
Application of Bio-Friendly Formulations of Chitinase-Producing Streptomyces cellulosae Actino 48 for Controlling Peanut Soil-Borne Diseases Caused by Sclerotium rolfsii

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal and Actinobacterial Isolates
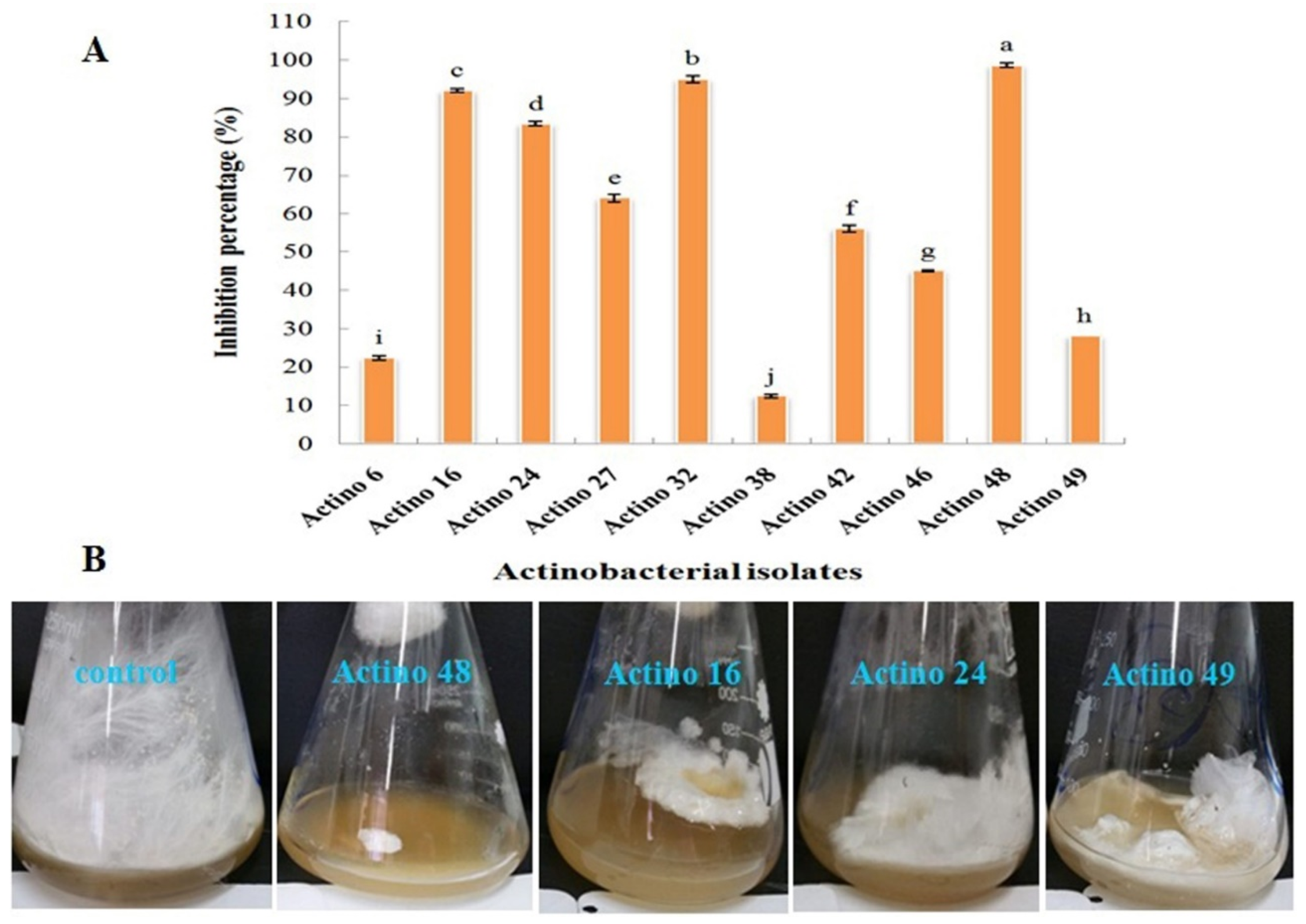
2.2. Antagonistic Effect of Actinobacterial Isolates against S. rolfsii
2.3. Qualitative and Quantitative Evaluation of Chitinase Production from S. cellulosae Actino 48
2.3.1. Detection of Chitinase Production
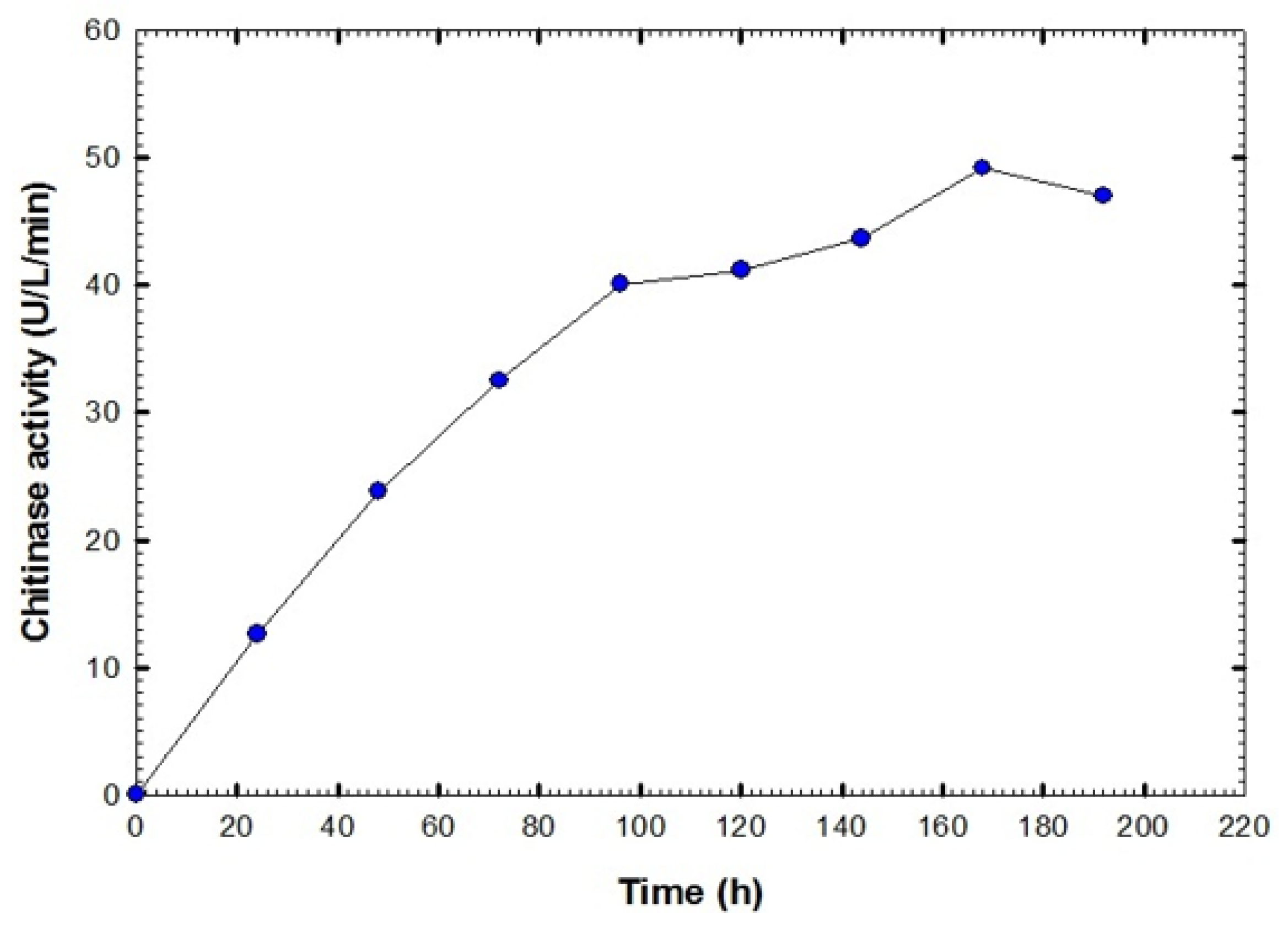
2.3.2. Chitinase Assay
2.4. Detection of Interaction between S. cellulosae Actino 48 and S. rolfsii
2.5. Formulation of Culture Broth, Cell-Free Supernatant, and Cell Pellet Suspension of Chitinase-Producing S. cellulosae Actino 48
2.6. Characterizations of Talc-Based Bioformulations of Chitinase-Producing S. cellulosae Actino 48
2.6.1. Scanning Electron Microscopy
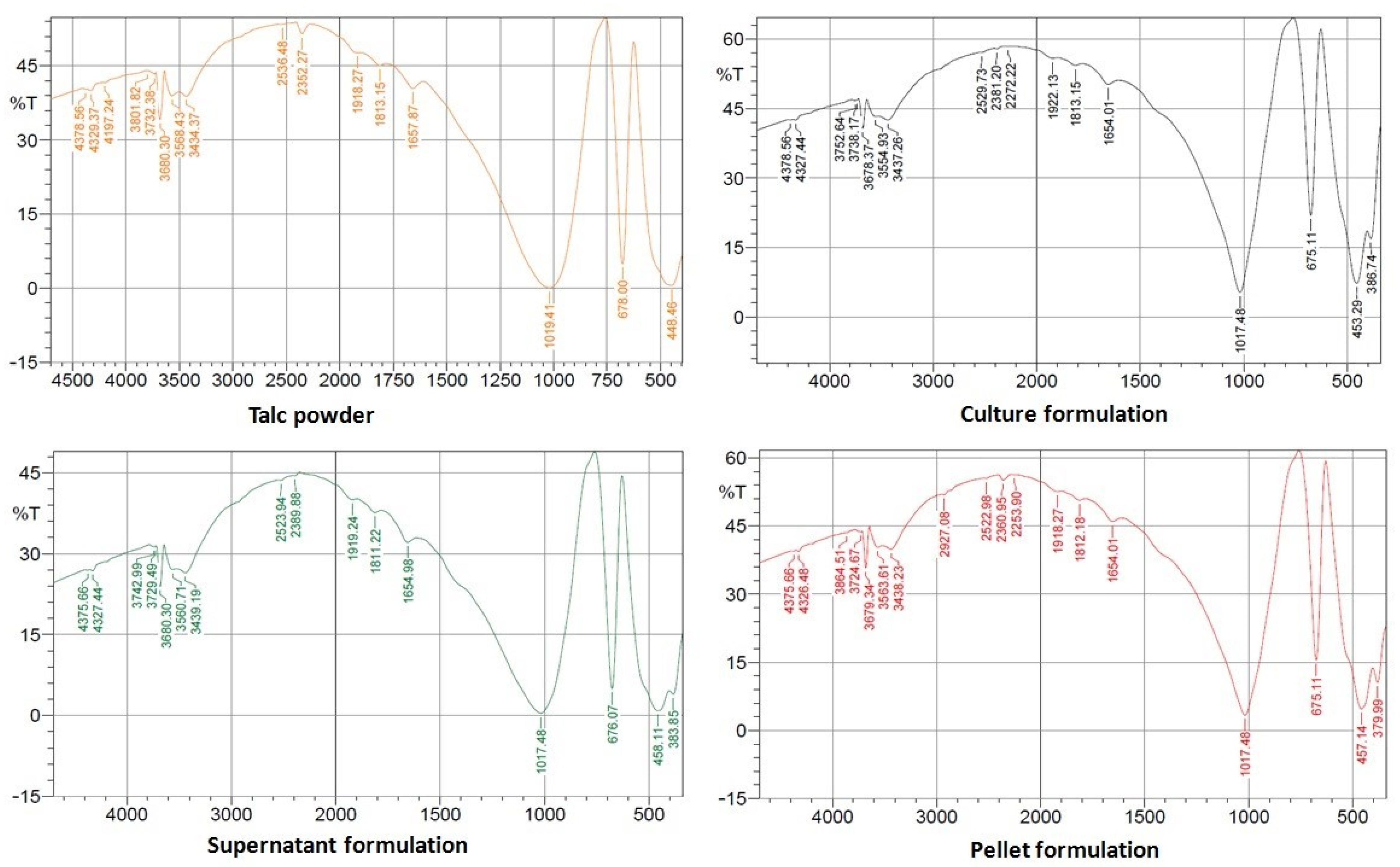
2.6.2. Fourier Transform Infrared (FTIR) Spectroscopy
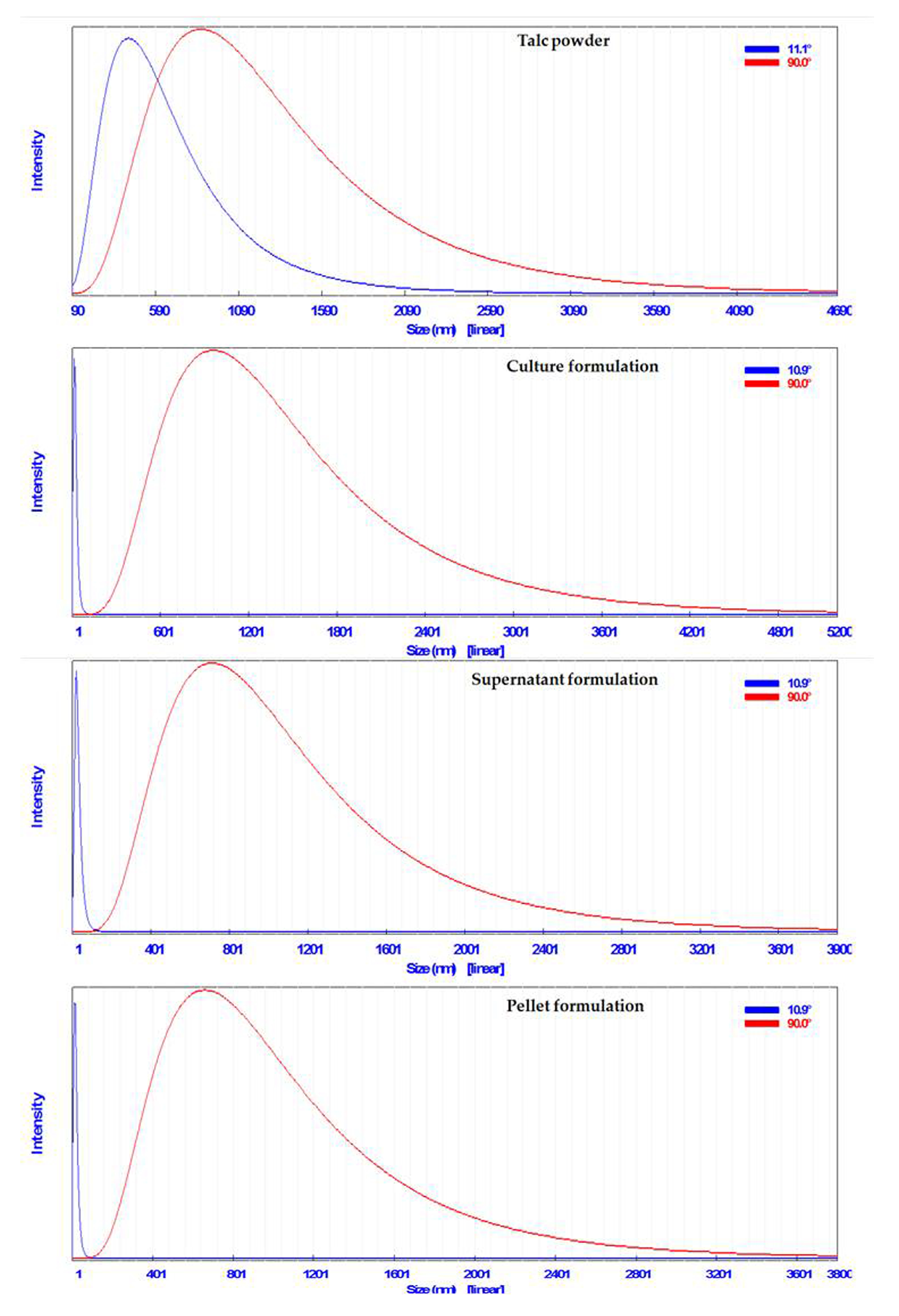
2.6.3. Particle Size Analysis
2.7. Application of Bio-Friendly Formulations of Chitinase-Producing S. cellulosae Actino 48 as Biocontrol Agents against S. rolfsii on Peanut
2.7.1. Preparation of Fungal Inoculum
2.7.2. Soil Infestation
2.7.3. Application Dose of Bio-Friendly Formulations and Recommended Fungicide Rizolex-T 50% Wettable Powder (WP)
2.7.4. Greenhouse Experiment
2.7.5. Open-Field Experiment
2.7.6. Disease Evaluation
2.8. Quantitative Real-Time PCR Analysis of the Defense-Related Genes
2.8.1. Plant Total RNA Extraction and cDNA Synthesis
2.8.2. qRT-PCR Assay and Data Analysis
2.9. Statistical Analysis
3. Results
3.1. Antagonistic Effect of Actinobacterial Isolates against S. rolfsii
3.2. Qualitative and Quantitative of Chitinase Production from S. cellulosae Actino 48
3.3. Detection of Interaction between Actinobacteria and S. rolfsii
3.4. Characterization of Talc-Based Formulations of S. cellulosae Actino 48
3.4.1. SEM
3.4.2. FTIR Spectroscopy
3.4.3. Particle Size Analysis
3.5. Application of Bio-Friendly Formulations of Chitinase-Producing S. cellulosae Actino 48
3.5.1. Greenhouse Experiment
3.5.2. Open-Field Experiment
3.6. Quantitative Real-Time PCR Analysis of the Defense-Related Genes
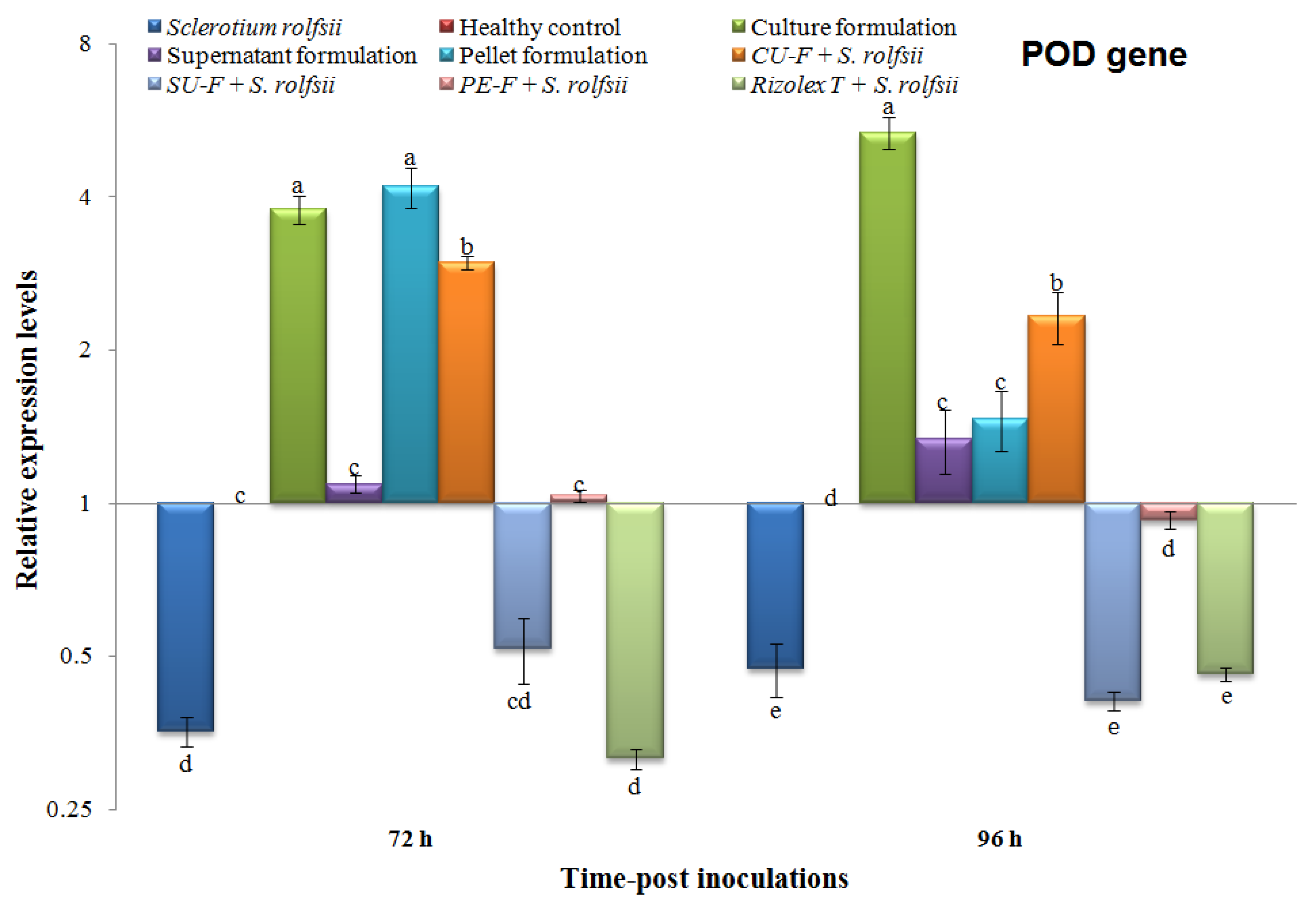
3.6.1. Effects on the Transcriptional Level of Peroxidase (POD)
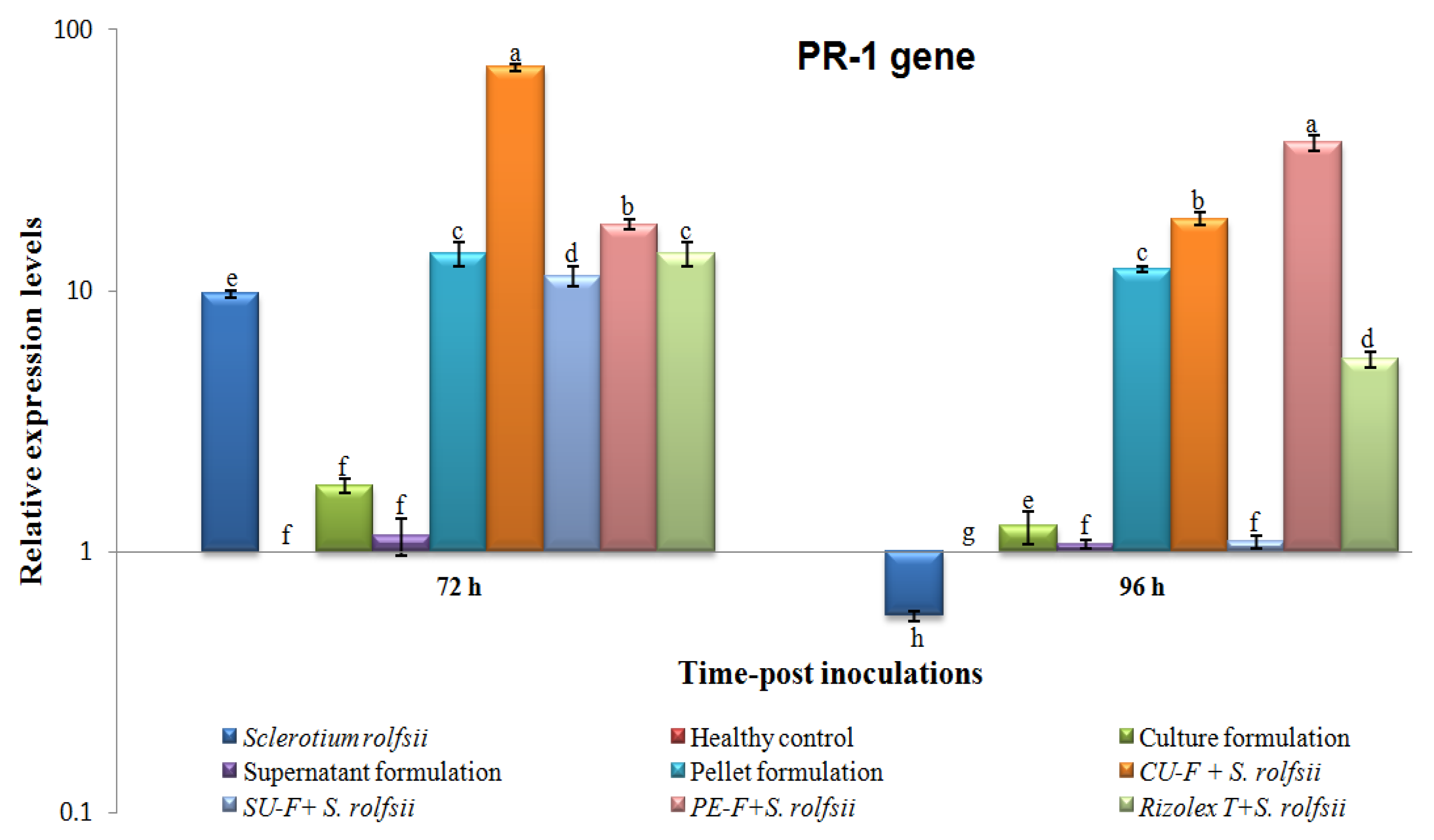
3.6.2. Effects on Transcriptional Level of Pathogenesis-Related Protein 1 (PR-1)
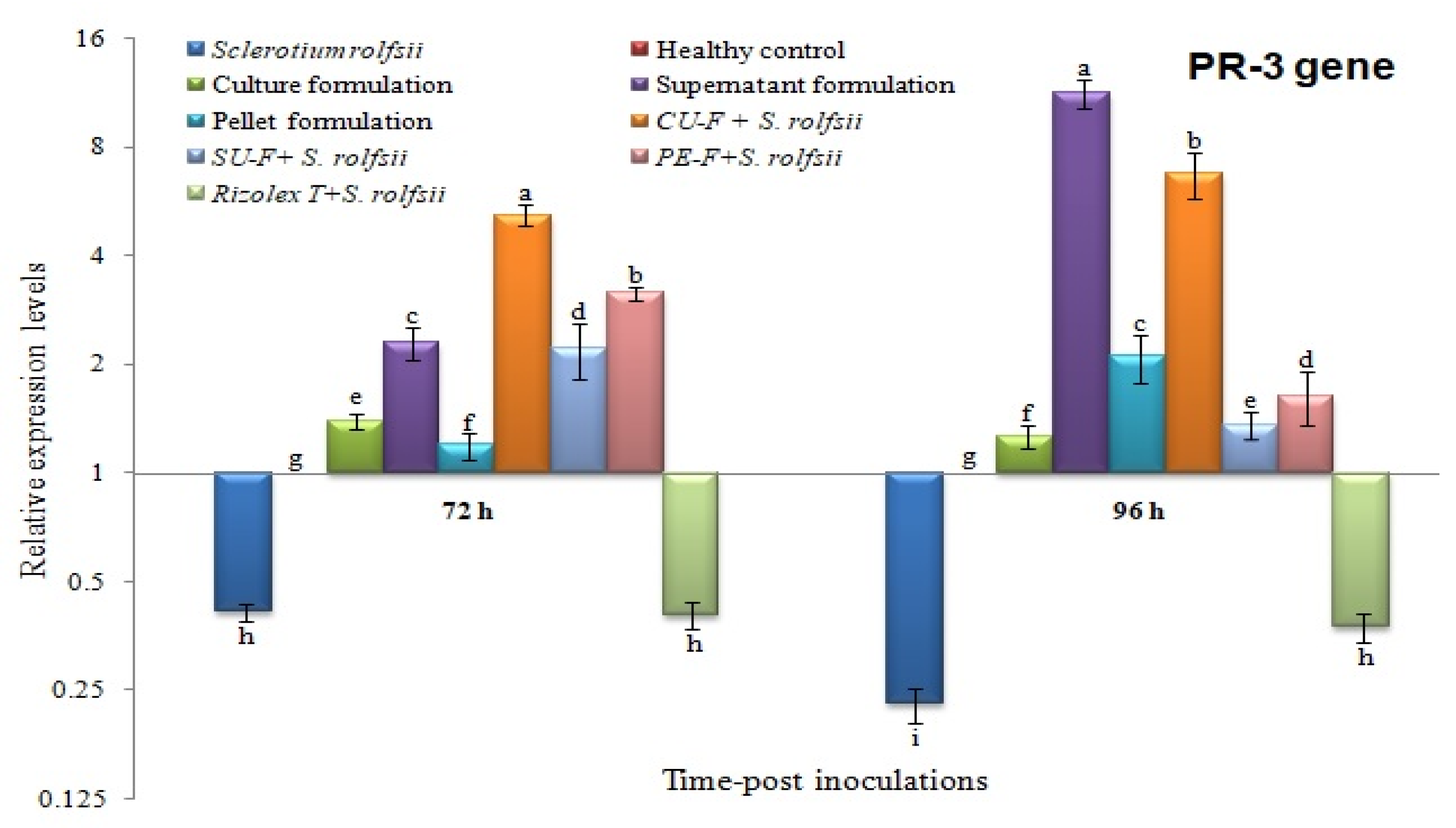
3.7. Effects on Transcriptional Level of Chitinase (PR-3)
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Aycock, R. Stem Rot and other Diseases Caused by Sclerotium Rolfsii or the Status of Rolfs’ Fungus after 70 Years; North Carolina State University: Raleigh, NC, USA, 1966. [Google Scholar]
- Punja, Z.K. Sclerotium rolfsii, a pathogen of many plant species. In Advances in Plant Pathology; Sidhu, G.S., Ed.; Academic Press: London, UK, 1988; pp. 523–534. [Google Scholar]
- Chet, I. Ultrastructural basis of sclerotial survival in soil. Microbiol. Ecol. 1975, 2, 194–200. [Google Scholar] [CrossRef]
- Li, Q.; Ning, P.; Zheng, L.; Huang, J.; Li, G.; Hsiang, T. Effects of volatile substances of Streptomyces globisporus JK-1 on control of Botrytis cinerea on tomato fruit. Biol. Control 2012, 61, 113–120. [Google Scholar] [CrossRef]
- On, A.; Wong, F.; Ko, Q.; Tweddell, R.J.; Antoun, H.; Avis, T.J. Antifungal effects of compost tea microorganisms on tomato pathogens. Biol. Control 2015, 80, 63–69. [Google Scholar] [CrossRef]
- Jiménez-Díaz, R.M.; Castillo, P.; del Mar Jiménez-Gasco, M.; Landa, B.B.; Navas-Cortés, J.A. Fusarium wilt of chickpeas: Biology, ecology and management. Crop Protec. 2015, 73, 16–27. [Google Scholar] [CrossRef]
- Antoraz, S.; Santamaría, R.I.; Díaz, M.; Sanz, D.; Rodríguez, H. Toward a new focus in antibiotic and drug discovery from the Streptomyces arsenal. Front. Microbiol. 2015, 6, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Errakhi, R.; Lebrihi, A.; Barakate, M. In vitro and in vivo antagonism of actinomycetes isolated from Moroccan rhizospherical soils against Sclerotium rolfsii: A causal agent of root rot on sugar beet (Beta vugaris L.). J. Appl. Microbiol. 2009, 107, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Bubici, G.; Marsico, A.D.; D’Amico, M.; Amenduni, M.; Cirulli, M. Evaluation of Streptomyces spp. for the biological control of corky root of tomato and Verticillium wilt of eggplant. Appl. Soil Ecol. 2013, 72, 128–134. [Google Scholar] [CrossRef]
- Tamreihao, K.; Ningthoujam, D.S.; Nimaichand, S.; Singh, E.S.; Reena, P.; Singh, S.H.; Nongthomba, U. Biocontrol and plant growth promoting activities of a Streptomyces corchorusii strain UCR3-16 and preparation of powder formulation for application as biofertilizer agents for rice plant. Microbiol. Res. 2016, 192, 260–270. [Google Scholar] [CrossRef]
- Singh, S.P.; Gupta, R.; Gaur, R.; Srivastava, A.K. Antagonistic actinomycetes mediated resistance in Solanum lycopersicon Mill. against Rhizoctonia solani Kühn. Proc. Natl. Acad. Sci. India. Sect. B Biol. Sci. 2017, 87, 789–798. [Google Scholar] [CrossRef]
- Abo-Zaid, G.A.; Soliman, N.A.; Abdullah, A.S.; El-Sharouny, E.E.; Matar, S.M.; Sabry, S.A. Maximization of siderophores production from biocontrol agents, Pseudomonas aeruginosa f2 and Pseudomonas fluorescens JY3 using batch and exponential fed-batch fermentation. Processes 2020, 8, 455. [Google Scholar] [CrossRef]
- Intra, B.; Mungsuntisuk, I.; Nihira, T.; Igarashi, Y.; Panbangred, W. Identification of actinomycetes from plant rhizospheric soils with inhibitory activity against Colletotrichum spp., the causative agent of anthracnose disease. BMC Res. Notes 2011, 4, 98. [Google Scholar] [CrossRef] [Green Version]
- Vurukonda, S.K.; Giovanardi, D.; Stefani, E. Plant Growth Promoting and Biocontrol Activity of Streptomyces spp. as Endophytes. Inter. J. Molecu. Sci. 2018, 19, 952. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Saxena, R.K.; Chaturvedi, P.; Virdi, J.S. Chitinase production by Streptomyces viridificans: Its potential in fungal cell wall lysis. J. Appl. Bacteriol. 1995, 78, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M. Endophytic actinomycetes: Biocontrol agents and growth promoters. In Bacteria in Agrobiology: Plant Growth Responses; Maheshwari, D.K., Ed.; Elsevier Academic Press: San Diego, CA, USA, 2011; pp. 201–220. [Google Scholar]
- Reddy, K.R.K.; Jyothi, G.; Sowjanya, C.; Kusumanjali, K.; Malathi, N.; Reddy, K.R.N. Plant Growth-Promoting Actinomycetes: Mass Production, Delivery systems, and commercialization. In Plant Growth Promoting Actinobacteria; Subramaniam, G., Ed.; Springer: Singapore, 2016; pp. 287–298. [Google Scholar]
- El-Sayed, M.A.; Valadon, L.R.G.; El-Shanshoury, A. Biosynthesis and metabolism of indole-3-acetic acid in Streptomyces mutabilis and Streptomyces atroolivaceus. Microbiol. Lett. 1987, 36, 85–95. [Google Scholar]
- El-Shanshoury, A.R. Biosynthesis of indole-3-acetic acid in Streptomyces atroolivaceus and its changes during spore germination and mycelial growth. Microbiol. Lett. 1991, 67, 159–164. [Google Scholar]
- El-Tarabily, K.A. Promotion of tomato (Lycopersicon esculentum Mill.) plant growth by rhizosphere competent 1-aminocyclopropane-1-carboxylic acid deaminase-producing streptomycete actinomycetes. Plant Soil 2008, 308, 161–174. [Google Scholar] [CrossRef]
- Minuto, A.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Control of soilborne pathogens of tomato using a commercial formulation of Streptomyces griseoviridis and solarization. Crop Protect. 2006, 25, 468–475. [Google Scholar] [CrossRef]
- Berg, G.; Marten, P.; Minkwitz, A.; Brückner, S. Efficient biological control of plant fungal diseases by Streptomyces sp. DSMZ 12424. J. Plant Dis. Protect. 2010, 108, 1–10. [Google Scholar]
- Zeng, W.; Wang, D.; Kirk, W.; Hao, J. Use of Coniothyrium minitans and other microorganisms for reducing Sclerotinia sclerotiorum. Biol. Control 2012, 60, 225–232. [Google Scholar] [CrossRef]
- Boukaew, S.; Chuenchit, S.; Petcharat, V. Evaluation of Streptomyces spp. for biological control of Sclerotium root and stem rot and Ralstonia wilt of chili pepper. BioControl 2011, 56, 365–374. [Google Scholar] [CrossRef]
- Jacob, S.; Sajjalaguddam, R.R.; Sudini, H.K. Streptomyces sp. RP1A-12 mediated control of peanut stem rot caused by Sclerotium rolfsii. J. Integ. Agri. 2018, 174, 892–900. [Google Scholar] [CrossRef]
- Trivedi, P.; Pandey, A.; Palni, L.M.S. In vitro evaluation of antagonistic properties of Pseudomonas corrugate. Microbiol. Res. 2008, 163, 329–336. [Google Scholar] [CrossRef]
- Boller, T.; Mauch, F. Colourimetric assay for chitinase. Method. Enzymol. 1988, 161, 430–435. [Google Scholar]
- Vidhyasekaran, P.; Muthamilan, M. Development of formulation of Pseudomonas fluorescens for control of chickpea wilt. Plant Dis. 1995, 79, 782–786. [Google Scholar] [CrossRef]
- Hussien, Z.N.; Mahmoud, E.Y.; Metwaly, A.H.; Sobhy, H.M. Effect of some antagonistic bacteria in reducing of peanut damping-off, root and pod rot incidence caused by Rhizoctonia solani. J. Plant Protec. Path. 2012, 3, 1173–1187. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Sanan-Mishra, N. Differential expression profiles of tomato miRNAs induced by Tobacco mosaic virus. J. Agric. Sci. Technol. 2019, 21, 475–485. [Google Scholar]
- Abo-Zaid, G.A.; Matar, S.M.; Abdelkhalek, A. Induction of plant resistance against tobacco mosaic virus using the biocontrol agent Streptomyces cellulosae isolate Actino 48. Agronomy 2020, 10, 1620. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Al-Askar, A.A.; Hafez, E. Differential induction and suppression of the potato innate immune system in response to Alfalfa mosaic virus infection. Physiol. Mol. Plant Pathol. 2020, 110, 101485. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Gayed, M.A.; Abo-Zaid, G.A.; Matar, S.M.; Hafez, E.E. Fermentation, formulation and evaluation of PGPR Bacillus subtilis isolate as a bioagent for reducing occurrence of peanut soil-borne diseases. J. Integ. Agric. 2019, 18, 2080–2092. [Google Scholar]
- Martinez-Alvarez, J.C.; Castro-Martiez, C.; Sanchez-Pena, P.; Gutierrez-Dorado, R.; Maldonado- Mendoza, I.E. Development of a powder formulation based on Bacillus cereus sensulato strain B25 spores for biological control of Fusarium verticillioides in maize plants. World J. Microb. Biotechn. 2016, 33, 1–10. [Google Scholar]
- De-Oliveira, M.F.; Da Silva, M.G.; Van Der Sand, S.T. Anti-phytopathogen potential of endophytic actinobacteria isolated from tomato plants (Lycopersicon esculentum) in southern Brazil, and characterization of Streptomyces sp. R18, a potential biocontrol agent. Res. Microb. 2010, 161, 565–572. [Google Scholar] [CrossRef]
- Passari, A.K.; Mishra, V.K.; Gupta, V.K.; Yadav, M.K.; Saikia, R.; Singh, B.P.; Virolle, M.J. In vitro and in vivo plant-growth-promoting activities and DNA fingerprinting of antagonistic endophytic actinomycetes associates with medicinal plants. PLoS ONE 2015, 10, e0139468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conn, V.M.; Walker, A.R.; Franco, C.M.M. Endophytic actinobacteria induce defense pathways in Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2008, 21, 208–218. [Google Scholar] [CrossRef] [Green Version]
- Adhilakshmi, M.; Latha, P.; Paranidharan, V.; Balachandar, D.; Ganesamurthy, K.; Velazhahan, R. Biological control of stem rot of groundnut (Arachis hypogaea L.) caused by Sclerotium rolfsii Sacc. with actinomycetes. Arch. Phytopathol. Plant Protect. 2014, 47, 298–311. [Google Scholar] [CrossRef]
- Ensign, J.C. Introduction to the actinomycetes. In The prokaryotes; Balows, A., Trijper, H.G., Dworkin, M., Harder, W., Schleifer, K.H., Eds.; Springer: New York, NY, USA, 1992; pp. 811–815. [Google Scholar]
- Behal, V. Bioactive products from Streptomyces. Adv. Appl. Microbiol. 2000, 47, 113–157. [Google Scholar]
- Pattanapipitpaisal, P.; Kamlandharn, R. Screening of chitinolytic actinomycetes for biological control of Sclerotium rolfsii stem rot disease of chilli. Songklanakarin J. Sci. Technol. 2012, 34, 387–393. [Google Scholar]
- Singh, S.P.; Gaur, R. Evaluation of antagonistic and plant growth promoting activities of chitinolytic endophytic actinomycetes associated with medicinal plants against Sclerotium rolfsii in chickpea. J. Appl. Microb. 2016, 121, 506–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ningthoujam, D.S.; Sanasam, S.; Tamreihao, K.; Nimaichand, S. Antagonistic activities of local actinoimycete isolates against rice fungal pathogens. Afr. J. Microbiol. Res. 2009, 3, 737–742. [Google Scholar]
- Errakhi, R.; Bouteau, F.; Lebrihi, A.; Barakate, M. Evidences of biological control capacities of Streptomyces spp. against Sclerotium rolfsii responsible for damping-off disease in sugar beet (Beta vulgaris L.). World J. Microb. Biotech. 2007, 23, 1503–1509. [Google Scholar] [CrossRef]
- Zacky, F.M.; Ting, A.S.Y. Biocontrol of Fusarium oxysporum f.sp. cubense tropical race 4 by formulated cells and cell-free extracts of Streptomyces griseus in sterile soil environment. Bioc. Sci. Tech. 2015, 25, 685–696. [Google Scholar] [CrossRef]
- Pornsuriya, C.; Sunpapao, A. Formulations of Streptomyces philanthi RL-1-178 biocontrol agent against Sclerotium root and stem rot of chili pepper (Capsicum annuum L.). Philipp. Agric. Scient. 2014, 97, 273–279. [Google Scholar]
- Yingsanga, P.; Srilaong, V.; Kanlayanarat, S.; Noichinda, S.; McGlasson, W.B. Relationship between browning and related enzymes (PAL, PPO and POD) in rambutan fruit (Nephelium lappaceum Linn.) cvs. Rongrien and See-Chompoo. Posthar. Bio. Techno. 2008, 50, 164–168. [Google Scholar] [CrossRef]
- Lin, J.; Gong, D.; Zhu, S.; Zhang, L.; Zhang, L. Expression of PPO and POD genes and contents of polyphenolic compounds in harvested mango fruits in relation to Benzothiadiazole-induced defense against anthracnose. Scient. Horticult. 2011, 130, 85–89. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoegen, E.; Strömberg, A.; Pihlgren, U.; Kombrink, E. Primary structure and tissue-specific expression of the pathogenesis-related protein PR-1b in potato. Mol. Plant Pathol. 2002, 3, 329–345. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Al-Askar, A. Green synthesized ZnO nanoparticles mediated by Mentha Spicata extract induce plant systemic resistance against tobacco mosaic virus. Appl. Sci. 2020, 10, 5054. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Salem, M.Z.M.; Ali, H.M.; Kordy, A.M.; Salem, A.Z.M.; Behiry, S.I. Antiviral, antifungal, and insecticidal activities of Eucalyptus bark extract: HPLC analysis of polyphenolic compounds. Microb. Pathog. 2020, 147, 104383. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindschedler, L.V.; Dewdney, J.; Blee, K.A.; Stone, J.M.; Asai, T.; Plotnikov, J.; Denoux, C.; Hayes, T.; Gerrish, C.; Davies, D.R. Peroxidase-dependent apoplastic oxidative burst in Arabidopsis required for pathogen resistance. Plant J. 2006, 47, 851–863. [Google Scholar] [CrossRef] [Green Version]
- Xayphakatsa, K.; Tsukiyama, T.; Inouye, K.; Okumoto, Y.; Nakazaki, T.; Tanisaka, T. Gene cloning, expression, purification and characterization of rice (Oryza sativa L.) class II chitinase CHT11. Enz. Microb. Technol. 2008, 43, 19–24. [Google Scholar] [CrossRef]
- Schneider, K.T.; van de Mortel, M.; Bancroft, T.J.; Braun, E.; Nettleton, D.; Nelson, R.T.; Frederick, R.D.; Baum, T.J.; Graham, M.A.; Whitham, S.A. Biphasic gene expression changes elicited by Phakopsora pachyrhizi in soybean correlate with fungal penetration and haustoria formation. Plant Physiol. 2011, 157, 355–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Huang, Y.; Li, Y.; Dong, J.; Liu, X.; Li, C. Biocontrol of Rhizoctonia solani via induction of the defense mechanism and antimicrobial compounds produced by Bacillus subtilis SL-44 on Pepper (Capsicum annuum L.). Front. Microbiol. 2019, 10, 1–12. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Abbreviation | Direction | Sequence (5′…3′) |
|---|---|---|---|
| Pathogenesis related protein 1 | PR-1 | Forward | GTTCCTCCTTGCCACCTTC |
| Reverse | TATGCACCCCCAGCATAGTT | ||
| Chitinase | PR-3 | Forward | ATGGAGCATTGTGCCCTAAC |
| Reverse | TCCTACCAACATCACCACCA | ||
| Peroxidase | POD | Forward | GGAATGTTGGGTTAGGCAGA |
| Reverse | GCTTCCCCTGTTGTGTGAG | ||
| Beta-actin | β-actin | Forward | TGGCATACAAAGACAGGACAGCCT |
| Reverse | ACTCAATCCCAAGGCCAACAGAGA |
| Treatments | % Damping-Off | % Root Rot | % Survival |
|---|---|---|---|
| Sclerotium rolfsii | * 27.5 ± 5.00 a ** | 32.5 ± 9.57 a | 40.0 ± 14.14 f |
| Healthy control | 07.5 ± 5.00 de | 05.0 ± 5.77 de | 87.5 ± 05.00ab |
| Culture formulation (CU-F) | 05.0 ± 5.77 e | 02.5 ± 5.00 e | 92.5 ± 09.57 a |
| Supernatant formulation (SU-F) | 07.5 ± 5.00 de | 05.0 ± 5.77 de | 87.5 ± 05.00 ab |
| Pellet formulation (PE-F) | 07.5 ± 9.57 de | 07.5 ± 5.00 de | 85.0 ± 12.91 abc |
| CU-F + S. rolfsii | 15.0 ± 5.77 cd | 12.5 ± 5.00 cd | 72.5 ± 09.57 cd |
| SU-F + S. rolfsii | 17.5 ± 5.00 bc | 20.0 ± 8.16 bc | 62.5 ± 12.58 de |
| PE-F + S. rolfsii | 25.0 ± 5.77 ab | 25.0 ± 5.77 ab | 50.0 ± 00.00 ef |
| Rizolex-T + S. rolfsii | 12.5 ± 9.57 cde *** | 10.0 ± 8.16 de | 77.5 ± 12.58 bc |
| Least significant difference (LSD 5%) | 09.47 | 09.67 | 14.64 |
| Treatments | Dry Weight of Shoot System (g pot−1) | Dry weight of Root System (g pot−1) | Percentage of Infected Pods (%) | Percentage of Apparent Healthy Pods (%) |
|---|---|---|---|---|
| Sclerotium rolfsii | * 08.31 ± 1.47 g ** | 1.48 ± 0.28 f | 56.35 ± 5.33 a | 43.65 ± 5.33 e |
| Healthy control | 36.19 ± 3.23 bc | 3.23 ± 0.25 bcd | 10.69 ± 5.62 de | 89.31 ± 5.62 ab |
| Culture formulation (CU-F) | 41.56 ± 3.79 a | 4.59 ± 0.58 a | 06.45 ± 0.90 e | 93.55 ± 0.90 a |
| Supernatant formulation (SU-F) | 37.94 ± 2.90 ab | 3.52 ± 0.42 bc | 11.23 ± 5.04 cde | 88.77 ± 5.04 abc |
| Pellet formulation (PE-F) | 28.87 ± 2.16 e | 3.34 ± 0.42 bc | 10.67 ± 3.64 de | 89.33 ± 3.64 ab |
| CU-F + S. rolfsii | 29.70 ± 3.16 de | 3.01 ± 0.12 cde | 14.08 ± 2.33 bcd | 85.92 ± 2.33 bcd |
| SU-F+ S. rolfsii | 26.28 ± 2.54 e | 2.80 ± 0.18 de | 16.97 ± 1.99 b | 83.03 ± 1.99 d |
| PE-F + S. rolfsii | 19.95 ± 3.15 f | 2.57 ± 0.38 e | 16.50 ± 3.72 bc | 83.50 ± 3.72 cd |
| Rizolex-T + S. rolfsii | 33.17 ± 1.12 cd *** | 3.58 ± 0.41 b | 09.13 ± 3.14 de | 90.87 ± 3.14 ab |
| LSD 5% | 3.98 | 0.53 | 5.57 | 5.57 |
| Treatments | Dry weight of Infected Pods (g pot−1) | Dry Weight of Healthy Pods (g pot−1) | Dry Weight of Total Pods (g pot−1) |
|---|---|---|---|
| Sclerotium rolfsii | * 03.55 ± 1.60 a ** | 11.09 ± 6.88 c | 14.63 ± 7.92 d |
| Healthy control | 01.74 ± 1.22 bc | 24.68 ± 8.62 ab | 26.42 ± 7.88 abc |
| Culture formulation (CU-F) | 00.76 ± 0.05 c | 30.71 ± 4.71 a | 31.48 ± 4.73 ab |
| Supernatant formulation (SU-F) | 01.71 ± 0.43 bc | 24.35 ± 8.59 ab | 26.06 ± 8.78 abc |
| Pellet formulation (PE-F) | 01.73 ± 0.14 bc | 26.26 ± 9.02 ab | 27.99 ± 9.06 abc |
| CU-F + S. rolfsii | 01.39 ± 0.25 bc | 24.18 ± 1.79 ab | 25.57 ± 1.96 abcd |
| SU-F+ S. rolfsii | 02.27 ± 0.82 b | 18.65 ± 1.75 bc | 20.92 ± 1.79 bcd |
| PE-F + S. rolfsii | 01.68 ± 0.79 bc | 16.97 ± 4.83 bc | 18.64 ± 5.39 cd |
| Rizolex-T + S. rolfsii | 01.09 ± 0.46 c *** | 34.58 ± 13.61 a | 35.67 ± 13.58 a |
| LSD 5% | 1.17 | 10.96 | 11.10 |
| Treatments | Damping-off (%) | Efficacy (%) | Root Rot (%) | Efficacy (%) | Healthy Survival (%) |
|---|---|---|---|---|---|
| Trial field season 2018 | |||||
| Infected control | * 20.22 ± 1.93 a ** | 21.52 ± 9.45 a | 58.26 ± 1.88 d | ||
| Culture formulation | 12.61 ± 1.67 c | 37.63 | 11.74 ± 2.07 c | 45.45 | 75.65 ± 3.69 b |
| Supernatant formulation | 15.43 ± 2.29 bc | 23.69 | 12.83 ± 2.28 c | 40.41 | 71.74 ± 4.52 bc |
| Pellet formulation | 16.74 ± 1.31 b | 17.20 | 16.08 ± 1.12 b | 22.22 | 67.18 ± 0.44 c |
| Rizolex-T | 08.04 ± 1.09 d | 60.22 | 07.40 ± 0.50 d | 65.66 | 84.56 ± 1.09 a |
| LSD 5% | 2.87 | 2.89 | 4.64 | ||
| Trial field season 2019 | |||||
| Infected control | 22.18 ± 1.67 a | 21.31 ± 3.06 a | 56.51 ± 1.59 d | ||
| Culture formulation | 13.48 ± 2.30 c | 39.22 | 10.87 ± 1.66 cd | 48.98 | 75.65 ± 2.75 b |
| Supernatant formulation | 16.74 ± 0.83 b | 24.51 | 14.56 ± 1.65 b | 31.63 | 68.70 ± 1.42 c |
| Pellet formulation | 15.87 ± 1.65 bc | 28.43 | 13.26 ± 2.06 bc | 37.78 | 70.87 ± 1.12 c |
| Rizolex-T | 08.26 ± 1.51 d *** | 62.75 | 08.70 ± 1.23 d | 59.18 | 83.04 ± 2.70 a |
| LSD 5% | 2.67 | 3.27 | 3.41 | ||
| Treatments | Dry weight of shoot system (g plot−1) | Increase Z (%) | Dry weight of root system (g plot−1) | Increase Z (%) |
|---|---|---|---|---|
| Trial field season 2018 | ||||
| Infected control | * 095.81 ± 09.37 d ** | 19.27 ± 2.75 c | ||
| Culture formulation | 385.01 ± 09.90 a | 75.11 | 41.65 ± 1.81 a | 53.73 |
| Supernatant formulation | 289.05 ± 10.46 b | 66.87 | 32.14 ± 2.20 b | 40.03 |
| Pellet formulation | 265.91 ± 13.76 c | 63.97 | 34.60 ± 3.55 b | 44.30 |
| Rizolex-T | 380.20 ± 25.91 a | 74.80 | 40.92 ± 1.44 a | 52.87 |
| LSD 5% | 21.24 | 3.94 | ||
| Trial field season 2019 | ||||
| Infected control | 091.25 ± 07.96 c | 17.81 ± 2.34 c | ||
| Culture formulation | 370.47 ± 07.59 a | 75.37 | 39.62 ± 1.61 a | 55.06 |
| Supernatant formulation | 275.86 ± 22.62 b | 66.92 | 30.80 ± 1.80 b | 42.18 |
| Pellet formulation | 257.28 ± 15.66 b | 64.53 | 30.96 ± 1.95 b | 42.47 |
| Rizolex-T | 373.56 ± 25.85 a *** | 75.57 | 40.28 ± 2.28 a | 55.79 |
| LSD 5% | 27.89 | 3.22 | ||
| Treatments | Percentage of Infected Pods (%) | Percentage of Apparent Healthy Pods (%) |
|---|---|---|
| Trial field season 2018 | ||
| Infected control | * 47.22 ± 3.47 a ** | 52.78 ± 3.47 d |
| Culture formulation | 09.07 ± 1.05 c | 90.93 ± 1.05 b |
| Supernatant formulation | 12.43 ± 2.38 b | 87.57 ± 2.38 c |
| Pellet formulation | 13.74 ± 1.31 b | 86.26 ± 1.31 c |
| Rizolex-T | 05.74 ± 0.75 d | 94.26 ± 0.75 a |
| LSD 5% | 3.08 | 3.08 |
| Trial field season 2019 | ||
| Infected control | 39.09 ± 1.73 a | 60.91 ± 1.73 c |
| Culture formulation | 07.24 ± 0.78 c | 92.76 ± 0.78 a |
| Supernatant formulation | 10.61 ± 0.46 b | 89.39 ± 0.46 b |
| Pellet formulation | 10.61 ± 1.81 b | 89.39 ± 1.81 b |
| Rizolex-T | 05.31 ± 1.02 c *** | 94.69 ± 1.02 a |
| LSD 5% | 2.00 | 2.00 |
| Treatments | Dry Weight of Infected Pods (g plot−1) | Dry Weight of Healthy Pods (g plot−1) | Dry Weight of Total Pods (g plot−1) | Yield Increase Z (%) |
|---|---|---|---|---|
| Trial Field Season 2018 | ||||
| Infected control | * 37.31 ± 4.08 a ** | 194.51 ± 09.67 d | 231.82 ± 11.85 d | ----- |
| Culture formulation | 15.12 ± 1.68 c | 406.96 ± 07.15 b | 422.08 ± 08.00 b | 45.08 |
| Supernatant formulation | 19.35 ± 0.76 b | 333.81 ± 18.10 c | 353.16 ± 17.73 c | 34.36 |
| Pellet formulation | 21.64 ± 2.09 b | 350.37 ± 13.72 c | 372.01 ± 14.16 c | 37.68 |
| Rizolex-T | 13.32 ± 2.35 c | 489.94 ± 12.11 a | 503.26 ± 13.68 a | 53.94 |
| LSD 5% | 3.36 | 20.17 | 21.58 | |
| Trial Field Season 2019 | ||||
| Infected control | 33.27 ± 3.92 a | 202.14 ± 05.17 e | 235.41 ± 08.87 e | ----- |
| Culture formulation | 13.27 ± 1.52 c | 428.55 ± 20.66 b | 441.82 ± 21.33 b | 46.72 |
| Supernatant formulation | 17.08 ± 1.48 b | 361.86 ± 13.01 c | 378.94 ± 11.59 c | 37.88 |
| Pellet formulation | 19.17 ± 2.34 b | 328.66 ± 05.97 d | 347.83 ± 05.01 d | 32.32 |
| Rizolex-T | 10.98 ± 1.64 c *** | 482.23 ± 16.57 a | 493.21 ± 15.53 a | 52.27 |
| LSD 5% | 3.51 | 23.31 | 23.41 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abo-Zaid, G.; Abdelkhalek, A.; Matar, S.; Darwish, M.; Abdel-Gayed, M. Application of Bio-Friendly Formulations of Chitinase-Producing Streptomyces cellulosae Actino 48 for Controlling Peanut Soil-Borne Diseases Caused by Sclerotium rolfsii. J. Fungi 2021, 7, 167. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030167
Abo-Zaid G, Abdelkhalek A, Matar S, Darwish M, Abdel-Gayed M. Application of Bio-Friendly Formulations of Chitinase-Producing Streptomyces cellulosae Actino 48 for Controlling Peanut Soil-Borne Diseases Caused by Sclerotium rolfsii. Journal of Fungi. 2021; 7(3):167. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030167
Chicago/Turabian StyleAbo-Zaid, Gaber, Ahmed Abdelkhalek, Saleh Matar, Mai Darwish, and Muhammad Abdel-Gayed. 2021. "Application of Bio-Friendly Formulations of Chitinase-Producing Streptomyces cellulosae Actino 48 for Controlling Peanut Soil-Borne Diseases Caused by Sclerotium rolfsii" Journal of Fungi 7, no. 3: 167. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030167