Carbon and Nitrogen Sources Have No Impact on the Organization and Composition of Ustilago maydis Respiratory Supercomplexes

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Isolation of Mitochondria
2.3. Sample Preparation for Native Electrophoresis
2.4. In-Gel Enzyme Activity Assays
2.5. 2D Tricine-SDS Gel Electrophoresis and Western Blot Analysis
2.6. Tandem Mass Spectrometry (LC/ESI-MS/MS)
2.7. Determination of Protein Content
2.8. Materials
3. Results
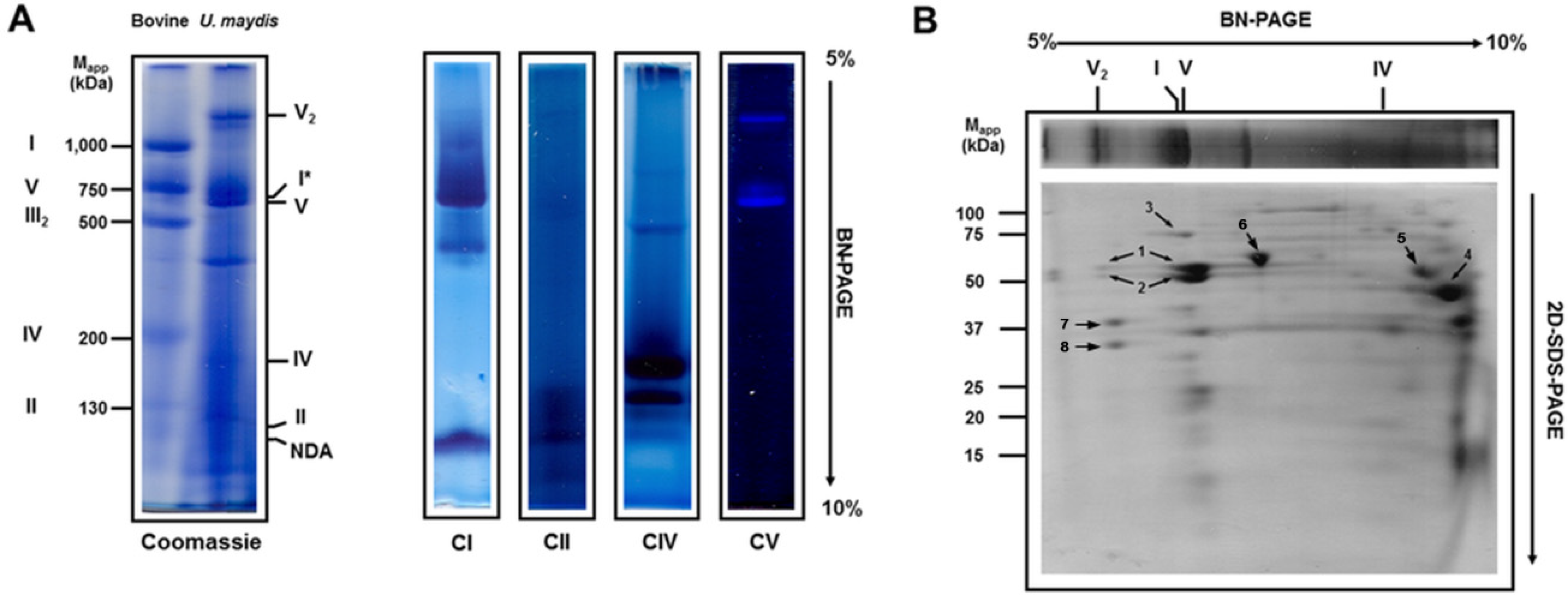
3.1. Solubilization of Mitochondrial Respiratory Proteins by Dodecyl-β-d-Maltoside
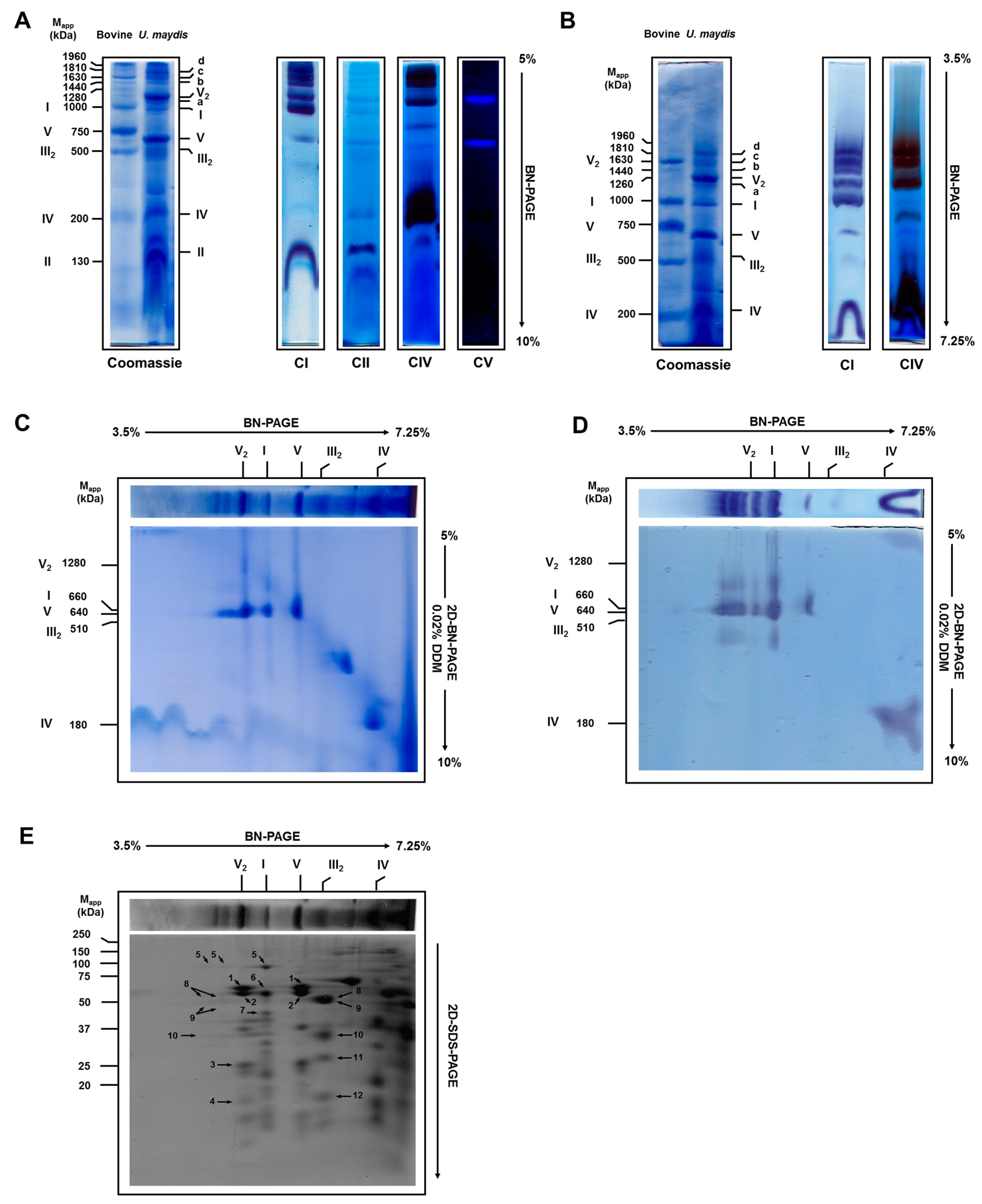
3.2. Solubilization of Mitochondrial Respiratory Complexes with Digitonin
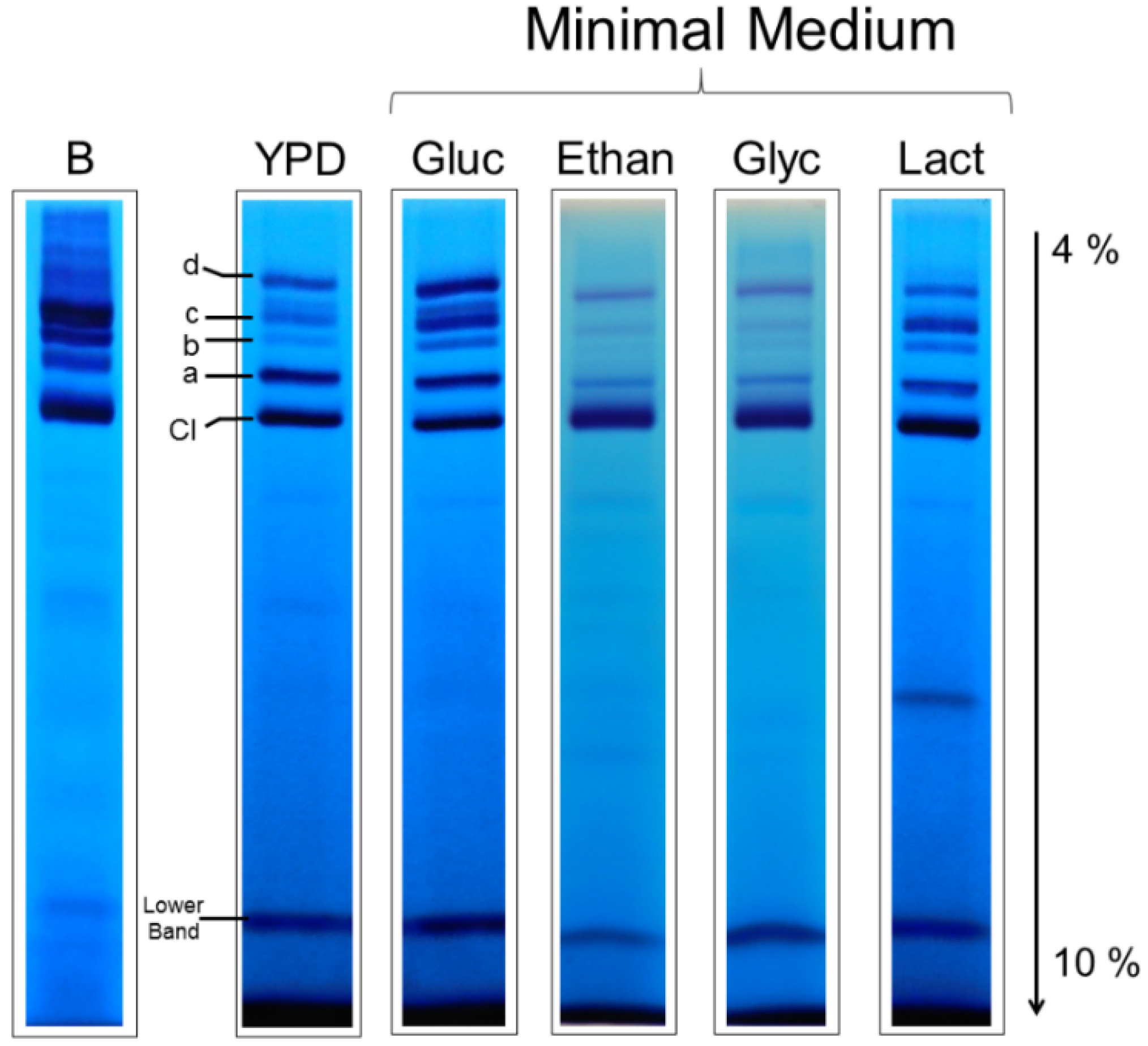
3.3. Respiratory Supercomplexes in Cells Growing at Different Rates
4. Discussion
4.1. The Respiratory Complexes in Ustilago maydis
4.2. Association of Other Elements with the Respiratory Complexes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Juarez, O.; Guerra, G.; Martinez, F.; Pardo, J.P. The Mitochondrial Respiratory Chain of Ustilago maydis. Biochim. Biophys. Acta 2004, 1658, 244–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matuz-Mares, D.; Matus-Ortega, G.; Cardenas-Monroy, C.; Romero-Aguilar, L.; Villalobos-Rocha, J.C.; Vazquez-Meza, H.; Guerra-Sanchez, G.; Pena-Diaz, A.; Pardo, J.P. Expression of Alternative NADH Dehydrogenases (NDH-2) in the Phytopathogenic Fungus Ustilago maydis. FEBS Open Bio 2018, 8, 1267–1279. [Google Scholar] [CrossRef] [PubMed]
- Sierra-Campos, E.; Velázquez, I.; Matuz-Mares, D.; Villavicencio-Queijeiro, A.; Pardo, J.P. Functional Properties of the Ustilago Maydis Alternative Oxidase under Oxidative Stress Conditions. Mitochondrion 2009, 9, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Vonck, J.; Schäfer, E. Supramolecular Organization of Protein Complexes in the Mitochondrial Inner Membrane. Biochim. Biophys. Acta 2009, 1793, 117–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schägger, H.; Pfeiffer, K. Supercomplexes in the Respiratory Chains of Yeast and Mammalian Mitochondria. Embo J. 2000, 19, 1777–1783. [Google Scholar] [CrossRef] [Green Version]
- Eubel, H.; Jänsch, L.; Braun, H.-P. New Insights into the Respiratory Chain of Plant Mitochondria. Supercomplexes and a Unique Composition of Complex II. Plant Physiol. 2003, 133, 274–286. [Google Scholar] [CrossRef] [Green Version]
- Marques, I.; Dencher, N.A.; Videira, A.; Krause, F. Supramolecular Organization of the Respiratory Chain in Neurospora Crassa Mitochondria. Eukaryot Cell 2007, 6, 2391–2405. [Google Scholar] [CrossRef] [Green Version]
- Schägger, H.; Pfeiffer, K. The Ratio of Oxidative Phosphorylation Complexes I-V in Bovine Heart Mitochondria and the Composition of Respiratory Chain Supercomplexes. J. Biol. Chem. 2001, 276, 37861–37867. [Google Scholar] [CrossRef]
- Guo, R.; Zong, S.; Wu, M.; Gu, J.; Yang, M. Architecture of Human Mitochondrial Respiratory Megacomplex I2III2IV2. Cell 2017, 170, 1247–1257.e12. [Google Scholar] [CrossRef] [Green Version]
- Letts, J.A.; Fiedorczuk, K.; Degliesposti, G.; Skehel, M.; Sazanov, L.A. Structures of Respiratory Supercomplex I+III2 Reveal Functional and Conformational Crosstalk. Mol. Cell 2019, 75, 1131–1146.e6. [Google Scholar] [CrossRef] [Green Version]
- Letts, J.A.; Fiedorczuk, K.; Sazanov, L.A. The Architecture of Respiratory Supercomplexes. Nature 2016, 537, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Gu, J.; Guo, R.; Huang, Y.; Yang, M. Structure of Mammalian Respiratory Supercomplex I1III2IV1. Cell 2016, 167, 1598–1609.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartley, A.M.; Lukoyanova, N.; Zhang, Y.; Cabrera-Orefice, A.; Arnold, S.; Meunier, B.; Pinotsis, N.; Maréchal, A. Structure of Yeast Cytochrome c Oxidase in a Supercomplex with Cytochrome Bc1. Nat. Struct. Mol. Biol. 2019, 26, 78–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiseman, B.; Nitharwal, R.G.; Fedotovskaya, O.; Schäfer, J.; Guo, H.; Kuang, Q.; Benlekbir, S.; Sjöstrand, D.; Ädelroth, P.; Rubinstein, J.L.; et al. Structure of a Functional Obligate Complex III2IV2 Respiratory Supercomplex from Mycobacterium Smegmatis. Nat. Struct. Mol. Biol. 2018, 25, 1128–1136. [Google Scholar] [CrossRef]
- Genova, M.L.; Baracca, A.; Biondi, A.; Casalena, G.; Faccioli, M.; Falasca, A.I.; Formiggini, G.; Sgarbi, G.; Solaini, G.; Lenaz, G. Is Supercomplex Organization of the Respiratory Chain Required for Optimal Electron Transfer Activity? Biochim. Biophys. Acta 2008, 1777, 740–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acín-Pérez, R.; Bayona-Bafaluy, M.P.; Fernández-Silva, P.; Moreno-Loshuertos, R.; Pérez-Martos, A.; Bruno, C.; Moraes, C.T.; Enríquez, J.A. Respiratory Complex III Is Required to Maintain Complex I in Mammalian Mitochondria. Mol. Cell 2004, 13, 805–815. [Google Scholar] [CrossRef]
- Maranzana, E.; Barbero, G.; Falasca, A.I.; Lenaz, G.; Genova, M.L. Mitochondrial Respiratory Supercomplex Association Limits Production of Reactive Oxygen Species from Complex I. Antioxid. Redox Signal. 2013, 19, 1469–1480. [Google Scholar] [CrossRef] [Green Version]
- Holliday, R. Ustilago maydis. In Handbook of Genetics: Volume 1 Bacteria, Bacteriophages, and Fungi; King, R.C., Ed.; Handbook of Genetics; Springer: New York, NY, USA, 1974; Volume I, pp. 575–595. ISBN 978-0-306-37611-5. [Google Scholar]
- Schagger, H.; Cramer, W.A.; von Jagow, G. Analysis of Molecular Masses and Oligomeric States of Protein Complexes by Blue Native Electrophoresis and Isolation of Membrane Protein Complexes by Two-Dimensional Native Electrophoresis. Anal. Biochem. 1994, 217, 220–230. [Google Scholar] [CrossRef]
- Jung, C.; Higgins, C.M.; Xu, Z. Measuring the Quantity and Activity of Mitochondrial Electron Transport Chain Complexes in Tissues of Central Nervous System Using Blue Native Polyacrylamide Gel Electrophoresis. Anal. Biochem. 2000, 286, 214–223. [Google Scholar] [CrossRef]
- Wittig, I.; Karas, M.; Schagger, H. High Resolution Clear Native Electrophoresis for In-Gel Functional Assays and Fluorescence Studies of Membrane Protein Complexes. Mol. Cell. Proteom. MCP 2007, 6, 1215–1225. [Google Scholar] [CrossRef] [Green Version]
- Wittig, I.; Carrozzo, R.; Santorelli, F.M.; Schägger, H. Functional Assays in High-Resolution Clear Native Gels to Quantify Mitochondrial Complexes in Human Biopsies and Cell Lines. Electrophoresis 2007, 28, 3811–3820. [Google Scholar] [CrossRef] [PubMed]
- Wittig, I.; Beckhaus, T.; Wumaier, Z.; Karas, M.; Schägger, H. Mass Estimation of Native Proteins by Blue Native Electrophoresis: Principles and Practical Hints. Mol. Cell. Proteom. 2010, 9, 2149–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Zamorano, M.; Mendoza-Hernández, G.; Xolalpa, W.; Parada, C.; Vallecillo, A.J.; Bigi, F.; Espitia, C. Mycobacterium tuberculosis Glycoproteomics Based on ConA-Lectin Affinity Capture of Mannosylated Proteins. J. Proteome Res. 2009, 8, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Bensadoun, A.; Weinstein, D. Assay of Proteins in the Presence of Interfering Materials. Anal. Biochem. 1976, 70, 241–250. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Claros, M.G.; Vincens, P. Computational Method to Predict Mitochondrially Imported Proteins and Their Targeting Sequences. Eur. J. Biochem. 1996, 241, 779–786. [Google Scholar] [CrossRef]
- Marques, I.; Duarte, M.; Assunção, J.; Ushakova, A.V.; Videira, A. Composition of Complex I from Neurospora Crassa and Disruption of Two “Accessory” Subunits. Biochim. Biophys. Acta 2005, 1707, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Wirth, C.; Brandt, U.; Hunte, C.; Zickermann, V. Structure and Function of Mitochondrial Complex I. Biochim. Biophys. Acta 2016, 1857, 902–914. [Google Scholar] [CrossRef]
- Castello, P.R.; Woo, D.K.; Ball, K.; Wojcik, J.; Liu, L.; Poyton, R.O. Oxygen-Regulated Isoforms of Cytochrome c Oxidase Have Differential Effects on Its Nitric Oxide Production and on Hypoxic Signaling. Proc. Natl. Acad. Sci. USA 2008, 105, 8203–8208. [Google Scholar] [CrossRef] [Green Version]
- Cardenas-Monroy, C.A.; Pohlmann, T.; Pinon-Zarate, G.; Matus-Ortega, G.; Guerra, G.; Feldbrugge, M.; Pardo, J.P. The Mitochondrial Alternative Oxidase Aox1 Is Needed to Cope with Respiratory Stress but Dispensable for Pathogenic Development in Ustilago maydis. PLoS ONE 2017, 12, e0173389. [Google Scholar] [CrossRef]
- Reyes-Galindo, M.; Suarez, R.; Esparza-Perusquia, M.; de Lira-Sanchez, J.; Pardo, J.P.; Martinez, F.; Flores-Herrera, O. Mitochondrial Respirasome Works as a Single Unit and the Cross-Talk between Complexes I, III2 and IV Stimulates NADH Dehydrogenase Activity. Biochim. Biophys. Acta Bioenerg. 2019, 1860, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Tuschen, G.; Sackmann, U.; Nehls, U.; Haiker, H.; Buse, G.; Weiss, H. Assembly of NADH: Ubiquinone Reductase (Complex I) in Neurospora Mitochondria: Independent Pathways of Nuclear-Encoded and Mitochondrially Encoded Subunits. J. Mol. Biol. 1990, 213, 845–857. [Google Scholar] [CrossRef]
- Lazarou, M.; Thorburn, D.R.; Ryan, M.T.; McKenzie, M. Assembly of Mitochondrial Complex I and Defects in Disease. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2009, 1793, 78–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero-Castillo, S.; Vazquez-Acevedo, M.; Gonzalez-Halphen, D.; Uribe-Carvajal, S. In Yarrowia Lipolytica Mitochondria, the Alternative NADH Dehydrogenase Interacts Specifically with the Cytochrome Complexes of the Classic Respiratory Pathway. Biochim. Biophys. Acta 2009, 1787, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Formosa, L.E.; Dibley, M.G.; Stroud, D.A.; Ryan, M.T. Building a Complex Complex: Assembly of Mitochondrial Respiratory Chain Complex I. Semin. Cell Dev. Biol. 2018, 76, 154–162. [Google Scholar] [CrossRef]
- Krause, F.; Scheckhuber, C.Q.; Werner, A.; Rexroth, S.; Reifschneider, N.H.; Dencher, N.A.; Osiewacz, H.D. Supramolecular Organization of Cytochrome c Oxidase- and Alternative Oxidase-Dependent Respiratory Chains in the Filamentous Fungus Podospora anserina. J. Biol. Chem. 2004, 279, 26453–26461. [Google Scholar] [CrossRef] [Green Version]
- Schägger, H. Respiratory Chain Supercomplexes. IUBMB Life 2001, 52, 119–128. [Google Scholar] [CrossRef]
- Davies, K.M.; Blum, T.B.; Kühlbrandt, W. Conserved in Situ Arrangement of Complex I and III2 in Mitochondrial Respiratory Chain Supercomplexes of Mammals, Yeast, and Plants. Proc. Natl. Acad. Sci. USA 2018, 115, 3024–3029. [Google Scholar] [CrossRef] [Green Version]
- Cabrera-Orefice, A.; Chiquete-Félix, N.; Espinasa-Jaramillo, J.; Rosas-Lemus, M.; Guerrero-Castillo, S.; Peña, A.; Uribe-Carvajal, S. The Branched Mitochondrial Respiratory Chain from Debaryomyces Hansenii: Components and Supramolecular Organization. Biochim. Biophys. Acta 2014, 1837, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Blaza, J.N.; Serreli, R.; Jones, A.J.Y.; Mohammed, K.; Hirst, J. Kinetic Evidence against Partitioning of the Ubiquinone Pool and the Catalytic Relevance of Respiratory-Chain Supercomplexes. Proc. Natl. Acad. Sci. USA 2014, 111, 15735–15740. [Google Scholar] [CrossRef] [Green Version]
- Fedor, J.G.; Hirst, J. Mitochondrial Supercomplexes Do Not Enhance Catalysis by Quinone Channeling. Cell Metab. 2018, 28, 525–531.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berndtsson, J.; Aufschnaiter, A.; Rathore, S.; Marin-Buera, L.; Dawitz, H.; Diessl, J.; Kohler, V.; Barrientos, A.; Büttner, S.; Fontanesi, F.; et al. Respiratory Supercomplexes Enhance Electron Transport by Decreasing Cytochrome c Diffusion Distance. EMBO Rep. 2020, e51015. [Google Scholar] [CrossRef] [PubMed]
- Stuchebrukhov, A.; Schäfer, J.; Berg, J.; Brzezinski, P. Kinetic Advantage of Forming Respiratory Supercomplexes. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148193. [Google Scholar] [CrossRef] [PubMed]
- Esparza-Perusquia, M.; Olvera-Sanchez, S.; Pardo, J.P.; Mendoza-Hernandez, G.; Martinez, F.; Flores-Herrera, O. Structural and Kinetics Characterization of the F1F0-ATP Synthase Dimer. New Repercussion of Monomer-Monomer Contact. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 975–981. [Google Scholar] [CrossRef] [PubMed]
- González-Barroso, M.M.; Ledesma, A.; Lepper, S.; Pérez-Magán, E.; Zaragoza, P.; Rial, E. Isolation and Bioenergetic Characterization of Mitochondria from Pichia Pastoris. Yeast 2006, 23, 307–313. [Google Scholar] [CrossRef]
- Martins, V.P.; Soriani, F.M.; Magnani, T.; Tudella, V.G.; Goldman, G.H.; Curti, C.; Uyemura, S.A. Mitochondrial Function in the Yeast Form of the Pathogenic Fungus Paracoccidioides Brasiliensis. J. Bioenerg. Biomembr. 2008, 40, 297–305. [Google Scholar] [CrossRef]
- Medentsev, A.G.; Arinbasarova, A.Y.; Golovchenko, N.P.; Akimenko, V.K. Involvement of the Alternative Oxidase in Respiration of Yarrowia Lipolytica Mitochondria Is Controlled by the Activity of the Cytochrome Pathway. FEMS Yeast Res. 2002, 2, 519–524. [Google Scholar] [CrossRef] [Green Version]
- Ukolova, I.V.; Kondakova, M.A.; Kondratov, I.G.; Sidorov, A.V.; Borovskii, G.B.; Voinikov, V.K. New Insights into the Organisation of the Oxidative Phosphorylation System in the Example of Pea Shoot Mitochondria. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148264. [Google Scholar] [CrossRef]
- Eubel, H.; Heinemeyer, J.; Braun, H.-P. Identification and Characterization of Respirasomes in Potato Mitochondria. Plant Physiol. 2004, 134, 1450–1459. [Google Scholar] [CrossRef] [Green Version]
- Tisdale, H.D. [39] Preparation and properties of succinic—cytochrome c reductase (complex II–III). In Methods in Enzymology; Oxidation and Phosphorylation; Academic Press: Cambridge, MA, USA, 1967; Volume 10, pp. 213–215. [Google Scholar]
- Bruel, C.; Brasseur, R.; Trumpower, B.L. Subunit 8 of the Saccharomyces Cerevisiae Cytochrome Bc1 Complex Interacts with Succinate-Ubiquinone Reductase Complex. J. Bioenerg. Biomembr. 1996, 28, 59–68. [Google Scholar] [CrossRef]
- Grandier-Vazeille, X.; Bathany, K.; Chaignepain, S.; Camougrand, N.; Manon, S.; Schmitter, J.M. Yeast Mitochondrial Dehydrogenases Are Associated in a Supramolecular Complex. Biochemistry 2001, 40, 9758–9769. [Google Scholar] [CrossRef]
- Ardehali, H.; Chen, Z.; Ko, Y.; Mejía-Alvarez, R.; Marbán, E. Multiprotein Complex Containing Succinate Dehydrogenase Confers Mitochondrial ATP-Sensitive K+ Channel Activity. Proc. Natl. Acad. Sci. USA 2004, 101, 11880–11885. [Google Scholar] [CrossRef] [Green Version]
- Acín-Pérez, R.; Fernández-Silva, P.; Peleato, M.L.; Pérez-Martos, A.; Enriquez, J.A. Respiratory Active Mitochondrial Supercomplexes. Mol. Cell 2008, 32, 529–539. [Google Scholar] [CrossRef]
- Boumans, H.; Grivell, L.A.; Berden, J.A. The Respiratory Chain in Yeast Behaves as a Single Functional Unit. J. Biol. Chem. 1998, 273, 4872–4877. [Google Scholar] [CrossRef] [Green Version]
- Matus-Ortega, M.G.; Cardenas-Monroy, C.A.; Flores-Herrera, O.; Mendoza-Hernandez, G.; Miranda, M.; Gonzalez-Pedrajo, B.; Vazquez-Meza, H.; Pardo, J.P. New Complexes Containing the Internal Alternative NADH Dehydrogenase (Ndi1) in Mitochondria of Saccharomyces Cerevisiae. Yeast 2015, 32, 629–641. [Google Scholar] [CrossRef]
- Nübel, E.; Wittig, I.; Kerscher, S.; Brandt, U.; Schägger, H. Two-Dimensional Native Electrophoretic Analysis of Respiratory Supercomplexes from Yarrowia Lipolytica. Proteomics 2009, 9, 2408–2418. [Google Scholar] [CrossRef]
- Navet, R.; Jarmuszkiewicz, W.; Douette, P.; Sluse-Goffart, C.M.; Sluse, F.E. Mitochondrial Respiratory Chain Complex Patterns from Acanthamoeba Castellanii and Lycopersicon Esculentum: Comparative Analysis by BN-PAGE and Evidence of Protein-Protein Interaction between Alternative Oxidase and Complex III. J. Bioenerg. Biomembr. 2004, 36, 471–479. [Google Scholar] [CrossRef]
- Duarte, M.; Videira, A. Effects of Mitochondrial Complex III Disruption in the Respiratory Chain of Neurospora Crassa. Mol. Microbiol. 2009, 72, 246–258. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Band | Experimental Molecular Mass (kDa) | Theoretical Molecular Mass (kDa) | Subunit Identity | Gene Name |
|---|---|---|---|---|
| I | 980 | 877 | 75 kDa subunit (5) 51 kDa subunit (6) 40 kDa subunit (7) | UMAG_10695 UMAG_11170 UMAG_00381 |
| II | 139 | 117 | ||
| III | 510 | 474 | QCR2 (8) QCR1 (9) Cytochrome c1 (10) Rieske protein (11) QCR7 (12) | UMAG_01478 UMAG_11590 UMAG_11534 UMAG_10507 UMAG_04237 |
| IV | 240 | 203 | ||
| V | 640 | 599 | α subunit (1) β subunit (2) d subunit (3) g subunit (4) δ subunit (4) | UMAG_10213 UMAG_10397 UMAG_12050 UMAG_00975 UMAG_01103 |
| Band * | Experimental Molecular Mass (kDa) | Calculated Molecular Mass (kDa) | Suggested Stoichiometry |
|---|---|---|---|
| a | 1200 | 1220 | I1:IV1 |
| V2 | 1260 | 1280 | |
| b | 1440 | 1490 | I1:III2 |
| c | 1630 | 1730 | I1:III2:IV1 |
| d | 1810 | 1970 | I1:III2:IV2, I2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matuz-Mares, D.; Flores-Herrera, O.; Guerra-Sánchez, G.; Romero-Aguilar, L.; Vázquez-Meza, H.; Matus-Ortega, G.; Martínez, F.; Pardo, J.P. Carbon and Nitrogen Sources Have No Impact on the Organization and Composition of Ustilago maydis Respiratory Supercomplexes. J. Fungi 2021, 7, 42. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010042
Matuz-Mares D, Flores-Herrera O, Guerra-Sánchez G, Romero-Aguilar L, Vázquez-Meza H, Matus-Ortega G, Martínez F, Pardo JP. Carbon and Nitrogen Sources Have No Impact on the Organization and Composition of Ustilago maydis Respiratory Supercomplexes. Journal of Fungi. 2021; 7(1):42. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010042
Chicago/Turabian StyleMatuz-Mares, Deyamira, Oscar Flores-Herrera, Guadalupe Guerra-Sánchez, Lucero Romero-Aguilar, Héctor Vázquez-Meza, Genaro Matus-Ortega, Federico Martínez, and Juan Pablo Pardo. 2021. "Carbon and Nitrogen Sources Have No Impact on the Organization and Composition of Ustilago maydis Respiratory Supercomplexes" Journal of Fungi 7, no. 1: 42. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010042