
Halophytophthora fluviatilis Pathogenicity and Distribution along a Mediterranean-Subalpine Gradient

 , , and
, , and 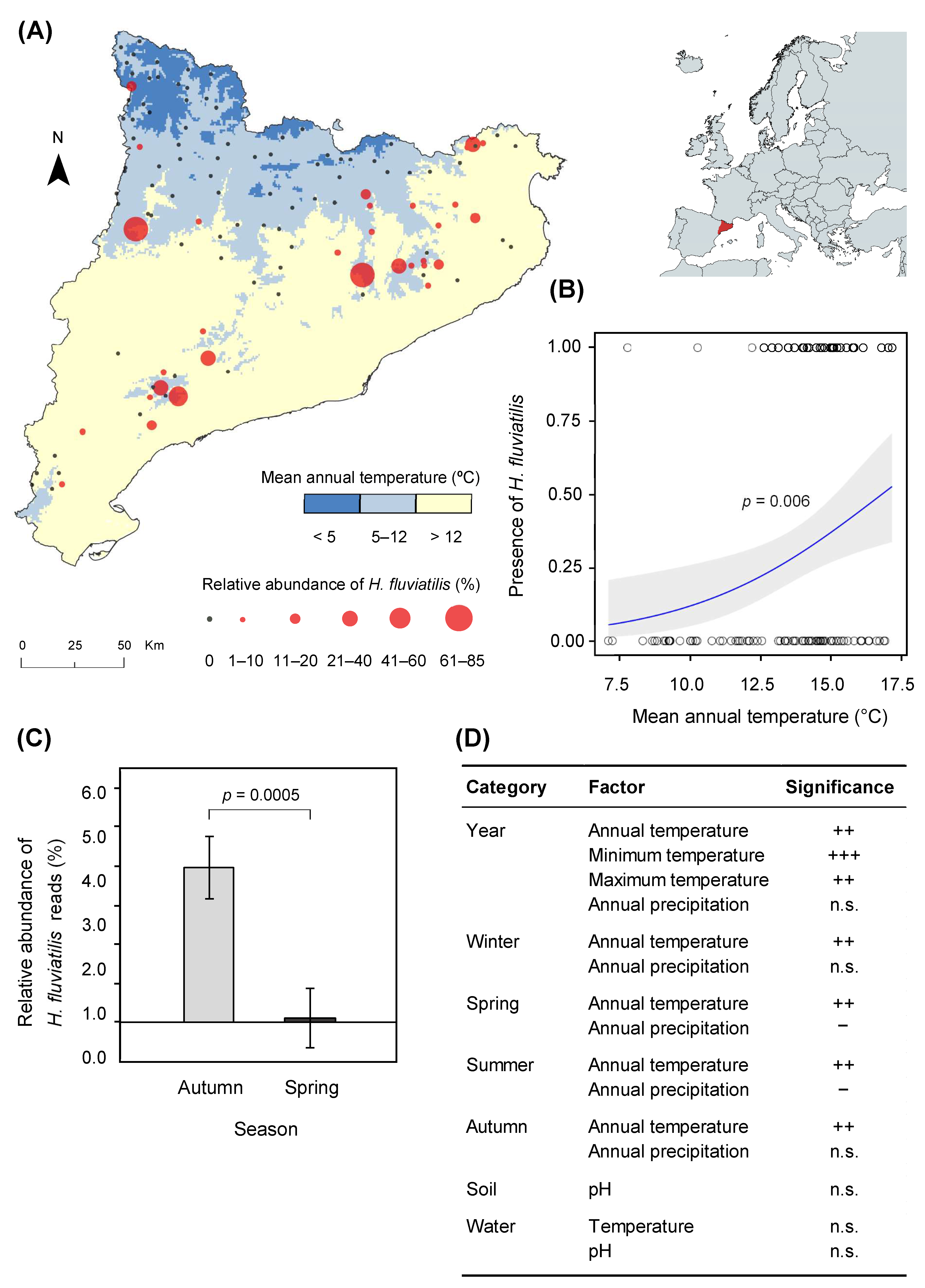{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Streams Survey
2.2. Library Preparation for Metabarcoding
2.3. Quality Control and Bioinformatic Analysis of Metabarcoding Data
2.4. Isolation From Leaves
2.5. Molecular Identification of Isolates
2.6. Pathogenicity Tests
2.7. Climatic and Soil Data
2.8. Statistical Analysis
3. Results
3.1. Halophytophthora fluviatilis Distribution
3.2. Pathogenicity Assessment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Nakagiri, A. Ecology and biodiversity of Halophytophthora species. Fungal Divers. 2000, 5, 153–164. [Google Scholar]
- Leaño, E.M.; Jones, E.B.G.; Vrijmoed, L.L.P. Why are Halophytophthora species well adapted to mangrove habitats? Fungal Divers. 2000, 5, 131–151. [Google Scholar]
- Su, C.-J.; Hsieh, S.-Y.; Chiang, M.W.-L.; Pang, K.-L. Salinity, pH and temperature growth ranges of Halophytophthora isolates suggest their physiological adaptations to mangrove environments. Mycology 2020, 11, 256–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeser, P.W.; Sutton, W.; Hansen, E.M.; Remigi, P.; Adams, G.C. Phytophthora species in forest streams in Oregon and Alaska. Mycologia 2011, 103, 22–35. [Google Scholar] [CrossRef]
- Ďatková, H. Phytophthora Diversity in Forest Streams of Moravia and Slovakia. Master’s Thesis, Mendel University in Brno, Brno, Czech Republic, 2020. [Google Scholar]
- Yang, X.; Hong, C. Halophytophthora fluviatilis sp. nov. from freshwater in Virginia. FEMS Microbiol. Lett. 2014, 352, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Nakagiri, A.; Ito, T.; Manoch, L.; Tanticharoen, M. A new Halophytophthora species, H. porriogvesica, from subtropical and tropical mangroves. Mycoscience 2001, 42, 33–41. [Google Scholar] [CrossRef]
- Jesus, A.L.; Marano, A.V.; Jerônimo, G.H.; de Souza, J.I.; Leaño, E.M.; Pires-Zottarelli, C.L.A. The genus Halophytophthora (Peronosporales, Straminipila) in Brazil: First descriptions of species. Braz. J. Bot. 2016, 39, 729–739. [Google Scholar] [CrossRef]
- Jesus, A.L.; Marano, A.V.; Gonçalves, D.R.; Jerônimo, G.H.; Pires-Zottarelli, C.L.A. Two new species of Halophytophthora from Brazil. Mycol. Prog. 2019, 18, 1411–1421. [Google Scholar] [CrossRef]
- Català, S. Development of DNA massive sequencing techniques and Real-Time PCR for the Detection, Identification and Quantitation of Phytophthora spp. in Environmental Samples. Ph.D. Thesis, Universitat Politècnica de València, Valencia, Spain, 2017. [Google Scholar]
- Nigrelli, L.; Thines, M. Tropical oomycetes in the German Bight—Climate warming or overlooked diversity? Fungal Ecol. 2013, 6, 152–160. [Google Scholar] [CrossRef]
- Man in ’t Veld, W.A.; Rosendahl, K.C.H.M.; van Rijswick, P.C.J.; Meffert, J.P.; Boer, E.; Westenberg, M.; van der Heide, T.; Govers, L.L. Multiple Halophytophthora spp. and Phytophthora spp. including P. gemini, P. inundata and P. chesapeakensis sp. nov. isolated from the seagrass Zostera marina in the Northern hemisphere. Eur. J. Plant Pathol. 2019, 153, 341–357. [Google Scholar] [CrossRef]
- Herrero, M.-L.; Sundheim, L.; Brevik, A.; Tojo, M.; Brurberg, M.B. Oomycetes survey in northern Norway. In Phytophthora in Forests & Natural Ecosystems, Proceedings of the 7th Meeting of the International Union of Forest Research Organization (IUFRO) Working Party S07.02.09, Esquel, Argentina, 9–14 November 2014; Sutton, W., Reeser, P.W., Hansen, E.M., Eds.; Available online: http://forestphytophthoras.org/sites/default/files/proceedings/IUFRO_Proceedings_2014.pdf (accessed on 3 December 2020).
- Newell, S.Y.; Miller, J.D.; Fell, J.W. Rapid and pervasive occupation of fallen mangrove leaves by a marine zoosporic fungus. Appl. Environ. Microbiol. 1987, 53, 2464–2469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marano, A.V.; Jesus, A.L.; de Souza, J.I.; Jerônimo, G.H.; Gonçalves, D.R.; Boro, M.C.; Rocha, S.C.O.; Pires-Zottarelli, C.L.A. Ecological roles of saprotrophic Peronosporales (Oomycetes, Straminipila) in natural environments. Fungal Ecol. 2016, 19, 77–88. [Google Scholar] [CrossRef]
- Govers, L.L.; Man in ’t Veld, W.A.; Meffert, J.P.; Bouma, T.J.; van Rijswick, P.C.J.; Heusinkveld, J.H.T.; Orth, R.J.; van Katwijk, M.M.; van der Heide, T. Marine Phytophthora species can hamper conservation and restoration of vegetated coastal ecosystems. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160812. [Google Scholar] [CrossRef] [PubMed]
- Redondo, M.A.; Boberg, J.; Stenlid, J.; Oliva, J. Contrasting distribution patterns between aquatic and terrestrial Phytophthora species along a climatic gradient are linked to functional traits. ISME J. 2018, 12, 2967–2980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drenth, A.; Wagels, G.; Smith, B.; Sendall, B.; O’Dwyer, C.; Irvine, G.; Irwin, J.A.G. Development of a DNA-based method for detection and identification of Phytophthora species. Australas. Plant Pathol. 2006, 35, 147–159. [Google Scholar] [CrossRef]
- Jeffers, S.N.; Martin, S.B. Comparison of two media selective for Phytophthora and Pythium species. Plant Dis. 1986, 70, 1038–1043. [Google Scholar] [CrossRef]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; APS Press: St. Paul, MN, USA, 1996. [Google Scholar]
- Wang, H.; Qi, M.; Cutler, A.J. A simple method of preparing plant samples for PCR. Nucleic Acids Res. 1993, 21, 4153–4154. [Google Scholar] [CrossRef] [Green Version]
- Samils, B.; Ihrmark, K.; Kaitera, J.; Stenlid, J.; Barklund, P. New genetic markers for identifying Cronartium flaccidum and Peridermium pini and examining genetic variation within and between lesions of Scots pine blister rust in Sweden. Fungal Biol. 2011, 115, 1303–1311. [Google Scholar] [CrossRef]
- De Cáceres, M.; Martin-StPaul, N.; Turco, M.; Cabon, A.; Granda, V. Estimating daily meteorological data and downscaling climate models over landscapes. Environ. Model. Softw. 2018, 108, 186–196. [Google Scholar] [CrossRef]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Redondo, M.A.; Boberg, J.; Olsson, C.H.B.; Oliva, J. Winter conditions correlate with Phytophthora alni subspecies distribution in southern Sweden. Phytopathology 2015, 105, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Oliva, J.; Redondo, M.Á.; Stenlid, J. Functional ecology of forest disease. Annu. Rev. Phytopathol. 2020, 58, 343–361. [Google Scholar] [CrossRef] [PubMed]
- Redondo, M.A.; Boberg, J.; Stenlid, J.; Oliva, J. Functional traits associated with the establishment of introduced Phytophthora spp. in Swedish forests. J. Appl. Ecol. 2018, 55, 1538–1552. [Google Scholar] [CrossRef]
- Crone, M.; McComb, J.A.; O’Brien, P.A.; Hardy, G.E.S.J. Survival of Phytophthora cinnamomi as oospores, stromata, and thick-walled chlamydospores in roots of symptomatic and asymptomatic annual and herbaceous perennial plant species. Fungal Biol. 2013, 117, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Colquhoun, I.J.; Hardy, G.E.S.J. New insights into the survival strategy of the invasive soilborne pathogen Phytophthora cinnamomi in different natural ecosystems in Western Australia. For. Pathol. 2013, 43, 266–288. [Google Scholar] [CrossRef]
- Caballol, M.; Štraus, D.; Macia, H.; Ramis, X.; Redondo, M.Á.; Oliva, J. Halophytophthora fluviatilis pathogenicity and distribution along a Mediterranean-subalpine gradient. Figshare 2021. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caballol, M.; Štraus, D.; Macia, H.; Ramis, X.; Redondo, M.Á.; Oliva, J. Halophytophthora fluviatilis Pathogenicity and Distribution along a Mediterranean-Subalpine Gradient. J. Fungi 2021, 7, 112. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7020112
Caballol M, Štraus D, Macia H, Ramis X, Redondo MÁ, Oliva J. Halophytophthora fluviatilis Pathogenicity and Distribution along a Mediterranean-Subalpine Gradient. Journal of Fungi. 2021; 7(2):112. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7020112
Chicago/Turabian StyleCaballol, Maria, Dora Štraus, Héctor Macia, Xavier Ramis, Miguel Á. Redondo, and Jonàs Oliva. 2021. "Halophytophthora fluviatilis Pathogenicity and Distribution along a Mediterranean-Subalpine Gradient" Journal of Fungi 7, no. 2: 112. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7020112