Phytophthora palmivora–Cocoa Interaction

1
School of Life and Environmental Sciences, The University of Sydney, LEES Building (F22), Camperdown, NSW 2006, Australia
2
The University of Sydney Institute of Agriculture, 1 Central Avenue, Australian Technology Park, Eveleigh, NSW 2015, Australia
J. Fungi 2020, 6(3), 167; https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030167
Submission received: 2 June 2020
/
Revised: 28 August 2020
/
Accepted: 7 September 2020
/
Published: 9 September 2020
(This article belongs to the Special Issue Plant Fungal Pathogenesis)
Abstract
:Phytophthora palmivora (Butler) is an hemibiotrophic oomycete capable of infecting over 200 plant species including one of the most economically important crops, Theobroma cacao L. commonly known as cocoa. It infects many parts of the cocoa plant including the pods, causing black pod rot disease. This review will focus on P. palmivora’s ability to infect a plant host to cause disease. We highlight some current findings in other Phytophthora sp. plant model systems demonstrating how the germ tube, the appressorium and the haustorium enable the plant pathogen to penetrate a plant cell and how they contribute to the disease development in planta. This review explores the molecular exchange between the oomycete and the plant host, and the role of plant immunity during the development of such structures, to understand the infection of cocoa pods by P. palmivora isolates from Papua New Guinea.
1. Introduction
Within the order Peronosporales, the largest genus with over 120 described species, Phytophthora is a hemibiotrophic phytogen capable of infecting a wide range of hosts, including many agricultural crops, worldwide [1,2]. One of the most economically important and delicious crops affected is cocoa (Theobroma cacao L.). At a global scale, black pod or pod rot is the most important cocoa disease, caused by several Phytophthora species (Table 1) and contributing to significant pod losses of up to 30% and killing up to 10% of trees annually [1,3,4]. Some black pod-causing Phytophthora species have distinct geographical distributions (Table 1) while Phytophthora palmivora (Ppal, Butler) [5], which was originally isolated from Palmyra palm (Borassus flabellifer) in 1907, has a pantropical geographical distribution and is found in virtually all cocoa production areas [1,4,6]. In addition, it has a wide host range of over 200 plant species in the tropics [1,4,6]. This has serious implications for smallholder farmers who produce over 80% of all cocoa, as cocoa trees are mainly grown under shade trees, either in an inter-cropped or in semi-natural agro-forestry systems [3].
2. Morphology of P. palmivora (Ppal)
Phytophthora, as an oomycete, is part of a distinct group of fungus-like eukaryotic microbes. It shares a range of morphological features with fungi, but it possesses other features unique to plants, such as the major component of its cell wall being cellulose, unlike true fungi, which consists mainly of chitin [11]. Another feature is that its mycelium is composed of hyaline, branched, non-septate filaments, while fungal hyphae have septate.
The dispersal of Phytophthora by wind or water is achieved by asexual sporangia (Figure 1A), which develop at the ends of specialized hyphal tips [12]. Sporangia morphology can be quite diverse but the shapes of Ppal sporangia range from ovoid-ellipsoid to obpyriform, and they are papillate and cadacous, i.e., short pedicels [1,12,13]. Sporangia can germinate directly forming germ tubes and hyphae, or they release motile asexual spores called zoospores (Figure 1D–G). Anodotactic Ppal zoospores actively swim with the aid of two flagella on the wet surface of plant tissues or in flooded soil by negative geotaxis [14], by electrotaxis in natural root-generated electric fields [15] and by chemotaxis [16,17,18]. In addition, high humidity/moisture and splashes of water help in the spread of such zoospores from plant to plant. Therefore, the reduction of high humidity and avoidance of excess water are some of the practices in greenhouses/glasshouses for Phytophthora disease control [19].
Ppal reproduces both asexually, via the motile zoospores, and sexually, via the formation of oospores caused by the contact of two structures found at the mycelium tips: the female oogonium (the sac which contains the developing oospore) and the male structure, the antheridium. Phytophthora species can be described as homothallic (self-fertile) or heterothallic (self-sterile) where the latter requires the mating of compatible A1 and A2 types. Ppal is heterothallic and oospores can be produced only when A1 and A2 types are grown together on agar plates or on infected plants. Interestingly, in Ppal, the A2 compatibility type is predominant on cocoa throughout the world [20,21].
Chlamydospores are usually globose and can be intercalarily or terminally located on the mycelium. They can be distinguished from hyphal swellings due to the presence of septate (Figure 1B–D). They are recognized as resistant, long-term survival structures [22]. It was shown that Ppal storage cultures can remain viable in water at room temperature for up to 23 years and that Ppal colonies developed from chlamydospore-like structures that were produced in the absence of adequate nutrition and aeration [23].
3. P. palmivora (Ppal)’s Infection Process in Cocoa
Ppal belongs to Clade 4, whose species form papillate sporangia and are known to be pathogenic to plant roots [7], causing root rot disease in many plants [24]. It can infect other plants tissues such as the stems, leaves and fruits of many economically important tropical plants such as breadfruit (Artocarpus altilis), coconut (Cocos nucifera) and durian (Durio zibethinus) [1,23], including both monocots and dicots. Infection studies have been done in model plants such as Medicago truncatula [25,26], Nicotiana benthamiana [27,28] and in the model liverwort, Marchantia polymorpha [29], as well as on coconut [30], oil palm [31,32,33], betelvine [34], citrus hosts [35,36], rubber [37], and papaya [38].
In cocoa, Purwantara [39] demonstrated that soil from cocoa plantations in West Java was a massive and consistent source of Ppal inocula and that Ppal infection from soil to the cocoa pods appears to be mainly through contact or rain splash. Caducous sporangia or motile zoospores adhere to the plant surface. Though a single sporangium could germinate to start the infection cycle within cocoa, our focus will be on the infectious agent, a single motile zoospore. The zoospore adheres to the plant surface where it sheds its flagella and forms a non-motile spherical cyst (Figure 1H–I and Figure 2B).
Encystment and cyst germination are two important developmental stages required for Ppal to adhere or to dock [40,41] on the surface of cocoa plant tissues. Studies in various oomycetes have demonstrated that the zoospores docked precisely on the root surface at its ventral face with the help of the posterior flagellum, allowing the deposition of adhesive contents during encystment on the plant host and orientating Phytophthora to germinate toward the host [42,43,44]. Transient leaching treatments of encysting zoospores, which involved leaching solutions at various time intervals underneath polycarbonate membranes, calcium, pectin and various other molecules as well as mechanical agitation, affect the ability of Ppal to dock and to form germ tubes [45,46,47]. Further work by Zhang and colleagues [48] found that methylation destroyed the capacity of the pectin to induce germination, but its methylated form induced zoospore rounding and partial encystment at low concentrations. This is important as the outer surface of most aerial organs of plants such as leaves, flowers, fruits and non-woody stems are covered with the cuticle, which consists of cellulose, hemicelluloses and pectins [49]. Using functional and structural analyses, pectin methylesterase-coding genes have been found in various Phytophthora species [50,51,52] as well as polygalacturonase and pectate lyase in P. capsici [53], capable of degrading pectin.
Bimpong and Clerk [16] demonstrated that Ppal zoospores responded chemotactically to an extract of cocoa pod where the cyst germ tubes grew towards the stimulus but not to the exudate. The germ tube grows on the plant surface (Figure 2C) and various environmental cues induce the formation of an appressorium for the subsequent entry into the plant host cell [54]. Studies by Ali et al. [55] and Tey [56] demonstrated that Ppal cysts formed germ tubes and appressoria on cocoa pod husks and leaf tissues. Entry via wounding and stomatal pores has been observed by Ppal in cocoa and in other Phytophthora–plant infection studies by microscopy. Studies in chickpea showed that P. megasperma f. sp. medicaginis vacuolated zoospore cysts formed germ tubes to gain entry into stomatal pore and after septum formation, differentiated into the primary hypha within the hypocotyl region [57]. Widmer et al. [36] found that P. palmivora gained entry in a natural wound site on a root of tolerant trifoliate orange (Poncirus trifoliata) via a germ tube.
There are differences in the structure and organization of various plant tissues; for example, the presence of the cuticle on the aerial parts of plants [49] i.e., the leaf, the non-woody stem and pod, but not in the roots. Figure 2 is a schematic diagram focusing on the general infection process of Phytophthora in plant tissues (Figure 2D–F). The appressorium forms a penetration peg to penetrate the cuticle layer or the cell wall of an epidermal cell [58,59,60]. In ground tissues, germ tubes that emerged from cysts penetrated the root epidermis, usually by intercellular growth along the anticlinal cell walls [61] or by appressorium-mediated penetration via a penetration peg between two rhizodermis cells by P. parasitica [62]. Intracellular penetration can occur and germ tubes from encysted zoospores can become swollen and produce a penetration peg [57]. Then, specialised hyphae invade plant cells to form haustoria (Figure 2D–E) [26,63,64,65,66,67]. Histological studies in Quercus ilex roots during P. cinnamomi infection found haustoria-like structures in the cortical root and phloem cells [63]. Haustoria have been observed in Medicago root epidermal cells [64]. The Ppal haustorium is a short, swollen, anucleate hyphal branch, which protrudes into the peripheral cytoplasm of the host cell [66,68]. The haustorium is surrounded by a specialized host-derived membrane, the extrahaustorial membrane (EHM), which is distinct from the plant plasma membrane. In fungi, haustoria function as feeding structures [69]. During this phase of growth, Ppal interaction with its host is biotrophic and secreted effectors and enzymes targeting the apoplastic and cytoplasmic sites in the plant host have been shown to play a role in plant cellular reprogramming/rearrangement and in reducing plant immunity [65,70,71,72,73,74,75,76,77,78,79,80]. Therefore, the development of the haustorium plays a critical role in the successful parasitic infection of Phytophthora.
Intercellular infection by Ppal can be observed in planta as well [33,81]. To complete its lifecycle, the hemibiotrophic Ppal switches from a biotrophic to a necrotrophic lifestyle highlighted by the presence of necrotic plant tissues, prolific hyphal growth and the formation of sporangia as well as chlamydospores in plant tissues (Figure 2F) [28,29,60,82].
4. Overcoming Plant Host Immunity by Ppal and Other Oomycetes
Phytophthora, along with other plant pathogens, needs to overcome the plant host’s immunity. In the first line of defense, the cocoa plant would use pattern-recognition receptors (PRRs) found on the plant cell membrane. These detect microbe- and pathogen-associated molecular pattern (MAMP and PAMP) molecules leading to pattern-triggered immunity (PTI). In addition, the cocoa plant needs a secondary line of defense, as Phytophthora can overcome PTI by secreting effectors that suppress PTI responses, resulting in effector-triggered susceptibility. These effectors can act within the apoplastic and symplastic regions of the plant cell, where a secretory system would enable the delivery of such effectors via the appressorium and the haustorium respectively [59,83]. Plants possess cytoplasmic resistance (R) proteins that recognize such effectors. These R proteins are intracellular receptor proteins of the nucleotide binding–leucine-rich repeat (NB-LRR) type [84,85,86], which are activated in the presence of key effectors to trigger a hypersensitive response (HR) and systemic acquired resistance (SAR) in the plant host [87,88,89]. This is termed effector-triggered immunity (ETI). It is pathogen strain- or race-specific and associated with programmed cell death [90]. Furthermore, Thomma et al. [90] proposed that PAMP receptors and R proteins are part of the plant’s surveillance mechanism and that both PTI and ETI are used for effective immunity.
Under a hypersensitive defense response, a rapid plant cell death occurs at the point of pathogen ingress and is generally associated with ETI. Recent work by Gu et al. [91] observed the upregulation of multiple plant NB-LRR genes in Mexican wild potato species, Solanum pinnatisectum against P. infestans, where hyphal expansion was significantly restricted in epidermal cells and mesophyll cell death was predominant at 12 hours post inoculation (hpi), thus indicating that the HR was induced upon infection. Under SAR, a localised response due to a pathogen induces resistance at sites remotely located from the initial infection, and this is associated with the transport of defense signals such as salicylic acid throughout the plant, resulting in broad-spectrum disease resistance against secondary infections. Recent work in potato has shown a link between microRNAs i.e., non-coding RNAs that act as negative regulators of gene expression, in SAR response [92]. The knockdown (KD) of miR160 compromised SAR response to P. infestans in miR160 KD lines of S. tuberosum cv. Désirée [92]. miRNAs also affected NB-LRR genes in tomatoes [93] and in soybean [94] during P. infestans and P. sojae infection respectively. In addition, it has been shown that P. sojae secreted effectors to suppress RNA silencing in plants by inhibiting the biogenesis of small RNAs [95], thus promoting infection. Recent evidence suggests that miRNAs repression of NB-LRR resistance genes in plants is not only used by plant pathogenic oomycetes such as Phytophthora, but could play a role in the infection of leguminous plants by symbiotic bacteria. miRNAs repressed NB-LRR resistance genes to promote Sinorhizobium meliloti’s colonization and the development of nitrogen-fixing nodules in Medicago truncatula [96]. Sós-Hegedűs et al. [96] proposed a model that a subset of NB-LRR-targeting plant miRNAs (miR482/2118 superfamily, miR1507, miR2109) could tip the balance in NB-LRR proteins in the M. truncatula, affecting the perception of S. meliloti as a pathogen or a symbiont [96].
Hardham and Blackman [97] and Wang and Jiao [98] highlighted PAMPs and effectors used in other characterized plant pathogenic Phytophthora species such as P. infestans, P. capsici, P. cinnamomic, and P. parasitica and some of the approaches used to understand their role in PTI and ETI. Furthermore, Raaymakers and Van den Ackerveken [99] listed several oomycete-derived patterns known to activate plant immunity. In the case of Ppal and cocoa interaction, Ppal success in establishing disease would rely on avoiding detection of PAMPs by PRRs or the secretion of effectors within the plant’s apoplast and symplast to interfere with PTI or ETI to support its infection and promote disease development. The following section of this review will focus on some specific Ppal-derived patterns such as lectins and Ppal RxLR effectors and their functions during infection in cocoa and in other model plants.
4.1. Necrosis and Ethylene-Inducing Peptide 1 (Nep1)-Like Proteins
Necrosis and ethylene-inducing peptide 1 (Nep1)-like proteins (NLPs), which were first identified in P. parasitica, have been shown to induce necrosis in planta [100,101]. Work by Schumacher et al. [102] identified NLPs in the obligate biotrophic oomycete Plasmopara viticola, which causes grapevine downy mildew. In addition, NLPs are secreted by bacteria and fungi and come in two forms, those that are cytotoxic to eudicot plants and those that are noncytotoxic [103]. Within 24 h of application of Nep1 purified from Fusarium oxysporum f. sp. erythroxyli culture filtrates (at 5 μg ml−1 plus 0.2% Silwet-L77), the majority of stomata guard cells and two or more neighboring epidermal cells around each affected stomata on the abaxial leaf surface in mature green cocoa leaves were killed, with the microscopic necrotic flecks and darkly pigmented necrotic lesions developed on Nep1-treated field-grown Amelonado cocoa pods (at the same concentration) [104]. It was suggested that lesion development in cocoa pods was due to Nep1 entry via the stomata on the cocoa pod surface [104]. In addition, the expression of cocoa genes involved in defense gene regulation, cell wall development and energy production were different in young red leaves and mature green leaves of cocoa in response to the application of Nep1 [104]. Bae et al. [105] demonstrated that six of the nine NEP1 orthologues, which had a similar sequence to the NEP1 of F. oxysporium, were expressed in P. megakarya mycelium and in P. megakarya zoospore-infected cocoa leaf tissue using leaf disc assays. Evangelisti et al. [28] identified 24 putative NLPs in the Ppal secretome study in N. benthamiana. Ali and colleagues [106] identified several NPP1-type necrosis inducing-like proteins and NPP1-like proteins, a necrosis-inducing protein NPP8 and a Suppressor of Necrosis 1 protein (SNE1) in Ppal–cocoa infection studies. The latter, SNE1, previously characterized in P. infestans, was shown to translocate to the plant nucleus and suppressed the action of secreted NLPs from Phytophthora that are expressed during the necrotrophic growth phase, as well as programmed cell death mediated by the Avr3a/R3a protein interaction [107].
4.2. Lectins and Cellulose-Binding Elicitor Lectins (CBELs)
Plant lectins play a signaling role to modulate plant immunity responses to various plant pathogens via lectin receptor kinases [108,109]. Previous work in potatoes demonstrated that lectins lysed P. infestans zoospores and mediated the binding of cell membranes of potato to cell wall surfaces of infecting hyphae of both compatible and incompatible races of P. infestans in vivo [110,111]. By expressing the Arabidopsis lectin receptor kinase LecRK-I.9 gene in potato and N. benthamiana, late blight resistance to P. infestans was significantly enhanced [112]. Cellulose-binding elicitor lectins (CBELs) are cell wall-localized glycoproteins involved in cell wall organization and the adhesion of the mycelium to cellulosic substrates [113,114,115]. They have also been shown to aid in Phytophthora’s penetration into its plant host by mediating the oomycete’s attachment to the host surface [113,114,115]. According to Khatib and colleagues [116], this glycoprotein is widespread in the genus Phytophthora. Secretome work on Ppal identified 24 lectins including one CBEL [28]. Infection work in Ppal on cocoa found eight CBELs and a putative CBEL-like protein transcribed in the Ppal mycelia, zoospores and in planta [106]. Work by Laroque and colleagues [117] found that CBEL played a role in triggering immunity in the P. parasitica–Arabidopsis interaction, showing that BRASSINOSTEROID INSENSITIVE 1-associated kinase 1 (BAK1) and NADPH oxidase genes were required for CBEL-induced oxidative burst and defense responses but not for necrosis.
4.3. Elicitins
Lack of extracellular 10-kDa elicitins have been correlated with virulence in most P. parasitica isolates of tobacco [118]. Work by Huitema et al. [119] identified two classes of elicitins that are secreted such as INF1 (class I) and the cell-surface-anchored polypeptides, INF2A and INF2B (Class III) in P. infestans. Coexpression of INF1 and the NLP protein PiNPP1.1 from P. infestans led to synergistic enhancement of cell-death elicitation in N. benthamiana [120]. Work by Le Fevre and colleagues [60] demonstrated that PAL1, the Ppal homolog of P. infestans inf1, was transcriptionally induced in barley roots and leaves during Ppal infection. Ppal produces a 10-kDa protein, palmivorein [121,122] and a 75 kDa elicitor, which triggered defense responses in rubber plants [122]. In another study, a crude elicitor from culture filtrates of Ppal was applied to rubber tree leaves and this pretreatment significantly increased Ppal infection in such leaves [123]. In addition, infiltration of this crude elicitor promoted cell death and increased salicylic acid (SA), abscisic acid (ABA) and the phytoalexin, scopoletin (Scp) content in tobacco and rubber tree leaves [123]. Recent work by Pettongkhao and colleagues [124] isolated a secreted glycoprotein of 15 kDa from a papaya Ppal isolate and suggested that Ppal15kDa played an important role in normal development of Ppal infection structures. All Ppal15kDa mutants generated via CRISPR/Cas9-mediated gene editing, were compromised in infectivity on N. benthamiana and papaya [124]. In addition, the mutants’ development was also affected as they produced smaller sporangia, shorter germ tubes, and fewer appressoria, leading to reduced levels of pathogenicity [124].
4.4. Glycoside Hydrolase 12 Proteins
Ma et al. [125] showed that the P. sojae glycoside hydrolase 12 protein, PsXEG1, acted as a PAMP in soybean (Glycine max) and solanaceous species and, by both silencing and overexpression of XEG1 in P. sojae, severely reduced virulence. Later, Ma et al. [126] demonstrated that P. sojae secreted a paralogous PsXEG1-like protein, PsXLP1, that had lost enzyme activity. The latter could bind to a soybean apoplastic glucanase inhibitor protein, GmGIP1, more tightly than did PsXEG1, thus freeing PsXEG1 to assist P. sojae infection [126]. P. parasitica orthologs PpXEG1 and PpXLP1 were found to have similar functions and both genes were found to be conserved in other Phytophthora species [126]. Use of the Carbohydrate-Active EnZymes (CAZy) database enabled Zerillo et al. [127] to identify xyloglucan-β-1, 4-D-endoglucanase genes in family GH12 in Pythium sp. and various oomycetes. Evangelisti et al. [28] identified putative glycosyl hydrolases in Ppal in N. benthamiana infection studies. Only nine belonged to GH12 family, where PLTG_13824/PEX_0219 was described as a cell 12A endoglucanase and the remaining eight as hypothetical proteins [28]. However, work by Ali et al. [106] identified two candidate genes in Ppal and three candidate genes in P. megakyara belonging to the glycoside hydrolase 12 family.
However, recent work by Ochola and colleagues [128] may provide some clues as to how Avr gene expression impacts the compatibility of plant disease. By using the CRISPR/Cas9 engineering technique, PsAvr3b promoter sequences from P. sojae were substituted in situ with promoter sequences from Actin (constitutive expression), PsXEG1 (early expression), and PsNLP1 (later expression). Compared to the wild type and the unedited mutant (T1) i.e., with the native PsAvr3b promoter as controls, PsAvr3b expression was significantly reduced when the PsAvr3b promoter was substituted with PsXEG1 (early expression) or PsNLP1 (late expression) promoters [128]. When these promoter mutants carrying PsXEG1 (X02 and X03) or PsNLP1 (N02 and N10) were tested on Williams (susceptible) and two resistant (Rps3b and Rps3c) soybean cultivars, these mutants gained virulence against the resistant Rps3b cultivar while mutants containing the PsACT promoter (A24 and A26) were unable to infect soybean cultivars carrying Rps3b [128]. No infection was observed with the WT and T1 control on soybean cultivars carrying Rps3b [128]. Further transcriptomic studies with these promoter mutants highlighted a difference in gene expression in the resistant Rps3b cultivar such as the wound-inducible, jasmonate synthesis-degradation lipoxygenase (LOX-1) and the proline extensin-like receptor kinase 1 (PERK1) [128]. Compared to the WT strain, LOX-1 and PERK1 were upregulated in soybean cultivars (carrying Rps3b) infected with the mutant with the PsACT promoter (A24) expressed, while they were downregulated in those infected with the promoter mutants, PsXEG1 (X03) and PsNLP1 (N02), respectively, at 24 hpi [128].
4.5. Transglutaminases (Pep-13)
The calcium-dependent cell wall transglutaminase (TGase), GP42 from P. sojae, consists of a peptide fragment/domain (Pep-13), which activates plant defense in parsley and potato [129]. GP42 belongs to a group of enzymes that catalyzes the post-translational modification of proteins by the formation of isopeptide bonds [130]. In a proteome study of P. infestans membrane, two transglutaminases were encoded by PITG_22117 and PITG_16956, respectively [131]. PITG_22117 was detected in both non-sporulating mycelium and germinating cysts with appressoria while PITG_16956 was identified from sporulating mycelium [131]. Potato plants treated with Pep-13 not only were able to mount a salicylic acid (SA)- and jasmonic acid (JA)-dependent defense response, but were also found to activate the co-receptor BAK1 [132]. Recent work by Wang et al. [133] demonstrated that the Phytophthora MAMP Pep-13 triggered SOMATIC EMBROYOGENESIS KINASE 3 (SERK3)/BAK1-independent PTI. In wild potato (Solanum microdontum), a receptor-like protein ELR (elicitin response) mediated extracellular recognition of the elicitin domain, a domain known to be conserved in Phytophthora species. ELR also was associated with the immune co-receptor BAK1/SERK3 and the transfer of ELR into cultivated potato resulted in enhanced resistance to P. infestans [134]. Previous work by Brunner et al. [129] found a GP42-like protein containing the Pep-13 motif in Ppal and in the Ppal secretome study; Evangelisti et al. [28] identified five out of six transglutaminases carrying the conserved Pep-13 motif.
4.6. RxLR and CRN Effectors
Many effectors are known to act in the apoplastic and symplastic region of plant cells during the appressorium and the haustorium development [59,83]. Two classes of effectors are known: RxLR where N terminus of such effectors have a conserved arginine-any amino acid-leucine-arginine motifs usually linked with a glutamic acid-glutamic acid-arginine domain (RxLR-dEER). The other class is CRinkling- and Necrosis-inducing proteins (CRNs), which contain a LFLAK motif. These are involved in manipulating many functions linked to the host immunity such as cell protease function, phytohormone signaling and RNA silencing effectors [97,98]. Secretome studies in Ppal identified putative secreted proteins such as RXLR effectors [28]. Transcriptomic work found four RxLR effectors (REX1-4) to be upregulated during Ppal infection in N. benthamiana roots, and REX2 and REX3 effectors were found to suppress host secretion [28]. In P. parasitica, the Penetration-Specific Effector 1 (PSE1) protein is a secreted RxLR effector protein whose expression is induced during appressorium-mediated penetration of the host roots, but declines during early biotrophy and cannot be detected during the necrotrophic phase of infection [59,135]. PSE1 abolished cell death in tobacco plants triggered by the P. cryptogea elicitin cryptogein and the Pseudomonas syringae AvrPto avirulence protein and increased susceptibility of A. thaliana to P. parasitica by altering the distribution of key auxin efflux transporters in PSE1 transgenic A. thaliana lines [135]. Genome, transcriptome and secretome studies combined with RNA-sequencing and RT-PCR identified RXLR effectors and crinklers in Ppal and P. megakarya, which were differentially expressed in mycelia, zoospores, and in planta (infected pod husks) [106]. Furthermore, recent work by Morales–Cruz et al. [136] predicted that Ppal had 717 RxLR effectors compared to P. megakarya, which had 1,382 effectors due to genome duplication and expansion in the latter. In addition, 251 “putative effectors” in Ppal had shared homology and often bordered RxLRs [136]. More work would be needed to understand the functions of these effectors in Ppal and how they aid in infection and in manipulating cocoa’s immunity to cause disease.
5. Cocoa Diseases by P. palmivora (Ppal)
T. cacao is the only species within the Theobroma genus that is cultivated by about 6 million farmers globally [137]. The species is divided into three main recognized genetic groups: Criollo, Forastero and Trinitario [138,139]. The latter is a hybrid from crosses between the Criollo and Forastero varieties and is cultivated in many parts of the world due to its aromatic, high-yielding and disease-resistant characteristics [137,139]. Some cocoa breeding programs have been focused on selecting lines resistant to many plant pathogens as well as Ppal [140].
Ppal causes two main types of disease on cocoa trees: black pod and stem canker. Figure 3 shows a mature healthy cocoa plant growing in a glasshouse, highlighting the target sites of Ppal infection. In black pod, pods or cherelles (immature pods, Figure 3E,F) can be infected at any place on the surface, however, initial infection is usually at the tip or stem end.
Studies in 12 diverse cocoa genotypes demonstrated that germinating zoospores of Ppal could penetrate through stomata, epidermal hair base, scar and by direct penetration of pods [141,142]. Symptoms are a brown or black spot on the pod, which spreads to cover the whole pod.
In stem canker, Ppal mycelia spread from infected pods [143] along the stalk into the flower cushions (Figure 3B) and further along the stem or via direct infection in wounds along the stem. Newly infected bark may not show any external symptom, but the cambial layer would be infected [144]. Symptoms of canker are the formation of reddish water-soaked lesions with dark brown to black margins, and in some cases, reddish-brown liquid oozed from these lesions, usually through cracks in the bark [143,144]. In Sulawesi, incidence and severity of stem canker in cocoa by Ppal increases during the wet season, especially in more susceptible genotypes [145]. Okey and colleagues [146] demonstrated a strong correlation between bark hardness and moisture content with canker resistance to Ppal in greenhouse studies. The same authors [146] proposed that extra-xylary tissue hardness associated with fiber content or deposition of suberin, callose and lignin could hinder the progress of fungal pathogens and that bark hardness, acting as a mechanical barrier, had contributed to the slow rate of tissue colonization of Ppal in the canker-resistant cocoa line used in greenhouse studies, leading to the use and selection of resistant cultivars with acceptable horticultural traits [147]. Such traits in Ppal resistant lines were related to lignin concentration in cocoa stems [148] and the high activities of plant enzymes such as peroxidase (PO) and polyphenoloxidase (PPO), which are involved in phenol oxidation and lignin production, and phenylalanine ammonia-lyase (PAL) in lignin and phenol biosynthesis in response to Ppal infection in cocoa-resistant clones [149]. In another model system, it has been shown that both peroxidase activity and lignin deposition increased in the cell suspensions of the resistant Capsicum annuum (pepper) variety to P. capsici elicitors compared to the susceptible or intermediate pepper varieties [150].
Other approaches to controlling stem canker in cocoa have involved the application by trunk injection of potassium phosphonate (phosphite) [151]. Potassium phosphonate has been used as a systemically translocated chemical to protect plants against oomycetes due to its ability to induce rapid and localized defense responses similarly observed in phosphonate-treated A. thaliana seedlings inoculated with Ppal zoospores [67].
Other parts of the cocoa plants can be infected by Ppal i.e., the flower cushion, the chupons, leaves (Figure 3A–F) and seedlings, as well as the roots [20,82,152,153,154]. Work on cocoa roots by Oppoku and Wheeler [155] demonstrated that Ppal persisted in association with roots for at least 6 months, and the recovery of the oomycete generally declined with time.
However, cocoa as a perennial plant takes a long time to grow and the selection of Ppal-resistant lines or germplasm requires quick, easy and cheap inoculation testing methods [141,142]. Detached or attached cocoa leaves and pods are some of the materials used to determine resistance [141,142]. Work by Iwaro and colleagues [142] tested leaves and pods of various clones for resistance and demonstrated that there were two levels of resistance in both organs. Their studies showed a poor relationship between pod and leaf reaction at Ppal penetration stage of infection while a high positive correlation was observed between pod and leaf resistance at the post-penetration stage of infection, suggesting the role of a systemic mechanism in post-penetration resistance [141,142]. Resistance can be effective from the point of entry of the pathogen (penetration) or at a later stage during its development within the host tissue (post-penetration) [156]; thus, penetration and post-penetration resistance can both be used as selection criteria in breeding to improve the existing levels of cocoa resistance to Ppal. In expression pattern studies in susceptible cocoa pods, Ali and colleagues [9,55] highlighted the differences between Ppal and P. megakarya, especially in Ppal-inoculated wounded pod pieces, where Ppal is known for its rapid progression when penetrating through wounds. Previous studies in betelvine and papaya have shown that there is a synergistic effect with plant pathogenic nematodes, Rotylenchulus reniformis and Meloidogyne incognita, which predispose these plants to attack by Ppal [157,158,159].
6. Ppal Isolates from Papua New Guinea Cocoa Plantations
Analyses of random amplified microsatellites (RAMs) of 263 of the Phytophthora isolates demonstrated that there was limited morphological, physiological and genetic diversity of Ppal isolates from cocoa pods in Papua New Guinea (PNG), and that Ppal from cocoa in PNG formed a single, continuous largely asexual population [13,160] (Figure 1A–N). Recent studies in the genetic diversity among 81 Ppal isolates from various host plants and geographical regions in Indonesia and Japan using rep-PCR (BOX, ERIC, REP and M13) and microsatellite markers demonstrated that the isolates clustered into six groups, which corresponded more to geographic regions rather than host plants or mating types [161]. These studies highlighted the importance of implementing key quarantine measures to prevent the spread of Ppal-contaminated plant materials to different geographical regions [160,161].
However, work by Appiah et al. [162] demonstrated that Ppal isolates from different geographical sources associated with black pod disease in cocoa showed considerable inter- and intra-specific morphometric variation. This is important as correct identification of the pathogen is crucial, since Ppal can be controlled by crop sanitation alone, whereas Phytophthora megakarya (Table 1) cannot [9,163,164]. Through sexual reproduction or interspecific hybridization, Phytophthora could gain allelic diversity and achieve large sexual/clonal population sizes through rapid proliferation [165]. These would enhance pathogen fitness by generating recombinant genotypes that may be more pathogenic or resistant to crop protection chemicals [166].
7. Conclusions
The question remains as to why these Ppal isolates formed clusters based on their geographic regions and what are the characteristics that have allowed such isolates to infect and be pathogenic to current cocoa lines. Goodwin [167] presented many factors that could contribute to the genetic variation in Phytophthora population. Migration of Phytophthora, via the introduction of contaminated plant materials to different geographical regions or from centers of origins, would put pressure on founder Phytophthora populations [167]. Such populations would be subjected to genetic drift due to changes in environmental conditions; selection would contribute to overall fitness and, mating as heterothallic species, should contain high level of heterozygosity [167]. Brasier [168] proposed that the soil, with Phytophthora resting inoculum oospores and chlamydospores, would be a reservoir of genetic variation but the reinfection of the hosts would exert strong directional selection on such variation, favoring genotypes capable of infecting a particular host species or part of the host. Furthermore, successful pathogen genotypes could be maintained by asexual reproduction by directional and stabilizing selection as long as the host is still available [168]. Under episodic selection during widespread and continuous crop monoculture or following the introduction of a new and susceptible host population for example, rapid speciation could occur, increasing specialization on a single host species [168,169]. Combined with asexual reproduction and pathogenic feedback, this would lead to a reduction in genetic variability and to the emergence of a clone [169].
In the case of the characterized PNG cocoa Ppal isolates [160], a study investigating the differences in gene expression related to PTI and ETI during cocoa pod and stem infections would be useful in understanding the differences in pathogenicity observed in cocoa plantation fields at different locations in PNG.
Funding
This research received no external funding.
Acknowledgments
F.P.-W. would like to thank David Guest for providing the P. palmivora isolates from cocoa (Papua New Guinea) culture collection and cocoa plants used to generate micrographs for this review. She also thanks M.L. Walker for careful reading of the manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Erwin, D.; Ribeiro, O. Phytophthora Diseases Worldwide; APS Press: St. Paul, MN, USA, 1996. [Google Scholar]
- McCarthy, C.G.P.; Fitzpatrick, D.A. Phylogenomic reconstruction of the oomycete phylogeny derived from 37 genomes. mSphere 2017, 2, e00095-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acebo-Guerrero, Y.; Hernández-Rodríguez, A.; Heydrich-Pérez, M.; El Jaziri, M.; Hernández-Lauzardo, A. Management of black pod rot in cacao (Theobroma cacao L.): A review. Fruits 2012, 67, 41–48. [Google Scholar] [CrossRef]
- Guest, D. Black pod: Diverse pathogens with a global impact on cocoa yield. Phytopathology 2007, 97, 1650–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, E.J. Report of the imperial mycologist 1918–1919. Sci. Rep. Agric. Res. Inst. Pusa 1919, 1–82. [Google Scholar]
- Ploetz, R. The Impact of Diseases on Cacao Production: A Global Overview. In Cacao Diseases. A History of Old Enemies and New Encounters, 1st ed.; Bailey, B., Meinhardt, L., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 33–39. [Google Scholar]
- Kroon, L.; Brouwer, H.; de Cock, A.; Govers, F. The genus Phytophthora anno 2012. Phytopatholoy 2012, 102, 348–364. [Google Scholar] [CrossRef] [Green Version]
- Lamour, K.H.; Mudge, J.; Gobena, D.; Hurtado-Gonzales, O.P.; Schmutz, J.; Kuo, A.; Miller, N.A.; Rice, B.J.; Raffaele, S.; Cano, L.M.; et al. Genome sequencing and mapping reveal loss of heterozygosity as a mechanism for rapid adaptation in the vegetable pathogen Phytophthora capsici. Mol. Plant Microbe Interact. 2012, 25, 1350–1360. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Shao, J.; Lary, D.; Strem, M.; Meinhardt, L.; Bailey, B. Phytophthora megakarya and P. palmivora, Causal Agents of Black Pod Rot, Induce Similar Plant Defense Responses Late during Infection of Susceptible Cacao Pods. Front. Plant Sci. 2017, 8, 169. [Google Scholar] [CrossRef] [Green Version]
- Mao, Y.; Tyler, B.M. Genome organization of Phytophthora megasperma f.sp. glycinea. Exp. Mycol. 1991, 15, 283–291. [Google Scholar] [CrossRef]
- Mélida, H.; Sandoval-Sierra, J.V.; Diéguez-Uribeondo, J.; Bulonea, V. Analyses of extracellular carbohydrates in oomycetes unveil the existence of three different cell wall types. Eukaryot. Cell 2013, 12, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Christen, J.; Hohl, H.R. Growth and ultrastructural differentiation of sporangia in Phytophthora palmivora. Can. J. Microbiol. 1972, 18, 1959–1964. [Google Scholar] [CrossRef]
- Saul Maora, J. Diversity of Phytophthora palmivora on Cocoa in Papua New Guinea. Ph.D. Thesis, University of Sydney, Sydney, Australia, August 2008. [Google Scholar]
- Cameron, J.N.; Carlile, M.J. Negative geotaxis of zoospores of the fungus Phytophthora. J. Gen. Microbiol. 1977, 98, 599–602. [Google Scholar] [CrossRef] [Green Version]
- Van West, P.; Morris, B.M.; Reid, B.; Appiah, A.A.; Osborne, M.C.; Campbell, T.A.; Shepherd, S.J.; Gow, N.A.R. Oomycete Plant Pathogens Use Electric Fields to Target Roots. MPMI 2002, 15, 790–798. [Google Scholar] [CrossRef] [Green Version]
- Bimpong, C.E.; Clerk, G.C. Motility and chemotaxis in zoospores of Phytophthora palmivora (Butl.) Butl. Ann. Bot. 1970, 34, 617–624. [Google Scholar] [CrossRef]
- Cameron, J.; Carlile, M. Fatty acids, aldehydes and alcohols as attractants for zoospores of Phytophthora palmivora. Nature 1978, 271, 448–449. [Google Scholar] [CrossRef]
- Cameron, J.N.; Carlile, M.J. Negative chemotaxis of zoospores of the fungus Phytophthora palmivora. J. Gen. Microbiol. 1980, 120, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Uchida, J.Y.; Aragaki, M. Phytophthora Diseases of Orchids in Hawaii; HITAHR Research Extension Series 129; University of Hawaii: Honolulu, HI, USA, 1991; pp. 1–11. [Google Scholar]
- Surujdeo-Maharaj, S.; Sreenivasan, T.N.; Motilal, L.A.; Umaharan, P. Black Pod and Other Phytophthora Induced Diseases of Cacao: History, Biology, and Control. In Cacao Diseases. A History of Old Enemies and New Encounters, 1st ed.; Bailey, B., Meinhardt, L., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 213–266. [Google Scholar]
- Zentmyer, G.A.; Mitchell, D.J.; Jefferson, L.; Roheim, J.; Carnes, D. Distribution of mating types of Phytophthora palmivora. Phytopathology 1973, 63, 663–667. [Google Scholar] [CrossRef]
- Hemmes, D.; Lerma, A. The Ultrastructure of developing and germinating chlamydospores of Phytophthora palmivora. Mycologia 1985, 77, 743–755. [Google Scholar] [CrossRef]
- Ko, W. Long-term storage and survival structure of three species of Phytophthora in water. J. Gen. Plant. Pathol. 2003, 69, 186–188. [Google Scholar]
- Bodah, E.T. Root rot diseases in plants: A review of common causal agents and management strategies. Agric. Res. Technol. Open Access J. 2017, 5, 555661. [Google Scholar] [CrossRef]
- Rey, T.; Chatterjee, A.; Buttay, M.; Toulotte, J.; Schornack, S. Medicago truncatula symbiosis mutants affected in the interaction with a biotrophic root pathogen. New Phytol. 2015, 206, 497–500. [Google Scholar] [CrossRef] [Green Version]
- Rey, T.; Schornack, S. Interactions of beneficial and detrimental root colonizing filamentous microbes with plant hosts. Genome Biol. 2013, 14, 121. [Google Scholar] [CrossRef] [PubMed]
- Chaparro-Garcia, A.; Wilkinson, R.C.; Gimenez-Ibanez, S.; Findlay, K.; Coffey, M.D.; Zipfel, C.; Rathjen, J.P.; Kamoun, S.; Schornack, S. The receptor-like kinase SERK3/BAK1 is required for basal resistance against the late blight pathogen Phytophthora infestans in Nicotiana benthamiana. PLoS ONE 2011, 6, e16608. [Google Scholar] [CrossRef] [PubMed]
- Evangelisti, E.; Gogleva, A.; Hainaux, T.; Doumane, M.; Tulin, F.; Quan, C.; Yunusov, T.; Floch, K.; Schornack, S. Time-resolved dual root-microbe transcriptomics reveals early induced Nicotiana benthamiana genes and conserved infection-promoting Phytophthora palmivora effectors. BMC Biol. 2017, 15, 39. [Google Scholar] [CrossRef] [PubMed]
- Carella, P.; Gogleva, A.; Tomaselli, M.; Alfs, C.; Schornack, S. Phytophthora palmivora establishes tissue-specific intracellular infection structures in the earliest divergent land plant lineage. Proc. Natl. Acad. Sci. USA 2018, 115, E3846–E3855. [Google Scholar] [CrossRef] [Green Version]
- Harris, D.C.; Cardon, J.A.; Justin, S.H.F.W.; Passey, A.J. Phytophthora palmivora on cultured roots of coconut. Trans. Brit. Mycol. Soc. 1984, 82, 249–255. [Google Scholar] [CrossRef]
- Mohamed Azni, I.; Sundram, S.; Ramachandran, V. Pathogenicity of Malaysian Phytophthora palmivora on cocoa, durian, rubber and oil palm determines the threat of bud rot disease. For. Pathol. 2019, 49, e12557. [Google Scholar] [CrossRef]
- Ochoa, J.C.; Herrera, M.; Navia, M.; Romero, H.M. Visualization of Phytophthora palmivora infection in oil palm leaflets with fluorescent proteins and cell viability markers. Plant. Pathol. J. 2019, 35, 19–31. [Google Scholar]
- Sarria, G.; Martinez, G.; Varon, F.; Drenth, A.; Guest, D. Histopathological studies of the process of Phytophthora palmivora infection in oil palm. Eur. J. Plant Pathol. 2016, 145, 39–51. [Google Scholar] [CrossRef]
- Johri, J.K.; Devi, S. Ultrastructural studies on Phytophthora palmivora infection on betelvine (Piper Betle, L.). Arch. Phytopathol. Pflanzenschutz. 1998, 31, 233–240. [Google Scholar] [CrossRef]
- Widmer, T.L. Phytophthora palmivora. For. Phytophthoras 2014, 4. [Google Scholar] [CrossRef]
- Widmer, T.L.; Graham, J.H.; Mitchell, D.J. Histological comparison of fibrous root infection of disease-tolerant and susceptible citrus hosts by Phytophthora nicotianae and P. palmivora. Phytopathology 1998, 88, 389–395. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, A.; Joseph, L.; Bindu, R.C. An insight into Hevea—Phytophthora interaction: The story of Hevea defense and Phytophthora counter defense mediated through molecular signalling. Curr. Plant Biol. 2019, 17, 33–41. [Google Scholar] [CrossRef]
- Hamill, S.D. Fruit rot of papaya caused by Phytophthora palmivora in Queensland. Australas. Plant Pathol. 1987, 16, 22. [Google Scholar] [CrossRef]
- Purwantara, A. Infection of Phytophthora palmivora from soil in cocoa plantation. Pelita Perkeb. 2008, 24, 205–218. [Google Scholar] [CrossRef]
- Bimpong, C.E.; Hickman, C.J. Ultrastructural and cytochemical studies of zoospores, cysts, and germinating cysts of Phytophthora palmivora. Can. J. Bot. 1975, 13, 1310–1327. [Google Scholar] [CrossRef]
- Sing, V.O.; Bartnicki-Garcia, S. Adhesion of zoospores of Phytophthora palmivora to solid surfaces. Phytopathology 1972, 62, 790. [Google Scholar]
- Deacon, J.W. Ecological implications of recognition events in the pre-infection stages of root pathogens. New Phytol. 1996, 133, 135–145. [Google Scholar] [CrossRef]
- Hardham, A.R.; Gubler, F. Polarity of attachment of zoospores of a root pathogen and prealignment of the emerging germ-tubes. Cell Biol. Int. 1990, 14, 947–956. [Google Scholar] [CrossRef]
- Islam, M.T.; Ito, T.; Tahara, S. Microscopic studies on attachment and differentiation of zoospores of the phytopathogenic fungus Aphanomyces cochlioides. J. Gen. Plant Pathol. 2002, 68, 111–117. [Google Scholar] [CrossRef]
- Dijksterhuis, J.; Deacon, J. Defective zoospore encystment and suppressed cyst germination of Phytophthora palmivora caused by transient leaching treatments. Antonie Van Leeuwenhoek 2003, 83, 235–243. [Google Scholar] [CrossRef]
- Grant, B.R.; Irving, H.R.; Radda, M. The effect of pectin and related compounds on encystment and germination of Phytophthora palmivora zoospores. J. Gen. Microbiol. 1984, 131, 669–676. [Google Scholar] [CrossRef] [Green Version]
- Irving, H.R.; Griffith, J.M.; Grant, B.R. Calcium efflux associated with encystment of Phytophthora palmivora zoospores. Cell Calcium 1984, 5, 487–500. [Google Scholar] [CrossRef]
- Zhang, Q.; Griffith, J.; Moore, J.; Iser, J.; Grant, B. The effect of modified pectin, pectin fragments and cations on Phytophthora palmivora zoospores. Phytochemistry 1990, 29, 695–700. [Google Scholar] [CrossRef]
- Guzmán, P.; Fernández, V.; García, M.; Khayet, M.; Fernández, A.; Gil, L. Localization of polysaccharides in isolated and intact cuticles of eucalypt, poplar and pear leaves by enzyme-gold labelling. Plant Physiol. Biochem. 2014, 76, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horowitz, B.; Ospina-Giraldo, M. The pectin methylesterase gene complement of Phytophthora sojae: Structural and functional analyses, and the evolutionary relationships with its oomycete homologs. PLoS ONE 2015, 10, e0142096. [Google Scholar] [CrossRef]
- Li, P.; Feng, B.; Wang, H.; Tooley, P.; Zhang, X.; Li, P. Isolation of nine Phytophthora capsici pectin methylesterase genes which are differentially expressed in various plant species. J. Basic. Microbiol. 2011, 51, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Mingora, C.; Ewer, J.; Ospina-Giraldo, M. Comparative structural and functional analysis of genes encoding pectin methylesterases in Phytophthora spp. Gene 2014, 538, 74–83. [Google Scholar] [CrossRef]
- Feng, B.; Li, P.; Wang, H.; Zhang, X. Functional analysis of Pcpme6 from oomycete plant pathogen Phytophthora capsici. Microb. Pathog. 2010, 49, 23–31. [Google Scholar] [CrossRef]
- Bircher, U.; Hohl, H.R. Environmental signalling during induction of appressorium formation in Phytophthora. Mycol. Res. 1997, 101, 395–402. [Google Scholar] [CrossRef]
- Ali, S.S.; Amoako-Attah, I.; Bailey, R.A.; Strem, M.D.; Schmidt, M.; Akrofi, A.Y.; Surujdeo-Maharaj, S.; Kolawole, O.O.; Begoude, B.A.D.; ten Hoopen, G.M.; et al. PCR-based identification of cacao black pod causal agents and identification of biological factors possibly contributing to Phytophthora megakarya’s field dominance in West Africa. Plant Pathol. 2016, 65, 1095–1108. [Google Scholar] [CrossRef]
- Tey, C.C. Control of Phytophthora palmivora (Butl.) Butl. on Cocoa. Ph.D. Thesis, University of London, London, UK, March 1982. [Google Scholar]
- Dale, M.L.; Irwin, J.A.G. Stomata as an infection court for Phytophthora megasperma f. sp. medicaginis in chickpea and a histological study of infection. Phytopathology 1991, 81, 375–379. [Google Scholar] [CrossRef]
- Howard, R.J. Breaching the Outer Barriers—Cuticle and Cell Wall Penetration. In Plant Relationships. The Mycota (A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); Carroll, G.C., Tudzynski, P., Eds.; Springer: Berlin/Heidelberg, Germany, 1997; Volume 5. [Google Scholar]
- Kebdani, N.; Pieuchot, L.; Deleury, E.; Panabières, F.; Le Berre, J.-Y.; Gourgues, M. Cellular and molecular characterization of Phytophthora parasitica appressorium-mediated penetration. New Phytologist. 2010, 185, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Le Fevre, R.; O’Boyle, B.; Moscou, M.; Schornack, S.; Le Fevre, R. Colonization of barley by the broad-host hemibiotrophic pathogen Phytophthora palmivora uncovers a leaf development-dependent involvement of Mlo. MPMI 2016, 29, 385–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardham, A. The cell biology behind Phytophthora pathogenicity. Australas. Plant Pathol. 2001, 30, 91–98. [Google Scholar] [CrossRef]
- Attard, A.; Gourgues, M.; Callemeyn-Torre, N.; Keller, H. The immediate activation of defense responses in Arabidopsis roots is not sufficient to prevent Phytophthora parasitica infection. New Phytologist. 2010, 187, 449–460. [Google Scholar] [CrossRef]
- Redondo, M.; Pérez-Sierra, A.; Abad-Campos, P.; Torres, L.; Solla, A.; Reig-Armiñana, J.; García-Breijo, F. Histology of Quercus ilex roots during infection by Phytophthora cinnamomi. Trees 2015, 29, 1943–1957. [Google Scholar] [CrossRef]
- Huisman, R.; Bouwmeester, K.; Brattinga, M.; Govers, F.; Bisseling, T.; Limpens, E. Haustorium formation in Medicago truncatula roots infected by Phytophthora palmivora does not involve the common endosymbiotic program shared by AM fungi and rhizobia. MPMI 2015, 8, 1271–1280. [Google Scholar] [CrossRef] [Green Version]
- Bozkurt, T.; Kamoun, S. The plant–pathogen haustorial interface at a glance. J. Cell Sci. 2020, 133, jcs237958. [Google Scholar] [CrossRef] [Green Version]
- Calonge, F. Ultrastructure of the haustoria or intracellular hyphae in four different fungi. Archiv. Für. Mikrobiol. 1969, 67, 209–225. [Google Scholar] [CrossRef]
- Daniel, R.; Guest, D. Defence responses induced by potassium phosphonate in Phytophthora palmivora-challenged Arabidopsis thaliana. Physiol. Mol. Plant Pathol. 2006, 67, 194–201. [Google Scholar] [CrossRef]
- Ehrlich, M.; Ehrlich, H. Fine structure of the host-parasite interfaces in mycoparasitism. Annu. Rev. Phytopathol. 1971, 9, 155–184. [Google Scholar] [CrossRef]
- Szabo, L.; Bushnell, W. Hidden Robbers: The role of fungal haustoria in parasitism of plants. Proc. Natl. Acad. Sci. USA 2001, 98, 7654–7655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozkurt, T.; Richardson, A.; Dagdas, Y.; Mongrand, S.; Kamoun, S.; Raffaele, S. The plant membrane-associated remorin1.3 accumulates in discrete perihaustorial domains and enhances susceptibility to Phytophthora infestans. Plant Physiol. 2014, 165, 1005–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhari, P.; Ahmed, B.; Joly, D.L.; Germain, H. Effector biology during biotrophic invasion of plant cells. Virulence 2014, 5, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Chepsergon, J.; Motaung, T.; Bellieny-Rabelo, D.; Moleleki, L. Organize, Don’t Agonize: Strategic success of Phytophthora species. Microorganisms 2020, 8, 917. [Google Scholar] [CrossRef]
- Fawke, S.; Doumane, M.; Schornack, S. Oomycete interactions with plants: Infection strategies and resistance principles. MMBR 2015, 79, 263–280. [Google Scholar] [CrossRef] [Green Version]
- Judelson, H.S.; Ah-Fong, A.M.V. Exchanges at the plant-oomycete interface that influence disease. Plant Physiol. 2019, 179, 1198–1211. [Google Scholar] [CrossRef] [Green Version]
- Kamoun, S. A catalogue of the effector secretome of plant pathogenic oomycetes. Annu. Rev. Phytopathol. 2006, 44, 41–60. [Google Scholar] [CrossRef] [Green Version]
- Schornack, S.; Huitema, E.; Cano, L.M.; Bozkurt, T.O.; Oliva, R.; Van Damme, M.; Schwizer, S.; Raffaele, S.; Chaparro-Garcia, A.; Farrer, R.; et al. Ten things to know about oomycete effectors. Mol. Plant Pathol. 2009, 10, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Stassen, J.H.M.; Van den Ackerveken, G. How do oomycete effectors interfere with plant life? Curr. Opin. Plant Biol. 2011, 14, 407–414. [Google Scholar] [CrossRef]
- Takemoto, D.; Yuri Mizuno, Y. Belowground and Aboveground Strategies of Plant Resistance against Phytophthora Species. In Belowground Defence Strategies in Plants. Signaling and Communication in Plants, 1st ed.; Vos, C., Kazan, K., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 151–169. [Google Scholar] [CrossRef]
- Tyler, B.M. Molecular basis of recognition between Phytophthora species and their hosts. Annu. Rev. Phytopathol. 2002, 40, 137–167. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Welsh, L.; Thorpe, P.; Whisson, S.C.; Boevink, P.C.; Birch, P. The Phytophthora infestans haustorium is a site for secretion of diverse classes of infection-associated proteins. MBio 2018, 9, e01216-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuechtbauer, W.; Yunusov, T.; Bozsóki, Z.; Gavrin, A.; James, E.K.; Stougaard, J.; Schornack, S.; Radutoiu, S. LYS 12 LysM Receptor decelerates Phytophthora palmivora disease progression in Lotus japonicus. Plant J. 2018, 93, 297–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orellana, R.G. Infection and tissue changes of Theobroma cacao L. by Phytophthora palmivora Butl. Turrialba 1953, 3, 167–172. [Google Scholar]
- Wang, S.; Boevink, P.C.; Welsh, L.; Zhang, R.; Whisson, S.C.; Birch, P. Delivery of cytoplasmic and apoplastic effectors from Phytophthora infestans haustoria by distinct secretion pathways. New Phytol. 2017, 216, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Caplan, J.; Padmanabhan, M.; Dinesh-Kumar, S. Plant NB-LRR immune receptors: From recognition to transcriptional reprogramming. Cell Host Microbe 2008, 3, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Joshi, R.K.; Nayak, S. Functional characterization and signal transduction ability of nucleotide-binding site-leucine-rich repeat resistance genes in plants. GMR 2011, 10, 2637–2652. [Google Scholar] [CrossRef]
- Zhang, R.; Zheng, F.; Wei, S.; Zhang, S.; Li, G.; Cao, P.; Zhao, S. Evolution of disease defense genes and their regulators in plants. Int. J. Mol. Sci. 2019, 20, 335. [Google Scholar] [CrossRef] [Green Version]
- De Araújo, A.C.; Campos De Assis Fonseca, F.; Cotta, M.G.; Alves, G.S.C.; Miller, R.N.G. Plant NLR receptor proteins and their potential in the development of durable genetic resistance to biotic stresses, Biotechnol. Res. Innov. 2019, 3, 80–94. [Google Scholar]
- Balint-Kurti, P. The plant hypersensitive response: Concepts, control and consequences. Mol. Plant Pathol. 2019, 20, 1163–1178. [Google Scholar] [CrossRef] [Green Version]
- Eitas, T.K.; Dangl, J.L. NB-LRR proteins: Pairs, pieces, perception, partners, and pathways. Curr. Opin. Plant Biol. 2010, 13, 472–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomma, B.P.H.J.; Nürnberger, T.; Joosten, M.H.A.J. Of PAMPs and effectors: The blurred PTI-ETI Dichotomy. Plant. Cell 2011, 23, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Gu, B.; Cao, X.; Zhou, X.; Chen, Z.; Wang, Q.; Liu, W.; Chen, Q.; Zhao, H. The histological, effectoromic, and transcriptomic analyses of Solanum pinnatisectum reveal an upregulation of multiple NBS-LRR genes suppressing Phytophthora infestans infection. Int. J. Mol. Sci. 2020, 21, 3211. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, B.; Kalsi, H.; Godbole, P.; Malankar, N.; Thiagarayaselvam, A.; Siddappa, S.; Thulasiram, H.; Chakrabarti, S.; Banerjee, A. MiRNA160 is associated with local defense and systemic acquired resistance against Phytophthora infestans infection in potato. J. Exp. Bot. 2018, 69, 2023–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, N.; Cui, J.; Shi, Y.; Yang, G.; Zhou, X.; Hou, X.; Meng, J.; Luan, Y. Tomato lncRNA23468 functions as a competing endogenous RNA to modulate genes by decoying miR482b in the tomato interaction. Hortic. Res. 2019, 6, 28. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Yan, Q.; Gan, S.; Xue, D.; Dou, D.; Guo, N.; Xing, H. Overexpression of gma-miR1510a/b suppresses the expression of a NB-LRR domain gene and reduces resistance to Phytophthora sojae. Gene 2017, 621, 32–39. [Google Scholar] [CrossRef]
- Qiao, Y.; Liu, L.; Xiong, Q.; Flores, C.; Wong, J.; Shi, J.; Wang, X.; Liu, X.; Xiang, Q.; Jiang, S.; et al. Oomycete pathogens encode RNA silencing suppressors. Nat. Genet. 2013, 45, 330–333. [Google Scholar] [CrossRef] [Green Version]
- Sós-Hegedűs, A.; Domonkos, Á.; Tóth, T.; Gyula, P.; Kaló, P.; Szittya, G. Suppression of NB-LRR genes by miRNAs promotes nitrogen-fixing nodule development in Medicago truncatula. Plant. Cell Environ. 2020, 43, 1117–1129. [Google Scholar] [CrossRef] [Green Version]
- Hardham, A.R.; Blackman, L.M. Phytophthora cinnamomi. Mol. Plant Pathol. 2018, 19, 260–285. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Jiao, F. Effectors of Phytophthora pathogens are powerful weapons for manipulating host immunity. Planta 2019, 250, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Raaymakers, T.M.; Van den Ackerveken, G. Extracellular recognition of oomycetes during biotrophic infection of plants. Front. Plant Sci. 2016, 7, 906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fellbrich, G.; Romanski, A.; Varet, A.; Blume, B.; Brunner, F.; Engelhardt, S.; Felix, G.; Kemmerling, B.; Krzymowska, M.; Nürnberger, T. NPP1, a Phytophthora-associated trigger of plant defense in parsley and Arabidopsis. Plant J. 2002, 32, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Qutob, D.; Kemmerling, B.; Brunner, F.; Küfner, I.; Engelhardt, S.; Gust, A.A.; Luberacki, B.; Seitz, H.U.; Stahl, D.; Rauhut, T.; et al. Phytotoxicity and innate immune responses induced by Nep1-like proteins. Plant Cell 2006, 18, 3721–3744. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, S.; Grosser, K.; Voegele, R.T.; Kassemeyer, H.-H.; Fuchs, R. Identification and characterization of Nep1-like proteins from the grapevine downy mildew pathogen Plasmopara viticola. Front. Plant Sci. 2020, 11, 65. [Google Scholar] [CrossRef] [PubMed]
- Seidl, M.F.; Van den Ackerveken, G. Activity and phylogenetics of the broadly occurring family of microbial Nep1-like proteins. Annu. Rev. Phytopathol. 2019, 57, 367–386. [Google Scholar] [CrossRef]
- Bailey, B.A.; Bae, H.; Strem, M.D.; Antúnez de Mayolo, G.; Guiltinan, M.J.; Verica, J.A.; Maximova, S.N.; Bowers, J.H. Developmental expression of stress response genes in Theobroma cacao leaves and their response to Nep1 treatment and a compatible infection by Phytophthora megakarya. Plant Physiol. Biochem. 2005, 43, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.; Bowers, J.H.; Tooley, P.W.; Bailey, B.A. NEP1 orthologs encoding necrosis and ethylene inducing proteins exist as a multigene family in Phytophthora megakarya, causal agent of black pod disease on cacao. Mycol. Res. 2005, 109, 1373–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.S.; Shao, J.; Lary, D.J.; Kronmiller, B.A.; Shen, D.; Strem, M.D.; Amoako-Attah, I.; Akrofi, A.Y.; Begoude, B.A.D.; ten Hoopen, G.M.; et al. Phytophthora megakarya and Phytophthora palmivora, closely related causal agents of cacao black pod rot, underwent increases in genome sizes and gene numbers by different mechanisms. Genome Biol. Evol. 2017, 9, 536–557. [Google Scholar] [CrossRef]
- Kelley, B.S.; Lee, S.-J.; Damasceno, C.M.B.; Chakravarthy, S.; Kim, B.-D.; Martin, G.B.; Rose, J.K.C. A secreted effector protein (SNE1) from Phytophthora infestans is a broadly acting suppressor of programmed cell death. Plant J. 2010, 62, 357–366. [Google Scholar] [CrossRef]
- Gouget, A.; Senchou, V.; Govers, F.; Sanson, A.; Barre, A.; Rouge, P.; Pont-Lezica, R.; Canut, H.; Gouget, A. Lectin receptor kinases participate in protein-protein interactions to mediate plasma membrane-cell wall adhesions in Arabidopsis. Plant Physiol. 2006, 140, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Van Holle, S.; Van Damme, E.J.M. Signaling through plant lectins: Modulation of plant immunity and beyond. Biochem. Soc. Trans. 2018, 46, 217–233. [Google Scholar] [CrossRef] [PubMed]
- Furuichi, N.; Tomiyama, K.; Doke, N. The role of potato lectin in the binding of germ tubes of Phytophthora infestans to potato cell membrane. Physiol. Plant Pathol. 1980, 16, 249–256. [Google Scholar] [CrossRef]
- Garas, N.; Kuć, J. Potato lectin lyses zoospores of Phytophthora infestans and precipitates elicitors of terpenoid accumulation produced by the fungus. Physiol. Plant Pathol. 1981, 18, 227–237. [Google Scholar] [CrossRef]
- Bouwmeester, K.; Han, M.; Blanco-Portales, R.; Song, W.; Weide, R.; Guo, L.; Van Der Vossen, E.; Govers, F. The Arabidopsis lectin receptor kinase Lec RK-I.9 enhances resistance to Phytophthora infestans in Solanaceous plants. Plant Biotechnol. J. 2014, 12, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Gaulin, E.; Jauneau, A.; Villalba, F.; Rickauer, M.; Esquerré-Tugayé, M.T.; Bottin, A. The CBEL glycoprotein of Phytophthora parasitica var-nicotianae is involved in cell wall deposition and adhesion to cellulosic substrates. J. Cell Sci. 2002, 115, 4565–4575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.W.; Ospina-Giraldo, M. Novel cellulose-binding-domain protein in Phytophthora is cell wall localized. PLoS ONE 2011, 6, e23555. [Google Scholar] [CrossRef] [PubMed]
- Martinez, T.; Texier, H.; Nahoum, V.; Lafitte, C.; Cioci, G.; Heux, L.; Dumas, B.; O’Donohue, M.; Gaulin, E.; Dumon, C. Probing the functions of carbohydrate binding modules in the CBEL protein from the oomycete Phytophthora parasitica. PLoS ONE 2015, 10, e0137481. [Google Scholar] [CrossRef]
- Khatib, M.; Lafitte, C.; Esquerré-Tugayé, M.-T.; Bottin, A.; Rickauer, M. The CBEL elicitor of Phytophthora parasitica var. nicotianae activates defence in Arabidopsis thaliana via three different signalling pathways. New Phytol. 2004, 162, 501–510. [Google Scholar] [CrossRef]
- Larroque, M.; Belmas, E.; Martinez, T.; Vergnes, S.; Ladouce, N.; Lafitte, C.; Gaulin, E.; Dumas, B. Pathogen-associated molecular pattern-triggered immunity and resistance to the root pathogen Phytophthora parasitica in Arabidopsis. J. Exp. Bot. 2013, 64, 3615–3625. [Google Scholar] [CrossRef] [Green Version]
- Kamoun, S.; Young, M.; Förster, H.; Coffey, M.D.; Tyler, B.M. Potential role of elicitins in the interaction between Phytophthora species and tobacco. Appl. Environ. Microbiol. 1994, 60, 1593–1598. [Google Scholar] [CrossRef] [Green Version]
- Huitema, E.; Vleeshouwers, V.G.; Cakir, C.; Kamoun, S.; Govers, F. Differences in intensity and specificity of hypersensitive response induction in Nicotiana spp. by INF1, INF2A, and INF2B of Phytophthora infestans. MPMI 2005, 18, 183–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanneganti, T.D.; Huitema, E.; Cakir, C.; Kamoun, S. Synergistic interactions of the plant cell death pathways induced by Phytophthora infestans Nepl-like protein PiNPP1.1 and INF1 elicitin. MPMI 2006, 19, 854–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Churngchow, N.; Rattarasarn, M. The elicitin secreted by Phytophthora palmivora, a rubber tree pathogen. Phytochemistry 2000, 54, 33–38. [Google Scholar] [CrossRef]
- Dutsadee, C.; Nunta, C. Induction of peroxidase, scopoletin, phenolic compounds and resistance in Hevea brasiliensis by elicitin and a novel protein elicitor purified from Phytophthora palmivora. Physiol. Mol. Plant Pathol. 2008, 72, 179–187. [Google Scholar] [CrossRef]
- Pettongkhao, S.; Churngchow, N. Novel cell death-inducing elicitors from Phytophthora palmivora promote infection on Hevea brasiliensis. Phytopathology 2019, 109, 1769–1778. [Google Scholar] [CrossRef]
- Pettongkhao, S.; Navet, N.; Schornack, S.; Tian, M.; Churngchow, N. A secreted protein of 15 kDa plays an important role in Phytophthora palmivora development and pathogenicity. Sci. Rep. 2020, 10, 2319. [Google Scholar] [CrossRef]
- Ma, Z.; Song, T.; Zhu, L.; Ye, W.; Wang, Y.; Shao, Y.; Dong, S.; Zhang, Z.; Dou, D.; Zheng, X.; et al. A Phytophthora sojae glycoside hydrolase 12 protein is a major virulence factor during soybean infection and is recognized as a PAMP. Plant Cell 2015, 27, 2057–2072. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Zhu, L.; Song, T.; Wang, Y.; Zhang, Q.; Xia, Y.; Qiu, M.; Lin, Y.; Li, H.; Kong, L.; et al. A paralogous decoy protects Phytophthora sojae apoplastic effector PsXEG1 from a host inhibitor. Science 2017, 355, 710–714. [Google Scholar] [CrossRef]
- Zerillo, M.M.; Adhikari, B.N.; Hamilton, J.P.; Buell, C.R.; Lévesque, C.A.; Tisserat, N. Carbohydrate-active enzymes in Pythium and their role in plant cell wall and storage polysaccharide degradation. PLoS ONE 2013, 8, e72572. [Google Scholar] [CrossRef]
- Ochola, S.; Huang, J.; Ali, H.; Shu, H.; Shen, D.; Qiu, M.; Wang, L.; Li, X.; Chen, H.; Kange, A.; et al. Editing of an effector gene promoter sequence impacts plant-Phytophthora interaction. J. Integr. Plant Biol. 2020, 62, 378–392. [Google Scholar] [CrossRef] [Green Version]
- Brunner, F.; Rosahl, S.; Lee, J.; Rudd, J.J.; Geiler, C.; Kauppinen, S.; Rasmussen, G.; Scheel, D.; Nürnberger, T. Pep-13, a plant defense-inducing pathogen-associated pattern from Phytophthora transglutaminases. EMBO J. 2002, 21, 6681–6688. [Google Scholar] [CrossRef] [PubMed]
- Griffin, M.; Casadio, R.; Bergamini, C.M. Transglutaminases: Nature’s biological glues. Biochem. J. 2002, 368, 377–396. [Google Scholar] [CrossRef] [Green Version]
- Grenville-Briggs, L.; Avrova, A.; Hay, R.; Bruce, C.; Whisson, S.; van West, P. Identification of appressorial and mycelial cell wall proteins and a survey of the membrane proteome of Phytophthora infestans. Fungal Biol. 2010, 114, 702–723. [Google Scholar] [CrossRef]
- Nietzschmann, L.; Gorzolka, K.; Smolka, U.; Matern, A.; Eschen-Lippold, L.; Scheel, D.; Rosahl, S. Early Pep-13-induced immune responses are SERK3A/B-dependent in potato. Sci. Rep. 2019, 9, 18380. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; He, H.; Qi, Y.; McLellan, H.; Tian, Z.; Birch, P.; Tian, Z. The oomycete microbe-associated molecular pattern Pep-13 triggers SERK3/BAK1-independent plant immunity. Plant Cell Rep. 2019, 38, 173–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Verzaux, E.; Chaparro-Garcia, A.; Bijsterbosch, G.; Keizer, L.; Zhou, J.; Liebrand, T.; Xie, C.; Govers, F.; Robatzek, S.; et al. Elicitin recognition confers enhanced resistance to Phytophthora infestans in potato. Nat. Plants 2015, 1, 15034. [Google Scholar] [CrossRef] [PubMed]
- Evangelisti, E.; Govetto, B.; Minet-Kebdani, N.; Kuhn, M.L.; Attard, A.; Ponchet, M.; Panabières, F.; Gourgues, M. The Phytophthora parasitica RXLR effector penetration-specific effector 1 favours Arabidopsis thaliana infection by interfering with auxin physiology. New Phytol. 2013, 199, 476–489. [Google Scholar] [CrossRef]
- Morales-Cruz, A.; Ali, S.; Minio, A.; Figueroa-Balderas, R.; García, J.; Kasuga, T.; Puig, A.; Marelli, J.; Bailey, B.; Cantu, D. Independent whole-genome duplications define the architecture of the genomes of the devastating West African cacao black pod pathogen and its close relative. G3 (Bethesda Md.) 2020, 10, 2241–2255. [Google Scholar] [CrossRef]
- Wickramasuriya, A.M.; Dunwell, J.M. Cacao biotechnology: Current status and future prospects. Plant Biotechnol. J. 2018, 16, 4–17. [Google Scholar] [CrossRef]
- Bekele, F.; Bidaisee, G.; Singh, H.; Duraisamy, S. Morphological characterisation and evaluation of cacao (Theobroma cacao L.) in Trinidad to facilitate utilisation of Trinitario cacao globally. Genet. Resour. Crop. Evol. 2020, 67, 621–643. [Google Scholar] [CrossRef]
- Castro-Alayo, E.M.; Idrogo-Vásquez, G.; Siche, R.; Cardenas-Toro, F.P. Formation of aromatic compounds precursors during fermentation of Criollo and Forastero cocoa. Heliyon 2019, 5, e01157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thevenin, J.-M.; Rossi, V.; Ducamp, M.; Doare, F.; Condina, V.; Lachenaud, P. Numerous clones resistant to Phytophthora palmivora in the ‘‘Guiana’’ genetic group of Theobroma cacao L. PLoS ONE 2012, 7, e40915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwaro, A.D.; Sreenivasan, T.N.; Umahara, P. Phytophthora resistance in cacao (Theobroma cacao): Influence of pod morphological characteristics. Plant Pathol. 1997, 46, 557–565. [Google Scholar] [CrossRef]
- Iwaro, A.D.; Sreenivasan, T.N.; Umaharan, P. Foliar resistance to Phytophthora palmivora as an indicator of pod resistance in Theobroma cacao. Plant Dis. 1997, 81, 619–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appiah, A.; Opoku, I.; Akrofi, A. Natural occurrence and distribution of stem cankers caused by Phytophthora megakarya and Phytophthora palmivora on cocoa. Eur. J. Plant Pathol. 2004, 110, 983–990. [Google Scholar] [CrossRef]
- Vernon, A.J. Canker: The forgotten disease of cocoa. Cocoa Grow. Bull. 1971, 16, 9–14. [Google Scholar]
- Purwantara, A.; McMahon, P.; Susilo, A.W.; Sukamto, S.; Mulia, S.; Nurlaila; Saftar, A.; bin Purung, H.; Lambert, S.; Keane, P.; et al. Testing local cocoa selections in Sulawesi: (ii) resistance to stem canker and pod rot (black pod) caused by Phytophthora palmivora. Crop. Prot. 2015, 77, 18–26. [Google Scholar] [CrossRef]
- Okey, E.N.; Duncan, E.J.; Sirju-Charran, G.; Screenivasan, T.N. Factors affecting the susceptibility of six cocoa clones to Phytophthora palmivora (Butl) Butler bark canker in Trinidad. Plant Pathol. 1996, 45, 84–91. [Google Scholar] [CrossRef]
- Nyadanu, D.; Akromah, R.; Adomako, B.; Akrofi, A.; Dzahini-Obiatey, H.; Lowor, S.; Atta, O.; Kwoseh, C.; Awuah, R.; Adu-Dapaah, H.; et al. Genetic control, combining ability and heritability of resistance to stem canker in cacao (Theobroma cacao L.). Euphytica 2017, 213, 263. [Google Scholar] [CrossRef]
- Okey, E.N.; Duncan, E.J.; Sirju-Charran, G.; Sreenivasan, T.N. Wound-healing in cocoa (Theobroma cacao L.) stems and its effect on canker caused by phytophthora palmivora (Butl.) Butler. Int. J. Pest. Manag. 1995, 41, 224–228. [Google Scholar] [CrossRef]
- Okey, E.; Duncan, E.; Sirju-Charran, G.; Sreenivasan, T. Phytophthora canker resistance in cacao: Role of peroxidase, polyphenoloxidase and phenylalanine ammonia-lyase. J. Phytopathol. 1997, 145, 295–299. [Google Scholar] [CrossRef]
- Egea, C.; Ahmed, A.; Candela, M.; Candela, M. Elicitation of peroxidase activity and lignin biosynthesis in pepper suspension cells by Phytophthora capsici. J. Plant Physiol. 2001, 158, 151–158. [Google Scholar] [CrossRef]
- McMahon, P.; Purwantara, A.; Wahab, A.; Imron, M.; Lambert, S.; Keane, P.; Guest, D. Phosphonate applied by trunk injection controls stem canker and decreases Phytophthora pod rot (black pod) incidence in cocoa in Sulawesi. Australas. Plant Pathol. 2010, 39, 170–175. [Google Scholar] [CrossRef]
- Asomaning, E.J.A. Varietal resistance of young clones and seedlings of cocoa (Theobroma cacao L.) to root infection by Phytophthora palmivora. Trop. Agric. (Trinidad Tobago) 1964, 41, 251–256. [Google Scholar]
- Jackson, G.V.H.; Newhook, F.J. Sources of Phytophthora palmivora inoculum in Solomon Island cocoa plantations. Trans. Brit. Mycol. Soc. 1978, 71, 239–249. [Google Scholar] [CrossRef]
- Turner, P.D.; Asomaning, E.J.A. Root infection of Theobroma cacao by Phytophthora palmivora. Trop. Agric. Trin. 1962, 39, 339–343. [Google Scholar]
- Opoku, I.Y.; Wheeler, B.E.J. Survival of Phytophthora palmivora and Phytophthora megakarya on and in roots of cocoa seedlings. Cocoa Grow. Bull. 1999, 51, 33–41. [Google Scholar]
- Agrios, G.N. Plant. Pathology, 5th ed.; Elsevier Academic Press: Boston, MA, USA, 2015. [Google Scholar]
- Alston, D.G.; Sipes, B.S.; Uchida, J.; Schmitt, D.P.; Chia, C.L. Interactive effects of Rotylenchulus reniformis and Phytophthora palmivora on papaya (Carica papaya L.) survival and growth in greenhouse pots. Nematropica 2003, 33, 73–85. [Google Scholar]
- Jonathan, E.I.; Gajendran, G.; Arulmozhiyan, R. Interaction of Rotylenchulus reniformis and Phytophthora palmivora on betelvine. Nematol. Mediterr. 1997, 25, 191–194. [Google Scholar]
- Jonathan, E.I.; Sivakumar, M.; Padmanaban, D. Interaction of Meloidogyne incognita and Phytophthora palmivora on betelvine. Nematol. Mediterr. 1996, 24, 341–343. [Google Scholar]
- Saul Maora, J.; Liew, E.; Guest, D. Limited morphological, physiological and genetic diversity of Phytophthora palmivora from cocoa in Papua New Guinea. Plant Pathol. 2017, 66, 124–130. [Google Scholar] [CrossRef]
- Masanto, A.; Hieno, A.; Wibowo, S.; Subandiyah, M.; Shimizu, H.; Suga, K.; Kageyama, K. Genetic diversity of Phytophthora palmivora isolates from Indonesia and Japan using rep-PCR and microsatellite markers. J. Gen. Plant Pathol. 2019, 85, 367–381. [Google Scholar] [CrossRef]
- Appiah, A.A.; Flood, J.; Bridge, P.D.; Archer, S.A. Inter- and intraspecific morphometric variation and characterization of Phytophthora isolates from cocoa. Plant Pathol. 2003, 52, 168–180. [Google Scholar] [CrossRef]
- Opoku, I.Y.; Appiah, A.A.; Akrofi, A.Y.; Owusu, G.K. Phytophthora megakarya: A potential threat to the cocoa industry in Ghana. Ghana J. Agric. Sci. 2000, 33, 237–248. [Google Scholar] [CrossRef]
- Puig, A.; Ali, S.; Strem, M.; Sicher, R.; Gutierrez, O.; Bailey, B. The differential influence of temperature on Phytophthora megakarya and Phytophthora palmivora pod lesion expansion, mycelia growth, gene expression, and metabolite profiles. Physiol. Mol. Plant Pathol. 2018, 102, 95–112. [Google Scholar] [CrossRef]
- Derevnina, L.; Benjamin, P.; Kellner, R.; Dagdas, Y.; Sarowar, M.; Giannakopoulou, A.; De la Concepcion, J.; Concepcion, D.; Chaparro-Garcia, A.; Pennington, H.; et al. Emerging oomycete threats to plants and animals. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150459. [Google Scholar] [CrossRef] [Green Version]
- Gavino, P.D.; Smart, C.D.; Sandrock, R.W.; Miller, J.S.; Hamm, P.B.; Lee, T.Y.; Davis, R.M.; Fry, W.E. Implications of sexual reproduction for Phytophthora infestans in the United States: Generation of an aggressive lineage. Plant Dis. 2000, 84, 731–735. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, S.B. The population genetics of Phytophthora. Phytopathology 1997, 87, 462–473. [Google Scholar] [CrossRef] [Green Version]
- Brasier, C. Evolutionary biology of Phytophthora. Annu. Rev. Phytopathol. 1992, 30, 153–170. [Google Scholar] [CrossRef]
- Brasier, C.; Hansen, E. Evolutionary Biology of Phytophthora Part II: Phylogeny, speciation, and population Structure. Annu. Rev. Phytopathol. 1992, 30, 173–200. [Google Scholar] [CrossRef]
Figure 1.
The characteristic morphology of Phytophthora palmivora. These Ppal isolates MAG14, NSP11 and NSP19 were from infected cocoa pods from three different farms in Madang and Bougainville (PNG) in 2005 [13]. NSP11 and NSP19 were from two different districts, Sinai and Buin, respectively in Bougainville. (A) Papillate sporangium (mature); (B) terminal chlamydospore forming new hyphal extension; (C) intercalary chlamydospore; (D–F) release of zoospores within 1 min time period in one sporangium (black asterisks); (G) trapped zoospores; (H) trapped cysts germinating within a sporangium; (I) germinating cyst on a glass microscope slide; (J) sympodial branching of sporangiophore with papillate sporangia stained with lactophenol blue; (K) DAPI (4′,6-diamidino-2-phenylindole) staining of nuclei in hyphae, zoospores within sporangia and chlamydospores; (L) to (N) Stellate/striate to radiant colony types of MAG14 (l), NSP11 (M) and NSP19 (N) isolates on carrot agar post 7 d growth at 26 °C respectively. Black arrowheads highlight the presence of flagella in (F,G). (A,D–G,I–K) isolate MAG14 on carrot agar post 6 d growth at 26 °C; (B,C,H) isolate NSP11 on oatmeal agar post 12 d growth at 26 °C. Micrographs (A–K) were captured using an Olympus BX51 microscope equipped with DP74 Olympus camera under differential interference contrast (DIC) and ultraviolet (UV) fluorescence.
Figure 1.
The characteristic morphology of Phytophthora palmivora. These Ppal isolates MAG14, NSP11 and NSP19 were from infected cocoa pods from three different farms in Madang and Bougainville (PNG) in 2005 [13]. NSP11 and NSP19 were from two different districts, Sinai and Buin, respectively in Bougainville. (A) Papillate sporangium (mature); (B) terminal chlamydospore forming new hyphal extension; (C) intercalary chlamydospore; (D–F) release of zoospores within 1 min time period in one sporangium (black asterisks); (G) trapped zoospores; (H) trapped cysts germinating within a sporangium; (I) germinating cyst on a glass microscope slide; (J) sympodial branching of sporangiophore with papillate sporangia stained with lactophenol blue; (K) DAPI (4′,6-diamidino-2-phenylindole) staining of nuclei in hyphae, zoospores within sporangia and chlamydospores; (L) to (N) Stellate/striate to radiant colony types of MAG14 (l), NSP11 (M) and NSP19 (N) isolates on carrot agar post 7 d growth at 26 °C respectively. Black arrowheads highlight the presence of flagella in (F,G). (A,D–G,I–K) isolate MAG14 on carrot agar post 6 d growth at 26 °C; (B,C,H) isolate NSP11 on oatmeal agar post 12 d growth at 26 °C. Micrographs (A–K) were captured using an Olympus BX51 microscope equipped with DP74 Olympus camera under differential interference contrast (DIC) and ultraviolet (UV) fluorescence.

Figure 2.
Schematic diagram of the infection process of Phytophthora palmivora in planta. (A) Via splashing of water droplets, flowing surface water, wind-driven rain, zoospores are released from the caducous sporangia and they actively swim towards potential infection sites on both aerial and subterranean surfaces of plants; (B) Once at the site, zoospores lose motility by shedding their flagella, encyst, and the newly formed wall adheres to the surface of the plant; (C) Cyst germinates forming a germ tube and later an appressorium, which provides stronger adhesion to the host surface in preparation for subsequent invasion into the epidermis of some aerial tissues. In ground tissues, the germ tube can penetrate the root epidermis by growing intercellularly along the anticlinal cell walls; (D) At the appressorium adhesion site, P. palmivora hyphae grow to invade intracellularly, forming a haustorium—this is the biotrophic stage; (E) During the necrotrophic stage, secondary hyphae form that kill the host cell; and (F) new structure characteristic of the oomycete i.e., intercalary and terminal chlamydospores and sporangia develop, providing new inocula for future infection of other regions on the same host or other hosts in the field.
Figure 2.
Schematic diagram of the infection process of Phytophthora palmivora in planta. (A) Via splashing of water droplets, flowing surface water, wind-driven rain, zoospores are released from the caducous sporangia and they actively swim towards potential infection sites on both aerial and subterranean surfaces of plants; (B) Once at the site, zoospores lose motility by shedding their flagella, encyst, and the newly formed wall adheres to the surface of the plant; (C) Cyst germinates forming a germ tube and later an appressorium, which provides stronger adhesion to the host surface in preparation for subsequent invasion into the epidermis of some aerial tissues. In ground tissues, the germ tube can penetrate the root epidermis by growing intercellularly along the anticlinal cell walls; (D) At the appressorium adhesion site, P. palmivora hyphae grow to invade intracellularly, forming a haustorium—this is the biotrophic stage; (E) During the necrotrophic stage, secondary hyphae form that kill the host cell; and (F) new structure characteristic of the oomycete i.e., intercalary and terminal chlamydospores and sporangia develop, providing new inocula for future infection of other regions on the same host or other hosts in the field.

Figure 3.
Theobroma cacao L. (cocoa) plants (undefined mixture of Trinitario) in the glasshouse. (A) Dimorphous cocoa plant; (B) Unopened cauliflorous flowers at different stages of growth and development attached to swollen and enlarged regions of the trunk known as flower cushions (white arrows); (C) Detached cauliflorous, reddish-white color, odorless and self-fertile flower; (D) a sucker or chupon near the base of the trunk; (E) cherelles (immature pods) at different stages of growth and development arising from former flower cushions and (F) close-up of an immature pod. White asterisk (*) in (E) shows the same cherelle in (F) after 1 month aborted in growth. Scale bar in (E) represents 5 cm.
Figure 3.
Theobroma cacao L. (cocoa) plants (undefined mixture of Trinitario) in the glasshouse. (A) Dimorphous cocoa plant; (B) Unopened cauliflorous flowers at different stages of growth and development attached to swollen and enlarged regions of the trunk known as flower cushions (white arrows); (C) Detached cauliflorous, reddish-white color, odorless and self-fertile flower; (D) a sucker or chupon near the base of the trunk; (E) cherelles (immature pods) at different stages of growth and development arising from former flower cushions and (F) close-up of an immature pod. White asterisk (*) in (E) shows the same cherelle in (F) after 1 month aborted in growth. Scale bar in (E) represents 5 cm.


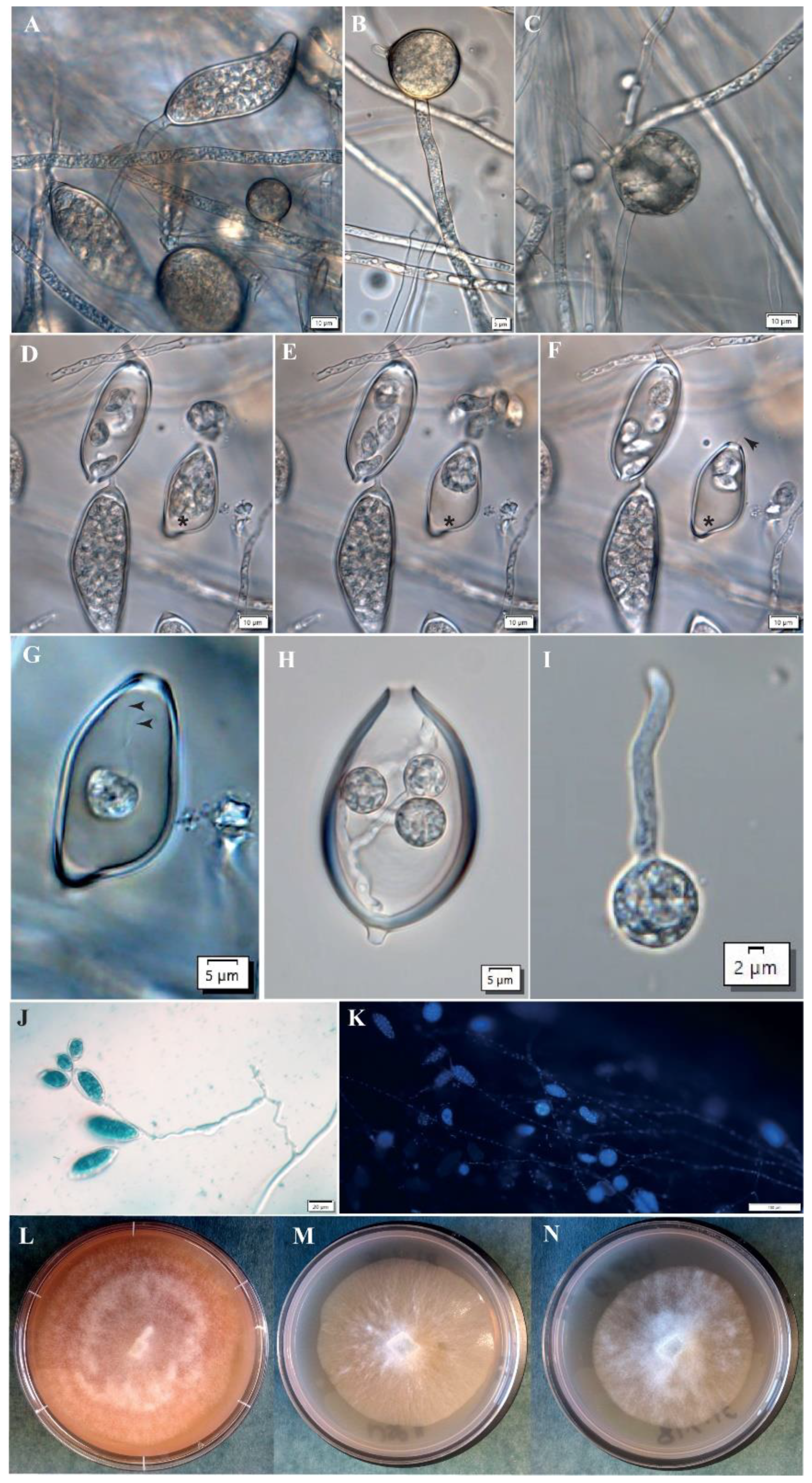{kind=link}
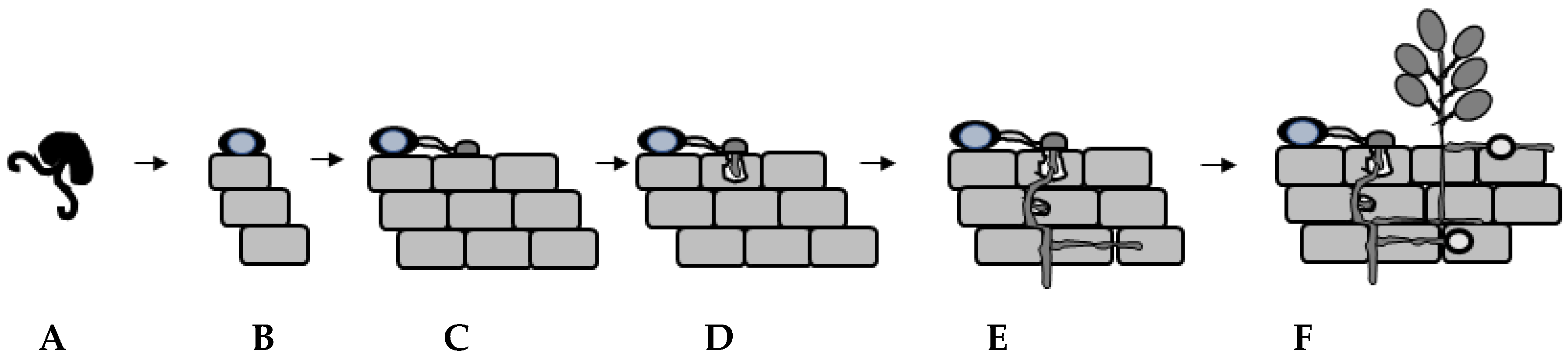{kind=link}
{kind=link}
Table 1.
List of Phytophthora species known to cause black pod in cocoa and characteristics related to their geographical distributions, host range, clade, sex, and genome size.
Table 1.
List of Phytophthora species known to cause black pod in cocoa and characteristics related to their geographical distributions, host range, clade, sex, and genome size.
| Species Name a | Geographical Distribution a | Clade b | Host b | Sex b | Papil. b | Genome Size (Mb) |
|---|---|---|---|---|---|---|
| Phytophthora capsici (Leonian) | Brazil, El Salvador, Guatemala, India, Jamaica, Mexico, Trinidad, Venezuela | 2 | Multiple | He | P | 64.00 [7] |
| P. citrophthora (RE Smith and EH Smith) | Brazil, India, Mexico | 2 | Multiple | He | P | n.d |
| P. heveae (Thompson) | Malaysia | 5 | Multiple | Ho | P | n.d |
| P. megakarya (Brasier and Griffin) | Cameroon, Côte d’Ivoire, Fernando Po (aka Bioko), Gabon, Ghana, Nigeria, São Tomé (islands of Principe and São Tomé), and Togo | 4 | T. cacao | He | P | 126.88 [8] |
| P. megasperma (Dreschler) | Venezuela | 6 | Multiple | Ho | NP | 62 [9] |
| P. nicotianae var. parasitica | Cuba | 1 | Multiple | He | P | 76.50 [2] |
| P. palmivora (Butler) | Pantropical | 4 | Multiple | He | P | 151.23 [10] |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Perrine-Walker, F. Phytophthora palmivora–Cocoa Interaction. J. Fungi 2020, 6, 167. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030167
AMA Style
Perrine-Walker F. Phytophthora palmivora–Cocoa Interaction. Journal of Fungi. 2020; 6(3):167. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030167
Chicago/Turabian StylePerrine-Walker, Francine. 2020. "Phytophthora palmivora–Cocoa Interaction" Journal of Fungi 6, no. 3: 167. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030167
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.