Characterization of Five New Monosporascus Species: Adaptation to Environmental Factors, Pathogenicity to Cucurbits and Sensitivity to Fungicides

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Monosporascus spp. Isolates
2.2. Effect of pH on the Mycelial Growth Rate of Monosporascus spp.
2.3. Effect of Salinity on the Mycelial Growth of Monosporascus spp.
2.4. Pathogenicity of Monosporascus spp. to Cucurbits
2.5. Sensitivity of Monosporascus spp. to Fungicides
3. Results
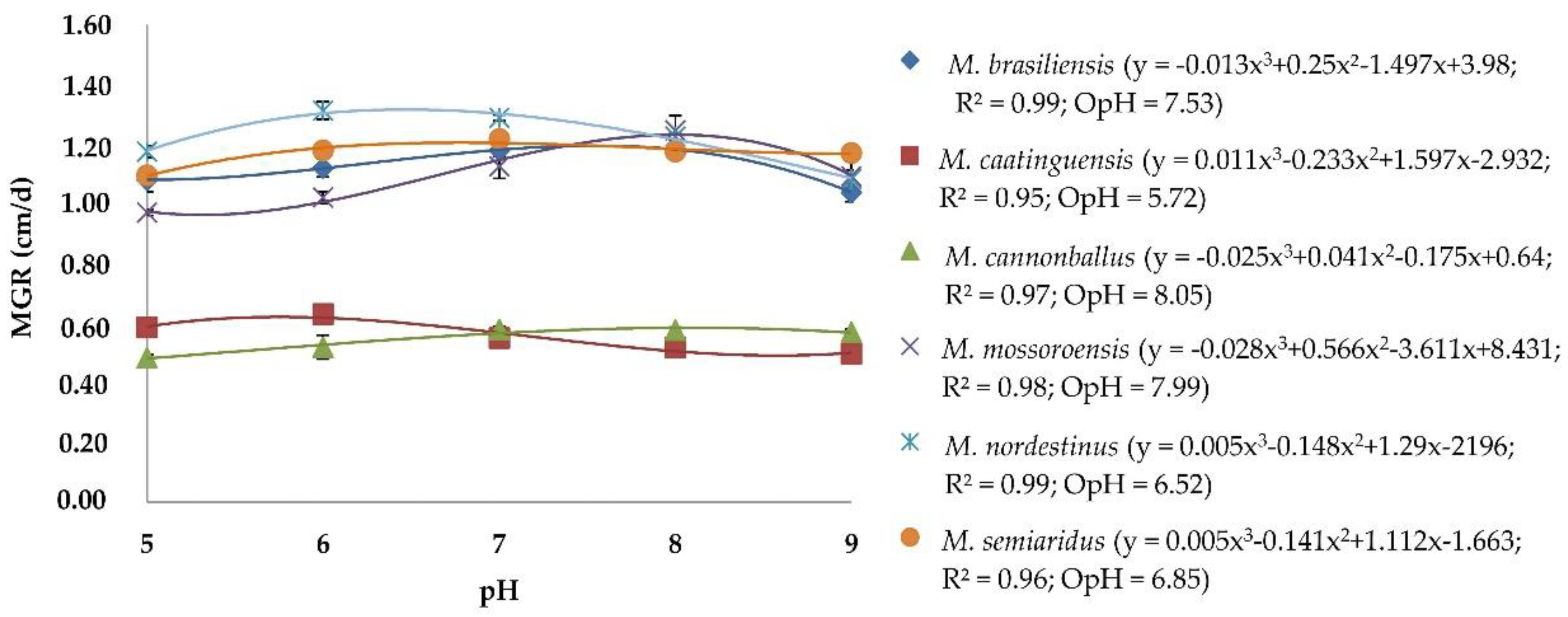
3.1. Effect of pH on Mycelial Growth of Monosporascus spp.
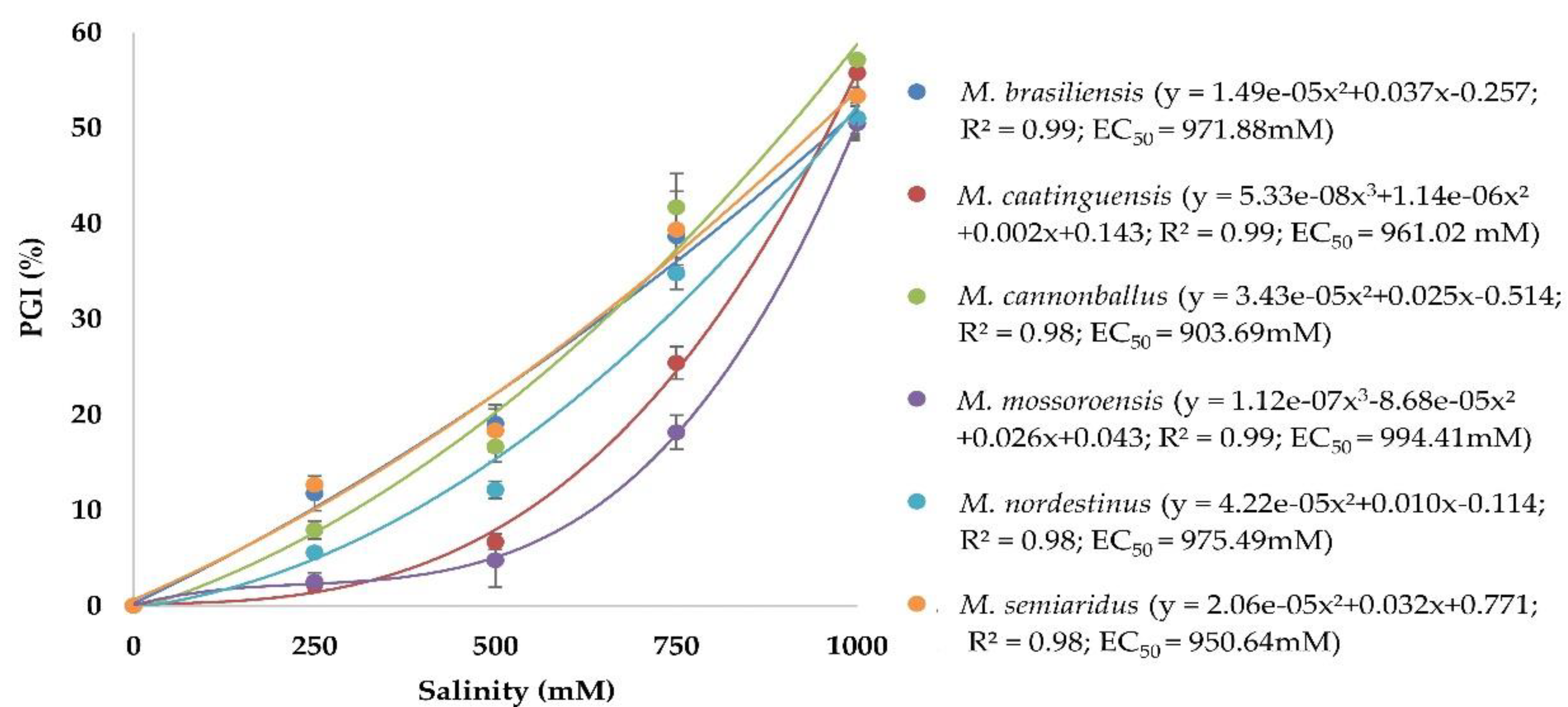
3.2. Effect of Salinity on Mycelial Growth of Monosporascus spp.
3.3. Pathogenicity of Monosporascus spp. to Cucurbits
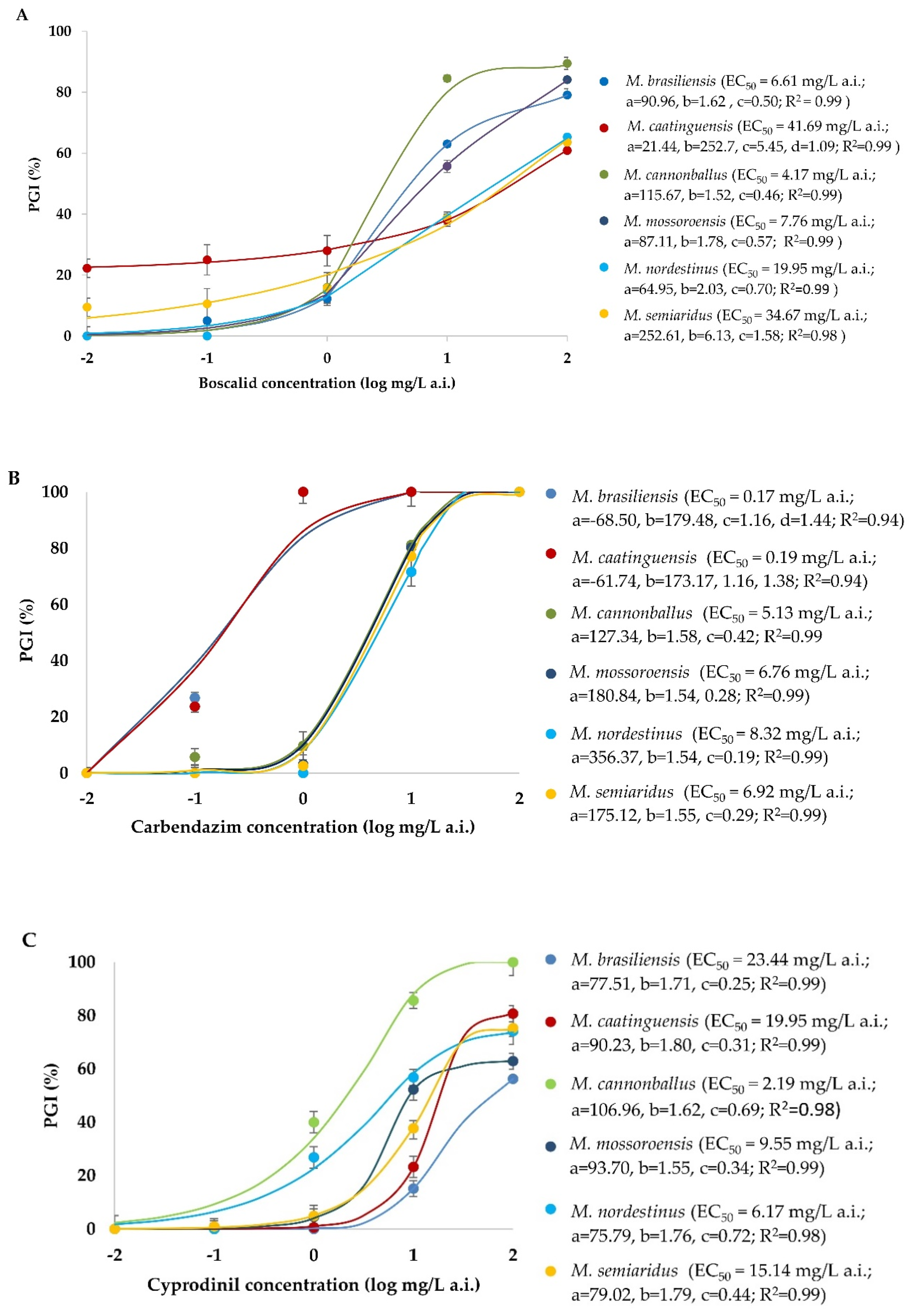
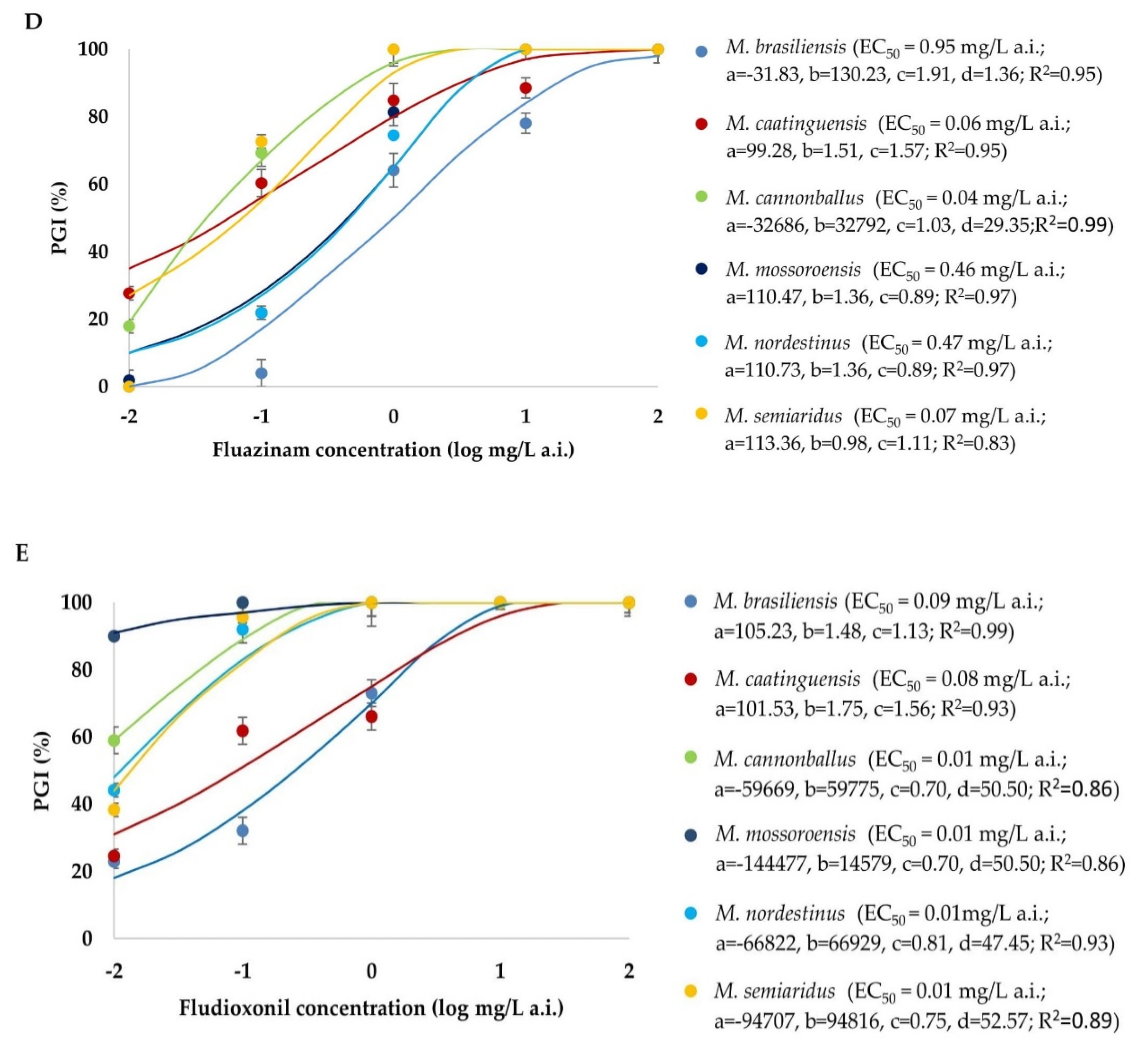
3.4. Sensitivity of Monosporascus spp. to Fungicides
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Martyn, R.D.; Miller, M.E. Monosporascus root rot and vine decline: An emerging disease of melons worldwide. Plant Dis. 1996, 80, 716–725. [Google Scholar] [CrossRef]
- Bruton, B.D. Soilborne diseases in cucurbitaceae: Pathogen virulence and host resistance. In Cucurbitaceae 98; McCreight, J., Ed.; International Society of Horticultural Science: Alexandria, Egypt, 1998; pp. 143–166. [Google Scholar]
- Cohen, R.; Pivonia, S.; Crosby, K.M.; Martyn, R.D. Advances in the biology and management of Monosporascus vine decline and wilt of melons and other cucurbits. Hortic. Rev. 2012, 39, 77–120. [Google Scholar] [CrossRef]
- Ben Salem, I.; Correia, K.C.; Boughalleb, N.; Michereff, S.J.; León, M.; Abad-Campos, P.; García-Jiménez, J.; Armengol, J. Monosporascus eutypoides, a cause of root rot and vine decline in Tunisia, and evidence that M. cannonballus and M. eutypoides are distinct species. Plant Dis. 2013, 97, 737–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sales Júnior, R.; Nascimento, I.J.B.; Freitas, L.S.; Beltrán, R.; Armengol, J.; Vicent, A.; García-Jiménez, J. First Report of Monosporascus cannonballus on melon in Brazil. Plant Dis. 2004, 88, 84. [Google Scholar] [CrossRef]
- Sales Júnior, R.; Santana, C.V.S.; Nogueira, D.R.S.; Silva, K.J.P.; Guimaraes, I.M.; Michereff, S.J.; Abad-Campos, P.; García-Jiménez, J.; Armengol, J. First Report of Monosporascus cannonballus on watermelon in Brazil. Plant Dis. 2010, 94, 278. [Google Scholar] [CrossRef]
- Al-Mawaali, Q.S.; Al-Sadi, A.M.; Al-Said, F.A.; Deadman, M.L. Etiology, development and reaction of muskmelon to vine decline under arid conditions of Oman. Phytopathol. Mediterr. 2013, 52, 457–465. [Google Scholar]
- Yan, L.Y.; Zang, Q.Y.; Huang, Y.P.; Wang, Y.H. First report of root rot and vine decline of melon caused by Monosporascus cannonballus in Easter Mainland China. Plant Dis. 2016, 100, 651. [Google Scholar] [CrossRef]
- Markakis, E.A.; Trantas, E.A.; Lagogianni, C.S.; Mpalantinaki, E.; Pagoulatou, M.; Ververidis, F.N.; Goumas, D.E. First report of root rot and vine decline of melon caused by Monosporascus cannonballus in Greece. Plant Dis. 2018, 102, 1036. [Google Scholar] [CrossRef]
- Negreiros, A.M.P.; Sales Júnior, R.; Rodrigues, A.P.M.S.; León, M.; Armengol, J. Prevalent weeds collected from cucurbit fields in Northeastern Brazil reveal new species diversity in the genus Monosporascus. Ann. Appl. Biol. 2019, 174, 349–363. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Herrera, J.; Sinsabaugh, R.L.; Odenbach, K.J.; Lowrey, T.; Natvig, D.O. Novel root fungal consortium associated with a dominant desert grass. Appl. Environ. Microbiol. 2008, 74, 2805–2813. [Google Scholar] [CrossRef] [Green Version]
- Herrera, J.; Khidir, H.H.; Eudy, D.M.; Porras-Alfaro, A.; Natvig, D.O.; Sinsabaugh, R.L. Shifting fungal endophyte communities colonize Bouteloua gracilis: Effect of host tissue and geographical distribution. Mycologia 2010, 102, 1012–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, S.L.; Warnock, D.D.; Litvak, M.E.; Porras-Alfaro, A.; Sinsabaugh, R.L. Root-associated fungal community response to drought-associated changes in vegetation community. Mycologia 2015, 107, 1089–1104. [Google Scholar] [CrossRef] [PubMed]
- Robinson, A.J.; Natvig, D.O.; Chain, P.S.G. Genomic analysis of diverse members of the fungal genus Monosporascus reveals novel lineages, unique genome content and a potential bacterial associate. G3-Genes Genom. Genet. 2020, 10, 2573–2583. [Google Scholar] [CrossRef]
- Kwon, M.K.; Hong, J.R.; Kim, Y.H.; Kim, K.C. Soil-environment factors involved in the development of root rot/vine decline of cucurbits caused by Monosporascus cannonballus. Plant Pathol. J. 2001, 17, 45–51. [Google Scholar]
- Martyn, R.D. Monosporascus root rot and vine decline of melons. Plant Health Instr. 2002. [Google Scholar] [CrossRef]
- Pivonia, S.; Cohen, R.; Kigel, J.; Katan, J. Effect of soil temperature on disease development in melon plants infected by Monosporascus cannonballus. Plant Pathol. 2002, 51, 472–479. [Google Scholar] [CrossRef]
- Waugh, M.M.; Kim, D.H.; Ferrin, D.M.; Stanghellini, M.E. Reproductive potential of Monosporascus cannonballus. Plant Dis. 2003, 87, 45–50. [Google Scholar] [CrossRef]
- Yasuaki, M.; Naoyuki, T.; Yoichi, K. The effect of temperature conditions on the death of Monosporascus cannonballus. Bull. Kochi Agr. Res. Center 2005, 14, 1–4. [Google Scholar]
- Hamza, H.; Belkadhi, M.S.; Triki, M.A.; Zouba, A. Morphological and biological studies of Monosporascus cannonballus, the cause of root rot and vine decline of melon in Southern Tunisia. Tunis. J. Plant Prot. 2007, 2, 71–77. [Google Scholar]
- Senhor, R.F.; Souza, P.A.; Andrade Neto, R.C.; Pinto, A.C.; Soares, S.R.F. Colapso do meloeiro associado a Monosporascus cannonballus. Rev. Verde 2009, 4, 6–14. [Google Scholar]
- Silva, K.J.P.; Cordeito, A.G.; Nogueira, D.R.S.; Sales Júnior, R. Monosporascus cannonballus: Agente causal do colapso ou morte súbita do meloeiro. Rev. Verde 2010, 5, 11–18. [Google Scholar]
- Armengol, J.; Alaniz, S.; Vicent, A.; Beltrán, R.; Abad-Campos, P.; Pérez-Sierra, A.; García-Jiménez, J.; Ben Salem, I.; Souli, M.; Boughalleb-M’Hamdi, N. Effect of dsRNA on growth rate and reproductive potential of Monosporascus cannonballus. Fungal Biol. 2011, 115, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Ben Salem, I.; Boughalleb-M’Hamdi, N.; Souli, M.; Cherif, M. Morphological and biological characterization of Monosporascus cannonballus isolates, responsible of watermelon decline in Kairouan’s area. Res. Plant Biol. 2011, 3, 28–37. [Google Scholar]
- Correia, K.C.; Silva, E.K.C.; Câmara, M.P.S.; Sales Júnior, R.; Mizubuti, E.S.G.; Armengol, J.; García-Jiménez, J.; Michereff, S.J. Fitness components of Monosporascus cannonballus isolates from northeastern Brazilian melon fields. Trop. Plant Pathol. 2014, 39, 217–223. [Google Scholar] [CrossRef]
- Rhouma, A.; Ben Salem, I.; M’hamdi, M.; Boughalleb-M’hamdi, N. Relationship study among soils physico-chemical properties and Monosporascus cannonballus ascospores densities for cucurbit fields in Tunisia. Eur. J. Plant Pathol. 2019, 153, 97–110. [Google Scholar] [CrossRef]
- Deacon, J. Fungal Biology, 4th ed.; Blackwell Publishing Ltd.: Oxford, UK, 2006; p. 371. [Google Scholar]
- Mertely, J.C.; Martyn, R.D.; Miller, M.E.; Bruton, B.D. An expanded host range for the muskmelon pathogen, Monosporascus cannonballus. Plant Dis. 1993, 77, 667–673. [Google Scholar] [CrossRef]
- Sales Júnior, R.; Balbino, D.A.D.; Negreiros, A.M.P.; Barboza, H.S.; Medeiros, E.V.; Armengol, J. Cotton, cowpea and sesame are alternative crops to cucurbits in soils naturally infested with Monosporascus cannonballus. J. Phytopathol. 2018, 166, 396–402. [Google Scholar] [CrossRef]
- Medeiros, E.V.; Silva, K.J.P.; Oliveira, L.A.; Ferreira, H.A.; Sales Júnior, R. Monosporascus cannonballus density in soils cultivated with different crops in Rio Grande do Norte State, Brazil. Rev. Bras. Ciênc. Agrár. 2008, 3, 1–5. [Google Scholar]
- Pivonia, S.; Gerstl, Z.; Maduel, A.; Levita, R.; Cohen, R. Management of Monosporascus sudden wilt of melon by soil application of fungicides. Eur. J. Plant Pathol. 2010, 128, 201–209. [Google Scholar] [CrossRef]
- Awad, H.M. Evaluation of plant extracts and essential oils for the control of sudden wilt disease of watermelon plants. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 949–962. [Google Scholar] [CrossRef]
- Sales Júnior, R.; Senhor, R.F.; Michereff, S.J.; Medeiros, E.V. Influência da adubação verde na severidade do declínio-de-Monosporascus em solo naturalmente infestado. Hortic. Bras. 2017, 35, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Cohen, R.; Pivonia, S.; Shtienberg, D.; Edelstein, M.; Raz, D.; Gerstl, Z. Efficacy of fluazinam in suppression of Monosporascus cannonballus, the causal agent of vine decline of melons. Plant Dis. 1999, 83, 1137–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimarães, I.M.; Sales Júnior, R.; Silva, K.J.P.; Michereff, S.J.; Nogueira, D.R.S. Efeito de fluazinam no controle Monosporascus cannonballus, agente causal do declínio de ramas em meloeiro. Rev. Caatinga 2008, 21, 147–153. [Google Scholar]
- Cervantes-García, D.C.; Ramirez, J.S.P.; Simpson, J.; Mayek-Perez, J. Osmotic potential effects on in vitro growth, morphology and pathogenicity of Macrophomina phaseolina. J. Phytopathol. 2003, 151, 456–462. [Google Scholar] [CrossRef]
- Ben Salem, I.; Armengol, J.; Berbegel, M.; Boughalleb-M’Hamdi, N. Development of a screening test for resistance of cucurbits and Cucurbita hybrid rootstocks to Monosporascus cannonballus. Tunis. J. Plant Prot. 2015, 10, 23–33. [Google Scholar]
- Aegerter, B.J.; Gordon, T.R.; Davis, R.M. Occurrence and pathogenicity of fungi associated with melon root rot and vine decline in California. Plant Dis. 2000, 84, 224–230. [Google Scholar] [CrossRef] [Green Version]
- Silva, F.A.Z.; Azevedo, C.A.V. The Assistat Software Version 7.7 and its use in the analysis of experimental data. Afr. J. Agr. Res. 2016, 11, 3733–3740. [Google Scholar] [CrossRef] [Green Version]
- Tonin, R.F.B.; Avozani, A.; Danelli, A.L.D.; Reis, E.M.; Zoldan, S.M.; Garcés-Fiallos, F.R. In vitro mycelial sensitivity of Macrophomina phaseolina to fungicides. Pesq. Agropec. Trop. 2013, 43, 460–466. [Google Scholar] [CrossRef]
- Mendes, A.M.S.; Faria, C.M.B.; Terao, D.; Silva, D.J.; Batista, D.C.; Moreira, F.R.B.; Resende, G.M.; Alencar, J.A.; Oliveira, J.E.M.; Araújo, J.L.P.; et al. A Cultura do Melão, 2nd ed.; Embrapa Informação Tecnológica: Brasília, DF, Brasil, 2008; p. 191. [Google Scholar]
- Tijerina-Ramírez, N.; Lira-Méndez, K.; Moreno-Medina, V.R.; González-Prieto, J.M.; Mayek-Pérez, N. Efecto del estrés osmótico in vitro en el crecimiento, patogenicidad y producción de osmolitos en Macrophomina phaseolina. Rev. Mex. Mic. 2014, 39, 31–39. [Google Scholar]
- Negreiros, A.M.P.; Melo, N.J.A.; Medeiros, H.L.S.; Silva, F.H.A.; Armengol, J.; Sales Júnior, R. Characterization of adaptability components of Brazilian isolates of Macrophomina pseudophaseolina. J. Phytopathol. 2020, 1–10. [Google Scholar] [CrossRef]
- Thangavelu, V.; Tang, J.; Ryan, D.; Valix, M. Effect of saline stress on fungi metabolism and biological leaching of weathered saprolite ores. Miner. Eng. 2006, 19, 1266–1273. [Google Scholar] [CrossRef]
- Andrade, D.E.G.T.; Michereff, S.J.; Borges, M.A.S.; Araújo, I.B.; Sales Júnior, R. Influência da densidade de inóculo e de isolados de Monosporascus cannonballus na severidade do colapso do meloeiro. Summa Phytopathol. 2005, 31, 173–180. [Google Scholar]
- Castro, G.; Perpiñá, G.; Esteras, C.; Armengol, J.; Picó, B.; Pérez-de-Castro, A. Resistance in melon to Monosporascus cannonballus and M. eutypoides: Fungal pathogens associated with Monosporascus root rot and vine decline. Ann. Appl. Biol. 2020, 101–111. [Google Scholar] [CrossRef]
- Mertely, J.C.; Martyn, R.D.; Miller, M.E.; Bruton, B.D. Role of Monosporascus cannonballus and other fungi in a root rot/vine decline disease of muskmelon. Plant Dis. 1991, 77, 667–673. [Google Scholar] [CrossRef]
- Lovic, B.R.; Valadez, V.A.; Martyn, R.D.; Miller, M.E. Association of dsRNA with reduced aggressiveness and phenotypic variability in Monosporascus cannonballus. Phytopathology 1994, 84, 776. [Google Scholar] [CrossRef]
- Reis, E.M.; Reis, A.C.; Forcelini, C.A. Manual de Fungicidas: Guia para o Controle Químico de Doenças de Plantas, 6th ed.; UPF Editora: Passo Fundo, MG, Brasil, 2010; p. 226. [Google Scholar]
- Leite, I.C.H.L.; Lopes, U.P. Controle químico de patógenos radiculares. In Desafios do Manejo de Doenças Radiculares Causadas por Fungos; Lopes, U.P., Michereff, S.J., Eds.; UFRPE Editora: Recife, PE, Brasil, 2018; pp. 179–192. [Google Scholar]
- Edgington, L.V.; Khew, K.L.; Barrow, G.L. Fungitoxic spectrum of benzimidazole compounds. Phytopathology 1971, 61, 42–44. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monosporascus Species | Code (CMM) 1 | Host | Location 2 | GenBank ITS Region 3 |
|---|---|---|---|---|
| M. brasiliensis | 4839 | Trianthema portulacastrum | Brazil, RN | MG 735234 |
| M. caatinguensis | 4833 | Boerhavia diffusa | Brazil, CE | MG 735228 |
| M. cannonballus | 2429 | Cucumis melo | Brazil, RN | JQ 762366 |
| M. mossoroensis | 4857 | Trianthema portulacastrum | Brazil, RN | MG 735252 |
| M. nordestinus | 4846 | Trianthema portulacastrum | Brazil, RN | MG 735241 |
| M. semiaridus | 4830 | Trianthema portulacastrum | Brazil, CE | MG 735222 |
| Cucumber | Melon | |||||||
|---|---|---|---|---|---|---|---|---|
| Treatments | Disease Incidence | Disease Severity | Disease Incidence | Disease Severity | ||||
| Rank 1 | Mean (%) | Rank 1 | Mean (%) | Rank 1 | Mean (%) | Rank 1 | Mean (%) | |
| M. brasiliensis | 32.50 ab | 60 | 39.35 bc | 2.10 | 40.50 b | 100 | 41.90 b | 1.60 |
| M. caatinguensis | 46.50 b | 100 | 37.75 abc | 1.30 | 40.50 b | 100 | 36.75 b | 1.30 |
| M. cannonballus | 46.50 b | 100 | 42.55 bc | 1.60 | 40.50 b | 100 | 43.45 b | 1.60 |
| M. mossoroensis | 25.50 ab | 40 | 20.90 ab | 0.40 | 40.50 b | 100 | 43.45 b | 1.60 |
| M. nordestinus | 39.50 b | 80 | 39.00 bc | 1.60 | 40.50 b | 100 | 34.00 b | 1.20 |
| M. semiaridus | 46.50 b | 100 | 57.45 c | 3.30 | 40.50 b | 100 | 43.45 b | 1.60 |
| Control | 11.50 a | 0 | 11.50 a | 0.00 | 5.50 a | 0 | 5.50 a | 0.00 |
| χ2 | 39.73 | 35.87 | 69.00 | 32.57 | ||||
| Pumpkin | Watermelon | |||||||
| M. brasiliensis | 29.50 b | 60 | 25.60 ab | 0.60 | 40.50 b | 100 | 38.00 b | 1.70 |
| M. caatinguensis | 43.50 b | 100 | 47.65 bc | 1.60 | 40.50 b | 100 | 41.20 b | 1.70 |
| M. cannonballus | 43.50 b | 100 | 37.00 bc | 1.00 | 40.50 b | 100 | 42.80 b | 1.80 |
| M. mossoroensis | 43.50 b | 100 | 48.30 bc | 1.80 | 40.50 b | 100 | 44.80 b | 1.80 |
| M. nordestinus | 43.50 b | 100 | 50.15 c | 1.70 | 40.50 b | 100 | 44.40 b | 1.90 |
| M. semiaridus | 36.50 b | 80 | 31.30 abc | 0.80 | 40.50 b | 100 | 31.80 b | 1.30 |
| Control | 8.50 a | 0 | 8.50 a | 0.00 | 5.50 a | 0 | 5.50 a | 0.00 |
| χ2 | 46.64 | 42.23 | 69.00 | 31.66 | ||||
| Treatments | Cucumber | |||||
|---|---|---|---|---|---|---|
| RL 1 (cm) | FRW 2 (g) | DRW 3 (g) | SL 4 (cm) | FSW 5 (g) | DSW 6 (g) | |
| M. brasiliensis | 21.30 c | 14.24 ab | 0.37 b | 46.90 b | 27.47 a | 2.95 bc |
| M. caatinguensis | 35.40 a | 12.65 ab | 0.62 b | 60.80 a | 36.26 a | 4.88 a |
| M. cannonballus | 30.80 ab | 9.90 bc | 0.47 b | 64.40 a | 31.33 a | 4.37 ab |
| M. mossoroensis | 32.08 ab | 12.27 abc | 0.56 b | 62.90 a | 33.79 a | 4.03 ab |
| M. nordestinus | 24.60 bc | 10.50 bc | 0.44 b | 68.00 a | 29.84 a | 3.91 ab |
| M. semiaridus | 17.40 c | 7.94 c | 0.45 b | 28.55 c | 11.94 b | 2.00 c |
| Control | 39.80 a | 17.52 a | 1.00 a | 67.12 a | 37.19 a | 4.90 a |
| CV (%) | 23.78 | 27.04 | 37.67 | 16.37 | 28.72 | 30.39 |
| Melon | ||||||
| M. brasiliensis | 27.00 ab | 3.84 b | 0.28 b | 97.20 a | 46.51 a | 6.06 a |
| M. caatinguensis | 25.46 abc | 7.08 b | 0.36 b | 97.22 a | 46.73 a | 6.86 a |
| M. cannonballus | 19.44 c | 4.32 b | 0.35 b | 89.82 ab | 50.60 a | 6.49 a |
| M. mossoroensis | 24.70 abc | 5.68 b | 0.31 b | 73.80 b | 38.47 a | 5.21 a |
| M. nordestinus | 22.90 abc | 6.43 b | 0.40 b | 106.75 a | 51.72 a | 7.67 a |
| M. semiaridus | 19.90 bc | 4.91 b | 0.36 b | 85.00 ab | 47.31 a | 5.99 a |
| Control | 28.80 a | 12.44 a | 1.32 a | 108.00 a | 43.05 a | 6.12 a |
| CV (%) | 22.07 | 38.89 | 35.21 | 18.05 | 22.66 | 29.03 |
| Pumpkin | ||||||
| M. brasiliensis | 36.50 b | 11.48 bc | 0.67 b | 16.10 ab | 32.41 b | 3.63 c |
| M. caatinguensis | 37.80 b | 8.69 c | 0.66 b | 18.75 a | 38.24 ab | 4.76 ab |
| M. cannonballus | 35.73 b | 9.47 c | 0.67 b | 13.62 b | 32.00 b | 3.40 c |
| M. mossoroensis | 36.11 b | 10.70 bc | 0.69 b | 16.76 ab | 32.22 b | 3.82 bc |
| M. nordestinus | 31.78 b | 10.11 c | 0.66 b | 17.18 ab | 36.35 ab | 4.23 abc |
| M. semiaridus | 35.80 b | 15.11 b | 0.81 b | 17.80 ab | 34.40 b | 3.49 c |
| Control | 50.60 a | 20.02 a | 2.57 a | 18.70 a | 45.78 a | 4.91 a |
| CV (%) | 22.32 | 26.58 | 34.20 | 18.57 | 21.83 | 18.81 |
| Watermelon | ||||||
| M. brasiliensis | 25.40 b | 4.39 c | 0.30 b | 96.90 a | 32.72 a | 4.55 a |
| M. caatinguensis | 28.96 ab | 4.01 c | 0.27 b | 100.05 a | 31.04 a | 3.94 a |
| M. cannonballus | 26.50 ab | 3.94 c | 0.29 b | 102.80 a | 35.87 a | 5.04 a |
| M. mossoroensis | 27.50 ab | 4.17 c | 0.30 b | 109.24 a | 36.50 a | 4.45 a |
| M. nordestinus | 29.80 ab | 7.26 b | 0.36 b | 120.20 a | 38.58 a | 4.84 a |
| M. semiaridus | 28.85 ab | 5.50 bc | 0.38 b | 103.70 a | 37.48 a | 5.20 a |
| Control | 34.11 a | 11.67 a | 1.57 a | 103.08 a | 36.44 a | 5.25 a |
| CV (%) | 19.62 | 35.32 | 36.64 | 31.14 | 33.00 | 20.85 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavalcante, A.L.A.; Negreiros, A.M.P.; Tavares, M.B.; Barreto, É.d.S.; Armengol, J.; Sales Júnior, R. Characterization of Five New Monosporascus Species: Adaptation to Environmental Factors, Pathogenicity to Cucurbits and Sensitivity to Fungicides. J. Fungi 2020, 6, 169. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030169
Cavalcante ALA, Negreiros AMP, Tavares MB, Barreto ÉdS, Armengol J, Sales Júnior R. Characterization of Five New Monosporascus Species: Adaptation to Environmental Factors, Pathogenicity to Cucurbits and Sensitivity to Fungicides. Journal of Fungi. 2020; 6(3):169. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030169
Chicago/Turabian StyleCavalcante, Allinny Luzia Alves, Andréia Mitsa Paiva Negreiros, Moisés Bento Tavares, Érica dos Santos Barreto, Josep Armengol, and Rui Sales Júnior. 2020. "Characterization of Five New Monosporascus Species: Adaptation to Environmental Factors, Pathogenicity to Cucurbits and Sensitivity to Fungicides" Journal of Fungi 6, no. 3: 169. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030169