Specificity Influences in (1→3)-β-d-Glucan-Supported Diagnosis of Invasive Fungal Disease


Associates of Cape Cod, Inc., 124 Bernard E. St. Jean Drive, East Falmouth, MA 02536, USA
J. Fungi 2021, 7(1), 14; https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010014
Submission received: 26 November 2020
/
Revised: 17 December 2020
/
Accepted: 21 December 2020
/
Published: 29 December 2020
(This article belongs to the Special Issue Fungal Biomarkers)
Abstract
:(1→3)-β-glucan (BDG) testing as an adjunct in the diagnosis of invasive fungal disease (IFD) has been in use for nearly three decades. While BDG has a very high negative predictive value in this setting, diagnostic false positives may occur, limiting specificity and positive predictive value. Although results may be diagnostically false positive, they are analytically correct, due to the presence of BDG in the circulation. This review surveys the non-IFD causes of elevated circulating BDG. These are in the main, iatrogenic patient contamination through the use of BDG-containing medical devices and parenterally-delivered materials as well as translocation of intestinal luminal BDG due to mucosal barrier injury. Additionally, infection with Nocardia sp. may also contribute to elevated circulating BDG. Knowledge of the factors which may contribute to such non-IFD-related test results can improve the planning and interpretation of BDG assays and permit investigational strategies, such as serial sampling and BDG clearance evaluation, to assess the likelihood of contamination and improve patient care.
1. Introduction
Over the last four decades, increasing numbers of fungal biomarkers have been added to the roster of tests available as aids to the diagnosis of invasive fungal disease (IFD) [1,2,3]. Among these is (1→3)-β-glucan, a cell wall component of almost all pathogenic fungi, with the exception of the Mucorales [4,5]. At this point, analysis of circulating titers of (1→3)-β-d-glucan (BDG) has been practiced for almost three decades, as an adjunct to the diagnosis of IFD [6]. The BDG test offers a relatively simple, non-invasive opportunity to obtain information on a near pan-fungal biomarker with demonstrated relevance to IFD diagnosis and, importantly, anti-fungal stewardship [7,8]. As such, over the years, it has been incorporated into an increasing number of clinical guidelines and routine practice algorithms [9,10].
Commercialization of BDG diagnostics has primarily involved reagent preparations from the blood cells of two genera of Horseshoe Crabs, Limulus polyphemus and Tachypleus tridentatus [11,12]. The former is found only on the east coast of North America while the latter is found off the coast of China and in coastal waters to its south. These preparations of BDG detection reagents utilize a protein zymogen extract from Horseshoe Crab granulocytes commonly referred to as Limulus amebocyte lysate or LAL [13]. The key detection components are referred to as the LAL cascade. The LAL cascade is comprised of two independent proteolytic activation pathways, one activated by bacterial endotoxin and the other by BDG, and a common terminal protease (Figure 1) [11].
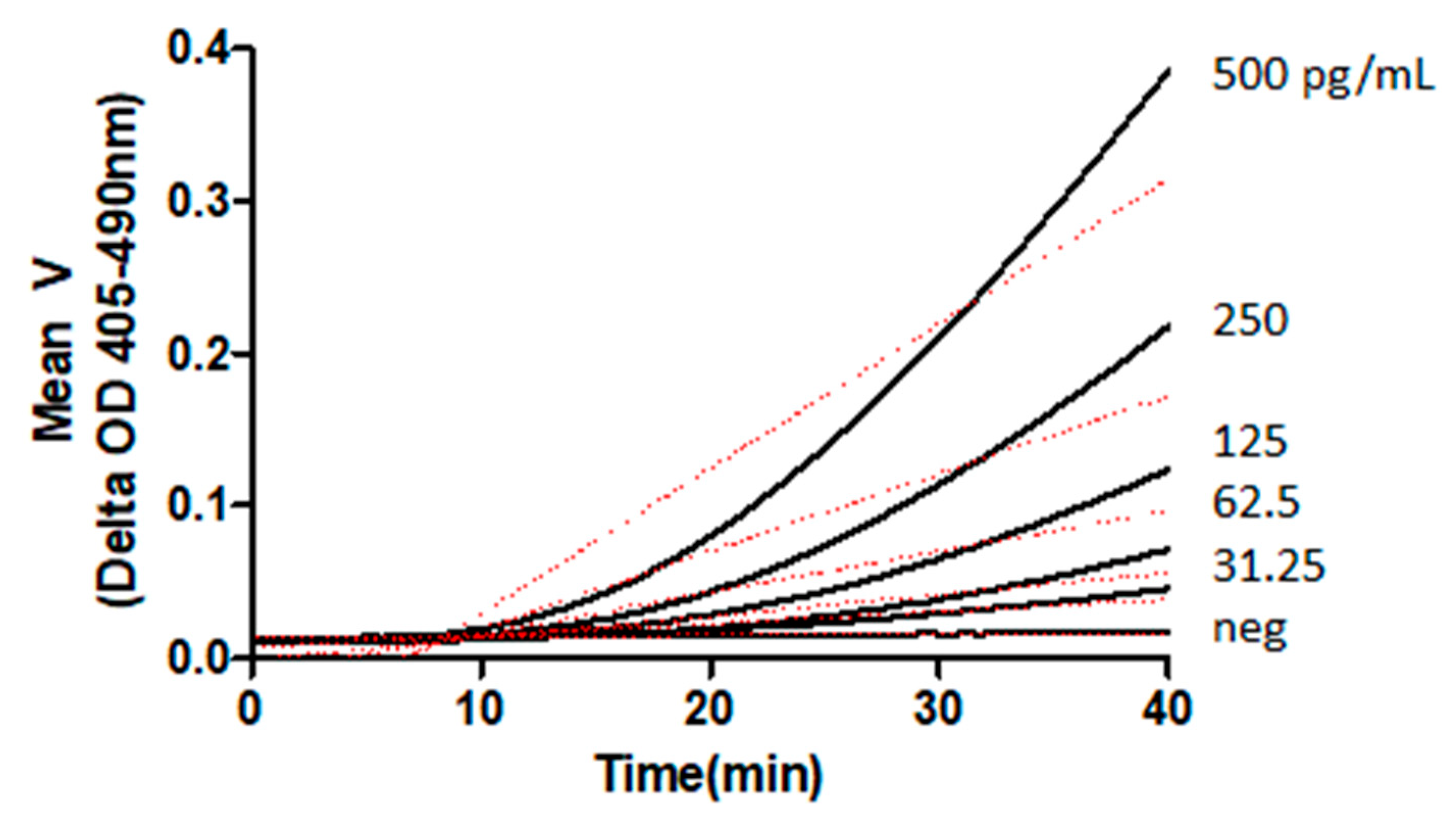
Inactivation, or elimination, of the endotoxin-specific components of the cascade renders it exclusively sensitive to BDG, permitting the creation of a reagent suitable for laboratory assays for BDG [14,15]. These assays have been established, commercially, using kinetic photometric methods [16]. Examples of chromogenic kinetic reactions of BDG are presented in Figure 2. The linear range of glucan concentrations accessible to LAL-based reagent BDG detection is dependent upon the formulation used and normally extends over a serum range that includes a negative, indeterminate, and positive range, with respect to invasive fungal disease [17].
2. (1→3)-β-Glucan Distribution and Structure
(1→3)-β-glucan is widely synthesized, for multiple purposes, among bacteria, fungi, algae, and plants [18,19,20,21]. It is not produced in mammals, nor are mammals capable of its enzymatic degradation [22]. Degradation of BDG has been observed in phagocytic cells and is presumed to occur chemically, through hydrolytic mechanisms [23]. Some of the myriad functions of BDG include use as a cell wall structural material in fungi [24], as extra-cellular and intra-cellular matrix material in certain bacteria [25], and algae [26], respectively. In plants, it appears as a wound repair and specialized structural material [27], typically in wound response tissue [28], vascular elements [29], and as intracellular pressure regulators in plasmodesmata [30]. In mammals and insects, which do not make BDG, it is recognized as an innate immune system-activating pathogen-associated molecular pattern and influences innate immuno-metabolism [31,32,33]. BDG is comprised of sequential d-glucose molecules linked by (1→3)-β-glycosidic linkages. The primary backbone structure of BDG is presented in Figure 3, with a fungi-typical (1→6)-β-linked glucose branch. The β linkage imposes steric constraints upon the orientation of the adjacent glucose molecules leading to the formation of a helical structure as the polyglucan chain becomes elongated [34,35,36]. Side chain addition, or branching, in multiple forms and with multiple moieties is also observed to occur, creating characteristic BDG types among different taxonomic groups [37]. Single helical BDG is observed to anneal through hydrogen bonding and hydrophobic interactions, resulting in native triple helical structures. Assemblages of triple helical structures also form, leading to ordered cable-like materials that contribute mechanical strength and structure to fungal cell walls [38].
3. Diagnostic Performance and the Role of Non-IFD Origin BDG
BDG can be contributed to the circulation during cell wall remodeling in the life cycle of fungal cells [39]. The extent to which this occurs is dependent upon myriad factors including fungal genus and species, site of infection, access to the vasculature, host factors including immune response, and clearance efficacy. Assay reactivity is dependent upon structural factors such as BDG molecular weight, branching, and single versus triple helical structure [40]. While commercial BDG assays are considered qualitative assays, due to the multiple factors that can contribute to titer, they have continuous numerical outputs which have permitted the validation of clinical thresholds for negative, indeterminate, and positive association with invasive fungal disease. In this context, the results of BDG testing are usually evaluated using the principles appropriate to quantitative assays [7].
The diagnostic performance of BDG assays has been assessed in approximately 200 publications, including multiple meta-analyses [7,41]. Although the heterogeneity of many studies is relatively high, diagnostic performance characteristics are reasonably reproducible. Typically, BDG negative predictive value is very high, often greater than 95%, while positive predictive value is lower, due to observations of BDG titers above the positive threshold, which are deemed to be diagnostic false positives [42]. Analytically, the BDG reagents are being activated by the actual presence of BDG in the serum samples. Accordingly, these findings beg the question of the source or sources of the BDG. At this point, three decades of clinical BDG testing experience as well as pre-clinical research suggest that three main sources of circulating BDG exist. These include IFD, iatrogenic contamination, and intestinal translocation. Non-specific pulmonary translocation due to fungal colonization is also a potential source, given the high levels of serum BDG observed to translocate from the alveoli in pneumocystosis [43], but non-IFD pulmonary translocation studies have not been performed and represent an area where data are needed [44].
4. Major Sources of Circulating BDG
4.1. Invasive Fungal Disease
The subject of fungal cell wall-originating BDG contribution to the circulation has been well described in numerous publications, including multiple meta-analyses [43,45,46,47,48]. This occurs as a result of the normal processes of the fungal life cycle in which the cyclical processes of wall polysaccharide synthesis and degradation occur [38,49]. Minute quantities of wall material, including BDG, are sloughed into the peri-fungal environment and migrate to the circulation. In the case of fungemia, in which the fungi are growing in the bloodstream, the contribution to the circulation is direct. The longitudinal characterization of circulating BDG titers typically demonstrates rising BDG levels early in infection, followed by slow declines which may only return to baseline well after successful disease resolution [50,51]. Another route of entry to the circulation was described by Hong et al. 2004, among others, in which phagocytosed fungal cells are degraded by hydrolytic processes within the phagolysosomes followed by externalization of degraded material [23]. BDG from phagocytosed yeast sacculi was demonstrated to be processed by macrophages both in vivo and in vitro. In the latter experiments, externalized BDG titers rose in the cell culture supernatants while declining within the cells. This may be a process that influences the observed slow decline of circulating BDG post-therapeutic success [52].
4.2. Iatrogenic Contamination
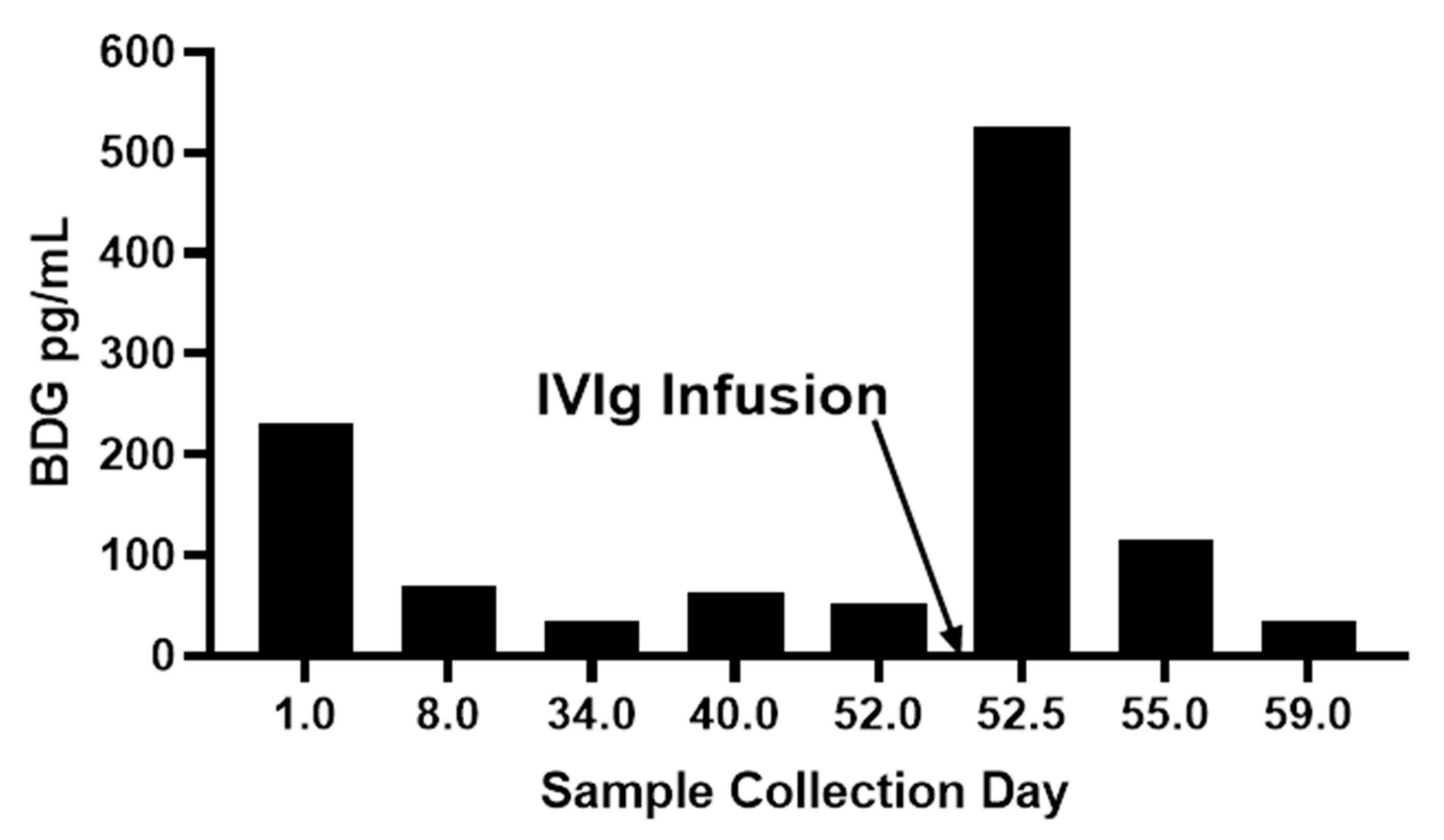
Medical treatment-related BDG contamination has been assessed in a variety of clinical contexts including multiple types of intravenously administered materials including drugs and blood fractionation products, invasive use of surgical materials, and cellulosic dialysis membranes Table 1. In the case of drug formulations, BDG may be present in the original source material itself, such as products made by fungal fermentation [53], in excipients added to the formulation [54], from media used in microbial or cell culture [55], or from process equipment, materials, and solutions [56]. In the case of blood fractionation products such as IVIg and serum albumin, the filtration of blood plasma through cellulosic depth filters can result in leached plant BDG (callose) [57,58]. It is fairly common to observe highly elevated BDG titers in patients receiving albumin or immunoglobulin (IVIg) infusions [59]. These high titers are usually observed to decline relatively quickly and such responses support suspicion of iatrogenic contamination. Figure 4 present observations for serum BDG pre-and post-infusion of an intravenous immunoglobulin product and the follow-up titers from subsequent blood draws two days and a week later. The very rapid rise and fall of serum BDG subsequent to its infusion is characteristic of such contamination events.
Surgical materials such as sponges and gauze permit large quantities of BDG (up to millions of pg/gm of gauze or sponge material) to be leached during intra-cavity or other invasive use, as well as contact with surgical solutions and bodily fluids such as with certain autologous blood recovery procedures [60,61]. Mohr et al. 2011, demonstrated that BDG diagnostic specificity in post-surgical patients improved, substantially, between sampling immediately post-surgery and after 3 days [62]. Similarly, the use of gauze in surgery has been observed to contribute to leaching and elevated circulating burdens of BDG [63]. BDG is also a licensed drug in certain countries, as an anticancer adjunctive therapy based upon its innate immune activation properties. Injected particulate BDG has been shown to generate elevated circulatory BDG for years after administration, making detailed medical history analysis critical to interpretation of unexpectedly high BDG titers in patients in whom IFD is unsuspected [64]. Renal replacement therapy utilizing regenerated cellulose was demonstrated to contribute to elevated BDG in hemodialysis patients [65]. As regenerated cellulose dialysis membranes have been replaced by non-BDG-leaching synthetic membranes, these are now an unlikely source [66].
Based upon these observations of patient contamination by multiple types of medical materials and procedures, blood draw planning for BDG titer determination should take these into account and unexpected BDG elevation should prompt chart review.
Another category of iatrogenic patient contamination, one that has had significant debate, is that of parenteral administration of potentially contaminated antibiotics. Multiple publications asserting the occurrence of patient false positives by this route as well as the opposite have been presented [67,68,69]. While this route of patient contamination is possible, the high level of dilution generated upon injection of relatively low volumes of antibiotic make this unlikely. Further, the high negative predictive value for IFD observed for patients receiving a vast array of antibiotics suggests that this is not a significant problem [70,71].
4.3. Intestinal Translocation
The past two decades have seen an explosion in data regarding intestinal barrier permeability, its myriad causes, and its relationship to inflammation-related pathophysiology [73,74,75]. Multiple biomarkers such as zonulin and various claudins have been identified as useful markers in characterizing the nature of barrier injury [76,77]. Data regarding elevated circulating BDG, its correlation with other markers of microbial translocation and inflammation, and orthogonal testing with enteral polysaccharides such as fluorescein labeled dextran demonstrate that it is also a translocated entity [78,79,80]. Ellis et al. 2008 observed that in a series of non-IFD hematological malignancy patients undergoing chemotherapy, those with enterocyte damage and/or mucositis had BDG titers that were persistently elevated compared to patients without those pathologies (p = 0.002) [78]. MAF: Sentence added with a new reference to restore numerical reference sequencing. Pre-clinical models of sepsis also demonstrate elevated levels of circulating BDG [79]. The kinds of insults that contribute to intestinal barrier permeability include intestinal ischemia, mesenteric hypoxia, microbial toxins, viral infection of intestinal tissue, metabolic toxicity such as in uremia, chemotherapy-associated mucositis, large total surface area burns, and protease-producing intestinal enterococci (Table 2). All of these conditions have been observed to be associated with elevated BDG titers. Thus, it is important to fully understand the host and clinical factors that could be contributing to intestinal barrier permeability as part of the consideration of BDG titer interpretation.
Several conditions may contribute to intestinal hypoxia, which is a causative factor in intestinal permeability barrier injury. These include large surface area burns, intestinal ischemia reperfusion injury, mesenteric hypoxia, and intra-dialytic hypotension. Studies have described the relationship between large surface area burns and BDG false positives [81,92]. Similarly, a Japanese study evaluated the use of BDG testing in the diagnosis of candidemia in severe burn patients and demonstrated high sensitivity, 100%, but lower specificity, 68%, when the test manufacturer’s positive cutoff of 11 pg/mL was used. A cutoff of 40 pg/mL generated a sensitivity of 100% with a specificity of 95% [93]. Validation of alternative cutoffs in specific clinical contexts known to contribute to elevated BDG titer may represent a means of dealing with specificity issues.
End stage renal disease is a condition associated with intestinal permeability barrier injury and microbial metabolite translocation from the lumen [94]. Additionally, hemodialysis is associated with intradialytic hypotension and attendant loss of intestinal permeability barrier patency [95]. It has been reported that hemodialysis within the previous 72 h was associated with BDG false positives, p = 0.011 [96].
4.4. Intestinal Contents: Mycobiome and BDG Translocation
Translocation of BDG from the intestinal lumen through the intestinal epithelium and then to the wider circulation raises questions concerning the sources of luminal BDG. Research over the last few decades suggests that both ingested foodstuffs and the intestinal mycobiome are potential contributors. With respect to the former, certain foods are extremely rich in BDG. Oat bran BDG content is in the range of 4.5%, by weight [97]. Similarly, foods rich in fungal mycelium, such as mushrooms, represent potential sources of BDG which might translocate. At this point, the case for translocating foodstuff-derived BDG remains to be more fully evaluated. A recent study evaluated a single bolus of extremely high BDG content foodstuff consumption in an HIV population and serial testing over 24 h failed to reveal meaningful serum BDG titer elevation [98]. The intestinal mycobiome represents another potential source of translocatable BDG and there are structural differences between fungal and plant-origin BDG which may affect translocation. This is of particular interest in patients receiving gut-active broad spectrum antibacterials. The administration of such drugs has been demonstrated to produce 1–2.5 log increases in gut Candida populations in patients [99]. It is possible that the release of mycobiome-origin BDG in the gut lumen, proximal to the luminal epithelium, may result in more efficient translocation. These are areas of continuing investigation with the potential to illuminate prognosis of gut-originating candidiasis and inflammatory disease. In this light, recently published data by Zhai et al. 2020 revealed an expansion of pathogenic Candida species in the intestinal mycobiota of post-stem cell transplant patients two–ten days prior to their developing candidemia. DNA analysis showed that the candidemia strains clustered tightly with the luminal contents strains [100]. Similarly, the role of the gut mycobiome has been examined in the context of alcohol-induced liver disease in a chimeric murine model. In this model, alcohol feeding resulted in intestinal Candida overgrowth and BDG translocation with resulting hepatic inflammation, thought to occur through BDG ligation of hepatic Kuppfer cell dectin-1. Dectin-1 knock-out mice failed to develop the inflammation, leading to the conclusion that translocated intestinal origin BDG played a major role [101].
4.5. Hepatic Function
In animal models, BDG is primarily cleared through the liver [102]. Human clinical conditions which result in reduced hepatic function have been shown to be associated with elevated serum BDG. Sanada et al. 2014 have described the relationship between serum BDG levels in the setting of pediatric end stage liver disease [103]. The liver clearance function was underscored by significantly higher BDG titers in portal blood relative to the peripheral circulation. There was a very significant negative correlation (p < 0.001) between the pediatric end-stage liver disease patients score and the hepatic clearance of BDG in the transplant patients. Sanada et al. 2012 also described elevated serum BDG in end-stage liver disease patients with and without fungal infections. Of note, median hepatic clearance of circulating BDG was measured as 87.9% [104]. Elevated patient peripheral blood levels of BDG pre-transplant predicted a much longer course of recovery (p < 0.001) and, post-operatively, fungal infections. The peripheral BDG titers in the infected patients were significantly higher than the uninfected patient levels, suggesting the potential of diagnostic utility despite compromised clearance. Recently, Moon et al. 2020 analyzed microbial translocation markers in the blood of hepatitis C virus infected patients [85]. Serum BDG was significantly elevated in hepatic fibrosis (Ishak Score 0–2 and 5–6), relative to controls (Mann–Whitney, p < 0.0001 and p < 0.001, respectively). That this may be due to inadequate clearance rather than intestinal translocation is suggested by observations that enhanced microbial translocation was not observed in HCV-infected chimpanzees which do not develop liver fibrosis [105]. Thus, hepatic function status is an additional factor to be considered in the evaluation of the significance of circulating BDG titers.
4.6. Bacterial Infections
As discussed earlier in this review, a number of publications have asserted that bacterial infections have been the source of elevated serum BDG [106]. While elevated BDG titers have been associated with bacterial infections in a number of studies, the demonstration of a contribution of BDG derived directly from the bacterium has not, in the main, been systematically demonstrated. Mennink-Kersten and colleagues described elevated BDG titers in a case series of bacteremic patients, particularly in infections with multiple Pseudomonas sp. and a Streptococcus pneumoniae [107]. Among the Streptococci, S. pneumoniae Type 37 has been described as producing a BDG with a (1→3)-β-backbone with each glucose moiety having a (1→2)-β-glucose side chain [108]. Held et al. 2013 described a mean serum BDG titer of 135 pg/mL in patients with enterococcemia while patients with other causes of bacteremia displayed a mean level of 15 pg/mL [84]. Enterococcal species, which produce exopolysaccharides of mixed monosaccharide and glycosidic linkage composition, are not known to produce BDG. As the etiology of enterococcal bacteremia includes overgrowth in the intestinal lumen followed by invasion through the intestinal barrier [109], the possibility of BDG translocation through a damaged intestinal barrier represents a plausible mechanism of its elevation in the circulation. Recently, the Enterococcal exopolysaccharide poly-N-acetyl glucosamine has been determined to be a virulence factor due to its role in breaching the intestinal permeability barrier [109]. In contrast to bacterial species not observed to directly produce BDG, circulating BDG derived from invasive Nocardia infections has been definitively demonstrated in a series of independent case reports. Elevated BDG titers were reported in infections caused by multiple species of Nocardia including N.abscessus, N. elegans, N. farcinica, and N. nova [110,111]. Additional evaluation to verify the presence of BDG involving the culturing of clinical isolates of N. asteroides, N. neocalidoniensis, and N. cyriacgeorgica, as well as the control species Staphylococcus aureus and Escherichia coli, and the semi-purification of the post-growth culture broth supernatants was conducted. The un-inoculated medium and the controls were negative for BDG while the Nocardia species’ supernatants were all over-range in the assay, confirming the source of the BDG as the Nocardia infections [112]. Thus, while a number of bacterial genera are known to produce BDG, very few are human pathogens. Of those pathogenic genera that do make beta-linked polyglucans, the genus Pseudomonas makes small, 2-12-mer, periplasmic osmoregulatory cyclic (1→2)-β-linked glucan sequences [113] which are broadly similar to the short (5-12-mer) branched (1→2)-β-linked glucans produced by E. coli [114]. Accordingly, while some bacteria are capable of producing (1→3)-β-linked beta-glucans, such as Agrobacterium tumefaciens [115], of those genera that are human pathogens it is rare that they make the (1→3)-β-linked glucan structures that are the only forms known to be capable of the activation of Limulus-based detection reagents.
5. Manufacturing-Associated BDG Contamination
Production processes for parenterals are another potential source of patient-contaminating BDG. During manufacturing, the introduction of BDG to parenterals may occur through multiple sources. These include fermentation media components such as yeast extracts, plant material extracts, sugars, as well as the use of fungal organisms as the source of fermentation products [53,55,57]. If downstream processing does not remove the contaminating BDG, it may result in patient exposure. Examples of patient contamination after infusions have been reported in the literature and represent a failure to control an easily measurable contaminant [59,116]. Another source of parenteral contamination is the use of cellulosic depth filters [57]. These devices are generally mixtures of cellulose and diatomaceous earth and are used to provide initial clarification of cell culture fluid, microbial fermentation broths, and blood plasma. As plant material contains small quantities of BDG (callose) in various plant tissues [117], BDG is readily leached from the depth filters to enter downstream processing as a contaminant [57,118], Blood fractionation products, whose starting material is depth filtered blood plasma, are recognized sources of product contamination with BDG [119,120].
In addition to process equipment, process solutions may contain BDG and introduce contamination. Vigor et al. 2017 reported two sources of process-related BDG contamination. One was BDG contamination in sucrose, used as a formulating excipient, and the other was a virus filter storage solution [56]. With its recognition as a bioactive contaminant, pharmaceutical manufacturing organizations are beginning to measure and control BDG contamination of parenterals [55,57].
6. False Positive Investigation
BDG is indicated as an adjunct to the diagnosis of invasive fungal disease. It is to be considered with reference to host, clinical, and other laboratory characteristics and findings. When an elevated BDG titer appears to be discordant with respect to these other observations, it is useful to evaluate potential sources of diagnostically false positive BDG presence. Table 3 provides a listing of a series of medical product and clinical condition-related questions which may help to clarify a previously unrecognized source of non-IFD-related BDG.
In addition to investigation of the above-listed potential sources of circulating BDG, support for the likely presence of a fungal source of the observed BDG may be found by utilizing additional tests. These include microscopic examination of appropriate patient tissue and fluid samples, as well as various PCR and antigen tests, as well as emerging novel diagnostic technologies [121]. Positive results on ancillary tests for the presence of fungi can add to support for ruling out BDG diagnostic false positivity. Multiple studies have reported improved specificity in the setting of multiple fungal marker utilization. Boch et al. 2016 reported the increase in specificity, in the setting of invasive aspergillosis, from 48% to 94% when serum BDG results were combined with BAL galactomannan results [122]. In the setting of acute leukemia, Qian et al. 2019 described specificity values for either BDG or GM alone, for invasive fungal disease, at 90% and 40%, respectively [123]. In combination, the specificity was 98%. Similarly, Urabe et al. 2017 demonstrated that combining Aspergillus PCR with BDG raised specificity from 48.2% in bronchoalveolar lavage fluid, to 96.7% and 99.2% with two different PCR assays, respectively [124]. In contrast, Held et al. 2013 reported that, in the setting of candidemia, combining mannan antigen testing with serum BDG lowered the specificity marginally, from 85.5% with BDG alone to 85.0% [84].
7. Discussion
As described above, circulating BDG titer is an important adjunct in the diagnostic process for patients suspected of invasive fungal infections. While the NPV is very high, optimizing the utility of positive BDG results requires consideration of both host and clinical factors, as well as potential sources of iatrogenic contamination. The latter factors may include introduction of BDG-contaminated materials in the course of medical care. In that circumstance, institutional observations suggesting an association between elevated BDG and certain parenterals or devices should prompt evaluation of BDG burdens present in those products. Prospective BDG testing practices should employ blood draws taken prior to introduction of materials considered suspect. Similarly, blood draws obtained during the initial 3 days post-surgery involving intra-cavity use of gauze and surgical sponges need to be interpreted cautiously and the observation of very rapidly declining serial sample values may indicate contamination rather than infection. In addition, BDG testing results of samples taken after intra-operative autologous blood recovery using surgical sponges must be similarly considered. In patients with intestinal permeability barrier injury risk factors, the presence of elevated BDG absent a diagnosis of IFD and iatrogenic contamination should prompt consideration of potential luminal contents translocation. This is also of significant importance given the association of intestinal permeability barrier injury and the potential for infection due to translocation of viable microorganisms, including Candida, to the circulation. Other potential sources of elevated BDG include hepatic insufficiency, potentially leading to reduced BDG clearance, and infection with Nocardiales sp.
Invasive fungal disease continues to have high morbidity and mortality and the development of adequate diagnostics has been challenging [3,125]. Fungal antigen tests, including BDG, represent advances toward the goal of improving the effective diagnosis of IFD. In order for them to be used effectively, it is critical that laboratories and health care practitioners are well acquainted with the factors that can influence results, within the patient’s clinical context.
Funding
This research received no external funding.
Institutional Review Board Statement
Not Applicable: Published literature review or de-identified clinical laboratory patient data, only.
Informed Consent Statement
Not Applicable: Published literature review or de-identified clinical laboratory patient data, only.
Data Availability Statement
All non-published literature sourced data is present in the manuscript.
Acknowledgments
The author would like to thank Robert D’Ordine for careful review of the documents and useful suggestions as well as assistance with the figures.
Conflicts of Interest
Malcolm A. Finkelman is an employee of Associates of Cape Cod, Inc., the manufacturer of a commercial BDG detection in vitro diagnostic kit.
References
- Bassetti, M.; Scudeller, L.; Giacobbe, D.R.; Lamoth, F.; Righi, E.; Zuccaro, V.; Grecchi, C.; Rebuffi, C.; Akova, M.; Alastruey-Izquierdo, A.; et al. Developing definitions for invasive fungal diseases in critically ill adult patients in intensive care units. Protocol of the FUNgal infections Definitions in ICU patients (FUNDICU) project. Mycoses 2019, 62, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Bouza, E.; Almirante, B.; Rodríguez, J.G.; Garnacho-Montero, J.; Salavert, M.; Muñoz, P.; Sanguinetti, M. Biomarkers of fungal infection: Expert opinion on the current situation. Rev. Esp. Quim. 2020, 33, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnelly, J.P.; Chen, S.C.; Kauffman, C.A.; Steinbach, W.J.; Baddley, J.W.; Verweij, P.E.; Clancy, C.J.; Wingard, J.R.; Lockhart, S.R.; Groll, A.H.; et al. Revision and Update of the Consensus Definitions of Invasive Fungal Disease from the European Or-ganization for Research and Treatment of Cancer and the Mycoses Study Group Education and Research Consortium. Clin. Infect. Dis. 2020, 71, 1367–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osibe, D.A.; Lei, S.; Wang, B.; Jin, C.; Fang, W. Cell wall polysaccharides from pathogenic fungi for diagnosis of fungal infectious disease. Mycoses 2020, 63, 644–652. [Google Scholar] [CrossRef]
- Patterson, T.F.; Donnelly, J.P. New Concepts in Diagnostics for Invasive Mycoses: Non-Culture-Based Methodologies. J. Fungi 2019, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Obayashi, T. The Plasma (1→3)-β-d-glucan Assay, a Japanese Contribution to the Diagnosis of Invasive Fungal Infection. Med. Mycol. J. 2017, 58, J141–J147. [Google Scholar] [CrossRef] [Green Version]
- Kethireddy, S.; Kumar, A. β-d-Glucan Assay for the Diagnosis of Invasive Fungal Infections: A Meta-analysis. Yearb. Crit. Care Med. 2012, 2012, 97–99. [Google Scholar] [CrossRef]
- Posteraro, B.; Tumbarello, M.; De Pascale, G.; Liberto, E.; Vallecoccia, M.S.; De Carolis, E.; Di Gravio, V.; Trecarichi, E.M.; Sanguinetti, M.; Antonelli, M. (1,3)-β-d-Glucan-based antifungal treatment in critically ill adults at high risk of candidaemia: An observational study. J. Antimicrob. Chemother. 2016, 71, 2262–2269. [Google Scholar] [CrossRef] [Green Version]
- Cuenca-Estrella, M.; Verweij, P.; Arendrup, M.C.; Arikan-Akdagli, S.; Bille, J.; Donnelly, J.P.; Jensen, H.E.; Lass-Flörl, C.; Richardson, M.D.; Akova, M.; et al. ESCMID* guideline for the diagnosis and management of Candida diseases 2012: Diagnostic procedures. Clin. Microbiol. Infect. 2012, 18 (Suppl. 7), 9–18. [Google Scholar] [CrossRef] [Green Version]
- Groll, A.H.; Castagnola, E.; Cesaro, S.; Dalle, J.H.; Engelhard, D.; Hope, W.; Roilides, E.; Styczynski, J.; Warris, A.; Lehrn-becher, T. Fourth European Conference on Infections in Leukaemia; Infectious Diseases Working Party of the European Group for Blood Marrow Transplantation (EBMT-IDWP); Infectious Diseases Group of the European Organisation for Re-search and Treatment of Cancer (EORTC-IDG); International Immunocompromised Host Society (ICHS); European Leu-kaemia Net (ELN). Fourth European Conference on Infections in Leukaemia (ECIL-4): Guidelines for diagnosis, prevention, and treatment of invasive fungal diseases in paediatric patients with cancer or allogeneic haemopoietic stem-cell trans-plantation. Lancet Oncol. 2014, 15, e327–e340. [Google Scholar]
- Kawabata, S.-I.; Muta, T. Sadaaki Iwanaga: Discovery of the lipopolysaccharide- and -1,3-d-glucan-mediated proteolytic cascade and unique proteins in invertebrate immunity. J. Biochem. 2010, 147, 611–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwanaga, S. Biochemical principle of Limulus test for detecting bacterial endotoxins. Proc. Jpn. Acad. 2007, 83, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Brandenburg, K.; Howe, J.; Gutsman, T.; Garidel, P. The expression of endotoxic activity in the Limulus test as compared to cytokine production in immune cells. Curr. Med. Chem. 2009, 16, 2653–2660. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Kohno, S.; Koga, H.; Kaku, M.; Mitsutake, K.; Maesaki, S.; Yasuoka, A.; Hara, K.; Tanaka, S.; Tamura, H. G test, a new direct method for diagnosis of Candida infection: Comparison with assays for beta-glucan and mannan antigen in a rabbit model of systemic candidiasis. J. Clin. Lab. Anal. 1992, 6, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, T.; Yoshida, M.; Tamura, H.; Aketagawa, J.; Tanaka, S.; Kawai, T. Determination of plasma (1→3)-beta-d-glucan: A new diagnostic aid to deep mycosis. J. Med. Vet. Mycol. 1992, 30, 275–280. [Google Scholar] [CrossRef]
- Yoshida, K.; Shoji, H.; Takuma, T.; Niki, Y. Clinical viability of Fungitell, a new (1→3)-β-d-glucan measurement kit, for diagnosis of invasive fungal infection, and comparison with other kits available in Japan. J. Infect. Chemother. 2011, 17, 473–477. [Google Scholar] [CrossRef]
- Patel, R. Faculty Opinions recommendation of Multicenter clinical evaluation of the (1→3) beta-d-glucan assay as an aid to diagnosis of fungal infections in humans. Fac. Opin. Post-Publ. Peer Rev. Biomed. Lit. 2005, 41. [Google Scholar] [CrossRef]
- Yagyu, K.; Nakatsuji, Y.; Matsushita, H. Elevated serum β-d-glucan levels in cavitary pulmonary nocardiosis. BMJ Case Rep. 2020, 13, e234738. [Google Scholar] [CrossRef]
- Ruiz-Herrera, J.; Ortiz-Castellanos, L. Cell wall glucans of fungi. A review. Cell Surf. 2019, 5, 100022. [Google Scholar] [CrossRef]
- Du, B.; Meenu, M.; Liu, H.; Xu, B. A Concise Review on the Molecular Structure and Function Relationship of β-Glucan. Int. J. Mol. Sci. 2019, 20, 4032. [Google Scholar] [CrossRef] [Green Version]
- Schneider, R.; Hanak, T.; Persson, S.; Voigt, C.A. Cellulose and callose synthesis and organization in focus, what’s new? Curr. Opin. Plant. Biol. 2016, 34, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Cortés, J.C.G.; Curto, M.-Á.; Carvalho, V.S.; Pérez, P.; Ribas, J.C. The fungal cell wall as a target for the development of new antifungal therapies. Biotechnol. Adv. 2019, 37, 107352. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.; Yan, J.; Baran, J.; Allendorf, D.J.; Hansen, R.D.; Ostroff, G.R.; Xing, P.X.; Cheung, N.-K.V.; Ross, G.D. Mechanism by which orally administered beta-1,3-glucans enhance the tumoricidal activity of antitumor monoclonal antibodies in murine tumor models. J. Immunol. 2004, 173, 797–804. [Google Scholar] [CrossRef] [Green Version]
- Wagener, J.; Striegler, K.; Wagener, N. α- and β-1,3-Glucan Synthesis and Remodeling. Curr. Top. Microbiol. Immunol. 2020, 425, 53–82. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Misaki, A.; Saito, H. Curdlan: A bacterial gel-forming beta-1,3-glucan. Arch. Biochem. Biophys. 1968, 124, 292–298. [Google Scholar] [CrossRef]
- Bobadilla, F.; Rodríguez-Tirado, C.; Imarai, M.; Galotto, M.J.; Andersson, R. Soluble β-1,3/1,6-glucan in seaweed from the southern hemisphere and its immunomodulatory effect. Carbohydr. Polym. 2013, 92, 241–248. [Google Scholar] [CrossRef]
- Burton, R.A.; Fincher, G.B. (1,3;1,4)-beta-d-glucans in cell walls of the poaceae, lower plants, and fungi: A tale of two linkages. Mol. Plant. 2009, 2, 873–882. [Google Scholar] [CrossRef] [Green Version]
- Savatin, D.V.; Gramegna, G.; Modesti, V.; Cervone, F. Wounding in the plant tissue: The defense of a dangerous passage. Front. Plant. Sci. 2014, 5, 470. [Google Scholar] [CrossRef] [Green Version]
- Giannoutsou, E.; Sotiriou, P.; Nikolakopoulou, T.L.; Galatis, B.; Apostolakos, P. Callose and homogalacturonan epitope dis-tribution in stomatal complexes of Zea mays and Vigna sinensis. Protoplasma 2020, 257, 141–156. [Google Scholar] [CrossRef]
- Liu, N.J.; Zhang, T.; Liu, Z.H.; Chen, X.; Guo, H.S.; Ju, B.H.; Zhang, Y.Y.; Li, G.Z.; Zhou, Q.H.; Qin, Y.M.; et al. Phyto-sphinganine Affects Plasmodesmata Permeability via Facilitating PDLP5-Stimulated Callose Accumulation in Arabidopsis. Mol. Plant. 2020, 13, 128–143. [Google Scholar] [CrossRef]
- Tsuzuki, A.; Tateishi, T.; Ohno, N.; Adachi, Y.; Yadomae, T. Increase of hematopoietic responses by triple or single helical conformer of an antitumor (1→3)-beta-d-glucan preparation, Sonifilan, in cyclophosphamide-induced leukopenic mice. Biosci. Biotechnol. Biochem. 1999, 63, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Underhill, D.M. β-Glucan signaling connects phagocytosis to autophagy. Glycobiology 2013, 23, 1047–1051. [Google Scholar] [CrossRef] [Green Version]
- Legentil, L.; Paris, F.; Ballet, C.; Trouvelot, S.; Daire, X.; Vetvicka, V.; Ferrières, V. Molecular Interactions of β-(1→3)-Glucans with Their Receptors. Molecules 2015, 20, 9745–9766. [Google Scholar] [CrossRef] [PubMed]
- Sanada, Y.; Matsuzaki, T.; Mochizuki, S.; Okobira, T.; Uezu, K.; Sakurai, K. β-1,3-d-glucan schizophyllan/poly(dA) triple-helical complex in dilute solution. J. Phys. Chem. B. 2012, 116, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sheng, X.; Shi, A.; Hu, H.; Yang, Y.; Liu, L.; Fei, L.; Liu, H. β-Glucans: Relationships between Modification, Conformation and Functional Activities. Molecules 2017, 22, 257. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Lan, P.; He, Y.; Li, C.; Ma, X. Effect of the Modifications on the Physicochemical and Biological Properties of β-Glucan—A Critical Review. Molecules 2019, 25, 57. [Google Scholar] [CrossRef] [Green Version]
- Adachi, Y.; Ishii, M.; Kanno, T.; Tetsui, J.; Ishibashi, K.; Yamanaka, D.; Miura, N.; Ohno, N. N-Terminal (1→3)-β-d-Glucan Recognition Proteins from Insects Recognize the Difference in Ultra-Structures of (1→3)-β-d-Glucan. Int. J. Mol. Sci. 2019, 20, 3498. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Herrera, J. Biosynthesis of beta-glucans in fungi. Antonie Van Leeuwenhoek 1991, 60, 72–81. [Google Scholar] [CrossRef]
- Guillen, A.; Leal, F.; Andaluz, E.; Larriba, G. Endogenous factors that modulate yeast glucan synthetase in cell-free extracts. Biochim. Biophys. Acta Gen. Subj. 1985, 842, 151–161. [Google Scholar] [CrossRef]
- Finkelman, M.A.; Tamura, H. Detection and measurement of (1→3)-β-d-glucan. In Toxicology of (1→3)-Beta-Glucans; Young, S.H., Castranova, V., Eds.; CRC Press: New York, NY, USA, 2005. [Google Scholar]
- Del Corpo, O.; Butler-Laporte, G.; Sheppard, D.C.; Cheng, M.P.; McDonald, E.G.; Lee, T.C. Diagnostic accuracy of serum (1-3)-β-d-glucan for Pneumocystis jirovecii pneumonia: A systematic review and meta-analysis. Clin. Microbiol. Infect. 2020, 26, 1137–1143. [Google Scholar] [CrossRef]
- Xiaoling, L.; Tingyu, T.; Caibao, H.; Tian, Z.; Changqin, C. Diagnostic efficacy of serum 1,3-β-d-glucan for invasive fungal infection: An update meta-analysis based on 37 case or cohort studies. Open Med. 2018, 13, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Karageorgopoulos, D.E.; Qu, J.M.; Korbila, I.P.; Zhu, Y.G.; Vasileiou, V.A.; Falagas, M.E. Accuracy of β-d-glucan for the diagnosis of Pneumocystis jirovecii pneumonia: A meta-analysis. Clin. Microbiol. Infect. 2013, 19, 39–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rautemaa, V.; Green, H.D.; Jones, A.M.; Rautemaa-Richardson, R. High level of β-(1,3)-d-glucan antigenaemia in cystic fi-brosis in the absence of invasive fungal disease. Diagn. Microbiol. Infect. Dis. 2017, 88, 316–321. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Hang, J.P.; Zhang, L.; Wang, F.; Zhang, D.C.; Gong, F.H. A systematic review and meta-analysis of diagnostic accuracy of serum 1,3-β-D-glucan for invasive fungal infection: Focus on cutoff levels. J. Microbiol. Immunol. Infect. 2015, 48, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Lamoth, F.; Cruciani, M.; Mengoli, C.; Castagnola, E.; Lortholary, O.; Richardson, M.; Marchetti, O. Third European Confer-ence on Infections in Leukemia (ECIL-3). β-Glucan antigenemia assay for the diagnosis of invasive fungal infections in pa-tients with hematological malignancies: A systematic review and meta-analysis of cohort studies from the Third European Conference on Infections in Leukemia (ECIL-3). Clin. Infect. Dis. 2012, 54, 633–643. [Google Scholar]
- Karageorgopoulos, D.E.; Vouloumanou, E.K.; Ntziora, F.; Michalopoulos, A.; Rafailidis, P.I.; Falagas, M.E. β-D-glucan assay for the diagnosis of invasive fungal infections: A meta-analysis. Clin. Infect. Dis. 2011, 52, 750–770. [Google Scholar] [CrossRef]
- Li, W.-J.; Guo, Y.-L.; Liu, T.-J.; Wang, K.; Kong, J.-L. Diagnosis of pneumocystis pneumonia using serum (1-3)-β-d-Glucan: A bivariate meta-analysis and systematic review. J. Thorac. Dis. 2015, 7, 2214–2225. [Google Scholar]
- Kottom, T.J.; Limper, A.H. ThePneumocystisAce2 Transcription Factor Regulates Cell Wall-remodeling Genes and Organism Virulence. J. Biol. Chem. 2013, 288, 23893–23902. [Google Scholar] [CrossRef] [Green Version]
- Held, J.; Wagner, D. β-d-Glucan kinetics for the assessment of treatment response in Pneumocystis jirovecii pneumonia. Clin. Microbiol. Infect. 2011, 17, 1118–1122. [Google Scholar] [CrossRef] [Green Version]
- Mikulska, M.; Furfaro, E.; Del Bono, V.; Gualandi, F.; Van Lint, M.T.; Miletich, F.; Bacigalupo, A.; Viscoli, C. Persistence of a positive (1,3)-beta-d-glucan test after clearance of candidemia in hematopoietic stem cell transplant recipients. Clin. Vaccine Immunol. 2011, 18, 518–519. [Google Scholar] [CrossRef] [Green Version]
- Angebault, C.; Lanternier, F.; Dalle, F.; Schrimpf, C.; Roupie, A.-L.; Dupuis, A.; Agathine, A.; Scemla, A.; Paubelle, E.; Caillot, D.; et al. Prospective Evaluation of Serum β-Glucan Testing in Patients with Probable or Proven Fungal Diseases. Open Forum Infect. Dis. 2016, 3, ofw128. [Google Scholar] [CrossRef] [PubMed]
- Behera, B.C. Citric acid from Aspergillus niger: A comprehensive overview. Crit. Rev. Microbiol. 2020, 46, 727–749. [Google Scholar] [CrossRef]
- Jacobs, S.; Bunt, C.R.; Wu, Z.; Lehr, C.M.; Rupenthal, I.D. Characterization and evaluationof β-glucan formulations as injectable implants for protein and peptide delivery. Drug Dev. Ind. Pharm. 2012, 38, 1337–1343. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Scherfner, S.; Dick, L.W., Jr.; Mahon, D.; Qiu, D.; Cheng, K.C.; Shukla, A.A. Demonstrating β-glucan and yeast peptide clearance in biopharmaceutical downstream processes. Biotechnol. Prog. 2011, 27, 442–450. [Google Scholar] [CrossRef]
- Vigor, K.; Emerson, J.; Scott, R.; Cheek, J.; Barton, C.; Bax, H.J.; Josephs, D.H.; Karagiannis, S.N.; Spicer, J.F.; Lentfer, H. Development of downstream processing to minimize beta-glucan impurities in GMP-manufactured therapeutic antibodies. Biotechnol. Prog. 2016, 32, 1494–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holstein, M.; Jang, D.; Urrea, C.; Botta, L.S.; Grimm, W.; Ghose, S.; Li, Z.J. Control of Leached Beta-Glucan Levels from Depth Filters by an Improved Depth Filtration Flush Strategy. Biotechnol. Prog. 2020, e3086. [Google Scholar] [CrossRef]
- Gefroh, E.; Hewig, A.; Vedantham, G.; McClure, M.; Krivosheyeva, A.; Lajmi, A.; Lu, Y. Multipronged approach to managing beta-glucan contaminants in the downstream process: Control of raw materials and filtration with charge-modified nylon 66 membrane filters. Biotechnol. Prog. 2013, 29, 672–680. [Google Scholar] [CrossRef]
- Duffner, U.; Abdel-Mageed, A.; Dahl, K.; Fogg, G.; Hester, J. Serum (1→3)-β-d-glucan levels (Fungitell assay) is not useful as a screening test for recipients of an allogeneic HSCT while on immunoglobulin replacement. Bone Marrow Transpl. 2011, 47, 151–152. [Google Scholar] [CrossRef] [Green Version]
- Styczynski, A.; Bonilla, H.; Treynor, E.; Shashank, J.; Zhang, Y.; Finkelman, M. Beta-Glucanemia after Coronary Artery By-pass Graft Surgery: A Case Report. J. Fungi 2018, 4, 114. [Google Scholar] [CrossRef] [Green Version]
- Ronai, A.K.; Glass, J.J.; Shapiro, A.S. Improving Autologous Blood Harvest: Recovery of Red Cells from Sponges and Suction. Anaesth. Intensiv. Care 1987, 15, 421–424. [Google Scholar] [CrossRef] [Green Version]
- Mohr, J.F.; Sims, C.; Paetznick, V.; Rodriguez, J.; Finkelman, M.A.; Rex, J.H.; Ostrosky-Zeichner, L. Prospective survey of (1→3)-beta-d-glucan and its relationship to invasive candidiasis in the surgical intensive care unit setting. J. Clin. Microbiol. 2011, 49, 58–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanamori, H.; Kanemitsu, K.; Miyasaka, T.; Ameku, K.; Endo, S.; Aoyagi, T.; Inden, K.; Hatta, M.; Yamamoto, N.; Kunishi-ma, H.; et al. Measurement of (1-3)-beta-d-glucan derived from different gauze types. Tohoku J. Exp. Med. 2009, 217, 117–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokuyasu, H.; Takeda, K.; Kawasaki, Y.; Sakaguchi, Y.; Isowa, N.; Shimizu, E.; Ueda, Y. High plasma concentration of beta-d-glucan after administration of sizofiran for cervical cancer. Int. J. Gen. Med. 2010, 3, 273–277. [Google Scholar] [PubMed] [Green Version]
- Kanda, H.; Kubo, K.; Hamasaki, K.; Kanda, Y.; Nakao, A.; Kitamura, T.; Fujita, T.; Yamamoto, K.; Mimura, T. Influence of various hemodialysis membranes on the plasma (1→3)-beta-d-glucan level. Kidney Int. 2001, 60, 319–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prattes, J.; Schneditz, D.; Prüller, F.; Jaindl, E.; Sauseng, N.; Hoenigl, M.; Schilcher, G.; Krause, R. 1,3-ß-d-Glucan testing is highly specific in patients undergoing dialysis treatment. J. Infect. 2017, 74, 72–80. [Google Scholar] [CrossRef]
- Liss, B.; Cornely, O.A.; Hoffmann, D.; Dimitriou, V.; Wisplinghoff, H. 1,3-β-d-Glucan contamination of common antimicrobials. J. Antimicrob. Chemother. 2016, 71, 913–915. [Google Scholar] [CrossRef]
- Furfaro, E.; Mikulska, M.; Del Bono, V.; Guolo, F.; Minetto, P.; Gobbi, M.; Ghiso, A.; Bacigalupo, A.; Viscoli, C. Bloodstream Infections Are an Improbable Cause of Positive Serum (1,3)-β-d-Glucan in Hematology Patients. Clin. Vaccine Immunol. 2014, 21, 1357–1359. [Google Scholar] [CrossRef] [Green Version]
- Metan, G.; Koc, A.N.; Ağkuş, Ç.; Kaynar, L.G.; Alp, E.; Eser, B. Can bacteraemia lead to false positive results in 1,3-beta-d-glucan test? Analysis of 83 bacteraemia episodes in high-risk patients for invasive fungal infections. Rev. Iberoam. Micol. 2012, 29, 169–171. [Google Scholar] [CrossRef]
- Hammarström, H.; Kondori, N.; Friman, V.; Wennerås, C. How to interpret serum levels of beta-glucan for the diagnosis of invasive fungal infections in adult high-risk hematology patients: Optimal cut-off levels and confounding factors. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 917–925. [Google Scholar] [CrossRef]
- Finkelman, M. Comment on: 1,3-β-d-Glucan contamination of common antimicrobials. J. Antimicrob. Chemother. 2016, 71, 2996–2997. [Google Scholar] [CrossRef] [Green Version]
- Nakae, H.; Tomida, K.; Kikuya, Y.; Okuyama, M.; Igarashi, T. Comparison of quality of human serum albumin preparations in two pharmaceutical products. Acute Med. Surg. 2017, 4, 251–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, P.; Braber, S.; Garssen, J.; Wichers, H.J.; Folkerts, G.; Fink-Gremmels, J.; Varasteh, S. Beyond Heat Stress: Intestinal In-tegrity Disruption and Mechanism-Based Intervention Strategies. Nutrients 2020, 12, 734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Vitetta, L. Gut Microbiota Metabolites in NAFLD Pathogenesis and Therapeutic Implications. Int. J. Mol. Sci. 2020, 21, 5214. [Google Scholar] [CrossRef] [PubMed]
- Ramendra, R.; Isnard, S.; Mehraj, V.; Chen, J.; Zhang, Y.; Finkelman, M.; Routy, J.P. Circulating LPS and (1→3)-β-d-Glucan: A Folie à Deux Contributing to HIV-Associated Immune Activation. Front. Immunol. 2019, 18, 465. [Google Scholar] [CrossRef]
- Kavanagh, K.; Hsu, F.C.; Davis, A.T.; Kritchevsky, S.B.; Rejeski, W.J.; Kim, S. Biomarkers of leaky gut are related to inflammation and reduced physical function in older adults with cardiometabolic disease and mobility limitations. Geroscience 2019, 41, 923–933. [Google Scholar] [CrossRef]
- González-González, M.; Díaz-Zepeda, C.; Eyzaguirre-Velásquez, J.; González-Arancibia, C.; Bravo, J.A.; Julio-Pieper, M. Investigating Gut Permeability in Animal Models of Disease. Front. Physiol. 2019, 9, 1962. [Google Scholar] [CrossRef] [Green Version]
- Ellis, M.; Al-Ramadi, B.; Finkelman, M.; Hedstrom, U.; Kristensen, J.; Ali-Zadeh, H.; Klingspor, L. Assessment of the clinical utility of serial beta-d-glucan concentrations in patients with persistent neutropenic fever. J. Med. Microbiol. 2008, 57 Pt 3, 287–295. [Google Scholar] [CrossRef]
- Panpetch, W.; Chancharoenthana, W.; Bootdee, K.; Nilgate, S.; Finkelman, M.; Tumwasorn, S.; Leelahavanichkul, A. Lactobacillus rhamnosus L34 Attenuates Gut Translocation-Induced Bacterial Sepsis in Murine Models of Leaky Gut. Infect. Immun. 2017, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Issara-Amphorn, J.; Surawut, S.; Worasilchai, N.; Thim-Uam, A.; Finkelman, M.; Chindamporn, A.; Palaga, T.; Hirankarn, N.; Pisitkun, P.; Leelahavanichkul, A. The Synergy of Endotoxin and (1→3)-β-d-Glucan, from Gut Translocation, Worsens Sepsis Severity in a Lupus Model of Fc Gamma Receptor IIb-Deficient Mice. J. Innate Immun. 2018, 10, 189–201. [Google Scholar] [CrossRef]
- Shupp, J.W.; Petraitiene, R.; Jaskille, A.D.; Pavlovich, A.R.; Matt, S.E.; do Nguyen, T.; Kath, M.A.; Jeng, J.C.; Jordan, M.H.; Finkelman, M.; et al. Early serum (1→3)-β-d-glucan levels in patients with burn injury. Mycoses 2012, 55, 224–227. [Google Scholar] [CrossRef]
- Wong, J.; Zhang, Y.; Swift, O.; Finkelman, M.; Patidar, A.; Ramanarayanan, S.; Vilar, E.; Farrington, K. Beta-glucans in ad-vanced CKD: Role in endotoxaemia and inflammation. BMC Nephrol. 2020, 21, 118. [Google Scholar] [CrossRef] [PubMed]
- Ramendra, R.; Isnard, S.; Lin, J.; Fombuena, B.; Ouyang, J.; Mehraj, V.; Zhang, Y.; Finkelman, M.; Costiniuk, C.; Lebouché, B.; et al. Cytomegalovirus Seropositivity Is Associated With Increased Microbial Translocation in People Living with Human Immunodeficiency Virus and Uninfected Controls. Clin. Infect. Dis. 2020, 71, 1438–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Held, J.; Kohlberger, I.; Rappold, E.; Busse Grawitz, A.; Häcker, G. Comparison of (1→3)-β-d-glucan, mannan/anti-mannan antibodies, and Cand-Tec Candida antigen as serum biomarkers for candidemia. J. Clin. Microbiol. 2013, 51, 1158–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, M.S.; Quinn, G.; Townsend, E.C.; Ali, R.O.; Zhang, G.Y.; Bradshaw, A.; Hill, K.; Guan, H.; Hamilton, D.; Kleiner, D.E.; et al. Bacterial Translocation and Host Immune Activation in Chronic Hepatitis C Infection. Open Forum Infect. Dis. 2019, 6. [Google Scholar] [CrossRef]
- Hoenigl, M.; de Oliveira, M.F.; Pérez-Santiago, J.; Zhang, Y.; Morris, S.; McCutchan, A.J.; Finkelman, M.; Marcotte, T.D.; Ellis, R.J.; Gianella, S. (1→3)-β-d-Glucan Levels Correlate with Neurocognitive Functioning in HIV-Infected Persons on Suppressive Antiretroviral Therapy: A Cohort Study. Medicine 2016, 95, e3162. [Google Scholar] [CrossRef]
- Hoenigl, M.; Moser, C.B.; Funderburg, N.; Bosch, R.; Kantor, A.; Zhang, Y.; Eugen-Olsen, J.; Finkelman, M.; Reiser, J.; Landay, A.; et al. Adult Clinical Trials Group NWCS 411 study team. Soluble Urokinase Plasminogen Activator Receptor Is Predictive of Non-AIDS Events During Antiretroviral Therapy-mediated Viral Sup-pression. Clin. Infect. Dis. 2019, 69, 676–686. [Google Scholar] [CrossRef]
- Heyland, D.; Jiang, X.; Day, A.G.; Laverdiere, M. Serum β-d-glucan of critically ill patients with suspected ventilator-associated pneumonia: Preliminary observations. J. Crit. Care. 2011, 26, 536. [Google Scholar] [CrossRef]
- Leelahavanichkul, A.; Worasilchai, N.; Wannalerdsakun, S.; Jutivorakool, K.; Somparn, P.; Issara-Amphorn, J.; Tachaboon, S.; Srisawat, N.; Finkelman, M.; Chindamporn, A. Gastrointestinal Leakage Detected by Serum (1→3)-β-D-Glucan in Mouse Models and a Pilot Study in Patients with Sepsis. Shock 2016, 46, 506–518. [Google Scholar] [CrossRef]
- White, P.L.; Posso, R.; Parr, C.; Price, J.S.; Finkelman, M.; Barnes, R.A. The Presence of (1→3)-β-d-Glucan as Prognostic Marker in Patients After Major Abdominal Surgery. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Shah, J.; Dutta, U.; Rudramurthy, S.; Chakrabarti, A.; Sinha, S.K.; Sharma, V.; Mandavdhare, H.; Sharma, P.; Kalsi, D.; Popli, P.; et al. Colonic mucosa-associated candida assessed by biopsy culture is associated with disease severity in ulcerative colitis: A prospective study. J. Dig. Dis. 2019, 20, 642–648. [Google Scholar] [CrossRef]
- Blyth, D.M.; Chung, K.K.; Cancio, L.C.; King, B.; Murray, C.K. Clinical utility of fungal screening assays in adults with severe burns. Burns 2013, 39, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Kaita, Y.; Tarui, T.; Otsu, A.; Tanaka, Y.; Suzuki, J.; Yoshikawa, K.; Yamaguchi, Y. The Clinical Significance of Serum 1,3-β-d-Glucan for the Diagnosis of Candidemia in Severe Burn Patients. J. Burn. Care Res. 2019, 40, 104–106. [Google Scholar] [CrossRef] [PubMed]
- Carron, C.; De Barros, J.-P.P.; Gaiffe, E.; Deckert, V.; Adda-Rezig, H.; Roubiou, C.; Laheurte, C.; Masson, D.; Simula-Faivre, D.; Louvat, P.; et al. End-Stage Renal Disease-Associated Gut Bacterial Translocation: Evolution and Impact on Chronic Inflammation and Acute Rejection After Renal Transplantation. Front. Immunol. 2019, 10, 1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuipers, J.; Oosterhuis, J.K.; Paans, W.; Krijnen, W.P.; Gaillard, C.A.J.M.; Westerhuis, R.; Franssen, C.F.M. Association between quality of life and various aspects of intradialytic hypotension including patient-reported intradialytic symptom score. BMC Nephrol. 2019, 20, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, A.; Ruhe, J.; Finkelman, M.A.; Bariola, R.; Woods, G. (1→3)-β-d-Glucan (BG) Assay for Diagnosis of Invasive Fungal Infections (IFI). In Proceedings of the IDSA Annual Meeting, San Diego, CA, USA, 4–7 October 2007. [Google Scholar]
- Khan, M.A.; Nadeem, M.; Rakha, A.; Shakoor, S.; Shehzad, A.; Khan, M.R. Structural Characterization of Oat Bran (1→3), (1→4)-β-d-Glucans by Lichenase Hydrolysis Through High-Performance Anion Exchange Chromatography with Pulsed Amperometric Detection. Int. J. Food Prop. 2015, 19, 929–935. [Google Scholar] [CrossRef] [Green Version]
- Hoenigl, M.; Lin, J.; Finkelman, M.; Zhang, Y.; Karris, M.Y.; Letendre, S.L.; Ellis, R.J.; Burke, L.; Richard, B.; Gaufin, T.; et al. Glucan rich nutrition does not increase gut translocation of beta-glucan. Mycoses 2021, 64, 24–29. [Google Scholar] [CrossRef]
- Samonis, G.; Kofteridis, D.P.; Maraki, S.; Alegakis, D.; Mantadakis, E.; Papadakis, J.A.; Gikas, A.H.; Falagas, M.E. Levofloxacin and moxifloxacin increase human gut colonization by Candida species. Antimicrob. Agents Chemother. 2005, 49, 5189. [Google Scholar] [CrossRef] [Green Version]
- Zhai, B.; Ola, M.; Rolling, T.; Tosini, N.L.; Joshowitz, S.; Littmann, E.R.; Amoretti, L.A.; Fontana, E.; Wright, R.J.; Miranda, E.; et al. High-resolution mycobiota analysis reveals dynamic intestinal translocation preceding invasive candidiasis. Nat. Med. 2020, 26, 59–64. [Google Scholar] [CrossRef]
- Yang, A.-M.; Inamine, T.; Hochrath, K.; Chen, P.; Wang, L.; Llorente, C.; Bluemel, S.; Hartmann, P.; Xu, J.; Koyama, Y.; et al. Intestinal fungi contribute to development of alcoholic liver disease. J. Clin. Investig. 2017, 127, 2829–2841. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Roth, R.I.; Grunfeld, C.; Feingold, K.R.; Levin, J. Pharmacokinetics, biological effects, and distribution of (1→3)-beta-d-glucan in blood and organs in rabbits. Mediat. Inflamm. 1997, 6, 279–283. [Google Scholar] [CrossRef]
- Sanada, Y.; Mizuta, K.; Urahashi, T.; Ihara, Y.; Wakiya, T.; Okada, N.; Yamada, N.; Yasuda, Y.; Kawarasaki, H. The efficacy of measurement of the serum beta-D glucan in thepatients with biliary atresia. Pediatr. Surg. Int. 2012, 28, 993–996. [Google Scholar] [CrossRef] [PubMed]
- Sanada, Y.; Urahashi, T.; Ihara, Y.; Wakiya, T.; Okada, N.; Yamada, N.; Hirata, Y.; Mizuta, K. Impact of 3-D glucan during liver transplantation. Hepatogastroenterology 2014, 61, 1368–1373. [Google Scholar] [PubMed]
- Verstrepen, B.E.; Nieuwenhuis, I.G.; Mooij, P.; Verschoor, E.J.; Fagrouch, Z.C.; Kondova, I.; Boonstra, A.; Koopman, G. Role of microbial translocation in soluble CD14m up-regulation in HIV-, but not in HCV-, infected chimpanzees. J. Gen. Virol. 2016, 97, 2599–2607. [Google Scholar] [CrossRef] [PubMed]
- Albert, O.; Toubas, D.; Strady, C.; Cousson, J.; Delmas, C.; Vernet, V.; Villena, I. Reactivity of (1→3)-β-d-glucan assay in bacterial bloodstream infections. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 1453–1460. [Google Scholar] [CrossRef]
- Mennink-Kersten, M.A.; Ruegebrink, D.; Verweij, P.E. Pseudomonas aeruginosa as a cause of 1,3-beta-d-glucan assay reactivity. Clin. Infect. Dis. 2008, 46, 1930–1931. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, S.M.; Falentin, H.; Dols-Lafargue, M.; Lapointe, G.; Roy, D. Capsular exopolysaccharide biosynthesis gene of Propionibacterium freudenreichii subsp.shermanii. Int. J. Food Microbiol. 2008, 125, 252–258. [Google Scholar] [CrossRef]
- Ramos, Y.; Rocha, J.; Hael, A.L.; van Gestel, J.; Vlamakis, H.; Cywes-Bentley, C.; Cubillos-Ruiz, J.R.; Pier, G.B.; Gilmore, M.S.; Kolter, R.; et al. PolyGlcNAc-containing exopolymers enable surface penetration by non-motile Enterococcus faecalis. PLoS Pathog. 2019, 15, e1007571. [Google Scholar] [CrossRef] [Green Version]
- Sawai, T.; Nakao, T.; Yamaguchi, S.; Yoshioka, S.; Matsuo, N.; Suyama, N.; Yanagihara, K.; Mukae, H. Detection of high se-rum levels of β-d-Glucan in disseminated nocardial infection: A case report. BMC Infect. Dis. 2017, 17, 272. [Google Scholar] [CrossRef]
- Nakamura, I.; Nagakura, T.; Fujita, H.; Fukusima, S.; Gonoi, T. Nocardia elegans infection: A case report and literature review. Int. J. Infect. Dis. 2017, 54, 15–17. [Google Scholar] [CrossRef] [Green Version]
- Koncan, R.; Favuzzi, V.; Ligozzi, M.; Sorrentino, A.; Cornaglia, G.; Cascio, G.L. Cross-reactivity of Nocardia spp. in the fungal (1-3)-β-d-glucan assay performed on cerebral spinal fluid. Diagn. Microbiol. Infect. Dis. 2015, 81, 94–95. [Google Scholar] [CrossRef]
- Lequette, Y.; Ödberg-Ferragut, C.; Bohin, J.-P.; Lacroix, J.-M. Identification of mdoD, an mdoG Paralog Which Encodes a Twin-Arginine-Dependent Periplasmic Protein That Controls Osmoregulated Periplasmic Glucan Backbone Structures. J. Bacteriol. 2004, 186, 3695–3702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debarbieux, L.; Bohin, A.; Bohin, J.P. Topological analysis of the membrane-bound glucosyltransferase, MdoH, required for osmoregulated periplasmic glucan synthesis in Escherichia coli. J. Bacteriol. 1997, 179, 6692–6698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntosh, M.; Stone, B.A.; Stanisich, V.A. Curdlan and other bacterial (1→3)-beta-d-glucans. Appl. Microbiol. Biotechnol. 2005, 68, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Buchacher, A.; Krause, D.; Wiry, G.; Weinberger, J. Elevated Endotoxin Levels in Human Intravenous Immunoglobulin Concentrates Caused by (1→3)-{beta}-d-Glucans. PDA J. Pharm. Sci. Technol. 2011, 64, 536–544. [Google Scholar]
- Voigt, C.A. Callose-mediated resistance to pathogenic intruders in plant defense-related papillae. Front. Plant. Sci. 2014, 5, 168. [Google Scholar] [CrossRef] [PubMed]
- Usami, M.; Ohata, A.; Horiuchi, T.; Nagasawa, K.; Wakabayashi, T.; Tanaka, S. Positive (1→3)-beta-d-glucan in blood com-ponents and release of (1→3)-beta-d-glucan from depth-type membrane filters for blood processing. Transfusion 2002, 42, 1189–1195. [Google Scholar] [CrossRef]
- Filtration of liquids using cellulose-based depth filters. Technical Report no 45. PDA J. Pharm. Sci. Technol. 2008, 62 (Suppl. 2), 2–35.
- Ohata, A.; Usami, M.; Horiuchi, T.; Nagasawa, K.; Kinoshita, K. Release of (13)-beta-d-Glucan from Depth-type Membrane Filters and Their In Vitro Effects on Proinflammatory Cytokine Production. Artif. Organs 2003, 27, 728–735. [Google Scholar] [CrossRef]
- Hussain, K.K.; Malavia, D.; Johnson, E.M.; Littlechild, J.; Winlove, C.P.; Vollmer, F.; Gow, N.A.R. Biosensors and Diagnostics for Fungal Detection. J. Fungi 2020, 6, 349. [Google Scholar] [CrossRef]
- Boch, T.; Spiess, B.; Cornely, O.; Vehreschild, J.; Rath, P.; Steinmann, J.; Heinz, W.; Hahn, J.; Krause, S.; Kiehl, M.; et al. Diagnosis of invasive fungal infections in haematological patients by combined use of galactomannan, 1,3-β-d-glucan, Aspergillus PCR, multifungal DNA-microarray, and Aspergillus azole resistance PCRs in blood and bronchoalveolar lavage samples: Results of a prospective multicentre study. Clin. Microbiol. Infect. 2016, 22, 862–868. [Google Scholar] [CrossRef] [Green Version]
- Qian, K.-Y.; Lyu, Q.-X.; Dai, L.-J.; Chang, H.-R.; Wu, D.-P.; Sun, A.-N. Diagnostic Value of G and GM Tests in Acute Leukemia Patients with Invasive Fungal Disease. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2019, 27, 1678–1681. [Google Scholar] [PubMed]
- Urabe, N.; Sakamoto, S.; Sano, G.; Suzuki, J.; Hebisawa, A.; Nakamura, Y.; Koyama, K.; Ishii, Y.; Tateda, K.; Homma, S. Use-fulness of Two Aspergillus PCR Assays and Aspergillus Galactomannan and β-d-Glucan Testing of Bronchoalveolar Lavage Fluid for Diagnosis of Chronic Pulmonary Aspergillosis. J. Clin. Microbiol. 2017, 55, 1738–1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenks, J.D.; Gangneux, J.P.; Schwartz, I.S.; Alastruey-Izquierdo, A.; Lagrou, K.; Thompson, G.R.; Lass-Flörl, C.; Hoenigl, M. Investigators ECOMMEC. Diagnosis of Breakthrough Fungal Infections in the Clinical Mycology Laboratory: An ECMM Consensus Statement. J. Fungi 2020, 6, 216. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Limulus amebocyte lysate cascade.

Figure 2.
Kinetic analysis of BDG activation of LAL cascade [Unpublished data, Associates of Cape Cod, Inc.]. BDG: (1→3)-β-glucan; LAL: Limulus amebocyte lysate.
Figure 2.
Kinetic analysis of BDG activation of LAL cascade [Unpublished data, Associates of Cape Cod, Inc.]. BDG: (1→3)-β-glucan; LAL: Limulus amebocyte lysate.

Figure 3.
Fungal BDG: (1→3)-β-glucan backbone with (1→6)-β-linked glucose.

Figure 4.
Pre-and post IVg infusion effect upon serum BDG titer [Unpublished data, Associates of Cape Cod, Inc.].
Figure 4.
Pre-and post IVg infusion effect upon serum BDG titer [Unpublished data, Associates of Cape Cod, Inc.].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Medical products and processes associated with patient/product contamination.
| Material | Reference |
|---|---|
| Gauze [63] | Kanamori, H. Tohoku J. Exp. Med. 2009, 217, 117–121. |
| Surgical sponge [60] | Stycznski, A. J. Fungi (Basel) 2018, 4, E114. |
| Process equipment [56] | Vigor, K. Biotechnol. Prog. 2016, 32, 1494–1502 |
| Fermentation media [58] | Gefroh, E. Biotechnol. Prog. 2013, 29, 672–680. |
| Cellulosic depth filters [57] | Holstein, M. Biotechnol. Prog. 2020, e3086 |
| Hemodialysis with cellulosic membranes [65] | Kanda, H. Kidney Int. 2001, 60, 319–323. |
| Intravenous immunoglobulin [52,59] | Angebault, C. Open Forum Infect. Dis. 2016, 3, ofw128.Duffner, U. Bone Marrow Transpl. 2012, 47, 151–152. |
| Human serum albumin [72] | Nakae, H. Acute Med. Surg. 2017, 4, 251–254 |
| Anti-tumor adjuvant [64] | Tokuyasu, U.H. Int. J. Gen. Med. 2010, 3, 273–277. |
| Antibiotics [67,68] | Liss, B. J. Antimicrob. Chemother. 2016, 71, 913–915.Furfaro, E. Clin. Vaccine Immunol. 2014, 21, 1357–1359. |
Table 2.
Conditions associated with elevated circulating BDG.
| Clinical Setting | Reference |
|---|---|
| Burns, large surface area [81] | Shupp, J. Mycoses 2011, 55, 224–227. |
| Chronic kidney disease [82] | Wong, J. BMC Nephrol. 2020, 21, 118. |
| Cystic fibrosis [44] | Rautemaa, V. DMID 2017, 88, 16–21. |
| Cytomegalovirus infection [83] | Ramendra, R. CID 2019, ePub. |
| Enteroccocemia [84] | Held, J. et al. J. Clin. Microbiol. 2013, 51, 1158–1164. |
| Hepatitis C virus [85] | Moon, M.S. Open Forum Inf. Dis. 2019, ePub. |
| HIV neurocognitive decline [86] | Hoenigl, M. Medicine 2016, 95. |
| HIV non-AIDS-related adverse events [87] | Hoenigl, M. CID 2019, 69, 676–686. |
| Invasive mechanical ventilation [88] | Heyland, D. J. Crit. Care 2011, 26, 536.e1–536.e9 |
| Lupus erythematosus [80] | Issara-Amphorn, J. J. Innate Immun. 2018, 10, 18. |
| Sepsis-septic shock transition [89] | Leelahavanichkul, A. Shock 2016, 46, 506–518. |
| Post-major abdom. Surgery [90] | White, P.L. CID 2020, In Press. |
| Ulcerative colitis [91] | Shah, J. J. Dig. Dis. 2019, 20, 642–648. |
Table 3.
Major factors to consider in investigating suspected BDG false positives.
| A. Medical Product-Related False Positives | |
| 1. | Has the patient received IVIg infusions? |
| 2. | Has the patient received human serum albumin infusions? |
| 3. | Has the patient received total parenteral nutrition? |
| 4. | Has the patient undergone invasive surgery within the past 4 days? |
| 5. | Does the patient have indwelling gauze, surgical sponges packings? |
| 6. | Are any other invasive cellulosic medical devices in use? |
| B. Medical Condition-Related False Positives | |
| 1. | Does the patient have evidence of severe mucositis or enterocolitis? |
| 2. | Is the patient on hemodialysis? |
| 3. | Does the patient have invasive nocardiosis? |
| 4. | Is gut ischemia or hypoxia suspected? |
| 5. | Has pneumocystosis been ruled out? |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Finkelman, M.A. Specificity Influences in (1→3)-β-d-Glucan-Supported Diagnosis of Invasive Fungal Disease. J. Fungi 2021, 7, 14. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010014
AMA Style
Finkelman MA. Specificity Influences in (1→3)-β-d-Glucan-Supported Diagnosis of Invasive Fungal Disease. Journal of Fungi. 2021; 7(1):14. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010014
Chicago/Turabian StyleFinkelman, Malcolm A. 2021. "Specificity Influences in (1→3)-β-d-Glucan-Supported Diagnosis of Invasive Fungal Disease" Journal of Fungi 7, no. 1: 14. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010014
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.