Chemical Characterization, Antioxidant, Enzyme Inhibition and Antimutagenic Properties of Eight Mushroom Species: A Comparative Study



Abstract
:1. Introduction
2. Materials and Methods
2.1. Mushrooms Material and Preparation of Extracts
2.2. Assay for Total Phenolic Content
2.3. LC–MS/MS Quantification of Phenolic Compounds
2.4. Determination of Antioxidant and Enzyme Inhibitory Effects
2.5. Antimutagenic/Mutagenic Properties
2.6. Data Analysis
3. Results and Discussion
3.1. Chemical Composition
3.2. Antioxidant Effects
3.3. Enzyme Inhibition Effects
3.4. Determination of Mutagenicity
3.5. Antimutagenic Evaluation
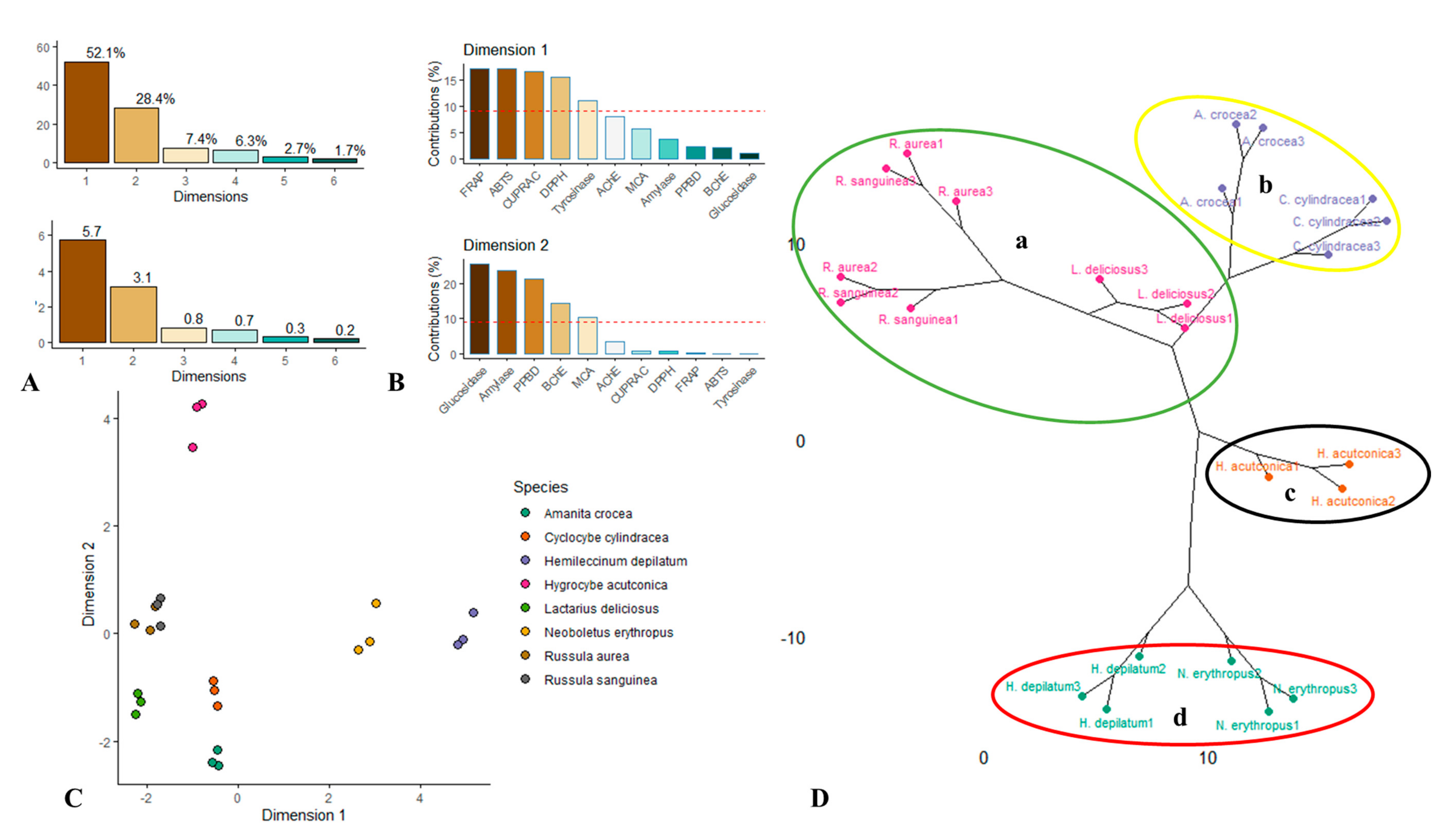
3.6. Unsupervised Multivariate Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zengin, G.; Sarikurkcu, C.; Gunes, E.; Uysal, A.; Ceylan, R.; Uysal, S.; Gungor, H.; Aktumsek, A. Two Ganoderma species: Profiling of phenolic compounds by HPLC–DAD, antioxidant, antimicrobial and inhibitory activities on key enzymes linked to diabetes mellitus, Alzheimer’s disease and skin disorders. Food Funct. 2015, 6, 2794–2802. [Google Scholar] [CrossRef] [PubMed]
- Georgescu, A.A.; Danet, A.F.; Radulescu, C.; Stihi, C.; Dulama, I.D.; Buruleanu, C.L. Nutritional and Food Safety Aspects Related to the Consumption of Edible Mushrooms from Dambovita County in Correlation with their Levels of Some Essential and Non-essential Metals. Rev. Chim. 2017, 68, 2402–2406. [Google Scholar] [CrossRef]
- Kavishree, S.; Hemavathy, J.; Lokesh, B.R.; Shashirekha, M.N.; Rajarathnam, S. Fat and fatty acids of Indian edible mushrooms. Food Chem. 2008, 106, 597–602. [Google Scholar] [CrossRef]
- Akyuz, M.; Kirbag, S. Antimicrobial activity of Pleurotus eryngii var. ferulae grown on various agro-wastes. EurAsian J. Biosci. 2009, 3, 58–63. [Google Scholar] [CrossRef]
- Breene, W.M. Nutritional and Medicinal Value of Specialty Mushrooms. J. Food Prot. 1990, 53, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Shao, Y.; Zhang, Z.; Wang, L.; Mariga, A.M.; Pang, G.; Geng, C.; Ho, C.-T.; Hu, Q.; Zhao, L. Regulation of human cytokines by Cordyceps militaris. J. Food Drug Anal. 2014, 22, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Kosanić, M.; Ranković, B.; Rančić, A.; Stanojković, T. Evaluation of metal concentration and antioxidant, antimicrobial, and anticancer potentials of two edible mushrooms Lactarius deliciosus and Macrolepiota procera. J. Food Drug Anal. 2016, 24, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Akata, I.; Seslİ, E. Four New Records of Basidiomycota for the Turkish Mycota From Trabzon and İstanbul Provinces. J. Fungus 2017, 8, 168–177. [Google Scholar] [CrossRef]
- IŞik, H.; TÜrkekul, İ. Leucopaxillus lepistoides: A New Record for Turkish Mycota from Yozgat Province. SDU J. Nat. Appl. Sci 2018, 22, 402. [Google Scholar] [CrossRef]
- Keleş, A. New records of Hymenoscyphus, Parascutellinia, and Scutellinia for Turkey. Mycotaxon 2019, 134, 169–175. [Google Scholar] [CrossRef]
- Uzun, Y.; Acar, İ. A New Inocybe (Fr.) Fr. Record for Turkish Macrofungi. Anatol. J. Bot. 2018, 2, 10–12. [Google Scholar] [CrossRef] [Green Version]
- Uzun, Y.; Kaya, A. First records of Hydnobolites and Pachyphlodes species from Turkey. Mycotaxon 2018, 133, 415–421. [Google Scholar] [CrossRef]
- Uzun, Y.; Kaya, A. New Additions to Turkish Pezizales from the Eastern Black Sea Region. Turk. J. Bot. 2019, 43, 262–270. [Google Scholar] [CrossRef]
- Vladimir-Knežević, S.; Blažeković, B.; Bival Štefan, M.; Alegro, A.; Kőszegi, T.; Petrik, J. Antioxidant activities and polyphenolic contents of three selected Micromeria species from Croatia. Molecules 2011, 16, 1454–1470. [Google Scholar] [CrossRef] [Green Version]
- Zengin, G.; Aktumsek, A. Investigation of antioxidant potentials of solvent extracts from different anatomical parts of Asphodeline anatolica E. Tuzlaci: An endemic plant to Turkey. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Orčić, D.; Francišković, M.; Bekvalac, K.; Svirčev, E.; Beara, I.; Lesjak, M.; Mimica-Dukić, N. Quantitative determination of plant phenolics in Urtica dioica extracts by high-performance liquid chromatography coupled with tandem mass spectrometric detection. Food Chem. 2014, 143, 48–53. [Google Scholar] [CrossRef]
- Grochowski, D.M.; Uysal, S.; Aktumsek, A.; Granica, S.; Zengin, G.; Ceylan, R.; Locatelli, M.; Tomczyk, M. In vitro enzyme inhibitory properties, antioxidant activities, and phytochemical profile of Potentilla thuringiaca. Phytochem. Lett. 2017, 20, 365–372. [Google Scholar] [CrossRef]
- Maron, D.M.; Ames, B.N. Revised methods for the Salmonella mutagenicity test. Mutat. Res. Environ. Mutagen. Relat. Subj. 1983, 113, 173–215. [Google Scholar] [CrossRef]
- Dean, B.; Brooks, T.; Hodson-Walker, G.; Hutson, D. Genetic toxicology testing of 41 industrial chemicals. Mutat. Res. Rev. Genet. Toxicol. 1985, 153, 57–77. [Google Scholar] [CrossRef]
- Zengin, G.; Uysal, A.; Gunes, E.; Aktumsek, A. Survey of Phytochemical Composition and Biological Effects of Three Extracts from a Wild Plant (Cotoneaster nummularia Fisch et Mey.): A Potential Source for Functional Food Ingredients and Drug Formulations. PLoS ONE 2014, 9, e113527. [Google Scholar] [CrossRef]
- Negi, P.; Jayaprakasha, G.; Jena, B. Antioxidant and antimutagenic activities of pomegranate peel extracts. Food Chem. 2003, 80, 393–397. [Google Scholar] [CrossRef]
- Rashmi, H.B.; Negi, P.S. Phenolic acids from vegetables: A review on processing stability and health benefits. Food Res. Int. 2020, 136, 109298. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.C.; Paiva de Sousa, C.; Fernandez-Prada, C.; Harel, J.; Dubreuil, J.D.; de Souza, E.L. A review of the current evidence of fruit phenolic compounds as potential antimicrobials against pathogenic bacteria. Microb. Pathog. 2019, 130, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Alu’datt, M.H.; Rababah, T.; Alhamad, M.N.; Al-Mahasneh, M.A.; Almajwal, A.; Gammoh, S.; Ereifej, K.; Johargy, A.; Alli, I. A review of phenolic compounds in oil-bearing plants: Distribution, identification and occurrence of phenolic compounds. Food Chem. 2017, 218, 99–106. [Google Scholar] [CrossRef]
- Leal, A.R.; Barros, L.; Barreira, J.C.M.; Sousa, M.J.; Martins, A.; Santos-Buelga, C.; Ferreira, I.C.F.R. Portuguese wild mushrooms at the “pharma–nutrition” interface: Nutritional characterization and antioxidant properties. Food Res. Int. 2013, 50, 1–9. [Google Scholar] [CrossRef]
- Onbaşılı, D.; Çelik, G.Y.; Katırcıoğlu, H.; Narin, I. Total phenolic, total flavonoid content and the DPPH free radical scavenging activity of Lactarius deliciosus (L.). Curr. Opin. Biotechnol. 2013, 24, S113. [Google Scholar] [CrossRef]
- Ferreira, I.C.F.R.; Baptista, P.; Vilas-Boas, M.; Barros, L. Free-radical scavenging capacity and reducing power of wild edible mushrooms from northeast Portugal: Individual cap and stipe activity. Food Chem. 2007, 100, 1511–1516. [Google Scholar] [CrossRef]
- Sánchez-Rangel, J.C.; Benavides, J.; Heredia, J.B.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. The Folin–Ciocalteu assay revisited: Improvement of its specificity for total phenolic content determination. Anal. Methods 2013, 5, 5990–5999. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Yan, Z.; Zhong, Y.; Duan, Y.; Chen, Q.; Li, F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim. Nutr. 2020, 6, 115–123. [Google Scholar] [CrossRef]
- Wong, F.-C.; Xiao, J.; Wang, S.; Ee, K.-Y.; Chai, T.-T. Advances on the antioxidant peptides from edible plant sources. Trends Food Sci. Technol. 2020, 99, 44–57. [Google Scholar] [CrossRef]
- Bahadori, M.B.; Sarikurkcu, C.; Yalcin, O.U.; Cengiz, M.; Gungor, H. Metal concentration, phenolics profiling, and antioxidant activity of two wild edible Melanoleuca mushrooms (M. cognata and M. stridula). Microchem. J. 2019, 150, 104172. [Google Scholar] [CrossRef]
- Krüzselyi, D.; Móricz, Á.M.; Vetter, J. Comparison of different morphological mushroom parts based on the antioxidant activity. LWT 2020, 127, 109436. [Google Scholar] [CrossRef]
- Garrab, M.; Edziri, H.; El Mokni, R.; Mastouri, M.; Mabrouk, H.; Douki, W. Phenolic composition, antioxidant and anticholinesterase properties of the three mushrooms Agaricus silvaticus Schaeff., Hydnum rufescens Pers. and Meripilus giganteus (Pers.) Karst. in Tunisia. S. Afr. J. Bot. 2019, 124, 359–363. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Farvin, K.H.S.; Surendraraj, A.; Al-Ghunaim, A.; Al-Yamani, F. Chemical profile and antioxidant activities of 26 selected species of seaweeds from Kuwait coast. J. Appl. Phycol. 2019, 31, 2653–2668. [Google Scholar] [CrossRef]
- Sevindik, M.; Akgul, H.; Bal, C.; Selamoglu, Z. Phenolic contents, oxidant/antioxidant potential and heavy metal levels in Cyclocybe cylindracea. Indian J. Pharm. Educ. Res. 2018, 52, 437–441. [Google Scholar] [CrossRef] [Green Version]
- Dorababu, A. Critical evaluation of current Alzheimer’s drug discovery (2018–19) and futuristic Alzheimer drug model approach. Bioorg. Chem. 2019, 93, 103299. [Google Scholar] [CrossRef]
- Ahangarpour, A.; Sayahi, M.; Sayahi, M. The antidiabetic and antioxidant properties of some phenolic phytochemicals: A review study. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 854–857. [Google Scholar] [CrossRef]
- Chinsembu, K.C. Diabetes mellitus and nature’s pharmacy of putative antidiabetic plants. J. Herb. Med. 2019, 15, 100230. [Google Scholar] [CrossRef]
- Mishra, P.; Kumar, A.; Panda, G. Anti-cholinesterase hybrids as multi-target-directed ligands against Alzheimer’s disease (1998–2018). Biorg. Med. Chem. 2019, 27, 895–930. [Google Scholar] [CrossRef] [PubMed]
- Akata, I.; Zengin, G.; Picot, C.M.N.; Mahomoodally, M.F. Enzyme inhibitory and antioxidant properties of six mushroom species from the Agaricaceae family. S. Afr. J. Bot. 2019, 120, 95–99. [Google Scholar] [CrossRef]
- Saidi, I.; Nimbarte, V.D.; Schwalbe, H.; Waffo-Téguo, P.; Harrath, A.H.; Mansour, L.; Alwasel, S.; Ben Jannet, H. Anti-tyrosinase, anti-cholinesterase and cytotoxic activities of extracts and phytochemicals from the Tunisian Citharexylum spinosum L.: Molecular docking and SAR analysis. Bioorg. Chem. 2020, 102, 104093. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G.; Uren, M.C.; Kocak, M.S.; Gungor, H.; Locatelli, M.; Aktumsek, A.; Sarikurkcu, C. Antioxidant and enzyme inhibitory activities of extracts from wild mushroom species from Turkey. Int. J. Med. Mushrooms 2017, 19, 327–336. [Google Scholar] [CrossRef]
- Yuan, Y.; Jin, W.; Nazir, Y.; Fercher, C.; Blaskovich, M.A.; Cooper, M.A.; Barnard, R.T.; Ziora, Z.M. Tyrosinase inhibitors as potential antibacterial agents. Eur. J. Med. Chem. 2020, 187, 111892. [Google Scholar]
- Muddathir, A.M.; Yamauchi, K.; Batubara, I.; Mohieldin, E.A.M.; Mitsunaga, T. Anti-tyrosinase, total phenolic content and antioxidant activity of selected Sudanese medicinal plants. S. Afr. J. Bot. 2017, 109, 9–15. [Google Scholar] [CrossRef]
- Kaewnarin, K.; Suwannarach, N.; Kumla, J.; Lumyong, S. Phenolic profile of various wild edible mushroom extracts from Thailand and their antioxidant properties, anti-tyrosinase and hyperglycaemic inhibitory activities. J. Funct. Foods 2016, 27, 352–364. [Google Scholar] [CrossRef]
- Azizuddin, S.; Khan, A.M.; Choudhary, M.I. Tyrosinase inhibitory potential of natural products isolated from various medicinal plants. Nat. Prod. Res. 2011, 25, 750–753. [Google Scholar] [CrossRef]
- Chen, Q.-X.; Song, K.-K.; Qiu, L.; Liu, X.-D.; Huang, H.; Guo, H.-Y. Inhibitory effects on mushroom tyrosinase by p-alkoxybenzoic acids. Food Chem. 2005, 91, 269–274. [Google Scholar] [CrossRef]
- Boo, Y.C. p-Coumaric Acid as An Active Ingredient in Cosmetics: A Review Focusing on its Antimelanogenic Effects. Antioxidants 2019, 8, 275. [Google Scholar] [CrossRef] [Green Version]
- Zengin, G.; Karanfil, A.; Uren, M.C.; Kocak, M.S.; Sarikurkcu, C.; Gungor, H.; Picot, C.M.N.; Mahomoodally, M.F. Phenolic content, antioxidant and enzyme inhibitory capacity of two Trametes species. RSC Adv. 2016, 6, 73351–73357. [Google Scholar] [CrossRef]
- Lo, H.C.; Wasser, S.P. Medicinal mushrooms for glycemic control in diabetes mellitus: History, current status, future perspectives, and unsolved problems (review). Int. J. Med. Mushrooms 2011, 13, 401–426. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-H.; Lai, M.-N.; Ng, L.-T. Inhibitory effects of medicinal mushrooms on α-amylase and α-glucosidase–enzymes related to hyperglycemia. Food Funct. 2013, 4, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Uskoković, A.; Jovanović, J.A.; Dinić, S.; Vidaković, M.; Mihailović, M.; Poznanović, G.; Grdović, N. Chapter 13—Mushroom and plant extracts as potential intervention supplements in diabetes management. In Biodiversity and Biomedicine; Ozturk, M., Egamberdieva, D., Pešić, M., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 247–256. [Google Scholar] [CrossRef]
- Gargano, M.L.; van Griensven, L.J.L.D.; Isikhuemhen, O.S.; Lindequist, U.; Venturella, G.; Wasser, S.P.; Zervakis, G.I. Medicinal mushrooms: Valuable biological resources of high exploitation potential. Plant Biosyst. 2017, 151, 548–565. [Google Scholar] [CrossRef]
- Lindequist, U.; Niedermeyer, T.H.; Julich, W.D. The pharmacological potential of mushrooms. Evid. Based Complement. Altern. Med. 2005, 2, 285–299. [Google Scholar] [CrossRef] [Green Version]
- Gruter, A.; Friederich, U.; Wurgler, F.E. Antimutagenic Effects of Mushrooms. Mutat. Res. 1990, 231, 243–249. [Google Scholar] [CrossRef]
- Ham, S.S.; Kim, S.H.; Moon, S.Y.; Chung, M.J.; Cui, C.B.; Han, E.K.; Chung, C.K.; Choe, M. Antimutagenic effects of subfractions of Chaga mushroom (Inonotus obliquus) extract. Mutat. Res. Gen. Toxicol. Environ. Mutagen. 2009, 672, 55–59. [Google Scholar] [CrossRef]
- Sugui, M.M.; de Lima, P.L.A.; Delmanto, R.D.; da Eira, A.F.; Salvadori, D.M.F.; Ribeiro, L.R. Antimutagenic effect of Lentinula edodes (BERK) Pegler mushroom and possible variation among lineages. Food Chem. Toxicol. 2003, 41, 555–560. [Google Scholar] [CrossRef]
- Shon, Y.-H.; Kim, S.-Y.; Lee, J.-S.; Lim, J.-K.; Nam, K.-S. Antimutagenic effect of polysaccharides extracted from soybeans fermented with basidiomycetes on 2-amino-3, 8-dimethylimidazo [4, 5-f] quinoxaline (MeIQx). J. Microbiol. Biotechnol. 2001, 11, 346–349. [Google Scholar]
- Laovachirasuwan, P.; Judprakob, C.; Sinaphet, B.; Phadungkit, M. In vitro antioxidant and antimutagenic activities of different solvent extracts of Phellinus spp. Int. Food Res. J. 2016, 23, 2608–2615. [Google Scholar]
- Morales, P.; Bermudez, E.; Sanz, B.; Hernandez, P.E. A Study of the Mutagenicity of Some Commercially Canned Spanish Mushrooms. Food Chem. Toxicol. 1990, 28, 607–611. [Google Scholar] [CrossRef]
- Mlinarič, A.; Kac, J.; Fatur, T.; Filipič, M. Anti-genotoxic activity of the mushroom Lactarius vellereus extract in bacteria and in mammalian cells in vitro. Die Pharm. Int. J. Pharm. Sci. 2004, 59, 217–221. [Google Scholar]
- Zhao, Y.Y.; Shen, X.F.; Chao, X.; Ho, C.C.; Cheng, X.L.; Zhang, Y.M.; Lin, R.C.; Du, K.J.; Luo, W.J.; Chen, J.Y.; et al. Ergosta-4,6,8(14),22-tetraen-3-one induces G2/M cell cycle arrest and apoptosis in human hepatocellular carcinoma HepG2 cells. BBA Gen. Subj. 2011, 1810, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Sun, J.; Wang, H.; Ng, T.B. First isolation and characterization of a novel lectin with potent antitumor activity from a Russula mushroom. Phytomedicine 2010, 17, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, H.F. The application of electronic computers to factor analysis. Educ. Psychol. Meas. 1960, 20, 141–151. [Google Scholar] [CrossRef]
- Sánchez, C. Reactive oxygen species and antioxidant properties from mushrooms. Synth. Syst. Biotechnol. 2017, 2, 13–22. [Google Scholar] [CrossRef]
- Liu, Y.-T.; Sun, J.; Luo, Z.-Y.; Rao, S.-Q.; Su, Y.-J.; Xu, R.-R.; Yang, Y.-J. Chemical composition of five wild edible mushrooms collected from Southwest China and their antihyperglycemic and antioxidant activity. Food Chem. Toxicol. 2012, 50, 1238–1244. [Google Scholar] [CrossRef]


{kind=link}
| Species | Habitat | Date | Edibility |
|---|---|---|---|
| Amanita crocea (Quél.) Singer | Pinus forest | June 2018 | Edible |
| Hemileccinum depilatum (Redeuilh) Šutara | Pinus forest | September 2018 | Edible |
| Cyclocybe cylindracea (DC.) Vizzini & Angelini | On Populus spp. stump | March 2018 | Edible |
| Lactarius deliciosus (L.) Gray | Pinus forest | June 2018 | Edible |
| Hygrocybe acutoconica (Clem.) Singer | Between meadows and grass | April 2018 | Inedible |
| Neoboletus erythropus (Pers.) C. Hahn | Pinus forest | October 2018 | Edible |
| Russula aurea Pers. | Pinus forest | June 2018 | Edible |
| Russula sanguinea Fr. | Pinus forest | May 2018 | Edible |
| Species | Total Phenolic Content (mg GAE/g) | DPPH (mg TE/g) | ABTS (mg TE/g) | CUPRAC (mg TE/g) | FRAP (mg TE/g) | Metal Chelating (mg EDTAE/g) | Phosphomolybdenum (mmol TE/g) |
|---|---|---|---|---|---|---|---|
| Amanita crocea | 9.33 ± 0.06 e | 26.09 ± 0.17 c | 36.59 ± 2.53 d | 39.57 ± 1.19 d,e | 37.40 ± 0.78 c | 15.06 ± 0.02 a | 0.35 ± 0.05 d |
| Hemileccinum depilatum | 20.10 ± 0.46 a | 41.89 ± 1.05 a | 84.33 ± 0.72 a | 116.44 ± 7.30 a | 86.23 ± 3.22 a | 13.85 ± 0.52 a,b,c | 0.47 ± 0.02 c |
| Cyclocybe cylindracea | 10.53 ± 0.13 d | 20.53 ± 0.21 d | 41.49 ± 0.34 c | 44.49 ± 0.15 d | 31.69 ± 0.12 d | 14.91 ± 0.04 a | 0.61 ± 0.03 a |
| Lactarius deliciosus | 7.42 ± 0.08 f | 5.66 ± 0.04 g | 17.53 ± 0.78 g | 31.90 ± 0.37 f | 16.65 ± 0.39 g | 14.82 ± 0.08 a | 0.46 ± 0.03 c |
| Hygrocybe acutoconica | 10.49 ± 0.22 d | 14.60 ± 1.12 e | 36.18 ± 1.59 d | 56.31 ± 0.06 c | 37.44 ± 1.69 c | 13.44 ± 0.81 b,c | 0.81 ± 0.04 a |
| Neoboletus erythropus | 16.52 ± 0.22 b | 30.36 ± 3.16 b | 70.96 ± 0.45 b | 103.87 ± 0.78 b | 65.76 ± 0.23 b | 12.85 ± 0.87 c | 0.41 ± 0.03 c,d |
| Russula aurea | 11.36 ± 0.19 c | 10.11 ± 0.41 f | 25.35 ± 0.15 f | 29.36 ± 0.05 f | 21.06 ± 0.68 f | 14.87 ± 0.39 a | 0.58 ± 0.04 b |
| Russula sanguinea | 10.89 ± 0.15 e,d | 9.28 ± 0.12 f | 30.07 ± 0.26 e | 34.40 ± 0.20 f | 27.12 ± 0.33 e | 14.75 ± 0.23 a,b | 0.66 ± 0.04 b |
| Compounds | Concentration of Compounds (µg/g) | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| p-Hydroxybenzoic acid | 56.10 | 14.31 | 105.73 | 63.32 | 22.77 | 12.64 | <9.77 | <9.77 |
| Cinnamic acid | 539.38 | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 |
| Protocatechuic acid | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 |
| 2,5-Dihydroxybenzoic acid | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 |
| umbelliferon | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 |
| p-Coumaric acid | 2.81 | 2.67 | 6.98 | <2.44 | 6.22 | <2.44 | <2.44 | <2.44 |
| o-Coumaric acid | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 |
| Vanillic acid | <156.3 | <156.3 | <156.3 | <156.3 | <156.3 | <156.3 | <156.3 | <156.3 |
| Gallic acid | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 |
| Esculetin | <19.5 | <19.5 | <19.5 | <19.5 | <19.5 | <19.5 | <19.5 | <19.5 |
| Caffeic acid | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 |
| Hinic acid | 36.40 | 19.4 | 26.81 | 14.17 | 14.60 | 8.50 | 32.48 | 23.32 |
| Scopolamine | <19.5 | <19.5 | <19.5 | <19.5 | <19.5 | <19.5 | <19.5 | <19.5 |
| Ferulic acid | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 |
| Sinapic acid | <78.1 | <78.1 | <78.1 | <78.1 | <78.1 | <78.1 | <78.1 | <78.1 |
| 3,4-Dimethoxycinnamic acid | <78.1 | <78.1 | <78.1 | <78.1 | <78.1 | <78.1 | <78.1 | <78.1 |
| Sinapic acid | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 |
| Daidzein | <19.5 | <19.5 | <19.5 | <19.5 | <19.5 | <19.5 | <19.5 | <19.5 |
| Genistein | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 |
| Apigenin | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 |
| Baicalin | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 |
| Naringenin | <1.22 | <1.22 | <1.22 | <1.22 | <1.22 | <1.22 | <1.22 | <1.22 |
| Luteolin | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 |
| kaempferol | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 |
| Catechin | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 |
| Epicatechin | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 |
| Chrysoeriol | <0.61 | <0.61 | <0.61 | <0.61 | <0.61 | <0.61 | <0.61 | <0.61 |
| Quercetin | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 |
| Isorhamnetin | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 |
| Myricetin | <625 | <625 | <625 | <625 | <625 | <625 | <625 | <625 |
| Chlorogenic acid | <1.22 | <1.22 | 6.81 | 5.49 | <1.22 | <1.22 | <1.22 | <1.22 |
| Matairesinol | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | 7.68 | <4.88 | <4.88 |
| Secoisolariciresinol | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 | <9.77 |
| Vitexin | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 |
| Apigenin-7-O-glc | <1.22 | <1.22 | <1.22 | <1.22 | <1.22 | <1.22 | <1.22 | <1.22 |
| Baicalin | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 | <39.1 |
| Luteolin-7-O-glc | <1.22 | <1.22 | <1.22 | <1.22 | <1.22 | <1.22 | <1.22 | <1.22 |
| Quercitrin | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 |
| Kaempferol 3-O-glucoside | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 |
| Epigallocatechin gallate | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 | <312.5 |
| Hyperoside | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 |
| Quercetin 3-O-glucoside | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 |
| Amentoflavone | <0.61 | <0.61 | <0.61 | <0.61 | <0.61 | 1.02 | <0.61 | <0.61 |
| Apiin | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 | <2.44 |
| Rutin | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 | <4.88 |
| Species | AChE (mg GALAE/g) | BChE (mg GALAE/g) | Tyrosinase (mg KAE/g) | Amylase (mmol ACAE/g) | Glucosidase (mmol ACAE/g) |
|---|---|---|---|---|---|
| Amanita crocea | 0.90 ± 0.01 b | 0.80 ± 0.02 b | 49.79 ± 0.15 d,e | 0.08 ± 0.01 f,g | NA |
| Hemileccinum depilatum | NA | NA | 54.18 ± 0.17 a | 0.07 ± 0.01 g | 0.17 ± 0.07 c |
| Cyclocybe cylindracea | 1.02 ± 0.02 a | 0.99 ± 0.07 a | 53.24 ± 0.12 a,b | 0.09 ± 0.01 e,f | 0.34 ± 0.01 b |
| Lactarius deliciosus | 0.90 ± 0.01 b | 0.98 ± 0.05 a | 50.47 ± 0.46 c,d | 0.10 ± 0.01 c,d | 0.36 ± 0.01 b |
| Hygrocybe acutoconica | 0.60 ± 0.06 d | NA | 50.97 ± 0.45 c | 0.17 ± 0.01 a | 1.86 ± 0.01 a |
| Neoboletus erythropus | 0.95 ± 0.04 b | NA | 52.64 ± 0.16 b | 0.09 ± 0.01 d,e | 0.22 ± 0.04 c |
| Russula aurea | 0.93 ± 0.01 b | NA | 49.66 ± 0.59 d,e | 0.12 ± 0.01 b | 0.36 ± 0.01 b |
| Russula sanguinea | 0.78 ± 0.03 c | NA | 48.83 ± 0.30 e | 0.11 ± 0.01 b,c | 0.36 ± 0.01 b |
| Concentration µg/plate | TA 98 | TA 100 | |||
|---|---|---|---|---|---|
| S9 (−) | S9 (+) | S9 (−) | S9 (+) | ||
| Positive control | 463 ± 42 | 2845 ± 109 | 1048 ± 131 | 4764 ± 129 | |
| Negative control | 100 µL | 32 ± 2 | 40 ± 3 | 139 ± 10 | 145 ± 6 |
| Bacteria control | 0 | 32 ± 1 | 36 ± 2 | 140 ± 15 | 151 ± 18 |
| Amanita crocea | 10,000 | 21 ± 1 | 29 ± 1 | 103 ± 3 | 138 ± 13 |
| 5000 | 23 ± 4 | 34 ± 4 | 123 ± 6 | 147 ± 16 | |
| 2500 | 29 ± 4 | 36 ± 2 | 148 ± 4 | 152 ± 8 | |
| Hemileccinum depilatum | 10,000 | 41 ± 3 | 41 ± 4 | 152 ± 13 | 163 ± 12 |
| 5000 | 33 ± 3 | 40 ± 3 | 162 ± 11 | 170 ± 11 | |
| 2500 | 34 ± 1 | 32 ± 0 | 171 ± 7 | 159 ± 7 | |
| Cyclocybe cylindracea | 10,000 | 37 ± 1 | 30 ± 2 | 172 ± 4 | 144 ± 8 |
| 5000 | 31 ± 4 | 37 ± 2 | 139 ± 10 | 142 ± 10 | |
| 2500 | 36 ± 0 | 34 ± 3 | 151 ± 1 | 138 ± 4 | |
| Lactarius deliciosus | 10,000 | 29 ± 1 | 40 ± 0 | 146 ± 4 | 155 ± 8 |
| 5000 | 27 ± 1 | 28 ± 1 | 154 ± 12 | 146 ± 2 | |
| 2500 | 27 ± 3 | 37 ± 2 | 134 ± 12 | 138 ± 7 | |
| Hygrocybe acutoconica | 10,000 | 43 ± 4 | 36 ± 3 | 147 ± 11 | 161 ± 14 |
| 5000 | 30 ± 1 | 36 ± 7 | 130 ± 7 | 135 ± 11 | |
| 2500 | 31 ± 4 | 33 ± 1 | 151 ± 7 | 148 ± 9 | |
| Neoboletus erythropus | 10,000 | 39 ± 3 | 30 ± 4 | 145 ± 8 | 129 ± 6 |
| 5000 | 34 ± 5 | 43 ± 3 | 152 ± 13 | 135 ± 18 | |
| 2500 | 32 ± 5 | 41 ± 3 | 137 ± 1 | 144 ± 12 | |
| Russula aurea | 10,000 | 28 ± 4 | 40 ± 5 | 135 ± 4 | 164 ± 9 |
| 5000 | 31 ± 0 | 39 ± 1 | 145 ± 1 | 149 ± 14 | |
| 2500 | 34 ± 3 | 38 ± 2 | 132 ± 8 | 119 ± 12 | |
| Russula sanguinea | 10,000 | 38 ± 4 | 37 ± 1 | 140 ± 3 | 128 ± 19 |
| 5000 | 26 ± 2 | 29 ± 1 | 160 ± 5 | 162 ± 5 | |
| 2500 | 33 ± 3 | 31 ± 3 | 130 ± 6 | 143 ± 8 | |
| Concentration (µg/plate) | TA 98 | TA 100 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| S9 (-) | % Inhibition | S9 (+) | % Inhibition | S9 (-) | % Inhibition | S9 (+) | % Inhibition | ||
| Negative Control | 100 µL/plate | 29 ± 2 | 41 ± 3 | 158 ± 8 | 170 ± 3 | ||||
| Positive Control | 667 ± 19 | 0 | 2796 ± 139 | 0 | 1730 ± 104 | 0 | 3631 ± 139 | 0 | |
| Bacteria Control | 0 | 31 ± 4 | 41 ± 4 | 157 ± 17 | 175 ± 9 | ||||
| Amanita crocea | 10,000 | 569 ± 17 | 15 | 203 ± 16 | 94 | 1633 ± 34 | 6 | 773 ± 26 | 83 |
| 5000 | 543 ± 20 | 19 | 1280 ± 25 | 55 | 1678 ± 28 | 3 | 3359 ± 49 | 8 | |
| 2500 | 524 ± 16 | 22 | 2457 ± 112 | 12 | 1407 ± 24 | 21 | 3519 ± 183 | 3 | |
| Hemileccinum depilatum | 10,000 | 588 ± 32 | 12 | 237 ± 22 | 93 | 1225 ± 4 | 32 | 880 ± 46 | 80 |
| 5000 | 553 ± 22 | 18 | 1679 ± 36 | 41 | 1233 ± 15 | 32 | 3382 ± 52 | 7 | |
| 2500 | 514 ± 25 | 24 | 2701 ± 126 | 3 | 1342 ± 36 | 25 | 3491 ± 103 | 4 | |
| Cyclocybe cylindracea | 10,000 | 551 ± 25 | 18 | 254 ± 2 | 92 | 1185 ± 53 | 35 | 739 ± 31 | 84 |
| 5000 | 560 ± 37 | 17 | 1697 ± 105 | 40 | 1241 ± 7 | 31 | 2904 ± 71 | 21 | |
| 2500 | 588 ± 42 | 12 | 2619 ± 39 | 6 | 1313 ± 34 | 27 | 3575 ± 151 | 2 | |
| Lactarius deliciosus | 10,000 | 535 ± 11 | 21 | 260 ± 21 | 92 | 1166 ± 30 | 36 | 1617 ± 45 | 58 |
| 5000 | 519 ± 10 | 23 | 1310 ± 118 | 54 | 1156 ± 57 | 36 | 3361 ± 61 | 8 | |
| 2500 | 580 ± 19 | 14 | 2760 ± 125 | 1 | 1281 ± 50 | 29 | 3312 ± 112 | 9 | |
| Hygrocybe acutoconica | 10,000 | 379 ± 35 | 45 | 156 ± 19 | 96 | 1127 ± 14 | 38 | 279 ± 5 | 97 |
| 5000 | 527 ± 16 | 22 | 242 ± 10 | 93 | 1188 ± 21 | 34 | 630 ± 54 | 87 | |
| 2500 | 565 ± 22 | 16 | 790 ± 46 | 73 | 1171 ± 4 | 36 | 1308 ± 102 | 67 | |
| Neoboletus erythropus | 10,000 | 513 ± 20 | 24 | 676 ± 46 | 77 | 1186 ± 33 | 35 | 1900 ± 75 | 50 |
| 5000 | 544 ± 25 | 19 | 2196 ± 102 | 22 | 1246 ± 14 | 31 | 3322 ± 114 | 9 | |
| 2500 | 571 ± 5 | 15 | 2594 ± 129 | 7 | 1292 ± 39 | 28 | 3527 ± 78 | 3 | |
| Russula aurea | 10,000 | 449 ± 13 | 34 | 262 ± 16 | 92 | 1189 ± 51 | 34 | 694 ± 11 | 85 |
| 5000 | 442 ± 6 | 35 | 847 ± 23 | 71 | 1171 ± 24 | 36 | 2306 ± 27 | 38 | |
| 2500 | 573 ± 11 | 15 | 2449 ± 131 | 13 | 1446 ± 49 | 18 | 2769 ± 46 | 25 | |
| Russula sanguinea | 10,000 | 489 ± 18 | 28 | 213 ± 23 | 94 | 1138 ± 33 | 38 | 705 ± 38 | 85 |
| 5000 | 461 ± 22 | 32 | 637 ± 26 | 78 | 1341 ± 16 | 26 | 2840 ± 44 | 23 | |
| 2500 | 591 ± 5 | 12 | 2025 ± 36 | 28 | 1287 ± 41 | 28 | 3386 ± 151 | 7 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkan, S.; Uysal, A.; Kasik, G.; Vlaisavljevic, S.; Berežni, S.; Zengin, G. Chemical Characterization, Antioxidant, Enzyme Inhibition and Antimutagenic Properties of Eight Mushroom Species: A Comparative Study. J. Fungi 2020, 6, 166. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030166
Alkan S, Uysal A, Kasik G, Vlaisavljevic S, Berežni S, Zengin G. Chemical Characterization, Antioxidant, Enzyme Inhibition and Antimutagenic Properties of Eight Mushroom Species: A Comparative Study. Journal of Fungi. 2020; 6(3):166. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030166
Chicago/Turabian StyleAlkan, Sinan, Ahmet Uysal, Giyasettin Kasik, Sanja Vlaisavljevic, Sanja Berežni, and Gokhan Zengin. 2020. "Chemical Characterization, Antioxidant, Enzyme Inhibition and Antimutagenic Properties of Eight Mushroom Species: A Comparative Study" Journal of Fungi 6, no. 3: 166. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030166