Proteomics Reveals Octyl Gallate as an Environmentally Friendly Wood Preservative Leading to Reactive Oxygen Species-Driven Metabolic Inflexibility and Growth Inhibition in White-Rot Fungi (Lenzites betulina and Trametes versicolor)

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Fungal Strain
2.2. Antifungal Assays
2.3. 1,1-Diphenyl-2-Picrylhydrazyl Assay
2.4. Ferrous Ion-Chelating Ability Assay
2.5. Determination of Laccase and Manganese Peroxidase Activities
2.6. Protein Sample Preparation
2.7. SDS-PAGE
2.8. Nanoflow Ultra-High-Performance Liquid Chromatography−Tandem Mass Spectrometry
2.9. Mass Spectrometric Data Analysis
2.10. Statistical Analysis
3. Results and Discussion
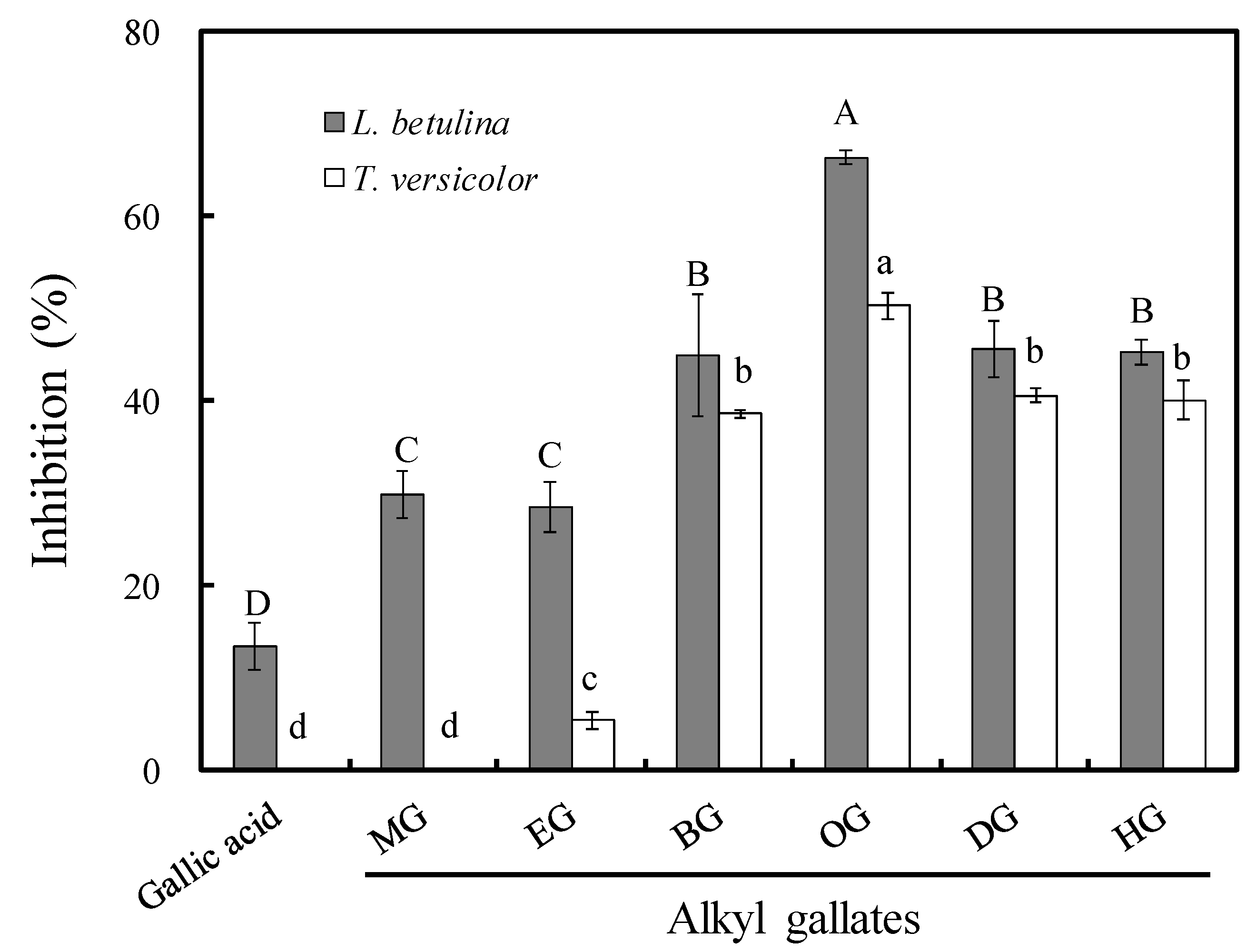
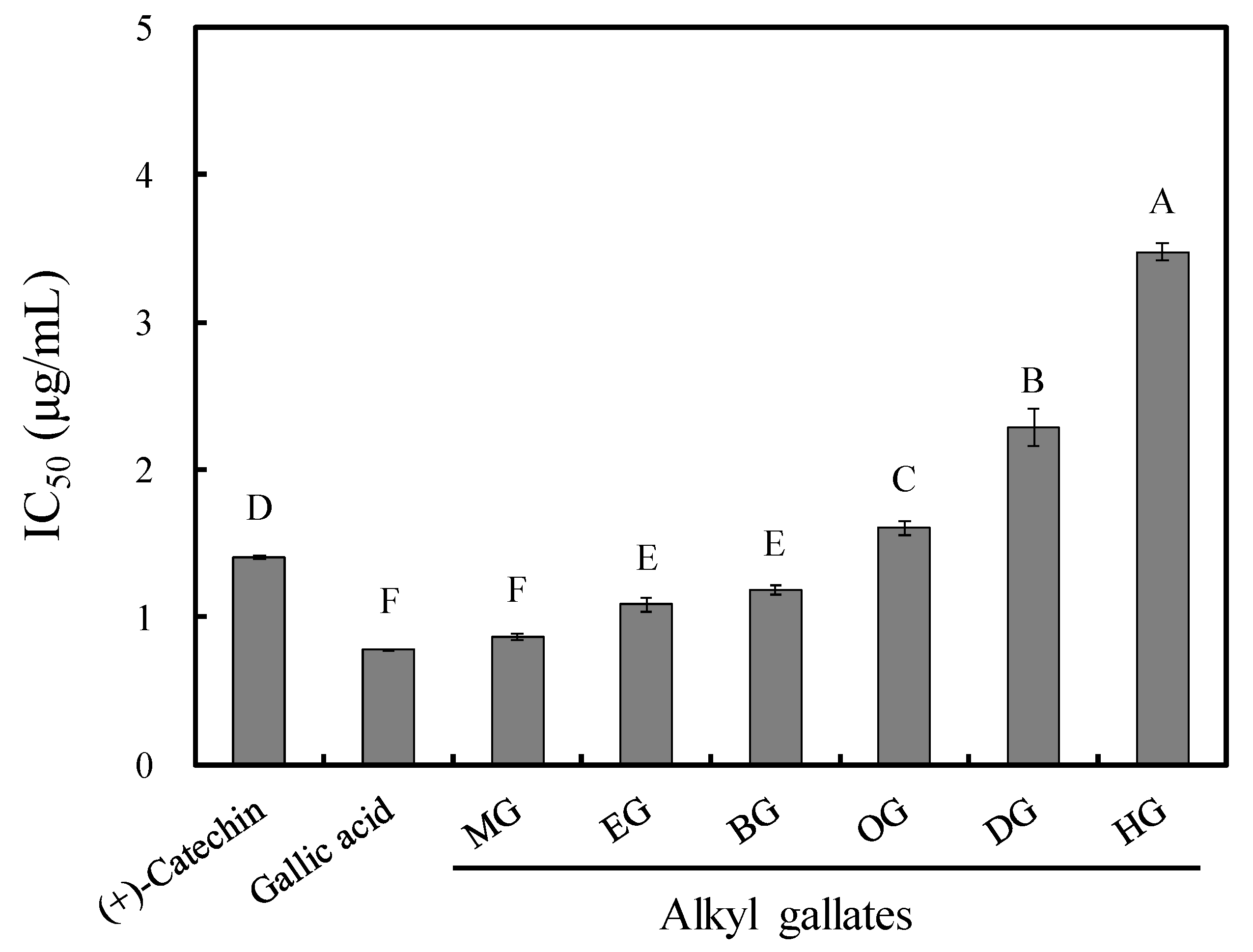
3.1. Antifungal Activity of Long-Chain Alkyl Gallates
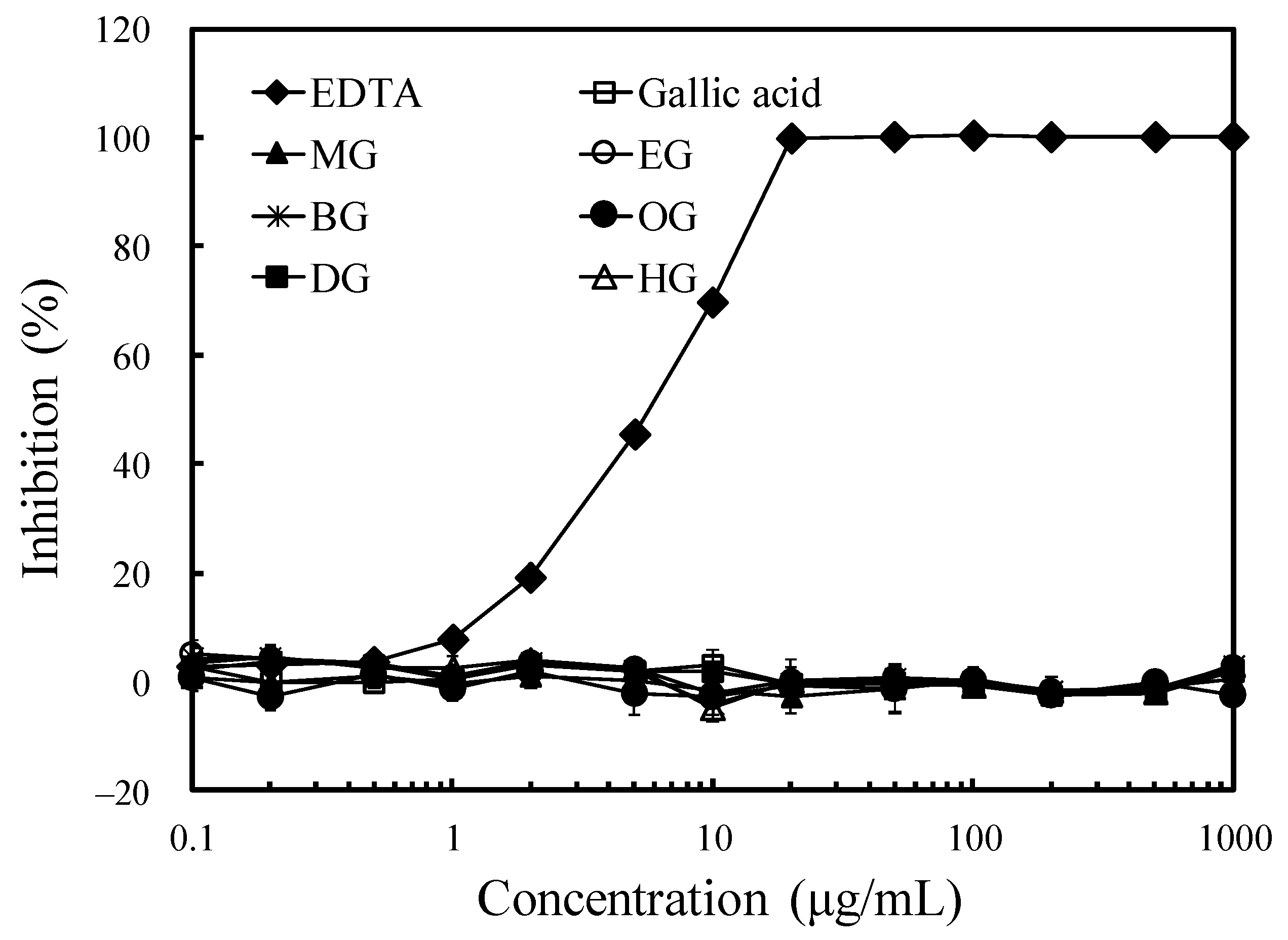
3.2. Antioxidant Activity and Ferrous Ion-Chelating Effect of Long-Chain Alkyl Gallates
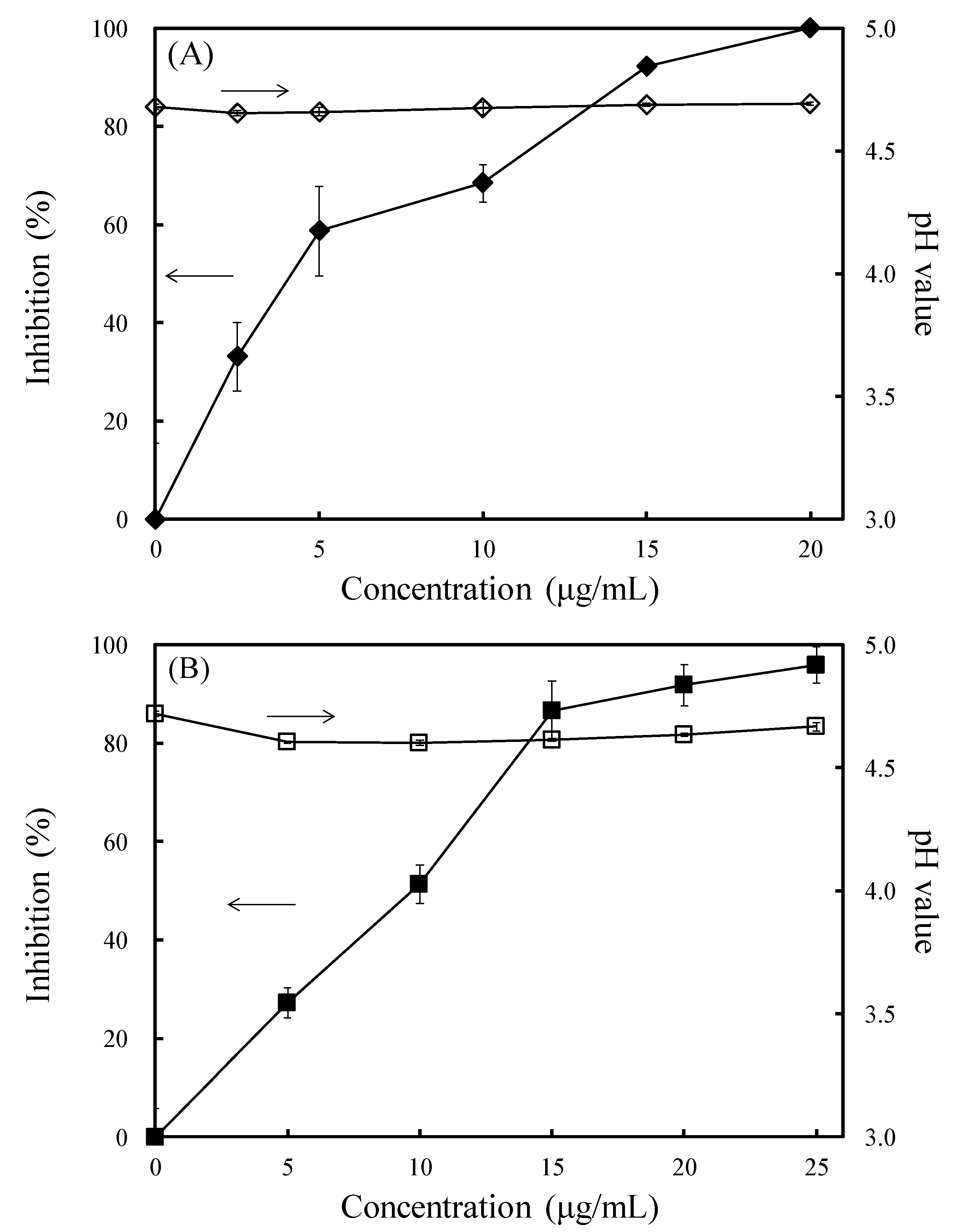
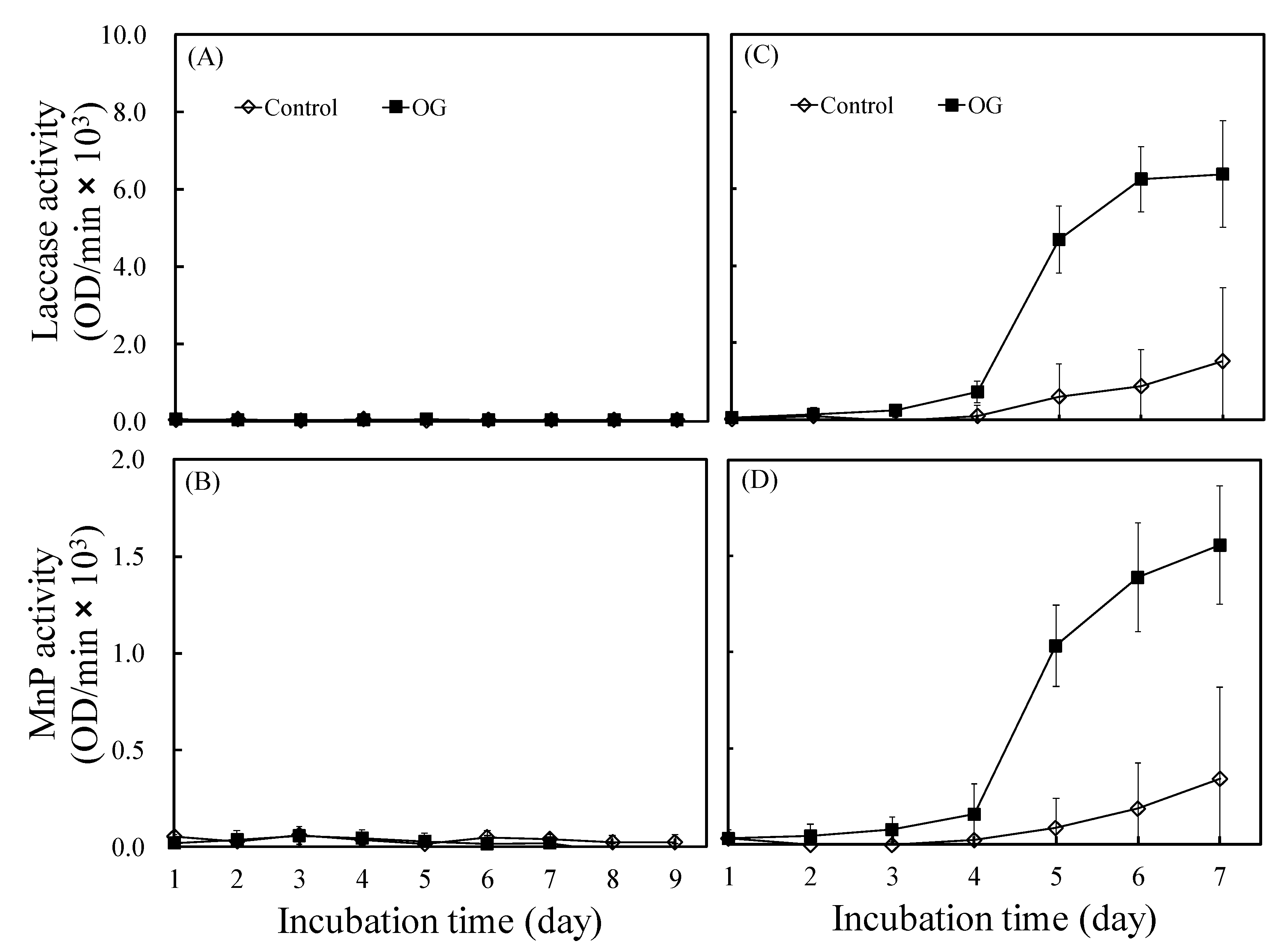
3.3. Effects of OG on the Laccase and Manganese Peroxidase Activities of L. betulina and T. versicolor
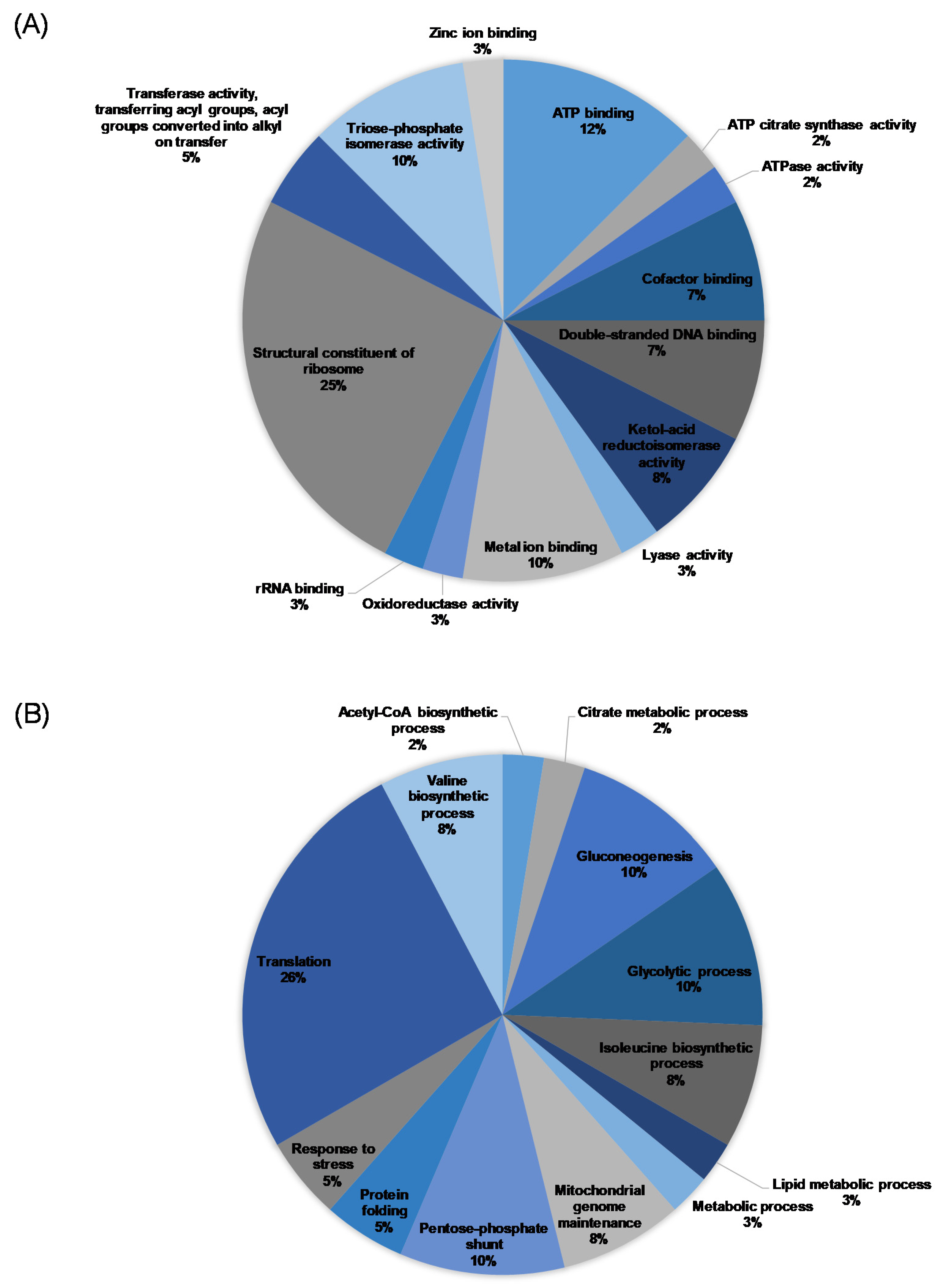
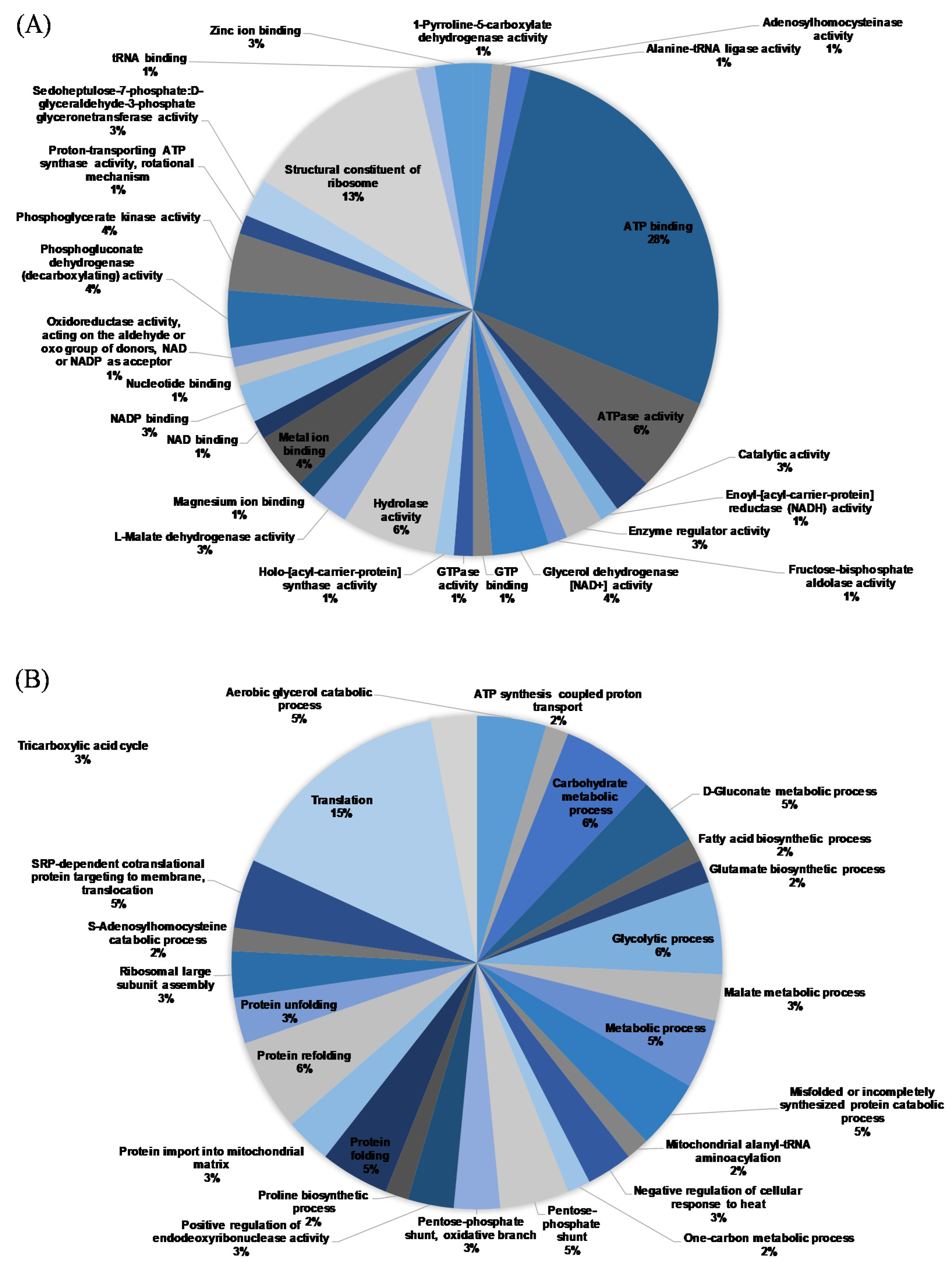
3.4. Proteomics Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hassan, E.B.; El-Giar, E.M.; Steele, P. Evaluation of the antioxidant activities of different bio-oils and their phenolic distilled fractions for wood preservation. Int. Biodeterior. Biodegrad. 2016, 110, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Goodell, B.; Nicholas, D.D.; Schultz, T.P. Introduction to wood deterioration and preservation. In Wood Deterioration and Preservation: Advances in Our Changing World; Goodell, B., Nicholas, D.D., Schultz, T.P., Eds.; American Chemical Society: Washington, DC, USA, 2003; pp. 2–9. [Google Scholar]
- Barbero-López, A.; Monzó-Beltrán, J.; Virjamo, V.; Akkanen, J.; Haapala, A. Revalorization of coffee silverskin as a potential feedstock for antifungal chemicals in wood preservation. Int. Biodeterior. Biodegrad. 2020, 152, 105011. [Google Scholar] [CrossRef]
- Lourençon, T.V.; Mattos, B.D.; Cademartori, P.H.; Magalhães, W.L. Bio-oil from a fast pyrolysis pilot plant as antifungal and hydrophobic agent for wood preservation. J. Anal. Appl. Pyrolysis 2016, 122, 1–6. [Google Scholar] [CrossRef]
- Yalcin, M.; Akcay, C.; Sahin, H.I. The effect of particle board industry waste tar on the physical and biological durability of wood. Bioresources 2020, 15, 1810–1827. [Google Scholar] [CrossRef]
- Valette, N.; Perrot, T.; Sormani, R.; Gelhaye, E.; Morel-Rouhier, M. Antifungal activities of wood extractives. Fungal Biol. Rev. 2017, 31, 113–123. [Google Scholar] [CrossRef]
- Saibabu, V.; Fatima, Z.; Ahmad, K.; Khan, L.A.; Hameed, S. Octyl gallate triggers dysfunctional mitochondria leading to ROS driven membrane damage and metabolic inflexibility along with attenuated virulence in Candida albicans. Med. Mycol. 2019, 58, 380–392. [Google Scholar] [CrossRef]
- Hsu, F.-L.; Chen, P.-S.; Chang, H.-T.; Chang, S.-T. Effects of alkyl chain length of gallates on their antifungal property and potency as an environmentally benign preservative against wood-decay fungi. Int. Biodeterior. Biodegrad. 2009, 63, 543–547. [Google Scholar] [CrossRef]
- Zacchino, S.A.; Butassi, E.; Di Liberto, M.; Raimondi, M.; Postigo, A.; Sortino, M. Plant phenolics and terpenoids as adjuvants of antibacterial and antifungal drugs. Phytomedicine 2017, 37, 27–48. [Google Scholar] [CrossRef]
- Badhani, B.; Sharma, N.; Kakkar, R. Gallic acid: A versatile antioxidant with promising therapeutic and industrial applications. RSC Adv. 2015, 5, 27540–27557. [Google Scholar] [CrossRef]
- Kubo, I.; Xiao, P.; Fujita, K. Antifungal activity of octyl gallate: Structural criteria and mode of action. Bioorg. Med. Chem. Lett. 2001, 11, 347–350. [Google Scholar] [CrossRef]
- Kubo, I.; Fujita, K.-I.; Nihei, K.-I. Anti-SalmonellaActivity of Alkyl Gallates†. J. Agric. Food Chem. 2002, 50, 6692–6696. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Lin, L.-C.; Ho, S.-T.; Tung, Y.-T.; Tseng, Y.-H.; Wu, J.-H. Antioxidant Activities and Phytochemicals of Leaf Extracts from 10 NativeRhododendronSpecies in Taiwan. Evid. Based Complement. Altern. Med. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tamagawa, Y.; Yamaki, R.; Hirai, H.; Kawai, S.; Nishida, T. Removal of estrogenic activity of natural steroidal hormone estrone by ligninolytic enzymes from white rot fungi. Chemosphere 2006, 65, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.-C.; Chiu, Y.-S.; Chiu, W.-C.; Tung, Y.-T.; Chuang, H.-L.; Wu, J.-H.; Huang, C.-C. Proteomics Analysis to Identify and Characterize the Molecular Signatures of Hepatic Steatosis in Ovariectomized Rats as a Model of Postmenopausal Status. Nutrients 2015, 7, 8752–8766. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.-C.; Chen, Y.-W.; Jeng, T.-L.; Li, C.-R.; Kuo, C.-F. Consumption of Purple Sweet Potato Affects Post-Translational Modification of Plasma Proteins in Hamsters. J. Agric. Food Chem. 2013, 61, 12450–12458. [Google Scholar] [CrossRef]
- Uen, Y.H.; Lin, K.Y.; Sun, D.P.; Liao, C.C.; Hsieh, M.S.; Huang, Y.K.; Chen, Y.W.; Huang, P.H.; Chen, W.J.; Tai, C.C.; et al. Comparative proteomics, network analysis and post-translational modification identification reveal differential profiles of plasma Con A-bound glycoprotein biomarkers in gastric cancer. J. Proteomics 2013, 83, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Andjelkovic, M.; Vancamp, J.; Demeulenaer, B.; Depaemelaere, G.; Socaciu, C.; Verloo, M.; Verhe, R. Iron-chelation properties of phenolic acids bearing catechol and galloyl groups. Food Chem. 2006, 98, 23–31. [Google Scholar] [CrossRef]
- Sergentani, A.G.; Gonou-Zagou, Z.; Kapsanaki-Gotsi, E.; Hatzinikolaou, D.G. Lignocellulose degradation potential of Basidiomycota from Thrace (NE Greece). Int. Biodeterior. Biodegrad. 2016, 114, 268–277. [Google Scholar] [CrossRef]
- Alberts, J.; Gelderblom, W.; Botha, A.; Van Zyl, W. Degradation of aflatoxin B1 by fungal laccase enzymes. Int. J. Food Microbiol. 2009, 135, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Tortella, G.R.; Rubilar, O.; Gianfreda, L.; Valenzuela, E.; Diez, M.C. Enzymatic characterization of Chilean native wood-rotting fungi for potential use in the bioremediation of polluted environments with chlorophenols. World J. Microbiol. Biotechnol. 2008, 24, 2805–2818. [Google Scholar] [CrossRef] [Green Version]
- Piscitelli, A.; Giardina, P.; Lettera, V.; Pezzella, C.; Sannia, G.; Faraco, V. Induction and Transcriptional Regulation of Laccases in Fungi. Curr. Genom. 2011, 12, 104–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaszek, M.; Grzywnowicz, K.; Malarczyk, E.; Leonowicz, A. Enhanced extracellular laccase activity as a part of the response system of white rot fungi: Trametes versicolor and Abortiporus biennis to paraquat-caused oxidative stress conditions. Pestic. Biochem. Physiol. 2006, 85, 147–154. [Google Scholar] [CrossRef]
- Sant, D.; Tupe, S.; Ramana, C.; Deshpande, M. Fungal cell membrane-promising drug target for antifungal therapy. J. Appl. Microbiol. 2016, 121, 1498–1510. [Google Scholar] [CrossRef] [Green Version]
- Takai, E.; Hirano, A.; Shiraki, K. Effects of alkyl chain length of gallate on self-association and membrane binding. J. Biochem. 2011, 150, 165–171. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, Y.; Yan, L.; Liang, R.-M.; Dai, B.-D.; Tang, R.-J.; Gao, P.-H.; Jiang, Y.-Y. Proteomic Analysis Reveals a Synergistic Mechanism of Fluconazole and Berberine against Fluconazole-ResistantCandida albicans: Endogenous ROS Augmentation. J. Proteome Res. 2009, 8, 5296–5304. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Grummt, I. Cellular Stress and Nucleolar Function. Cell Cycle 2005, 4, 1036–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, R.; Chandler-Brown, D.; Ramos, F.J.; Shamieh, L.S.; Kaeberlein, M. Regulation of mRNA Translation as a Conserved Mechanism of Longevity Control. Adv. Exp. Med. Biol. 2010, 694, 14–29. [Google Scholar] [CrossRef] [PubMed]
- Steffen, K.K.; MacKay, V.L.; Kerr, E.O.; Tsuchiya, M.; Hu, D.; Fox, L.A.; Dang, N.; Johnston, E.D.; Oakes, J.A.; Tchao, B.N.; et al. Yeast Life Span Extension by Depletion of 60S Ribosomal Subunits Is Mediated by Gcn4. Cell 2008, 133, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ren, A.; Li, M.-J.; Cao, P.-F.; Chen, T.-X.; Zhang, G.; Shi, L.; Jiang, A.-L.; Zhao, M.-W. Heat Stress Modulates Mycelium Growth, Heat Shock Protein Expression, Ganoderic Acid Biosynthesis, and Hyphal Branching of Ganoderma lucidum via Cytosolic Ca2+. Appl. Environ. Microbiol. 2016, 82, 4112–4125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamoth, F.; Juvvadi, P.R.; Fortwendel, J.R.; Steinbach, W.J. Heat Shock Protein 90 Is Required for Conidiation and Cell Wall Integrity in Aspergillus fumigatus. Eukaryot. Cell 2012, 11, 1324–1332. [Google Scholar] [CrossRef] [Green Version]
- Bedin, M.; Gaben, A.-M.; Mester, J. Geldanamycin, an inhibitor of the chaperone activity of HSP90, induces MAPK-independent cell cycle arrest. Int. J. Cancer 2004, 109, 643–652. [Google Scholar] [CrossRef]
- Grant, C.M. Metabolic reconfiguration is a regulated response to oxidative stress. J. Biol. 2008, 7, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grüning, N.M.; Rinnerthaler, M.; Bluemlein, K.; Mülleder, M.; Wamelink, M.M.C.; Lehrach, H.; Jakobs, C.; Breitenbach, M.; Ralser, M. Pyruvate kinase triggers a metabolic feedback loop that controls redox metabolism in respiring cells. Cell Metab. 2011, 14, 415–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perl, A.; Hanczko, R.; Telarico, T.; Oaks, Z.; Landas, S. Oxidative stress, inflammation and carcinogenesis are controlled through the pentose phosphate pathway by transaldolase. Trends Mol. Med. 2011, 17, 395–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotenberg, M.; Moritz, M.; Woolford, J.L. Depletion of Saccharomyces cerevisiae ribosomal protein L16 causes a decrease in 60S ribosomal subunits and formation of half-mer polyribosomes. Genes Dev. 1988, 2, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Jolly, C. Role of the Heat Shock Response and Molecular Chaperones in Oncogenesis and Cell Death. J. Natl. Cancer Inst. 2000, 92, 1564–1572. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Accession No. | pI | MW | GO | Expression Ratio |
|---|---|---|---|---|---|
| 60S ribosomal protein L9-B (Schizosaccharomyces pombe 972h-) | A0A0K6FS64_9HOMO | 9.38 | 21,383.81 | - | 3.92 |
| Ketol-acid reductoisomerase, mitochondrial | W4KC19_9HOMO | 8.85 | 44,553.29 | - | 2.17 |
| Ketol-acid reductoisomerase, mitochondrial | A0A0C3DUH5_9HOMO | 8.91 | 44,841.78 | - | 2.04 |
| Ketol-acid reductoisomerase, mitochondrial | A0A060SWV9_PYCCI | 8.98 | 44,706.68 | - | 1.98 |
| Triosephosphate isomerase | R4I570_9AGAR | 5.70 | 26,921.71 | - | 3.89 |
| Triosephosphate isomerase | A0A0C3PQJ3_PHLGI | 5.50 | 26,627.37 | - | 3.46 |
| Triosephosphate isomerase | A0A151V9C9_HYPMA | 5.97 | 26,765.62 | - | 3.46 |
| Triosephosphate isomerase | S8FUL8_FOMPI | 6.00 | 26,850.73 | - | 3.46 |
| Uncharacterized protein | A0A0C3S325_PHLGI | 11.17 | 20,963.75 | 0006412 | 3.84 |
| Protein Name | Accession No. | pI | MW | GO | Expression Ratio |
|---|---|---|---|---|---|
| 60S ribosomal protein L12 | A0A165F4X5_9APHY | 9.19 | 17,522.26 | 0006412 | 0.09 |
| 60S ribosomal protein L12 | A0A165RLF8_9APHY | 9.06 | 17,510.30 | 0006412 | 0.09 |
| 60S ribosomal protein L12 | A0A1C7LQZ3_GRIFR | 9.35 | 17,364.10 | - | 0.09 |
| 60S ribosomal protein L12 | B8P7S1_POSPM | 9.19 | 17,550.31 | 0006412 | 0.09 |
| ATP citrate lyase isoform 2 | A0A0D7B0G9_9HOMO | 6.55 | 125,756.11 | - | 0.11 |
| ATP citrate synthase | A0A165GIY9_9APHY | 8.33 | 125,805.87 | - | Control only |
| GroES-like protein | A0A166C2Q9_9HOMO | 6.37 | 37,305.72 | - | 0.25 |
| Heat shock protein 90 | R7S7P6_TRAVS | 4.95 | 79,624.25 | 0006457 0006950 | 0.39 |
| Heat shock protein HSP 90-alpha | A0A0B7FTU3_THACB | 4.94 | 79,834.83 | - | 0.36 |
| Mitochondrial glycoprotein | R7SPA0_DICSQ | 4.67 | 29,577.92 | - | 0.25 |
| Uncharacterized protein | J4HUC5_9APHY | 9.59 | 37,362.90 | 0006412 | 0.09 |
| Uncharacterized protein | M2R3S2_CERS8 | 9.06 | 17,655.46 | 0006412 | 0.09 |
| Uncharacterized protein | A0A060S428_PYCCI | 7.63 | 107,357.17 | - | 0.21 |
| Uncharacterized protein | A0A0C3P9N6_PHLGI | 9.32 | 17,499.22 | 0006412 | 0.09 |
| Uncharacterized protein | K5UVC6_PHACS | 9.19 | 17,396.05 | 0006412 | 0.09 |
| Uncharacterized protein | S8FGW3_FOMP | 9.06 | 17,496.28 | 0006412 | 0.09 |
| Protein Name | Accession No. | pI | MW | GO | Expression Ratio |
|---|---|---|---|---|---|
| 40S ribosomal protein | A0A146HFY9_9AGAR | 10.20 | 17,473.48 | 0006412 | 4.99 |
| 40S ribosomal protein S15 | A0A0B7FZY5_THACB | 10.24 | 17,610.61 | 0006412 | 3.78 |
| 40S ribosomal protein S15 | A0A151VTC7_HYPMA | 10.31 | 17,514.54 | 0006412 | 3.78 |
| 40S ribosomal protein S15 | L8X3B3_THACA | 9.90 | 16,454.27 | 0006412 | 3.78 |
| 6-phosphogluconate dehydrogenase, decarboxylating | A0A0C2XM62_AMAMU | 6.35 | 53,162.64 | - | 2.43 |
| 6-phosphogluconate dehydrogenase, decarboxylating | A0A0D7B7I4_9HOMO | 5.79 | 53,373.75 | 0019521 0009051 | 3.72 |
| 6-phosphogluconate dehydrogenase, decarboxylating | A0A146HJH8_9AGAR | 7.09 | 53,411.28 | - | 2.37 |
| ATP synthase subunit alpha | K5VTN2_PHACS | 9.15 | 58,843.66 | 0015986 | 1.99 |
| Chaperonin GroL | A0A165LVH4_9APHY | 5.79 | 62,855.91 | - | 2.14 |
| Heat shock protein | A8PB53_COPC7 | 5.62 | 63,344.06 | - | 2.20 |
| Uncharacterized protein | A0A060S4R5_PYCCI | 9.04 | 19,588.59 | 0006412 | 2.84 |
| Uncharacterized protein | A0A060SGS2_PYCCI | 9.74 | 13,299.48 | 0006412 | 7.99 |
| Uncharacterized protein | R7SX26_DICSQ | 10.51 | 17,474.38 | - | 4.66 |
| Protein Name | Accession No. | pI | MW | GO | Expression Ratio |
|---|---|---|---|---|---|
| 60S ribosomal protein L3 | B8PIG2_POSPM | 10.26 | 4,3761.83 | - | 0.45 |
| 60S ribosomal protein L4/L1/L2 | A0A165ZAV7_9HOMO | 11.12 | 40,412.82 | 0006412 | 0.23 |
| AAA ATPase | A0A165B9Y7_9APHY | 4.99 | 89,912.01 | - | 0.46 |
| Actin 1 | R7SI12_DICSQ | 5.44 | 41,692.58 | - | 0.37 |
| Actin-1 | A0A1C7LPB9_GRIFR | 5.35 | 42,816.81 | - | 0.37 |
| Adenosylhomocysteinase | M2QCQ6_CERS8 | 5.74 | 47,267.26 | - | 0.42 |
| Alanine--tRNA ligase | A0A060SL88_PYCCI | 5.75 | 106,172.69 | - | 0.46 |
| Beta-actin | V5W5W4_GANLU | 5.44 | 41,520.41 | - | 0.37 |
| Beta-actin-like protein | M2QI81_CERS8 | 5.44 | 41,692.58 | - | 0.37 |
| Delta-1-pyrroline-5-carboxylate dehydrogenase 1 | A0A165ZHP2_9HOMO | 6.48 | 59,553.60 | - | 0.32 |
| Fructose 1,6-bisphosphate aldolase | A0A1B7N8B6_9HOMO | 6.02 | 38,850.08 | - | Control only |
| Glycerol dehydrogenase | R7S6H2_TRAVS | 5.30 | 42,121.11 | - | 0.46 |
| Glycerol dehydrogenase | R7SKI4_DICSQ | 5.58 | 40,618.43 | - | 0.52 |
| Heat shock cognate 70 | A0A0H2RYR8_9HOMO | 5.13 | 70,997.65 | 1900035 | 0.45 |
| Heat shock protein 70 | R7SN52_DICSQ | 5.64 | 71,837.59 | - | 0.48 |
| Malate dehydrogenase | A0A165P5J0_9HOMO | 6.45 | 33,510.44 | 0005975 0006108 0006099 | 0.31 |
| Malate dehydrogenase | S7RXX1_GLOTA | 9.05 | 35,546.85 | 0005975 0006108 0006099 | 0.31 |
| Phosphoglycerate kinase | A0A0C2XI58_AMAMU | 5.97 | 44,767.53 | 0006096 | 0.16 |
| Phosphoglycerate kinase | A0A0C3S5Q2_PHLGI | 5.99 | 44,470.21 | 0006096 | 0.12 |
| Phosphoglycerate kinase | A0A146IDB4_9AGAR | 6.15 | 44,134.66 | - | Control only |
| Putative carboxymethylenebutenolidase | A0A137QS07_9AGAR | 6.75 | 28,102.12 | - | 0.33 |
| Related to HSP70 heat shock protein 70 (Hsp70) | G4T8Z4_SERID | 5.14 | 70,572.25 | 0006515 0006457 0006616 | 0.42 |
| Transaldolase | A0A060SMY4_PYCCI | 6.19 | 36,149.43 | 0005975 0006098 | 0.35 |
| Transaldolase | R7S6N3_TRAVS | 7.64 | 28,010.29 | 0005975 0006098 | 0.42 |
| Transketolase-like protein 2 | A0A1C7M6L6_GRIFR | 6.33 | 64,753.10 | - | 0.15 |
| Uncharacterized protein | A0A060S8Y5_PYCCI | 5.03 | 89,836.05 | - | 0.52 |
| Uncharacterized protein | J4G7W8_9APHY | 4.97 | 89,888.93 | - | 0.48 |
| Uncharacterized protein | J4GJQ5_9APHY | 5.56 | 72,198.74 | - | 0.51 |
| Uncharacterized protein | A0A060SCB9_PYCCI | 5.92 | 433,211.19 | - | 0.35 |
| Uncharacterized protein | A0A060SEM9_PYCC | 5.21 | 40,163.80 | - | 0.45 |
| Uncharacterized protein | A0A060SFD3_PYCCI | 5.13 | 70,262.90 | 0006515 0006457 0006616 | 0.44 |
| Uncharacterized protein | A0A060SNB2_PYCCI | 6.51 | 92,680.46 | - | 0.34 |
| Uncharacterized protein | A0A060SR94_PYCCI | 5.97 | 69,483.20 | 0008152 | 0.27 |
| Uncharacterized protein | A0A0C3NYK6_PHLGI | 5.37 | 41,591.47 | - | 0.17 |
| Uncharacterized protein | A0A0D2PLJ0_9AGAR | 6.31 | 30,884.26 | - | 0.34 |
| Uncharacterized protein | J4HZZ4_9APHY | 9.79 | 106,199.49 | - | 0.46 |
| Uncharacterized protein | S8ET35_FOMPI | 5.10 | 71,000.55 | 1900035 | 0.46 |
| Unplaced genomic scaffold CY34scaffold_2, whole genome shotgun sequence | A0A0D0AGV4_9HOMO | 5.12 | 70,384.87 | 0006515 0006457 0006616 | 0.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.-W.; Liao, C.-C.; Hung, K.-C.; Wang, Z.-Y.; Tung, Y.-T.; Wu, J.-H. Proteomics Reveals Octyl Gallate as an Environmentally Friendly Wood Preservative Leading to Reactive Oxygen Species-Driven Metabolic Inflexibility and Growth Inhibition in White-Rot Fungi (Lenzites betulina and Trametes versicolor). J. Fungi 2021, 7, 145. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7020145
Xu J-W, Liao C-C, Hung K-C, Wang Z-Y, Tung Y-T, Wu J-H. Proteomics Reveals Octyl Gallate as an Environmentally Friendly Wood Preservative Leading to Reactive Oxygen Species-Driven Metabolic Inflexibility and Growth Inhibition in White-Rot Fungi (Lenzites betulina and Trametes versicolor). Journal of Fungi. 2021; 7(2):145. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7020145
Chicago/Turabian StyleXu, Jin-Wei, Chen-Chung Liao, Ke-Chang Hung, Zhong-Yao Wang, Yu-Tang Tung, and Jyh-Horng Wu. 2021. "Proteomics Reveals Octyl Gallate as an Environmentally Friendly Wood Preservative Leading to Reactive Oxygen Species-Driven Metabolic Inflexibility and Growth Inhibition in White-Rot Fungi (Lenzites betulina and Trametes versicolor)" Journal of Fungi 7, no. 2: 145. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7020145