juice. In the production of marula wine, the fermentation process is mainly mediated by the natural microbiota associated with the marula fruit. The microbiota is mostly introduced by Drosophila fruit flies during ripening [2] and also picked from the soil with the marula fruits that drop to the ground when ripe. Yeast species such as Aureobasidium pullulans, Geotrichum capitatum, Trichosporon brassicae, Rhodotorula mucilaginosa, Wickerhamomyces anomalus (previously known as Hansenula anomala or Pichia anomala) and Hansenula jadinii were previously isolated from the marula fruit surface [11], while W. anomalus, Pichia guilliermondii, Candida intermedia, Candida tropicalis and Saccharomyces spp. were isolated from marula juice and wine [12,13]. Bacterial species, mainly Lactobacillus, were isolated from marula wine in Gwanda, Zimbabwe [12].
The yeasts were reported as the main drivers of alcoholic fermentation with the conversion of sugars to ethanol and other secondary metabolites [12]. Lactic acid bacteria present in marula juice metabolize the available sugars to mainly lactic acid. This activity by lactic acid bacteria has been shown to reduce the pH of the marula wine by at least one unit [2]. Furthermore, some of the lactic acid bacteria derived from marula have been shown to have probiotic properties and produced bacteriocins that were active against other lactic acid bacteria such as Escherichia coli, Pseudomonas aeruginosa, Klebsiella pneumoniae and Listeria spp. [14].
Previous studies reported on the microbial diversity associated with marula wine fermentation and the commercial potential of some of these yeasts and bacteria. The microorganisms were identified using phenotypic and biochemical tests [12] and were mainly performed on marula wines from Zimbabwe, Namibia and regions outside South Africa. Consequently, little is known about the microbial communities in marula wine that is fermented in South Africa. The quality of the final marula wine depends on the natural microbiota that is active during fermentation. Marula wine typically has a short shelf life that ranges from 2 to 4 days post fermentation depending on the storage temperature. Undoubtedly, a better understanding of the microbial diversity associated with this product would help to formulate better management strategies to control the fermentation process and prevent the proliferation of potential spoilage organisms.
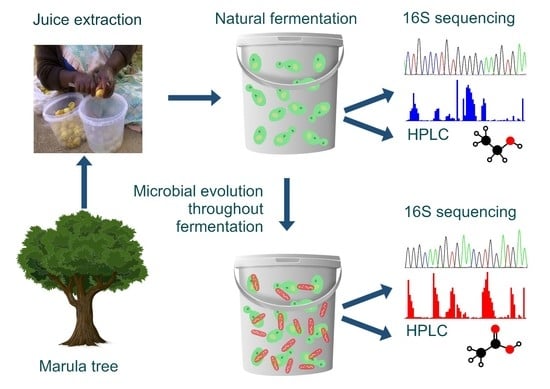
The purpose of this study was to characterize the microorganisms that contribute to the fermentation of marula juice. The different chemical metabolites, which were produced during marula wine fermentation and that typically contribute to the taste and aroma character were also investigated.
2. Materials and Methods
2.1. Marula Fruit Juice and Wine Preparations
All the fruit and wine used in this study were collected during the 2015 marula season (January to March). Approximately 20 kg of ripe marula fruits was collected from the grounds of the University of Limpopo (S 23°52′57.43″; E 29°43′1.77″) in South Africa, into plastic buckets. The juice was manually extracted and mixed with an equal part of tap water in accordance with the traditional recipe of preparing marula wine, and this is primarily for increasing the volume because the juice yield of the fruits is low. The mixture was divided into two equal portions. One portion was kept and allowed to ferment in the laboratory (the Lab brew), and the other was transported to the community partner’s home in Mentz (S 23°54′3.54″ E 29°46′28.02″). The Lab wine was fermented at a constant temperature of 25 °C, while the temperature of the Mentz fermentation was not controlled. The Mentz and Lab wines constituted biological replicates. All the marula wines were produced by spontaneous fermentation.
The Moshira-A and -B marula wines were prepared by the same brewer from Moshira village in the Sekhukhune district (S 24°20′35.84″ E 30°5′33.07″) of South Africa. Moshira-A was collected on Day 0 of marula wine preparation, whereas the Moshira-B wine was collected two days later. These wines represented two batches prepared from the same marula fruit set. Both wines were fermented for 11 days.
Marula fruits were also collected from Moshira village for content comparison with fruits collected on the grounds of the University of Limpopo.
All wines were sampled at 2-days intervals. At each sampling point, 1.5 mL of marula wine was drawn, transferred to a sterile micro-centrifuge tube and centrifuged at 10,000× g for 5 min at room temperature. The pellet was suspended in 50% glycerol in water and stored at −80 °C, and the liquid fraction was stored at −20 °C until needed.
2.2. Enumeration and Isolation of Bacteria and Yeasts
The yeast population was analyzed by direct plating on Wallerstein Laboratory Nutrient (WLN) agar (Sigma Aldrich, Steinheim, Germany) supplemented with 34 mg/L chloramphenicol (Sigma Aldrich) to suppress bacterial growth and 250 mg/L biphenyl (Riedel-de Haen AG, Seelze, Germany) to inhibit the growth of molds. Bacteria were isolated on de Man, Rogosa and Sharpe (MRS) medium (Biolab, Merck, East Rand, South Africa) supplemented with 100 mg/L Delvocid (DSM, Heerlen, The Netherlands) to inhibit fungal growth. One hundred microliters of ten-fold serial dilutions of the samples prepared in saline (0.9% w/v NaCl) was plated, and the plates were incubated at 30 °C for 72 h until clear colonies were obtained. Colonial morphology was used to determine relative abundance of microbial species at different sampling intervals. Ten percent of the viable counts for a particular colonial morphology was randomly sub-cultured for identification.
2.3. Identification by Biotyping of the Bacterial Isolates
The biotyper function of an Ultraflex MALDI-TOF (Bruker Daltonik GmbH, Bremen, Germany) was used for identification of purified bacterial isolates. Bacterial colonies were thoroughly suspended in 300 μL of distilled water, and 900 µL of absolute ethanol was added. The cell suspension was mixed and then centrifuged at 10,000× g for 2 min. The supernatant was removed, and 50 µL of formic acid was added to the cell pellet before vortexing. Acetonitrile (50 µL) was added, and the mixture was again vortexed vigorously, followed by centrifugation at 10,000× g for 2 min. One microliter of the supernatant was spotted onto the MALDI-TOF target plate and allowed to dry. The samples were overlaid with 1 µL of matrix (α-cyano-4-hydroxycinnamic acid (HCCA) in 50% acetonitrile and 1.5% trifluoroacetic acid) and dried at room temperature. The samples were applied to an Ultraflex LT mass spectrometer (Bruker Daltonik), and the results were analyzed by MALDI Biotyper 3.0 software (Bruker Daltonik), which was calibrated with bacterial test standard (BTS, Bruker Daltonik) in the automatic mode and using the manufacturer’s settings. Species identification was assigned and considered reliable when log scores ≥ 2, log scores < 2.0 but ≤1.7 were considered reliable identification at genus level, while log scores < 1.7 were not reliable for identification.
2.4. Amplification and Sequencing of rDNA Genes
Yeast genomic DNA was extracted following the method of Sambrook and Rusell [15], and the method described by Neumann et al. [16] was used to extract bacterial genomic DNA. The yeast ITS1-5.8S rRNA-ITS2 region was amplified using the primer set ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC) [17], and the bacterial 16S rDNA was amplified using the primers EUbB (27F) (5′-AGAGTTTGATCMTGGCTCAG-3′) and EUbA (1522R) (5′-AAGGAGGTGATCCANCCANCCRCA-3′) [18]. Polymerase chain reaction (PCR) was performed in a 25 μL reaction mixture containing 2.5 μL of 2.5 μmol/L of each primer, 2.5 μL of 10 × Ex Taq buffer, 1 unit of Ex Taq™ polymerase (TaKaRa Bio Inc., Olsu, Shiga, Japan), 1 µL of a 25 mmol/L MgCl2, 4 μL of 2.5 mmol/L dNTP, 1 μL of template DNA (100 ng/μL) and 11 μL distilled water.
For yeast 18S rDNA amplification, the PCR conditions consisted of an initial denaturation at 94 °C for 3 min, followed by 40 cycles at 94 °C for 30 s, 54 °C for 30 s and 72 °C for 45 s and a final extension at 72 °C for 10 min. The amplification of the bacterial 16S rDNA was carried out using the same cycling conditions except for 35 cycles and at an annealing temperature of 55 °C. The PCR amplicons were subjected to Sanger sequencing using the ITS4 primer for yeasts and the EUbA for bacteria 16S rRNA gene sequencing. The PCR amplicons of both bacteria and yeasts were purified using the ZymocleanTM Gel DNA recovery kit (Zymo Research Corporation, Irvine, CA, USA), following the manufacturer’s protocol, and the purified PCR products were sent to the Central Analytical Facility (CAF), Stellenbosch University, for sequencing. The resulting sequences were analyzed using Bio-Edit (Informer Technologies, Inc., Los Angeles, CA, USA), and the taxonomic assignment of individual isolates was performed by comparing with the relevant sequences available in the GenBank databases (NCBI) (http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed, accessed on 15 September 2018) using the Basic Local Alignment Search Tool (BLAST) algorithm [17]. Default settings were used. Identity was assigned at ≥97% sequence similarity.
2.5. Chemical Profiling of Marula Wine Samples
The wine samples collected at different fermentation stages were centrifuged at 10,000× g for 5 min, and the resulting supernatant was filtered through a 0.22 µm Whatman filter membrane into 1.5 mL HPLC vials.
Sugars (sucrose, glucose and fructose) in marula juice and wine samples were determined by HPLC using a Shimadzu Prominence 20 HPLC system. Samples of 20 µL were injected into a Rezex RHM-monosaccharide H+ column (300 × 7.8 mm) and eluted using water at a flow rate of 0.6 mL/min. The column temperature was kept at 85 °C. The separated components were detected using a Shimadzu RID10A refractive index detector. Data were processed using LC Solutions software. Sugars were identified and quantified by comparing with known standards of sucrose, glucose and fructose at a concentration range of 0.39–100 mg/mL.
The volatile organic compounds in marula juice and wine samples were analyzed by capillary gas chromatography using a Shimadzu GC2010plus gas chromatograph on a NukolTM capillary column (15 m × 0.53 mm × 0.5 µm). Nitrogen was used as the carrier gas. A sample of 1 µL was injected into the column at an injection temperature of 220 °C. The column temperature was kept at 200 °C with a flow rate of 1.33 mL/min. The volatile organic compounds were detected using a flame ionization detector (FID), with the detector temperature maintained at 220 °C. The peaks were processed using GC Solutions software and the pure compounds of organic acids (acetic acid, formic acid, propionic acid, isobutyric acid, butyric acid isovaleric acid, valeric acid isocaproic acid, caproic acid, heptanoic acid and nonanoic acid), alcohols (ethanol, n-butanol, 1-pentanol 1-propanol and 2 methyl-1-propanol), aldehydes (formaldehyde, acetaldehyde butyraldehyde and isobutyraldehyde) and the ester (isopentyl acetate). The standards were prepared and used at a concentration range of 0.125–1000 mg/L in order to assist with the identification of compounds present in the marula juice and wine. The National Institute of Standards and Technology (NIST) mass spectral library was used to compare the retention times.
2.6. Statistical Analysis
The statistical analysis of the sugar results was performed using GraphPad Instat version 3 software for analysis of variance using two-way ANOVA with Tukey test for comparison of any significant differences between the means of the concentrations of sugars of the marula wines from Mentz and Moshira. The significance of the differences was considered at p < 0.05.
3. Results
Marula wine samples were collected on alternating days to achieve a comprehensive spectrum of microorganisms and chemicals that contribute to the organoleptic properties of the wines.
3.1. Marula Fruit and Juice Analysis
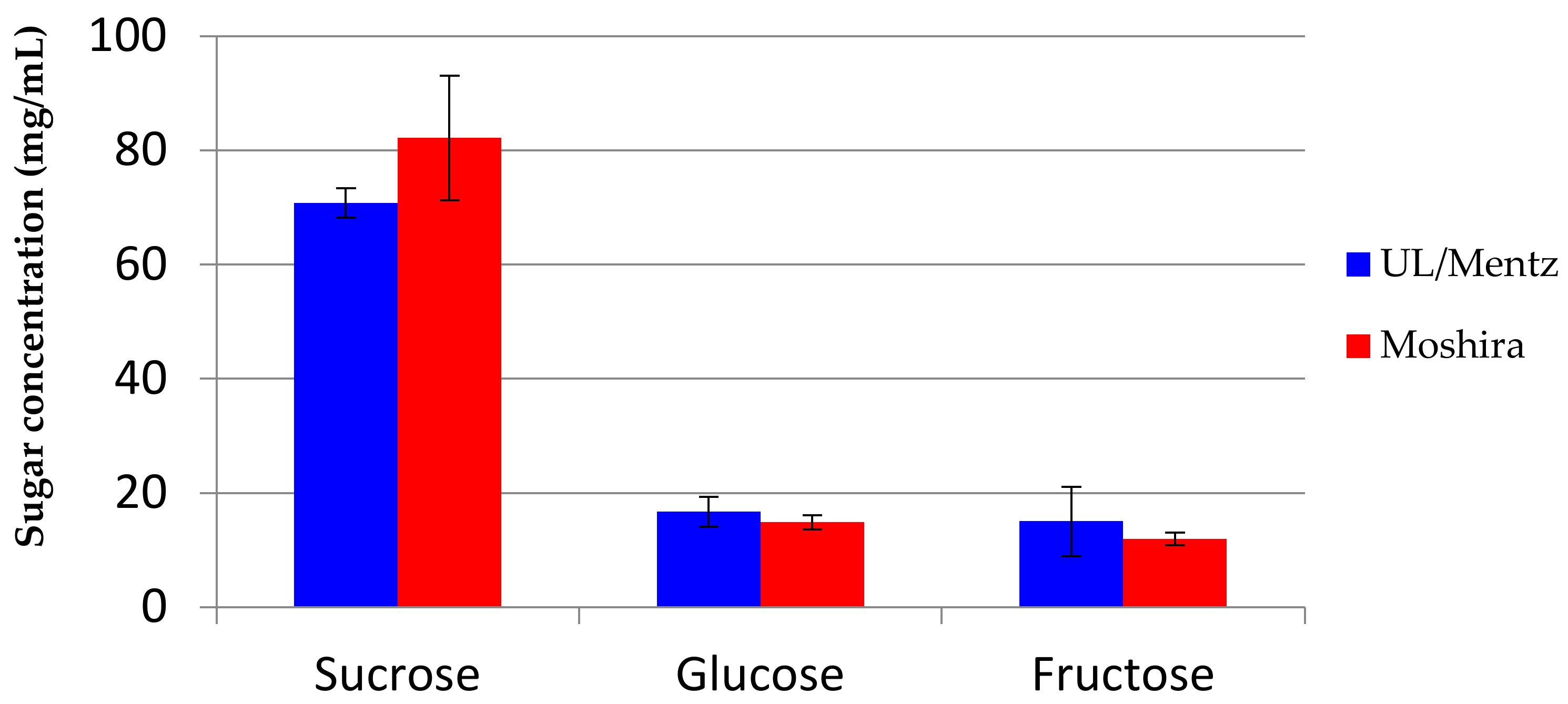
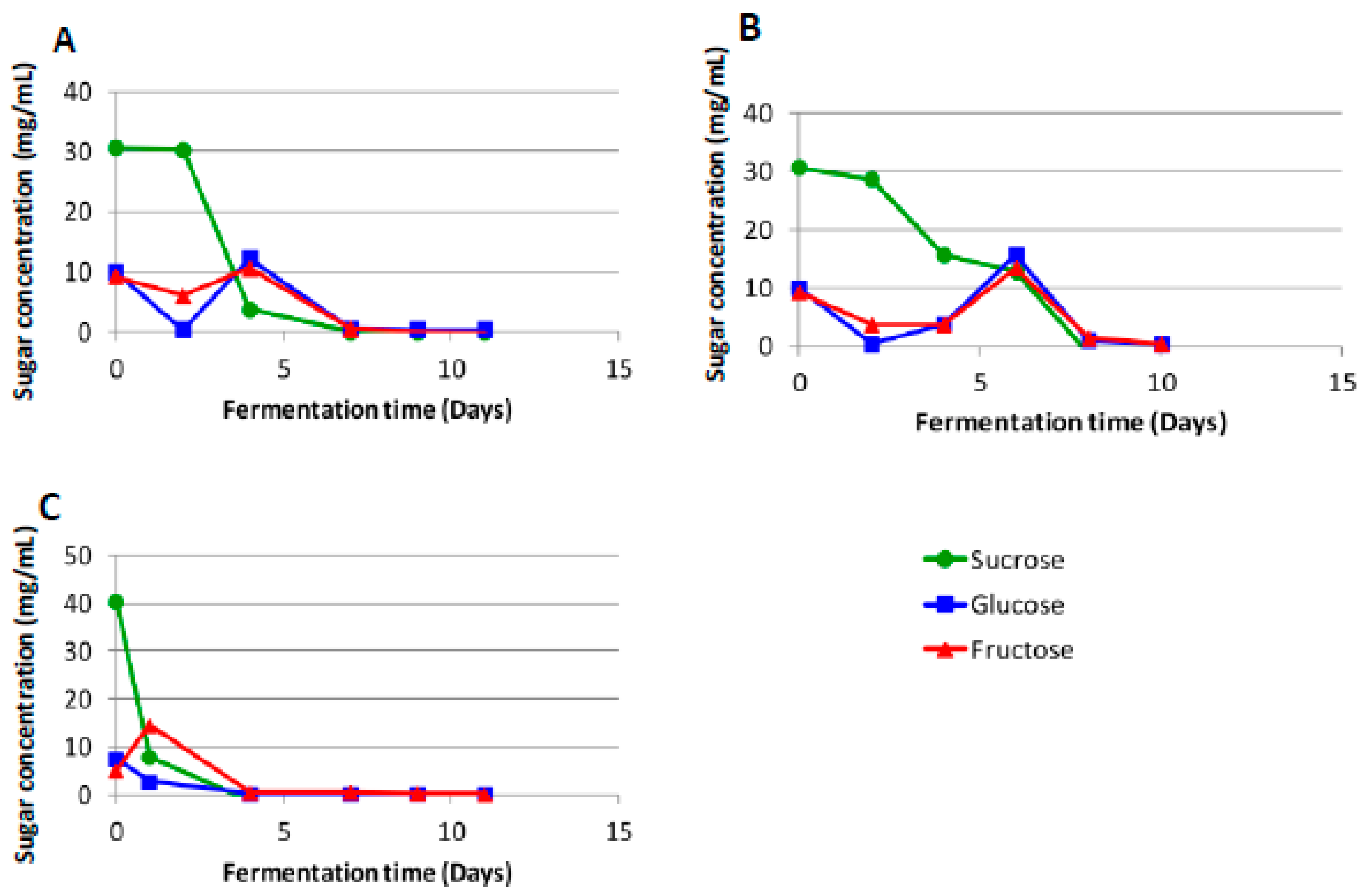
The ripe marula fruits contain sucrose, glucose and fructose as the major fermentable carbohydrates, with sucrose being the dominant sugar (Figure 1). Following dilution of the marula juice with water, the sugar content in the fresh pulp/water mixture used for producing the Lab and Mentz marula wines contained 31.4, 10.5 and 9.7 mg/mL of sucrose, fructose and glucose, respectively (Figure 2A,B).
3.2. Sugar Utilization during Fermentation
The Lab and Mentz fermentations, which were biological replicates, showed similar sugar consumption profiles, although the fermentations were performed in different environments. The Mentz wine fermentation temperature was not controlled and showed a slightly slower decline in the levels of sucrose (Figure 2A,B). The Moshira wines prepared from a different batch of juice displayed significantly faster sucrose utilization than the Lab and Mentz wines (Figure 2C). Interestingly, fructose consumption never decreased below 2–3 mg/mL for all the wines.
3.3. Microbial Growth during Fermentation
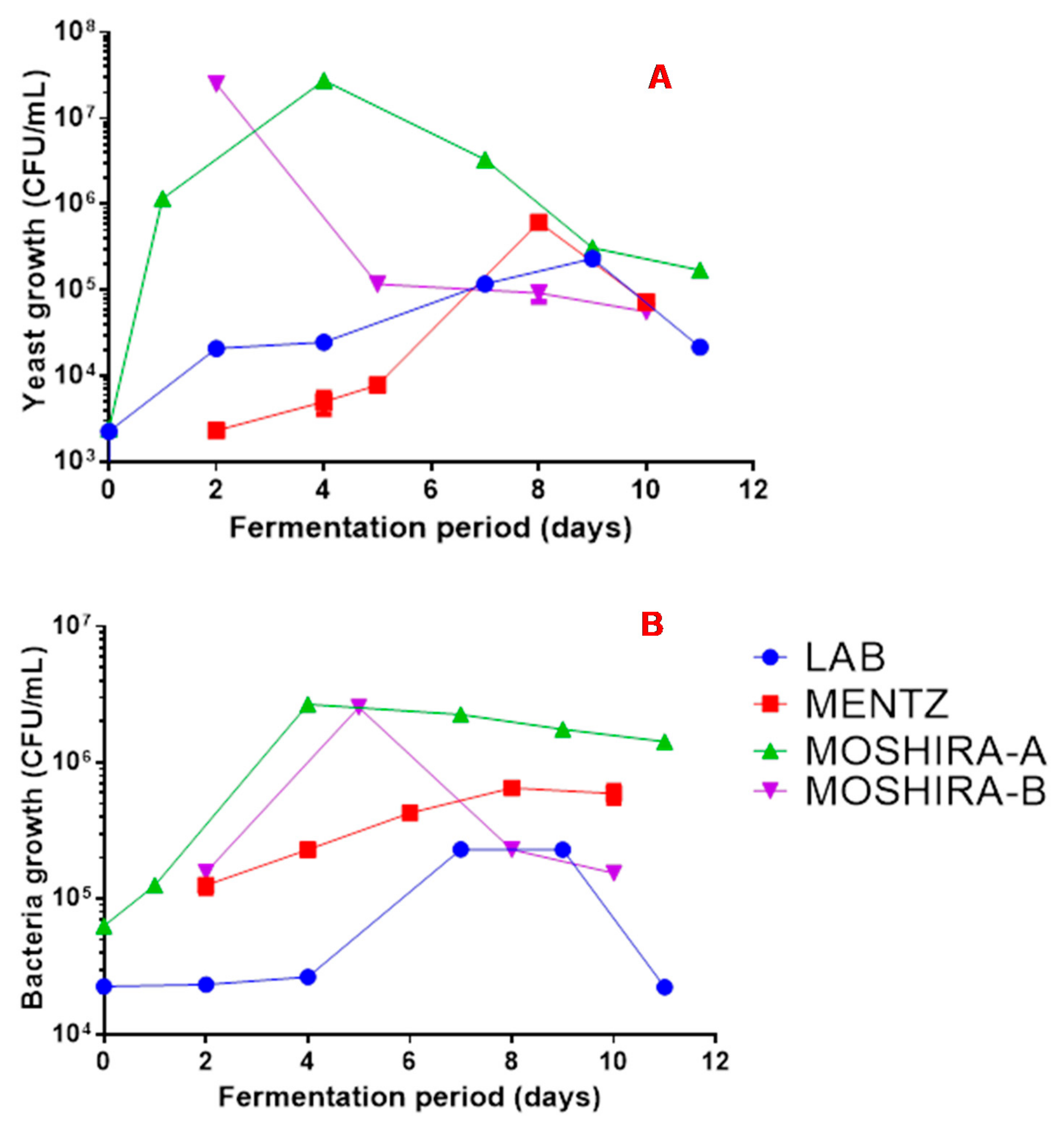
There were notable differences in the bacterial and yeast loads during fermentation of the Lab and Moshira marula wines. Both bacterial and yeast cell numbers reached higher levels in the wine prepared using the Moshira juice (Figure 3). Additionally, both bacterial and yeast numbers increased much faster in the initial stages of fermentation of the Moshira wine. The Moshira juice had a higher sucrose concentration at 80 mg/mL (Figure 1), and it declined to 40 mg/mL at the start of fermentation following dilution with water (Figure 2). Interestingly, faster sugar consumption was observed in the Moshira wine (Figure 2) within the first day of fermentation. This coincided with the rapid increase in microbial count relative to the observations made in the Mentz and Lab wines.
Figure 2.
Changes in the sucrose, glucose and fructose levels in the marula (A) Lab, (B) Mentz and (C) Moshira-A wines.
Figure 2.
Changes in the sucrose, glucose and fructose levels in the marula (A) Lab, (B) Mentz and (C) Moshira-A wines.

3.4. Yeast Dynamics during Fermentation
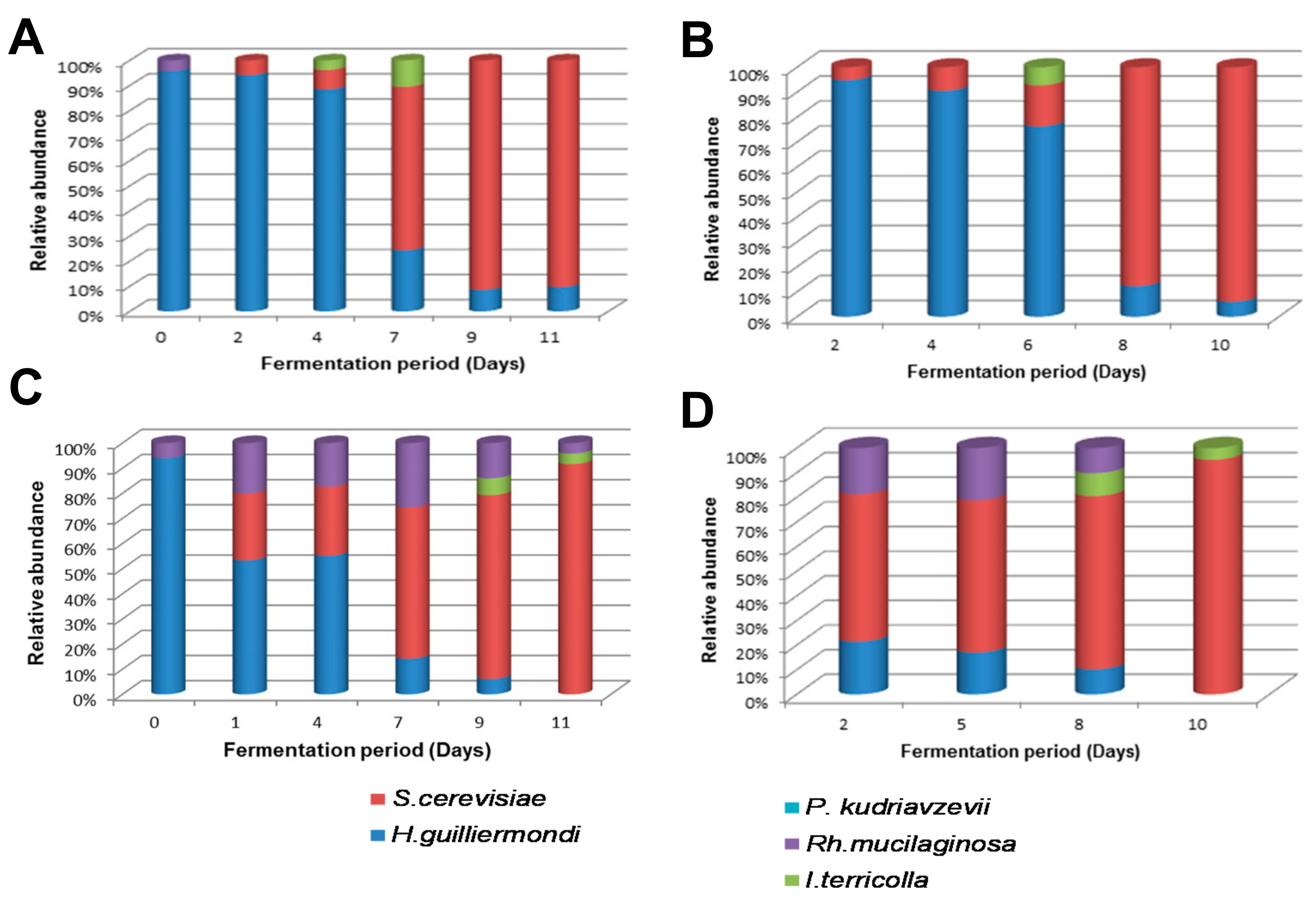
Fermenting and non-fermenting yeast species were isolated at various stages of the fermentation process, with clear evidence of displacement as fermentation progressed. The total yeast load for the Lab and Mentz wines was lower (2.22 × 104–2.31 × 104 CFU/mL) than the Moshira wines, which had higher yeast loads (1.17 × 106 CFU/mL and 2.50 × 107 CFU/mL) at Days 1 and 2 of fermentation, respectively. As expected, the profile of displacements of yeast species was the same for the Lab and Mentz marula wines that were produced from the same batch of marula juice. In the two Moshira wines, Saccharomyces cerevisiae started dominating earlier in the fermentation. Hanseniaspora guilliermondii, Saccharomyces cerevisiae, Issatchenkia terricola and Rhodotorula mucilaginosa were present in all the wines (Figure 4), with the non-fermenting H. guilliermondii dominating the early stages of fermentation, while the mid and late stages of fermentation were dominated by the fermenting yeast S. cerevisiae.
3.5. Bacterial Dynamics during Fermentation
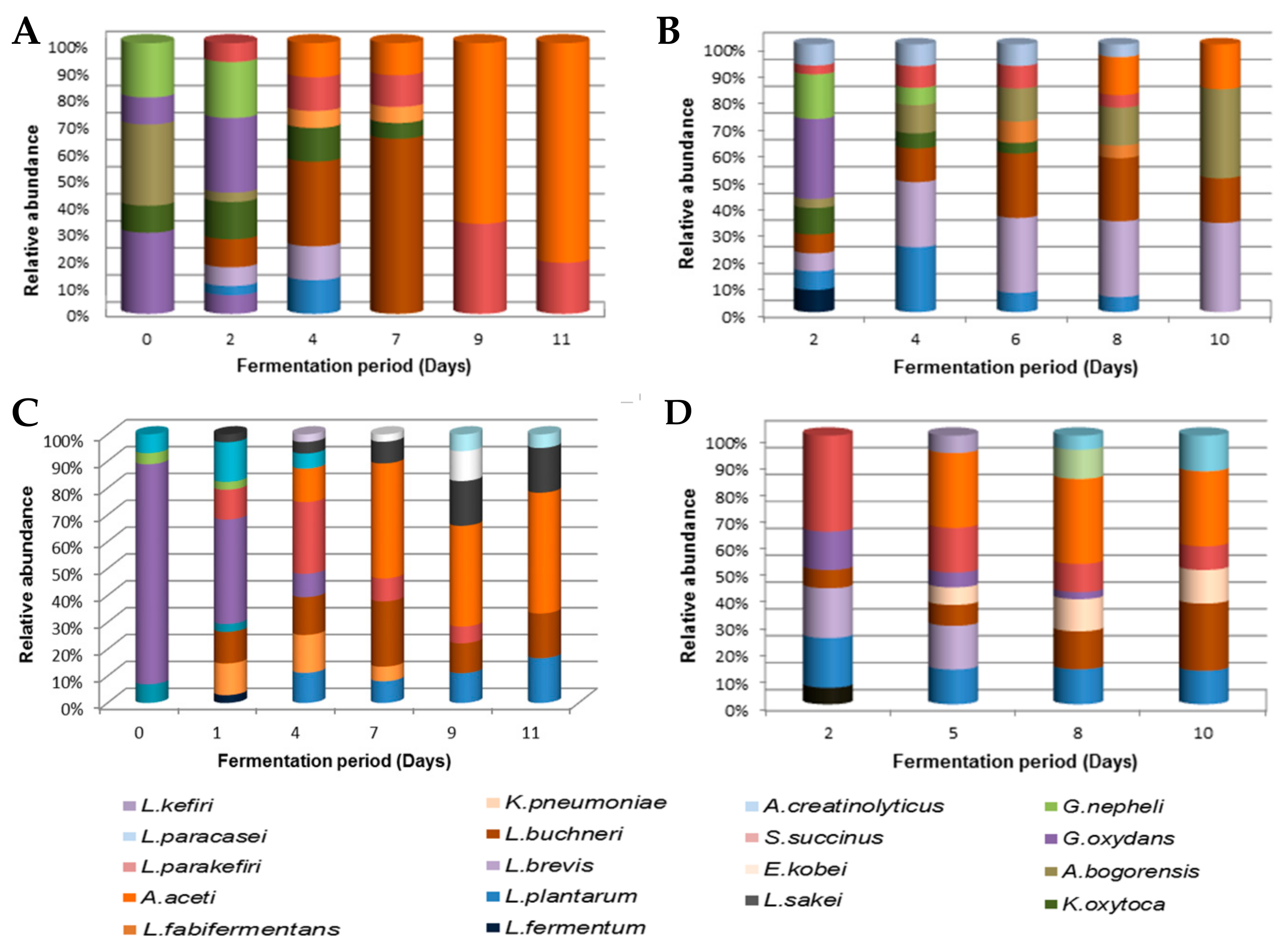
Unlike yeast profiles, there were no similarities in the bacterial profiles of wines prepared from the same batch of marula juice. Interestingly, the Lab wine that was kept under controlled laboratory conditions maintained a low bacterial load throughout, and the two Moshira wines both had higher bacterial loads of 1.26 × 105 CFU/mL and 1.57 × 105 CFU/mL, respectively. The spontaneously fermented marula wines were found to harbor various species of lactic acid bacteria and acetic acid bacteria. Lactic acid bacteria were dominated by Lactobacillus fermentum, Lactobacillus plantarum, Lactobacillus buchneri, Lactobacillus parabuchneri and Lactobacillus brevis, while acetic acid bacteria were dominated by Gluconobacter oxydans, Acetobacter pasteuriannus and Acetobacter aceti (Figure 5).
Lactobacillus spp. were present throughout the fermentation, with a particularly high prevalence during the first five days. Acetic acid bacteria such as G. nephelii and A. bogorensis were detected in low numbers during the early stages of fermentation, while the latter stages were dominated by A. aceti and A. pasteuriannus.
3.6. Volatile Compounds Evolution during Fermentation
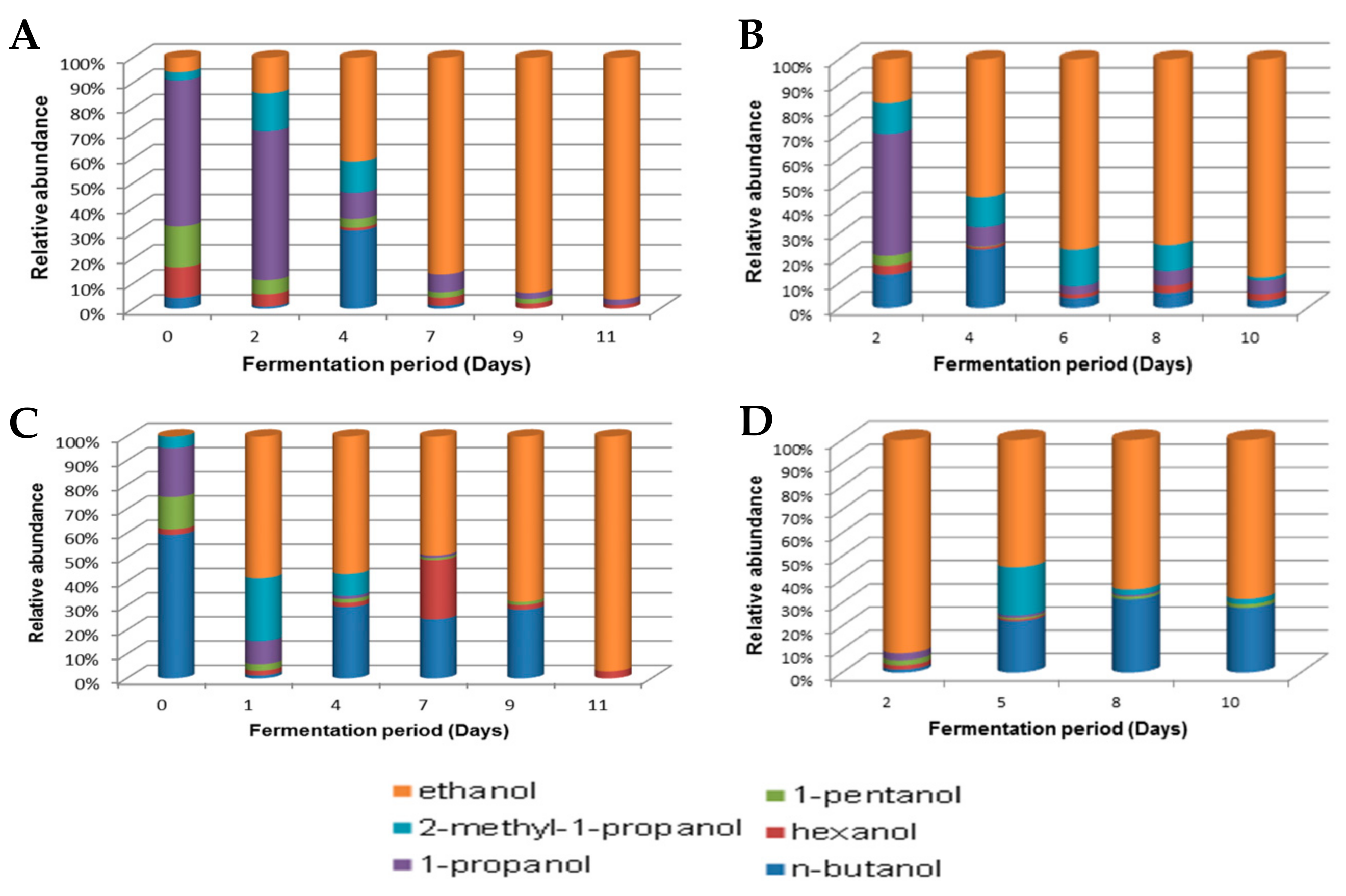
A number of organic compounds such as alcohols, volatile organic acids and aldehydes, which are commonly associated with fermented products, were observed during the fermentation of marula wines. The similarities between the Lab and Moshira wines in the evolution of compounds present in the respective wines were apparent (Figure 6, Figure 7 and Figure 8). Higher fusel alcohols such as 1-propanol, 1-pentanol, 2 methyl-1-propanol and hexanol, as well as acetate esters and aldehydes including isopentyl acetate and isobutyraldehyde, were present at relatively high concentrations during the early stages of the marula wine fermentations. Most of these compounds have been known to contribute to the aroma of the wine, and their levels decrease as fermentation time increases.
Alcoholic fermentation began on Day 2, and the ethanol content progressively increased and reached 90% abundance at the latter fermentation stage in all the wines (Figure 6). The dominance of ethanol expectedly coincided with the dominance of the fermenting yeast S. cerevisiae (Figure 4). It is noteworthy that a mixture of different alcohols was observed mainly on Days 0 to 2 of fermentation. The evolution of the different alcohols was similar in pattern in the Lab and Mentz wines, albeit at different amounts (Figure 6A,B). This was expected, as the two wines were biological duplicates. 1-Propanol was relatively the most abundant alcohol between Days 0 and 2 in the Lab and Mentz wines, although it diminished in concentration as fermentation progressed. On the contrary, the alcohol composition in the Moshira wines was different (Figure 6C,D); however, ethanol was abundant from the early fermentation stage.
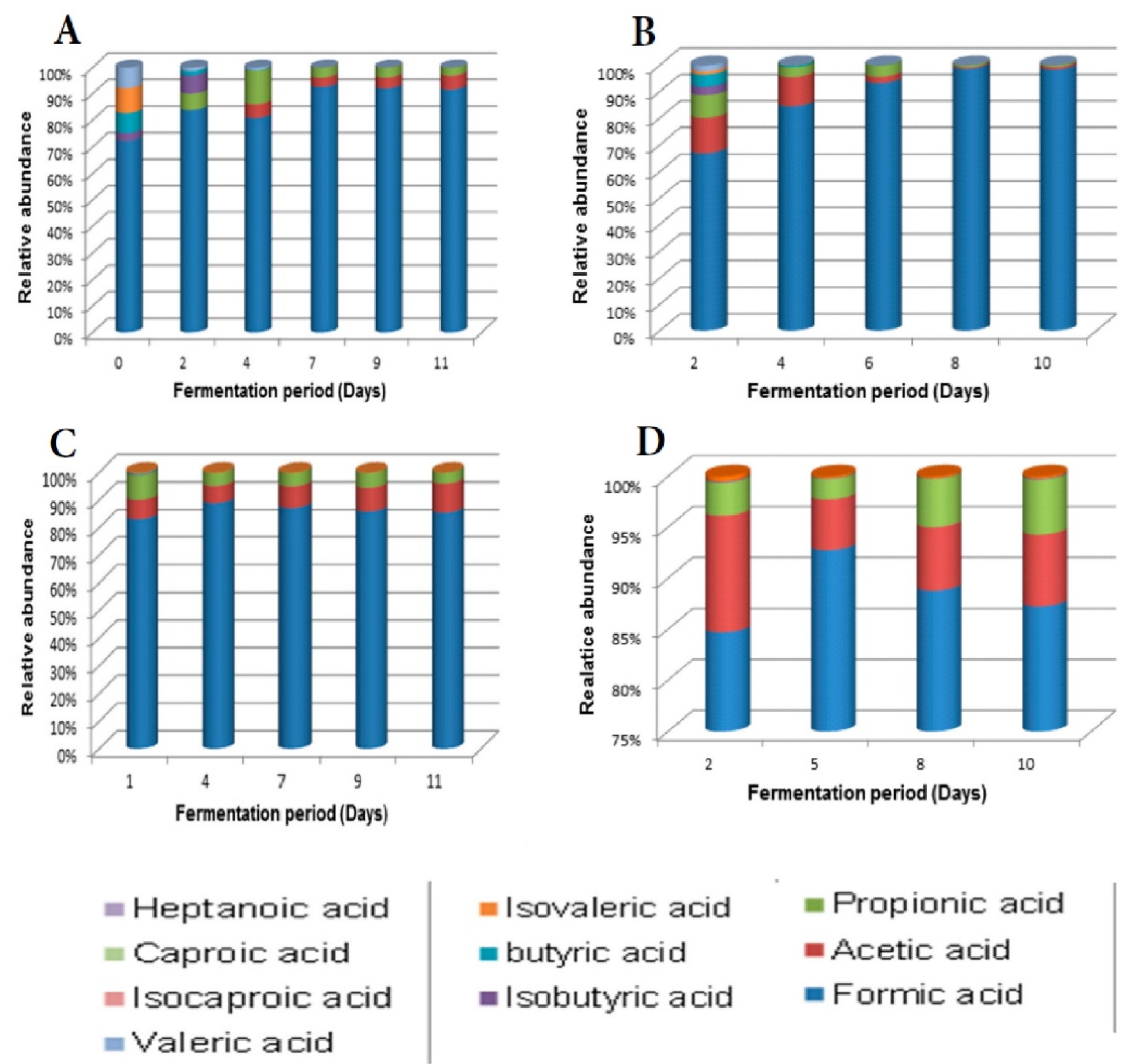
Of the organic acids tested, Mentz wine was the most divergent, with more VOCs than the other wines at the early fermentation stage. Acetic acid and formic acid were dominant throughout the fermentation period (Figure 7).
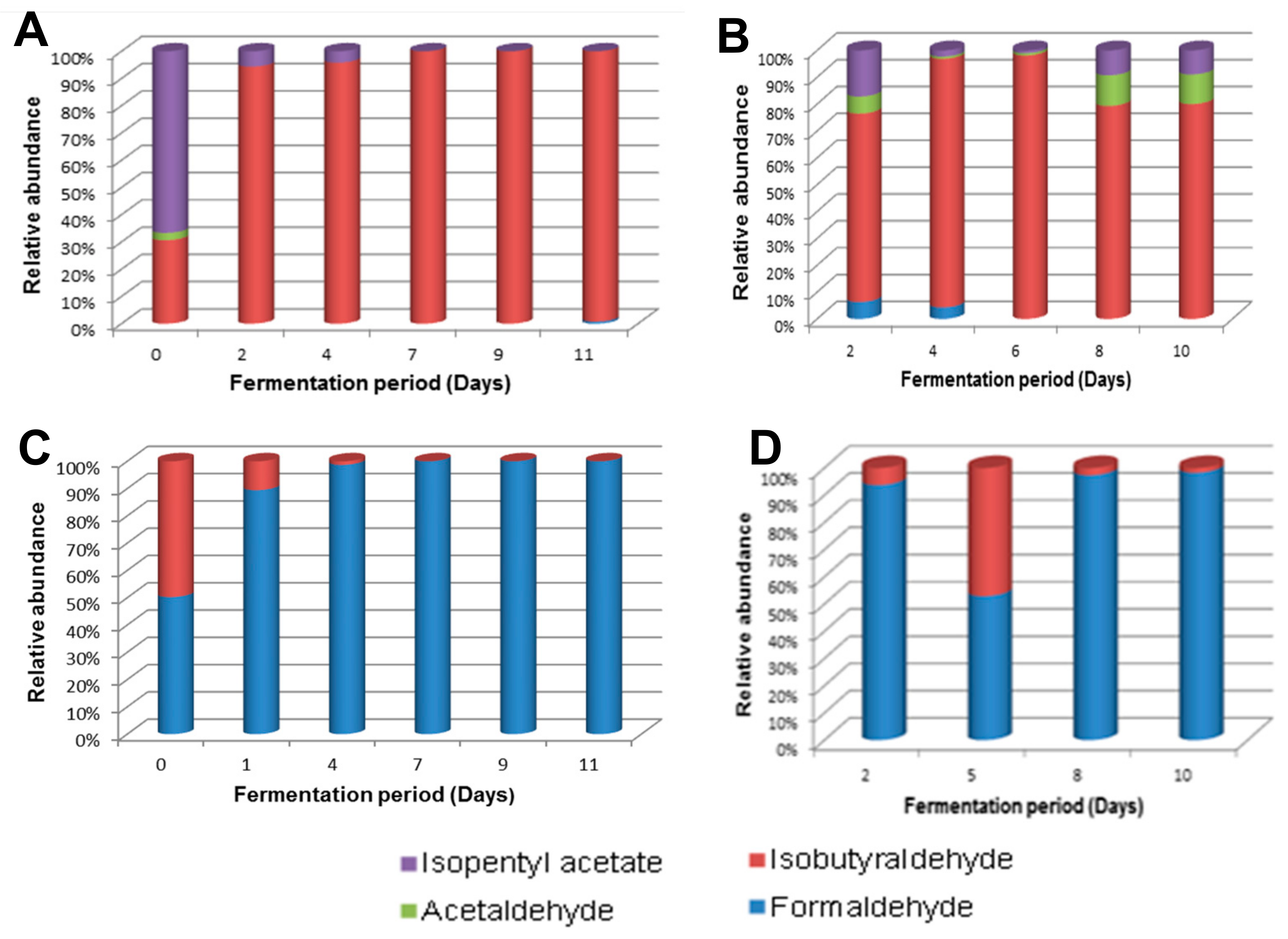
A clear distinction was observed between the Lab and Mentz wines, which are biological duplicates, and the two Moshira wines in terms of the tested aldehydes and esters. The Moshira wines contained solely isobutyraldehyde and formaldehyde, with the latter dominating throughout the fermentation period. In contrast, the Lab and Mentz wines contained isopentyl acetate and isobutyraldehyde as the dominant VOCs (Figure 8).
4. Discussion
The current study evaluated the chemical and microbial profiles of two sets of marula wines from juice to the stage where the wine was bitter and not palatable. This study sought to investigate the common contributing microbiota and chemicals to the characteristic taste and aroma of marula wine.
Sucrose has been reported to be the main sugar in the marula juice [18,19]. Climatic conditions and rainfall were reported to influence the juiciness and sugar content of the marula fruit [4], and subsequently, this will affect the organoleptic characteristics of the resulting wine, since the only ingredients used for making marula wine are marula fruit juice and water.
The decrease in the relative abundance of H. guilliermondii from the early stage of fermentation and the appearance of S. cerevisiae coincided with the transformation of sucrose to glucose and fructose from Day 4 onwards. S. cerevisiae is a glucophillic yeast [20]; hence, glucose depleted faster than fructose. The presence of the non-fermenting yeasts with low tolerance to ethanol [21], H. guilliermondii and I. terricola was reported previously by Wang and Liu [22]. They disappeared from the mid to the late stage of fermentation when ethanol production increased in the marula wine. A similar pattern of yeast evolution was reported in the alcoholic fermentation of grape juice where the non-Saccharomyces yeast species of Hanseniaspora, Candida, Pichia and Metschnikowia decreased as S. cerevisiae numbers increased [23]; however, some were known to persist throughout [24].
The succession of the bacteria observed in this study was similar to that reported by Dlamini and Dube [12]. It was apparent that the fermentation of the marula wine was mediated by a complex mixture of yeast and bacterial species. Bacteria such as the hetero-fermentative L. fermentum, L. plantarum, L. paracasei and L. brevis were present in high numbers in marula wines. Hetero-fermentative lactic acid bacteria have been reported previously in fermented foods [25]. A. aceti dominated in more acidic wines, whereas G. oxydans, which is often associated with sugar-rich environments [26], became undetectable when alcohol levels increased due to its low tolerance for ethanol [27]. Gluconobacter sp. prefers sugar as a carbon source, whereas Acetobacter sp. prioritizes the use of alcohol since it is moderately resistant to alcohol [28]. This explains the presence of A. aceti in higher numbers during the late stages of fermentation, although it was also detected in the early and mid-fermentation stages, similar to other studies on other types of wines [26,29,30]. Deemed the most active species in wine fermentation [28], Lactobacilli were reportedly responsible for acidification of other alcoholic beverages during the initial stages of fermentation [31,32], whilst the acetic acid bacteria were responsible for the acidification at the latter stages [31].
Generally, spontaneous fermentation of marula wine involves a complex interaction of the yeasts mainly Saccharomyces together with lactic acid and to a lesser extent acetic acid bacteria found on the surface of the fruits [12]. The yeasts and lactic acid bacteria have a symbiotic relationship in which the lactic acid bacteria produce a favorable acidic environment for the proliferation of various yeast genera [33], while the yeast contributes to the growth of the lactic acid bacteria through production of vitamin and other nutritional factors such as amino acids [34].
In alcoholic beverages, organic acids play a major role in contributing to flavor, color, and aromatic properties. Fusel alcohols such as 1-propanol, 1-pentanol, 2-methyl-1-propanol, n-butanol, hexanol were present in high proportions in all the marula wines during the early stages of fermentation, the period dominated by H. guilliermondii. H. guilliermondii and Pichia anomala are strong producers of aromatic compounds such as fusel alcohols and acetate esters 2-phenylethyl acetate and isoamyl acetate in wine [35,36]. Pretorius et al. [37] previously reported 1-pentanol and 1-hexanol to be the major alcohols in fresh marula pulp and a steady decline was observed with the progression of fermentation. Fusel alcohols are associated with off-flavors when present in large amounts, but they contribute to the aroma of the resulting wine in combination with their esters when present in low concentrations. These aroma characteristics can be used as an organoleptic fingerprint of a wine [38]. Ethanol was the major alcohol, and its production was associated with the presence of S. cerevisiae. Ethanol is an important volatile compound in wine and contributes to the wine’s strong and typical smell as well as taste [39]. Similarly, the bitter vinegary taste of unpalatable marula wine at the late stage of wine production was attributable to acetic acid produced by A. aceti and A. pasteuriannus [40].
Although different fruits may share most of the aromatic characteristics, differences also arise from the complex mixture of volatiles, which will inadvertently give a wine its distinctive aroma [41]. Compounds such as isopentyl acetate, isobutyraldehyde, formaldehyde and acetaldehyde have been reported to be present in small quantities in fermented beverages such as wine where they influence the flavor of the wine [39]. Such esters and aldehydes were noted at the initial stage and to a lesser extent at the later stage of the fermentation of marula wine. However, of the four VOCs tested, only isobutyraldehyde and formaldehyde were abundant throughout fermentation in all the wines, with a small amount of acetaldehyde in the Mentz wine only. Formaldehyde is produced during alcoholic fermentation through the oxidation of methanol [42]. While the process is regarded as complex, there are suggestions that Strecker aldehydes such as isobutyraldehyde are formed from their precursor alcohols through peroxidation [43]. The detection of formaldehyde in most alcoholic beverages such as whiskey, wine and beer does not preclude its undesirability as a health risk [42], and its presence should be regulated to prevent adverse effects if consumed in large quantities. On the other hand, the esters are produced from higher alcohols [44] and ethyl alcohol through catalysis by yeast alcohol acetyltransferase, alcohol dehydrogenase or other enzymatic activities by S. cerevisiae and yeasts such as the non-fermenting yeast species H. guilliermondii and P. kudriavzevii [45,46]. Esters are known to decline in concentration post fermentation and during storage due to non-enzymatic degradation [44]. This is congruent with the observation in this study, especially the Lab wine. Reduction of the amount of esters affects wine flavor and astringency during the long storage of wines [47]. Notably, the level of acetaldehyde can increase as the wine ages due to the oxidation of ethanol by the acetic acid bacteria such as A. aceti, which occur in the late stages of fermentation [40], as was observed in the marula wines in this current study. The acetate esters were reported to give a vinegar off-flavor [48,49] when present at levels above the sensory threshold. The common spoilage character in marula wines, which occurs generally after three weeks of production, is attributable to the presence of acetic acid at the latter fermentation stage. The presence of high acetic acid levels could have resulted from the oxidation of acetaldehyde, which was observed in the early stages of marula wine fermentation and from the oxidation of ethanol as well. The formation of these acetate esters is commonly attributed to the presence of oxygen in fermenting wines and the activities of yeasts such as H. guilliermondii, which produces larger amounts of acetate esters [46], whereas S. cerevisiae produces acetate esters in low amounts [35]. The corresponding presence of acetic acid bacterial species such as Acetobacter and Gluconobacter supports the formation of acetic acid in the marula wine.
5. Conclusions
This study provides information on the contributing microbiota in marula wine and their associated aromatic and flavor compounds. This information could be used in developing signature profiles of marula wine and would lay the foundation for the production of a good quality marula wine wherein the presence of microorganisms that give off undesirable characteristics can be regulated. The relationship revealed between the contributing and fermenting microorganisms and the flavor characteristics of marula wine is important for the regulation of good flavor quality. Consequently, the specific identification of functional strains from the traditional marula wine would be essential for establishing the commercial value in improving the overall flavor quality of marula wine.
Author Contributions
A.P. conducted the experimental work and drafted the manuscript; D.L.G. contributed to the conceptualization of the study and co-supervised the chemical profiling of the wines; K.M. conceptualized the study, supervised and prepared the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the National Research Foundation (NRF, South Africa) under Grant Number 93178. Any opinion, finding and conclusion or recommendation expressed in this material is that of the authors and the NRF does not accept any liability in this regard.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank the marula wine brewers R. Maipa and M. Kgoete for assistance with the production of the marula wine. Furthermore, the authors appreciate the traders of marula wine in Moshira in Sekhukhune for supporting this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Van Wyk, B.E. The potential of South African plants in the development of new food and beverage products. S. Afr. J. Botan. 2011, 77, 857–868. [Google Scholar] [CrossRef]
- Hiwilepo-van Hal, P.; Bille, P.G.; Verkerk, R.; Dekker, M. The effect of temperature and time on the quality of naturally fermented marula (Sclerocarya birrea subsp. caffra) juice. LWT-Food Sci. Technol. 2013, 53, 70–75. [Google Scholar] [CrossRef]
- Komane, B.; Vermaak, I.; Summers, B.; Viljoen, A. Safety and efficacy of Sclerocarya birrea (A.Rich.) Hochst (Marula) oil: A clinical perspective. J. Ethnopharmacol. 2015, 176, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Moganedi, K.; Sibara, M.; Grobler, P.; Goyvaerts, E. An assessment of genetic diversity among marula populations using the amplified fragment length polymorphism (AFLP) technique. Afr. J. Agric. Res. 2011, 6, 790–797. Available online: http://www.academicjournals.org/AJAR (accessed on 5 September 2018).
- Shackleton, S.E.; Shackleton, C.M.; Cunningham, T.; Lombard, C.; Sullivan, C.A.; Netshiluvhi, T.R. Knowledge on Sclerocarya birrea subsp. caffra with emphasis on its importance as a non-timber forest product in South and southern Africa: A Summary. S. Afr. For. J. 2002, 194, 27–41. [Google Scholar] [CrossRef]
- Dube, S.; Shereni, I.; Dlamini, N.R.; Sibanda, T. Extending the shelf life of fresh marula (Sclerocarya birrea) juice by altering its physico-chemical parameters. In Biochemical Testing; Jimenez-Lopez, J.C., Ed.; INTECH Open: London, UK, 2012; pp. 181–196. [Google Scholar] [CrossRef]
- Hiwilepo-van Hal, P.; Bosschaart, C.; van Twisk, C.; Verkerk, R.; Dekker, M. Kinetics of thermal degradation of vitamin C in marula fruit (Sclerocarya birrea subsp. caffra) as compared to other selected tropical fruits. LWT-Food Sci. Technol. 2012, 49, 188–191. [Google Scholar] [CrossRef]
- Simatende, P.; Gadaga, T.H.; Nkambule, S.J.; Siwela, M. Methods of preparation of Swazi traditional fermented foods. J. Eth. Foods 2015, 2, 119–125. [Google Scholar] [CrossRef]
- Leakey, R.R.B. Potential for novel food products from agroforestry trees. Food Chem. 1999, 66, 1–14. [Google Scholar] [CrossRef]
- Mokgolodi, N.C.; Ding, Y.; Setshogo, M.P.; Ma, C.; Liu, Y. The importance of an indigenous tree of Southern African communities with specific reference to its domestication and commercialization: A case of the marula tree. For. Stud. China 2011, 13, 36–44. [Google Scholar] [CrossRef]
- Okagbue, R.; Siwela, M. Yeasts and related microorganisms isolated from ripe marula fruits (Sclerocarya caffra) in Zimbabwe. S. Afr. J. Sci. 2002, 98, 551–552. Available online: https://hdl.handle.net/10520/EJC97428 (accessed on 9 August 2018).
- Dlamini, N.R.; Dube, S. Studies on the physico-chemical, nutritional and microbiological changes during the traditional preparation of marula wine in Gwanda, Zimbabwe. Nutr. Food Sci. 2008, 38, 61–69. [Google Scholar] [CrossRef]
- Mpofu, A.; Kock, J.L.F.; Pretorious, E.E.; Pohl, C.H.; Zvauya, R. Identification of yeast isolated from mukumbi, a Zimbabwean traditional wine. J. Sus. Dev. Afr. 2008, 10, 88–102. [Google Scholar]
- Prins, W.A.; Botha, M.; Botes, M.; de Kwaadsteniet, M.; Endo, A.; Dicks, L.M.T. Lactobacillus plantarum 24, isolated from the marula fruit (Sclerocarya birrea), has probiotic properties and harbours genes encoding the production of three bacteriocins. Curr. Microbiol. 2010, 61, 584–589. [Google Scholar] [CrossRef]
- Sambrook, J.; Rusell, D.W. Rapid isolation of yeast, DNA. In Molecular Cloning, a Laboratory Manual; Sambrook, J., Rusell, D.W., Eds.; Cold Harbor Laboratory Press: New York, NY, USA, 2001; pp. 631–632. [Google Scholar]
- Neumann, B.; Pospiech, A.; Schairer, H. Rapid isolation of genomic DNA from gram-negative bacteria. Trends Genet. 1992, 8, 332–333. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., Eds.; Academic Press Incorporated: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Fundira, M. Optimization of Fermentation Processes for the Production of Indigenous Fruit Wines (Marula). Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2001. [Google Scholar]
- Weinert, I.A.G.; van Wyk, P.J.; Holtzhausen, L.C. Marula. In Fruits of Tropical and Subtropical Origin: Composition, Properties and Uses; Nagy, S., Shaw, P.E., Wardowski, W.F., Eds.; Florida Science Source, Inc.: Lake Alfred, FL, USA, 1990; pp. 88–113. [Google Scholar]
- Berthels, N.; Cordero-Otero, R.; Bauer, F.; Thevelein, J.; Pretorius, I. Discrepancy in glucose and fructose utilization during fermentation by Saccharomyces cerevisiae wine yeast strains. FEMS Yeast Res. 2004, 4, 683–689. [Google Scholar] [CrossRef]
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The effect of non-Saccharomyces yeasts on fermentation and wine quality. S. Afri. J. Enol. Vitic. 2003, 24, 55–62. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Y. Dynamic study of yeast species and Saccharomyces cerevisiae strains during the spontaneous fermentations of Muscat blac in Jingayang, China. Food Microbiol. 2013, 33, 172–177. [Google Scholar] [CrossRef]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Díaz, C.; Molina, A.M.; Nähring, J.; Fischer, R. Characterization and dynamic behaviour of wild yeast during spontaneous fermentation in steel tanks and amphorae. Biomed. Res. Int. 2013, 2013, 540465. [Google Scholar] [CrossRef]
- Laureys, D.; De Vuyst, L. Microbial species diversity, community dynamics, and metabolite kinetics of water kefir fermentation. Appl. Environ. Microbiol. 2014, 80, 2564–2572. [Google Scholar] [CrossRef]
- Bartowsky, E.J.; Henschke, P.A. Acetic acid bacteria spoilage of bottled red wine—A review. Int. J. Food Microbiol. 2008, 125, 60–70. [Google Scholar] [CrossRef]
- De Ley, J.; Gillis, M.; Swings, J.; Family, V.I. Acetobacteraceae. In Bergey’s Manual of Systematic Bacteriology; Krieg, N.R., Holt, J.G., Eds.; Williams & Wilkins: Baltimore, MD, USA, 1984; pp. 267–278. [Google Scholar]
- Berbegal, C.; Peña, N.; Russo, P.; Grieco, F.; Pardo, I.; Ferrer, S.; Spano, G.; Capozzi, V. Technological properties of Lactobacillus plantarum strains isolated from grape must fermentation. Food Microbiol. 2016, 57, 187–194. [Google Scholar] [CrossRef]
- Pinto, L.; Malfeito-Ferreira, M.; Quintieri, L.; Silva, A.C.; Baruzzi, F. Growth and metabolite production of a grape sour rot yeast-bacterium consortium on different carbon sources. Int. J. Food Microbiol. 2019, 296, 65–74. [Google Scholar] [CrossRef]
- Xiangyu, Q.; Linlin, Y.; Weiying, W.; Riming, Y.; Zhibin, Z.; Huilin, Y.; Du, Z.; Bo, Z. Comparative Evaluation of Microbiota Dynamics and Metabolites Correlation between Spontaneous and Inoculated Fermentations of Nanfeng Tangerine Wine. Front. Microbiol. 2021, 12, 649978. [Google Scholar] [CrossRef]
- Amoa-Awua, W.K.; Sampson, E.; Tano-Debrah, K. Growth of yeasts, lactic and acetic acid bacteria in palm wine during tapping and fermentation from felled oil palm (Elaeis guineensis) in Ghana. J. Appl. Microbiol. 2007, 102, 599–606. [Google Scholar] [CrossRef]
- Atter, A.; Obiri-Danso, K.; Amoa-Awua, W.K. Microbiological and chemical processes associated with the production of burukutu a traditional fermented beer in Ghana. Int. Food Res. J. 2014, 21, 1769–1776. Available online: https://www.researchgate.net/publication/287292436 (accessed on 13 February 2018).
- Wilson, P.; David, T.; Sam, B. Microbial and chemical changes occuring during production of traditional Rwandese banana beer urwagwa. J. Ferm. Technol. 2012, 1, 104. [Google Scholar] [CrossRef]
- Jerpersen, L. Occurrence and taxonomic characteristics of strains of Saccharomyces cerevisiae predominant in African indigenous fermented foods and beverages. FEMS Yeast Res. 2003, 3, 191–200. [Google Scholar] [CrossRef]
- Gamero, A.; Quintilla, R.; Groenewald, M.; Alkema, W.; Boekhout, T.; Hazelwood, L. High-throughput of a large collection of non-conventional yeasts reveals their potential for aroma formation in food fermentation. Food Microbiol. 2016, 60, 147–159. [Google Scholar] [CrossRef]
- Rojas, V.; Gil, J.; Pinaga, F.; Manzanares, P. Studies on acetate ester production by non-Saccharomyces wine yeast. Int. J. Food Microbiol. 2001, 70, 283–289. [Google Scholar] [CrossRef]
- Pretorius, V.; Rohmer, E.; Rapp, A.; Holtzhausen, L.C.; Mandery, H. Volatile flavour components of marula juice. Z. Lebensmittel. Fors. 1985, 181, 458–461. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Esteban-Fernández, A.; Navascués, E.; Marquina, D.; Santos, A.; Moreno-Arribas, M.V. Microbial contribution to wine aroma and its extended use for wine quality improvement. Molecules 2017, 22, 189. [Google Scholar] [CrossRef]
- Shale, K.; Mukamugema, K.J.; Lues, R.J.; Venter, P.; Mokoena, K.K. Characterization of selected volatile organic compounds in Rwandan indigenous beer urwagwa by dynamic headspace gas chromatography-mass spectometry. Afr. J. Biotechnol. 2013, 12, 2990–2996. [Google Scholar] [CrossRef]
- Torija, M.J.; Mateo, E.; Guillamón, J.M.; Mas, A. Identification and quantification of acetic acid bacteria in wine and vinegar by TaqMan–MGB probes. Food Microbiol. 2010, 27, 257–265. [Google Scholar] [CrossRef]
- Tucker, G.A. Introduction. In Biochemistry of Fruit Ripening; Seymour, G.B., Taylor, J.E., Tucker, G.A., Eds.; Chapman & Hall: London, UK, 1993; pp. 1–55. [Google Scholar]
- Lago, L.O.; Welke, J.E. Carbonyl compounds in wine: Factors related to presence and toxic effects. Food Technol. 2019, 49, e20190349. [Google Scholar] [CrossRef]
- Bueno, M.; Marrufo-Curtido, A.; Carrascon, V.; Fernandez-Zurbano, P.; Escudero, A.; Ferreira, V. Formation and accumulation of acetaldehyde and Strecker aldehydes during red wine oxidation. Front. Chem. 2018, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Cordente, A.G.; Espinase Nandorfy, D.; Solomon, M.; Schulkin, A.; Kolouchova, R.; Francis, I.L.; Schmidt, S.A. Aromatic higher alcohols in wine: Implication on aroma and palate attributes during chardonnay aging. Molecules 2021, 26, 4979. [Google Scholar] [CrossRef]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of yeast species and strains in wine flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Badura, J.; van Wyk, N.; Brezina, S.; Pretorius, I.S.; Rauhut, D.; Wendland, J.; von Wallbrunn, C. Development of Genetic Modification Tools for Hanseniaspora uvarum. Int. J. Mol. Sci. 2021, 22, 1943. [Google Scholar] [CrossRef]
- Han, G.; Webb, M.R.; Waterhouse, A.L. Acetyladehyde reactions during wine bottle storage. Food Chem. 2019, 290, 208–215. [Google Scholar] [CrossRef]
- Guth, H. Quantification and sensory studies of character impact odorants of different white wine varieties. J. Agric. Food Chem. 1997, 45, 3027–3032. [Google Scholar] [CrossRef]
- Ferreira, V.; Lopéz, R.; Cacho, J.F. Quantification determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agric. 2000, 80, 1659–1667. [Google Scholar] [CrossRef]
Figure 1.
Sucrose, glucose and fructose content in marula juice from UL and Moshira.

Figure 3.
Yeast (A) and bacterial (B) content during marula wine fermentation.

Figure 4.
Yeast species identified in marula wines from Lab (A), Mentz (B), Moshira-A (C) and Moshira-B (D).
Figure 4.
Yeast species identified in marula wines from Lab (A), Mentz (B), Moshira-A (C) and Moshira-B (D).

Figure 5.
Bacterial species identified in marula wine from (A) Lab, (B) Mentz, (C) Moshira-A and (D) Moshira-B.
Figure 5.
Bacterial species identified in marula wine from (A) Lab, (B) Mentz, (C) Moshira-A and (D) Moshira-B.

Figure 6.
Alcohols during fermentation of the marula wines from Lab (A), Mentz (B), Moshira-A (C) and Moshira-B (D).
Figure 6.
Alcohols during fermentation of the marula wines from Lab (A), Mentz (B), Moshira-A (C) and Moshira-B (D).

Figure 7.
Volatile organic acids during fermentation of marula wine from (A) Lab, (B) Mentz, (C) Moshira-A and (D) Moshira-B.
Figure 7.
Volatile organic acids during fermentation of marula wine from (A) Lab, (B) Mentz, (C) Moshira-A and (D) Moshira-B.

Figure 8.
Aldehydes and esters during fermentation of marula wine from (A) Lab, (B) Mentz, (C) Moshira-A and (D) Moshira-B.
Figure 8.
Aldehydes and esters during fermentation of marula wine from (A) Lab, (B) Mentz, (C) Moshira-A and (D) Moshira-B.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Phiri, A.; La Grange, D.; Moganedi, K. Microbial and Chemical Dynamics during Marula Wine Fermentation. Beverages 2022, 8, 50. https://0-doi-org.brum.beds.ac.uk/10.3390/beverages8030050
AMA Style
Phiri A, La Grange D, Moganedi K. Microbial and Chemical Dynamics during Marula Wine Fermentation. Beverages. 2022; 8(3):50. https://0-doi-org.brum.beds.ac.uk/10.3390/beverages8030050
Chicago/Turabian StylePhiri, Archie, Daniel La Grange, and Kgabo Moganedi. 2022. "Microbial and Chemical Dynamics during Marula Wine Fermentation" Beverages 8, no. 3: 50. https://0-doi-org.brum.beds.ac.uk/10.3390/beverages8030050
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.