
Chaperone Networks in Fungal Pathogens of Humans


Michael Smith Laboratories, Department of Microbiology and Immunology, University of British Columbia, Vancouver, BC V6T 1Z4, Canada
*
Author to whom correspondence should be addressed.
J. Fungi 2021, 7(3), 209; https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030209
Submission received: 6 February 2021
/
Revised: 5 March 2021
/
Accepted: 10 March 2021
/
Published: 12 March 2021
(This article belongs to the Special Issue Systems Biology in Fungal Research)
{kind=link}
{kind=link}
{kind=link}
Abstract
:The heat shock proteins (HSPs) function as chaperones to facilitate proper folding and modification of proteins and are of particular importance when organisms are subjected to unfavourable conditions. The human fungal pathogens are subjected to such conditions within the context of infection as they are exposed to human body temperature as well as the host immune response. Herein, the roles of the major classes of HSPs are briefly reviewed and their known contributions in human fungal pathogens are described with a focus on Candida albicans, Cryptococcus neoformans, and Aspergillus fumigatus. The Hsp90s and Hsp70s in human fungal pathogens broadly contribute to thermotolerance, morphological changes required for virulence, and tolerance to antifungal drugs. There are also examples of J domain co-chaperones and small HSPs influencing the elaboration of virulence factors in human fungal pathogens. However, there are diverse members in these groups of chaperones and there is still much to be uncovered about their contributions to pathogenesis. These HSPs do not act in isolation, but rather they form a network with one another. Interactions between chaperones define their specific roles and enhance their protein folding capabilities. Recent efforts to characterize these HSP networks in human fungal pathogens have revealed that there are unique interactions relevant to these pathogens, particularly under stress conditions. The chaperone networks in the fungal pathogens are also emerging as key coordinators of pathogenesis and antifungal drug tolerance, suggesting that their disruption is a promising strategy for the development of antifungal therapy.
1. Introduction
Human fungal pathogens must be able to survive and proliferate in the host environment despite several unfavourable conditions. A major barrier to systemic infections in humans is the elevated mammalian body temperature compared to the ambient environment [1]. Human body temperature effectively restricts the growth of most fungi and, indeed, the majority of fungal infections are superficial and occur on the skin where temperatures are less restrictive [2,3]. Several human fungal pathogens also undergo morphological changes such as the transition between yeast and hyphae in response to temperature differences [4]. In order to survive at core human body temperature and to undergo these morphological changes, fungi must be able to cope with the proteotoxic stress induced at high temperature and upon flux in the demand for protein production. The heat shock proteins (HSPs) are one of the major groups of proteins which help respond to and mitigate these stresses. Herein, we will describe the general roles and classifications of HSPs, their known roles in the major human fungal pathogens, and the interactions between HSPs which coordinate chaperoning activity.
Classically, HSPs are defined as those proteins that are upregulated upon temperature upshift and stress induction as well as those which share a high degree of sequence similarity to established categories of HSPs [5]. Despite the general property of upregulation under stress conditions, there are also examples of HSPs that are constitutively expressed. Therefore, it is speculated that these proteins may have evolved to fulfill a proactive role of ensuring proper folding of nascent proteins and thus preventing the accumulation of proteotoxic stress rather than as a mechanism to respond to stresses such as heat shock [6]. The HSPs are involved in multiple processes including folding proteins de novo, stabilizing protein conformation under stress conditions, and modulating protein conformation to regulate their activity [7]. This modulation can involve individual proteins or multiprotein complexes which must be assembled or disassembled for their function [8].
HSPs have typically been named based on their sizes and classified based on their sequence similarity. This often reflects how they function, but provides limited information about their clients or the pathways in which they participate. In general, there are ATP-dependent HSPs including Hsp90, Hsp70, chaperonins, and disaggregases which undergo conformational changes upon ATP hydrolysis to facilitate protein folding, complex assembly, or disaggregation. There are also energy-independent passive chaperones such as the small HSPs which normally act as holdases to prevent protein aggregation. There are also many co-chaperones required to recruit clients as well as to facilitate the ATPase activity of their chaperones; however, we will focus on the Hsp40/J-domain co-chaperones of Hsp70s as they share a highly conserved domain and have emerging importance in fungal pathogens. These energy-dependent and -independent chaperones as well as their co-chaperones coordinately ensure that cells can function in normal and stressed conditions including those relevant to proliferation in a human host. The major classes of HSPs are reviewed below with descriptions of their major reported functions in the model species Saccharomyces cerevisiae to provide context, as well as current information on their known roles in the major human fungal pathogens (Figure 1).
2. Major Classes of HSPs
2.1. Hsp90s
Hsp90s are among the best studied HSPs due to the abundance of these proteins in cells. Hsp90 is an ATP-dependent chaperone which promotes substrate folding and participates in the activation of near native proteins through inducing conformational change [9,10]. Hsp90 has an N-terminal ATP-binding pocket, a C-terminal domain for homodimerization, and a C-terminal EEVD motif which promotes interactions with the tetratricopeptide repeat domains of several co-chaperones (Figure 2A) [11]. In S. cerevisiae, there are two paralogs encoding Hsp90 proteins and it is essential that cells have at least one functional copy. Hsp90s are among the most abundant proteins in the yeast cytosol under normal conditions [12]. One paralog (HSC82) is constitutively expressed and the other (HSP82) is induced upon heat shock; therefore, Hsp90 abundance is further increased upon heat shock [13]. Early reports on Hsp90s found that they interacted with steroid hormone receptors, kinases, actin, tubulin, calmodulin, and the SSA subfamily of Hsp70s [8,9,13,14,15]. Work using chemical genetic screens, synthetic genetic arrays, affinity purification, and yeast two hybrid analysis has expanded our understanding of the extensive nature of the Hsp90 interaction network. That is, Hsp90 interacts with approximately 10% of all yeast proteins, and was confirmed to interact with kinases, transcription factors, and other chaperones [16]. Chemical genetic screens under different conditions show that Hsp90 interacts genetically with the secretory pathway and cellular transport under normal growth conditions whereas Hsp90 interacted genetically with cell cycle, meiosis, and cytokinesis pathways upon elevated temperature (37 °C) [17]. More recently, proteomic approaches to identify interacting partners of the two paralogs encoding Hsp90 revealed that the interactomes of each protein are very similar in S. cerevisiae [18]. The clients that were identified reinforced previous studies which found that most client proteins were ligand-binding molecules. However, this approach revealed that most of these clients were enzymes involved in biosynthetic processes rather than kinases, as is the case with mammalian Hsp90 [18,19].
Despite the essentiality of Hsp90s in most fungal pathogens, several studies have been able to interrogate their contributions to pathogenesis using chemical inhibitors or strains with regulated Hsp90 expression. Early work using chemical inhibition established the importance of Hsp90 in the ability of fungi to gain resistance against antifungal drugs [20]. As expected for an essential protein, it was later shown that Hsp90 inhibition impairs fungal virulence [21] and influences the heat shock response through the Hsf1 transcription factor required for thermal adaptation after heat shock at 42 °C [22]. Additionally, combination therapy with anti-Hsp90 drugs and antifungal drugs of the azole class has broad therapeutic potential against multiple fungal pathogens [21,23]. In C. albicans, Hsp90 plays important roles in temperature-dependent morphological changes at 37 °C as well as in contributing to tolerance against the echinocandins [24,25]. Chemical genetic screens conducted under different conditions established the plasticity of the Hsp90 network in response to environmental stresses. These genetic screens also identified regulatory proteins which broadly interacted with Hsp90 under different conditions [26]. The physical interactions of Hsp90 and several of its co-chaperones have also been characterized in C. albicans revealing similarities with the interacting partners for S. cerevisiae Hsp90 under routine conditions, including clients participating in signal transduction. However, during exposure to antifungals, Hsp90 stabilizes P-bodies and stress granules thereby providing novel insights into the mechanisms by which Hsp90 promotes antifungal drug tolerance [27]. The roles of Hsp90 in C. albicans virulence have been reviewed in more detail elsewhere [28]. In A. fumigatus, Hsp90 also plays important roles in morphological transitions and has been suggested as a potential antifungal drug target in this context [29,30]. Furthermore, Hsp90 expression is upregulated upon cell wall stress in A. fumigatus and the kinases governing the cell wall integrity pathway such as MpkA and PkcA are clients of Hsp90 [31]. In C. neoformans, chemical inhibition of Hsp90 revealed its importance in capsule elaboration, thermotolerance at both 37 and 39 °C, tolerance to antifungal drugs, and virulence in a Caenorhabditis elegans model [32,33]. Overall, the importance of Hsp90 to fungal pathogenesis is well established making it one of the most promising candidates within the chaperone network as a target for antifungal therapies.
2.2. Hsp70s
Hsp70s have a “DnaK” domain, named after the dnaK gene first identified as necessary for bacteriophage λ DNA replication in E. coli [34]. Hsp70s promote folding of proteins through a cycle of substrate binding facilitated by co-chaperones and nucleotide exchange factors. The nucleotide binding domain of Hsp70 binds ATP, and co-chaperone-mediated ATP hydrolysis induces a conformational change in the substrate binding domain (SBD) closing the lid and allowing tight binding of client substrates (Figure 2B). Nucleotide exchange factors induce ADP release and ATP binding, which reverts the conformational change in the SBD to the open low affinity conformation ultimately resulting in substrate release [35,36]. Aside from this role in protein folding, Hsp70s participate in seemingly unrelated processes such as DNA replication, clathrin disassembly, and protein translocation across membranes, however these processes all require modulation of either protein conformation or protein-protein interactions [7,8,13]. Information from S. cerevisiae provides useful context for appreciating the roles of Hsp70 proteins. In this fungus, there are fourteen Hsp70 and Hsp70-like proteins in seven subfamilies including four typical subfamilies SSA, SSB, SSC, and KAR, and three atypical subfamilies, two of which function as nucleotide exchange factors (LHS, SSE) and one as a ribosome assembly protein (SSZ) [13,35,37]. There are also Hsp70 members in the endoplasmic reticulum (Kar2) and mitochondria (Ssc1), which are required for proteostasis within these organelles and are essential for viability [9,38]. Recent efforts have begun to elucidate the different roles of specific cytosolic Hsp70s. In S. cerevisiae, the Ssb1 and Ssb2 cytosolic Hsp70s associate with ribosomes and participate in the early folding of many nascent proteins [39]. Genetic studies also revealed that there are many genes encoding proteins that uniquely interact with each of the Hsp70s in the SSA subfamily [40]. Protein interaction studies show that the constitutive Hsp70s Ssa1 and Ssa2 interact with many more proteins than the inducible Ssa3 and Ssa4 proteins. However, this is based on information publicly available in the Saccharomyces genome database and it is likely that the full interactomes of Ssa3 and Ssa4 have not been characterized because the appropriate stress conditions have not been thoroughly tested [40]. Although the roles of Hsp70s have consistently been shown to relate to protein folding, the differences in interaction networks of cytosolic Hsp70s highlights their underappreciated specificity.
In the context of fungal pathogens, the Hsp70s have high levels of expression at elevated temperatures and also influence morphological transitions [41,42,43]. Hsp70s are found localized to the cell surfaces of C. albicans [44], C. neoformans [45], and A. fumigatus [46,47]. Accordingly, Hsp70s are immunogenic proteins in systemic infections caused by C. neoformans [48,49] or C. albicans [50]. The cytoplasmic Hsp70, Ssa1, contributes to virulence in C. albicans specifically through facilitating host cell invasion [51]. Furthermore, Ssa1 phosphorylation plays a role in regulating morphology and is required for thermotolerance at 42 °C in C. albicans [52]. In C. neoformans, Ssa1 contributes to melanin production in one serotype [53] and is required to promote non-protective M2 macrophage polarization in lung monocytes upon infection with a different serotype [54]; however, in both backgrounds, mutants lacking SSA1 had reduced virulence in murine models of infection. Some Hsp70s are essential in fungal pathogens which makes it difficult to study their roles in virulence using classic reverse genetics approaches. However, the use of strains with regulated expression of the genes encoding these Hsp70s has revealed that their roles extend beyond known essential functions. For example, the Hsp70 protein Msi3 contributes to fluconazole resistance and virulence in C. albicans [55]. Similarly, the essential endoplasmic reticulum (ER) Hsp70, Kar2, is also crucial for thermotolerance at 37 °C as well as resistance against cell wall stress and azoles in C. neoformans [56]. Further investigations of the other members of the Hsp70 family and their roles in these human fungal pathogens may uncover their contributions to virulence.
2.3. Hsp40s/JDPs
The Hsp40s are co-chaperones necessary for stimulating the ATPase activity of Hsp70s and are thereby required for the Hsp70 cycle of client protein tight binding and release [57,58,59]. The Hsp40s are also called J domain proteins (JDPs) as they have a conserved ~70 amino acid J domain which is required for the stimulation of Hsp70 ATPase [59,60]. This name also comes from the DnaJ protein in E. coli which is encoded by a gene located next to dnaK and that is also required for bacteriophage λ DNA replication [61,62]. JDPs are classified into three categories based on the presence of conserved regions in addition to their J domains: type I JDPs have glycine/phenylalanine (G/F) rich domains and zinc binding motifs, type II JDPs have G/F rich domains, and type III JDPs only have a J domain [58,63,64]. The type I and II JDPs are able to bind non-native proteins through their G/F rich regions allowing them to bring these substrates to Hsp70s [58]. Most organisms have more JDPs than Hsp70s [65]. The expansion of this family contributes to the ability of JDPs to direct the activity of Hsp70s to a wide range of substrates and to participate in diverse functions. In particular, some cytosolic and ER-resident JDPs in S. cerevisiae are considered generalists based on their ability to bind a wide range of substrates, whereas some are specific for a small number of substrates [58,65]. Knockouts of generalist JDPs, such as Ydj1, can be rescued by overexpression of other JDPs; however, knockouts of specialist JDPs, such as Cwc23, Sis1, Jjj1, and Jjj3 are not rescued by overexpression of other JDPs [66]. Specialist JDPs can be specific for a certain substrate or process, as illustrated by the activity of Swa2 in clathrin-mediated endocytosis or Jac1 in iron-sulfur cluster assembly. Specificity of JDPs is largely governed by regions outside of the J domain and can be provided through recognition of conserved motifs such as polyglutamine stretches, or based on their position on or near ribosomes, mitochondria membranes, or the ER [65,66]. The JDPs which provide specificity based on their position can recruit Hsp70s, increase their local concentration, and direct the ATPase activity of these chaperones towards certain processes [64]. Many type III JDPs are poorly studied even in S. cerevisiae and it is generally thought that they participate in diverse and unique functions.
JDPs continue to be actively studied because many of their functions are still unknown even in model organisms. In human fungal pathogens, there are examples of JDPs which contribute to virulence mainly through participating in specific processes. For example, the cytosolic JDP, Ydj1, contributes to the stress response, morphogenesis, and mitochondrial function of C. albicans [67]. Furthermore, the mitochondrial JDP, Mrj1, contributes to mitochondrial respiration, capsule formation, and ultimately virulence in C. neoformans [68]. Surprisingly there are few other reports on the roles of JDPs in the virulence of human fungal pathogens despite their well characterized roles in fungal pathogens of insects [69,70] and plants [71,72,73,74,75]. Further study of JDPs, particularly the divergent type III JDPs, in the context of fungal pathogens may uncover novel and divergent proteins with roles in pathogenesis and also provide insights into the roles of their Hsp70 partners.
2.4. Hsp100s
The Hsp100/ClpB family is a group of hexameric AAA+ ATPase chaperones (Figure 2C) conserved in bacteria, yeasts, and plants but absent in metazoans [76,77]. These proteins generally function as disaggregases by pulling and processing protein strands from their client proteins through a central channel and unfolding them [77]. Paradoxically, they also increase prion propagation, likely by pulling polypeptide strands through their central pore and producing seeds for prion propagation [78]. In S. cerevisiae, the AAA+ ATPase Hsp104 is not essential for viability, but it is important for the acquisition of thermotolerance [79]. Importantly, members of the Hsp100/ClpB family also associate with other proteins such as proteases or other chaperone machinery [80]. This class of heat shock proteins also interacts with the Hsp70/JDP machinery, as discussed later. Therefore, it links together other aspects of the HSP machinery to coordinately mitigate proteotoxic stress.
In the context of human fungal pathogens, relatively little is known about the roles of Hsp100/ClpB in pathogenesis. However, in C. albicans Hsp104 contributes to biofilm formation as well as virulence in a worm model of infection [81]. Furthermore, the sumoylation of Hsp104 contributes to the thermotolerance to heat shock at 42 °C of C. albicans [82]. Although the roles of these chaperones have not been characterized in other fungi, Hsp104 is required for the acquisition of thermotolerance in S. cerevisiae [79] suggesting that it could potentially contribute to fungal adaptation to a mammalian host.
2.5. Chaperonins
Chaperonins are oligomeric protein folding complexes that have two stacked rings which allow unfolded proteins to enter the lumen. When an unfolded protein is interacting with the chamber, the chaperonin binds ATP and adopts a closed conformation creating a protected environment for proteins to fold (Figure 2D). Upon ATP hydrolysis, the chaperonins open and release the folded protein from the lumen [83]. The chaperonins are divided into two distinct groups. The group I chaperonins include the Hsp60s and GroEL, which are found in prokaryotes as well as in eukaryotic organelles such as mitochondria. The group II chaperonins, CCT/TRiC, are found in archaea and in the eukaryotic cytosol [84,85]. The mitochondrial chaperonin GroEL is involved in the folding of many mitochondrial proteins, especially those with a size less than 60 kDa and with regions of exposed hydrophobic β-sheets [86]. Early reports described the eukaryotic CCT chaperonin as being involved in the folding of actin and tubulin [84,87]. More recently, the roles of this chaperonin have been characterized in S. cerevisiae using proteomic and structural approaches. This analysis revealed that CCT participates in folding of proteins related to the nuclear pore complex, chromatin remodeling, protein degradation, the anaphase promoting complex, and the mTOR complex [88,89]. The CCT chaperonins are not as abundant as other chaperones, but they facilitate the folding of as many as 15% of all newly synthesized proteins [90].
The chaperonins play roles in morphogenesis and virulence in C. albicans. In particular, a CCT complex protein, Cct8, suppresses hyphal formation in C. albicans [91]. The CCT complex is also important for echinocandin resistance in C. albicans through its role in chaperoning actin dynamics, promoting correct septin localization, and maintaining cell wall architecture [92]. Sumoylation of Hsp60 is also required for regulation of hyphal growth in C. albicans and thermotolerance to heat shock at 42 °C when mitochondrial respiration is inhibited with antimycin A [82]. In addition to these studies, Hsp60 is immunogenic and has been proposed as a candidate vaccine target to protect against other human fungal pathogens including Histoplasma capsulatum and Paracoccidioides brasiliensis [93,94].
2.6. Small HSPs
Small HSPs (sHsps) are passive, energy-independent chaperones which oligomerize in part through their conserved α-crystallin domains. The monomeric proteins range in size from 12 to 40kDa and are often named accordingly (e.g., Hsp12) [95]. The sHsps demonstrate binding capacity for denatured substrates and they often act as holdases immobilizing denatured proteins until an ATP-dependent chaperone such as Hsp70 can bind and reactivate them [96]. In S. cerevisiae, Hsp12 is a well-studied sHsp whose expression increases with a variety of stresses including exposure to NaCl or ethanol [97]. Importantly, Hsp12 also plays a role in the plasticity and flexibility of the yeast cell wall [98].
The sHsps also contribute to fungal pathogenesis. For example, the sHsp Hsp21 is required for growth at elevated temperatures between 39 and 42 °C and ultimately virulence in C. albicans [99]. Hsp21 is also required for resistance against the antifungal drugs caspofungin and the imidazoles [100]. Another sHsp, Hsp12, is upregulated upon stresses including heat shock at 45 °C, oxidative stress, and osmotic stress, although it is not required for virulence in C. albicans [101]. In C. neoformans, the sHsps Hsp12 and Hsp122 contribute to resistance against amphotericin B [102]. Overall, these data suggest that sHsps may broadly be important for resistance against antifungal drugs. However, since the sHsps are diverse, the sHsps that are as yet uncharacterized may have novel contributions in the human fungal pathogens.
3. Coordination of Chaperoning Activity across Different Families of HSPs
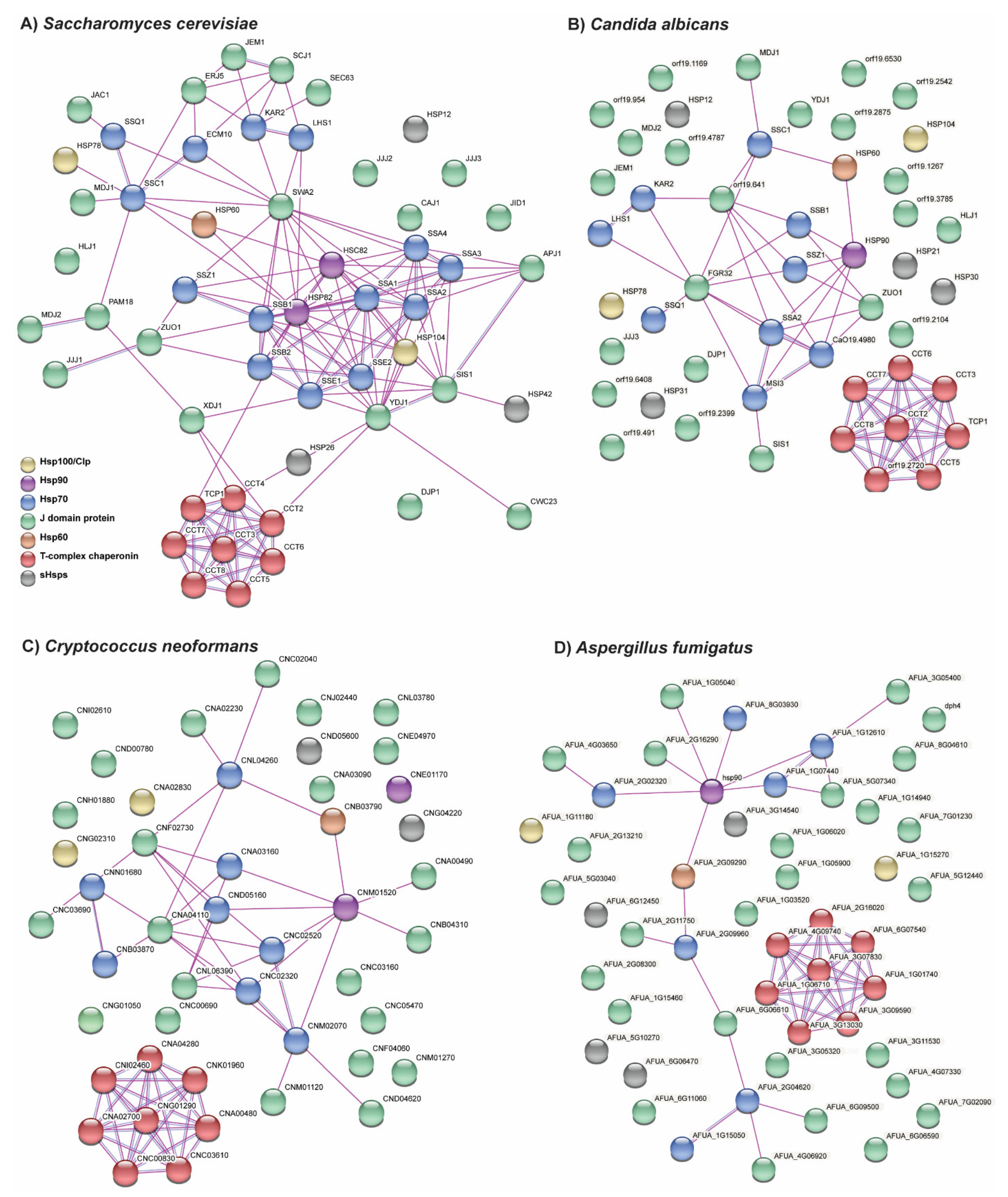
The families of heat shock proteins reviewed here do not act in isolation. It has already been discussed how the JDPs are co-chaperones necessary for the ATPase activity of the Hsp70s. However, many of the HSP families act within chaperone networks to facilitate proper protein folding, complex assembly, and protein degradation. Within the Kingdom Fungi, the connections between HSPs are best characterized in S. cerevisiae, however many of these connections are not characterized in other fungi including the major human fungal pathogens (Figure 3A–D). In the STRING database, many of the connections between HSPs in the fungal pathogens are inferred from interactions between orthologs encoding HSPs in S. cerevisiae. Therefore, the interactions between the divergent proteins in these fungal pathogens largely remain unknown. In particular, many of the proteins without connections in the chaperone networks of human fungal pathogens were JDPs or sHSPs (Figure 3B–D). This suggests that there are divergent proteins in these classes of HSPs which may have specific functions related to pathogenesis and which warrant further research to assess their roles and potential as drug targets.
Perhaps the best characterized interactions between chaperones are between the Hsp70s and Hsp90s. In S. cerevisiae, Hsp90 and Hsp70 orthologs (Hsp82 and Ssa1) interact with each other mediated by the tetratricopeptide repeat protein Sti1 and this interaction is enhanced by cytosolic JDPs [15,103,104,105]. In addition to its role in coordinating their interaction, Sti1 facilitates substrate transfer from Hsp70 to Hsp90 [106]. Since Hsp90s are so abundant and participate in many pathways, efforts have been made to characterize their co-chaperones through establishing their genetic interaction network [107]. The Hsp90 co-chaperones generally participate in different processes as suggested by a lack of synthetic lethality between co-chaperone knockouts [107]. Characterization of the physical interactions of Hsp90s in S. cerevisiae also confirmed the interactions between Hsp90s and other chaperones and co-chaperones as well as the interactions with ligand binding biosynthetic enzymes [16,18,108].
The Hsp70 chaperone network is extensive and has been reviewed elsewhere [35] with particular focus on the roles of the JDPs in directing Hsp70 activity towards specific functions [65]. The JDPs are structurally diverse outside the J domain and often participate in discrete functions by directing the activity of Hsp70s through binding Hsp70 substrates, recruiting Hsp70s to a particular subcellular compartment, or modulating the Hsp70-substrate interaction [109]. The Hsp70s and JDPs also transfer substrates to other components of the chaperone machinery including the disaggregases and chaperonins [35,65,110]. The disaggregation of proteins in vitro is enhanced when both a type I and type II JDP such as Ydj1 and Sis1 are added to purified Hsp104, Ssa1, Sse1, and aggregated substrate compared to either JDP alone [111]. The Hsp70s and JDPs also transfer non-native substrates to the chamber-type chaperonins (mitochondrial GroEL and cytosolic T-complex chaperonin) where they can fold in a protected environment [35,65].
The chaperone network also connects the HSPs to other proteins and pathways relevant to protein homeostasis, most notably functions for protein degradation. The JDPs can direct substrates to particular fates by tethering chaperone machinery to specific subcellular compartments or to other proteins such as ubiquitin ligases for subsequent degradation [65]. The Sse subfamily of Hsp70s act as nucleotide exchange factors for other Hsp70s and facilitate the transfer of substrates from Hsp70 to the proteasome for degradation in S. cerevisiae [112]. Another class of chaperone, the T complex cytosolic chaperonin, also has several genetic interactions with protein degradation networks including proteins involved in ubiquitination and sumoylation [88]. These data for S. cerevisiae fit well with the interactome studies in mammalian cells which show that Hsp90 co-chaperone networks are extensive and connect to protein degradation through Hsp90′s interactions with ~31% of all E3 ligases [19,113]. The connectivity of the HSP network suggests that disruption of specific interactions can fine tune the chaperone network to modulate the suite of clients or chaperone activities while retaining other essential functions.
In the fungal pathogens of humans, the best characterized chaperone network is focused on Hsp90 and its co-chaperones in C. albicans. The genetic interaction network of Hsp90 in C. albicans under different conditions indicated that Hsp90 interacts with different proteins depending on the environmental stress conditions. However, two proteins, the kinase CK2, and the transcription factor Ahr1, interacted with Hsp90 in five of the six conditions tested. Indeed, CK2 and Ahr1 regulate the phosphorylation of Hsp90 and the transcription of the gene, respectively. Interestingly only ~ 17% of the genetic interaction network in C. albicans was shared with S. cerevisiae [26]. Physical interactions between chaperones in C. albicans also revealed that Hsp90 interacts with many other chaperones including Hsp70s and the T-complex chaperonins. However, this interactome characterization also revealed novel interactions with Hsp90 that had not been characterized in S. cerevisiae such as a connection with the osmotic stress responsive kinase Pbs2 [27]. Interactions between Hsp90 and Hsp70 via the StiA co-chaperone have also been demonstrated in A. fumigatus to coordinately promote tolerance to the antifungal drug caspofungin [114]. Together, these studies show both conserved and novel elements of the Hsp90 network in the human fungal pathogens. The novel elements discovered in C. albicans highlight the importance of characterizing the interactions between chaperones in the context of fungal pathogens. Similar studies in the other pathogenic fungi discussed in this review would likely uncover more novel elements of the chaperone network as both C. albicans and S. cerevisiae are within the class Saccharomycetes, whereas A. fumigatus is in a different class, Eurotiomycetes, and C. neoformans is even more distantly related as a basidiomycete. Furthermore, the pathogenic mechanisms of these fungi differ and hence the context-dependent interactions may also be novel in these fungal pathogens.
4. Heat Shock Proteins as Drug Targets for Fungal Pathogens
The importance of HSPs to virulence factor production in fungal pathogens as outlined above makes them attractive as potential targets for antifungal drug development [115]. Drugs which impair the functions of Hsp90 such as radicicol and geldanamycin have been used to study the functions and genetic interactions of Hsp90s in the context of human fungal pathogens [21,26,32,33]. Derivatives of geldanamycin such as 17-N-allylamino-17-demethoxygeldanamycin (17-AAG) which are less toxic to mammalian hosts have also shown promising antifungal activity [21,33,116]. Recently, fungal selective inhibitors of Hsp90 were developed which have activity against both C. neoformans and C. albicans Hsp90s [117,118]. The development of these inhibitors, which was guided by structural differences in the nucleotide binding domains between human and fungal Hsp90s complexed with inhibitors, allowed the generation of inhibitors with lower toxicity to human cells and therefore greater promise for clinical use [117,118].
Another approach to targeting the chaperone network is through disruption of protein-protein interactions between chaperones using small molecules. The impact of this approach differs from direct inhibition of HSP functions as their activities can be fine-tuned by disrupting interactions with a specific subset of co-chaperones or clients [119,120]. This approach has been discussed in the context of anti-cancer strategies although similar approaches could be used for antifungal drug development. In particular, this approach may allow targeting of the Hsp70 and co-chaperone network. Although, to our knowledge, these approaches have not been used or optimized for fungal pathogens, small molecules modulating the Hsp70/JDP interaction such as the dihydropyrimidines influence the phenotypes of a ydj1∆ in S. cerevisiae suggesting that they are bioavailable to fungi [121].
Thus far, many of the HSP inhibitors in use or in development target the well-conserved Hsp90s. The ability to make fungal selective inhibitors of Hsp90s is critical to ensuring that they can be used safely without off target effects. Studies characterizing chaperones contributing to virulence in fungal pathogens have identified divergent components of the chaperone network in fungi such as the sHSPs [99] and JDPs [68]. This information may be exploited to prioritize these divergent targets for antifungal drug development. Finally, there are still many uncharacterized chaperones and co-chaperones in the fungal pathogens of humans which are divergent from characterized chaperones in S. cerevisiae. Further characterization of these divergent proteins may identify novel targets for antifungal drug development in these pathogens.
5. Conclusions
The HSPs generally allow organisms to maintain proteostasis and survive despite exposure to high temperatures and other environmental stresses. The ability to survive elevated temperatures is one of the major natural factors limiting fungi from surviving and proliferating in human hosts. In addition to the general role of facilitating growth at human body temperature, the HSPs in fungal pathogens play important roles in promoting morphological changes associated with pathogenesis, resistance against antifungal drugs, and virulence in several infection models. Currently, inhibitors of the Hsp90s, which are crucial for fungal virulence, are being developed with improved selectivity for fungal proteins. As outlined in this review, almost every major class of HSPs has representatives which contribute to fungal pathogenesis. This includes HSPs which lack well-conserved proteins in humans. Therefore, there may be underexploited chaperone targets, and inhibition of these HSPs or disruption of the protein–protein interactions within the HSP networks are promising strategies to develop novel antifungal drugs and therapeutics.
Author Contributions
Conceptualization, L.C.H.; writing—original draft preparation, L.C.H. and J.W.K.; writing—review and editing, L.C.H. and J.W.K.; funding acquisition, J.W.K. All authors have read and agreed to the published version of the manuscript.
Funding
Research in our laboratory on HSPs is supported by grants (MOP-13234 and PJT-166043) from the Canadian Institutes of Health Research (to JWK), and by a doctoral scholarship from the National Sciences and Engineering Research Council of Canada (to LCH).
Acknowledgments
The authors apologize to those researchers whose work we were unable to cite. JWK is a Burroughs Wellcome Fund Scholar in Molecular Pathogenic Mycology, and a fellow of the CIFAR program: Fungal Kingdom Threats & Opportunities.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Perfect, J.R. Cryptococcus neoformans: The yeast that likes it hot. Fems Yeast Res. 2006, 6, 463–468. [Google Scholar] [CrossRef] [Green Version]
- Robert, V.A.; Casadevall, A. Vertebrate Endothermy Restricts Most Fungi as Potential Pathogens. J. Infect. Dis. 2009, 200, 1623–1626. [Google Scholar] [CrossRef]
- Casadevall, A. Fungi and the Rise of Mammals. PLoS Pathog. 2012, 8, e1002808. [Google Scholar] [CrossRef]
- Gauthier, G.M. Dimorphism in Fungal Pathogens of Mammals, Plants, and Insects. PLoS Pathog. 2015, 11, e1004608. [Google Scholar] [CrossRef] [PubMed]
- De Maio, A. Heat shock proteins: Facts, thoughts, and dreams. Shock 1999, 11, 1–12. [Google Scholar] [CrossRef]
- Verghese, J.; Abrams, J.; Wang, Y.; Morano, K.A. Biology of the Heat Shock Response and Protein Chaperones: Budding Yeast (Saccharomyces cerevisiae) as a Model System. Microbiol. Mol. Biol. Rev. 2012, 76, 115–158. [Google Scholar] [CrossRef] [Green Version]
- Hendrick, J.; Hartl, F.U. Molecular Chaperone Functions of Heat-Shock Proteins. Annu. Rev. Biochem. 1993, 62, 349–384. [Google Scholar] [CrossRef]
- Parsell, D.; Lindquist, S. The Function of Heat-Shock Proteins in Stress Tolerance: Degradation and Reactivation of Damaged Proteins. Annu. Rev. Genet. 1993, 27, 437–496. [Google Scholar] [CrossRef] [PubMed]
- Georgopoulos, C.; Welch, W.J. Role of the major heat shock proteins as molecular chaperones. Annu. Rev. Cell Biol. 1993, 9, 601–634. [Google Scholar] [CrossRef] [PubMed]
- Wandinger, S.K.; Richter, K.; Buchner, J. The Hsp90 chaperone machinery. J. Biol. Chem. 2008, 283, 18473–18477. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Houry, W.A. Molecular interaction network of the Hsp90 chaperone system. In Molecular Aspects of the Stress Response: Chaperones, Membranes and Networks; Springer: New York, NY, USA, 2007; Volume 594, pp. 27–36. ISBN 9780387399744. [Google Scholar]
- Schopf, F.H.; Biebl, M.M.; Buchner, J. The HSP90 chaperone machinery. Nat. Rev. Mol. Cell Biol. 2017, 18, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, S.; Craig, E.A. THE HEAT-SHOCK PROTEINS. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, M.J. Heat Shock Proteins. J. Biol. Chem. 1990, 265, 12111–12114. [Google Scholar] [CrossRef]
- Chang, H.C.J.; Lindquist, S. Conservation of Hsp90 macromolecular complexes in Saccharomyces cerevisiae. J. Biol. Chem. 1994, 269, 24983–24988. [Google Scholar] [CrossRef]
- Zhao, R.; Davey, M.; Hsu, Y.C.; Kaplanek, P.; Tong, A.; Parsons, A.B.; Krogan, N.; Cagney, G.; Mai, D.; Greenblatt, J.; et al. Navigating the chaperone network: An integrative map of physical and genetic interactions mediated by the hsp90 chaperone. Cell 2005, 120, 715–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClellan, A.J.; Xia, Y.; Deutschbauer, A.M.; Davis, R.W.; Gerstein, M.; Frydman, J. Diverse Cellular Functions of the Hsp90 Molecular Chaperone Uncovered Using Systems Approaches. Cell 2007, 131, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Girstmair, H.; Tippel, F.; Lopez, A.; Tych, K.; Stein, F.; Haberkant, P.; Schmid, P.W.N.; Helm, D.; Rief, M.; Sattler, M.; et al. The Hsp90 isoforms from S. cerevisiae differ in structure, function and client range. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Taipale, M.; Krykbaeva, I.; Koeva, M.; Kayatekin, C.; Westover, K.D.; Karras, G.I.; Lindquist, S. Quantitative analysis of Hsp90-client interactions reveals principles of substrate recognition. Cell 2012, 150, 987–1001. [Google Scholar] [CrossRef] [Green Version]
- Cowen, L.E.; Lindquist, S. Hsp90 potentiates the rapid evolution of new traits: Drug resistance in diverse fungi. Science 2005, 309, 2185–2189. [Google Scholar] [CrossRef]
- Cowen, L.E.; Singh, S.D.; Köhler, J.R.; Collins, C.; Zaas, A.K.; Schell, W.A.; Aziz, H.; Mylonakis, E.; Perfect, J.R.; Whitesell, L.; et al. Harnessing Hsp90 function as a powerful, broadly effective therapeutic strategy for fungal infectious disease. Proc. Natl. Acad. Sci. USA 2009, 106, 2818–2823. [Google Scholar] [CrossRef] [Green Version]
- Leach, M.D.; Budge, S.; Walker, L.; Munro, C.; Cowen, L.E.; Brown, A.J.P. Hsp90 Orchestrates Transcriptional Regulation by Hsf1 and Cell Wall Remodelling by MAPK Signalling during Thermal Adaptation in a Pathogenic Yeast. PLoS Pathog. 2012, 8, 1003069. [Google Scholar] [CrossRef] [Green Version]
- Cowen, L.E. The fungal Achilles’ heel: Targeting Hsp90 to cripple fungal pathogens. Curr. Opin. Microbiol. 2013, 16, 377–384. [Google Scholar] [CrossRef]
- Singh, S.D.; Robbins, N.; Zaas, A.K.; Schell, W.A.; Perfect, J.R.; Cowen, L.E. Hsp90 governs echinocandin resistance in the pathogenic yeast Candida albicans via calcineurin. PLoS Pathog. 2009, 5, e1000532. [Google Scholar] [CrossRef]
- Shapiro, R.S.; Uppuluri, P.; Zaas, A.K.; Collins, C.; Senn, H.; Perfect, J.R.; Heitman, J.; Cowen, L.E. Hsp90 Orchestrates Temperature-Dependent Candida albicans Morphogenesis via Ras1-PKA Signaling. Curr. Biol. 2009, 19, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Diezmann, S.; Michaut, M.; Shapiro, R.S.; Bader, G.D.; Cowen, L.E. Mapping the Hsp90 Genetic Interaction Network in Candida albicans Reveals Environmental Contingency and Rewired Circuitry. PLoS Genet. 2012, 8, 1002562. [Google Scholar] [CrossRef] [Green Version]
- O’Meara, T.R.; O’Meara, M.J.; Polvi, E.J.; Pourhaghighi, M.R.; Liston, S.D.; Lin, Z.Y.; Veri, A.O.; Emili, A.; Gingras, A.C.; Cowen, L.E. Global proteomic analyses define an environmentally contingent Hsp90 interactome and reveal chaperone-dependent regulation of stress granule proteins and the R2TP complex in a fungal pathogen. PLoS Biol. 2019, 17, e3000358. [Google Scholar] [CrossRef]
- O’Meara, T.R.; Robbins, N.; Cowen, L.E. The Hsp90 Chaperone Network Modulates Candida Virulence Traits. Trends Microbiol. 2017, 25, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Lamoth, F.; Juvvadi, P.R.; Fortwendel, J.R.; Steinbach, W.J. Heat shock protein 90 is required for conidiation and cell wall integrity in Aspergillus fumigatus. Eukaryot. Cell 2012, 11, 1324–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamoth, F.; Juvvadi, P.R.; Steinbach, W.J. Heat shock protein 90 (Hsp90): A novel antifungal target against Aspergillus fumigatus. Crit. Rev. Microbiol. 2014, 42, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.C.; Minari, K.; Fabri, J.H.T.M.; Kerkaert, J.D.; Gava, L.M.; da Cunha, A.F.; Cramer, R.A.; Borges, J.C.; Malavazi, I. Aspergillus fumigatus Hsp90 interacts with the main components of the cell wall integrity pathway and cooperates in heat shock and cell wall stress adaptation. Cell. Microbiol. 2020, e13273. [Google Scholar] [CrossRef]
- De Cordeiro, R.A.; de Evangelista, A.J.J.; Serpa, R.; de Farias Marques, F.J.; de Melo, C.V.S.; de Oliveira, J.S.; da Silva Franco, J.; de Alencar, L.P.; de Jesus Pinheiro Gomes Bandeira, T.; Brilhante, R.S.N.; et al. Inhibition of heat-shock protein 90 enhances the susceptibility to antifungals and reduces the virulence of Cryptococcus neoformans/Cryptococcus gattii species complex. Microbiology 2016, 162, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Tatu, U. Heat shock protein 90 localizes to the surface and augments virulence factors of Cryptococcus neoformans. PLoS Negl. Trop. Dis. 2017, 11, e0005836. [Google Scholar] [CrossRef] [Green Version]
- Saito, H.; Uchida, H. Initiation of the DNA replication of bacteriophage lambda in Escherichia coli K12. J. Mol. Biol. 1977, 113, 1–25. [Google Scholar] [CrossRef]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef]
- Liu, Q.; Liang, C.; Zhou, L. Structural and functional analysis of the Hsp70/Hsp40 chaperone system. Protein Sci. 2020, 29, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Kominek, J.; Marszalek, J.; Neuvéglise, C.; Craig, E.A.; Williams, B.L. The complex evolutionary dynamics of Hsp70s: A genomic and functional perspective. Genome Biol. Evol. 2013, 5, 2460–2477. [Google Scholar] [CrossRef] [Green Version]
- Estruch, F. Stress-controlled transcription factors, stress-induced genes and stress tolerance in budding yeast. Fems Microbiol. Rev. 2000, 24, 469–486. [Google Scholar] [CrossRef] [PubMed]
- Döring, K.; Ahmed, N.; Riemer, T.; Suresh, H.G.; Vainshtein, Y.; Habich, M.; Riemer, J.; Mayer, M.P.; O’Brien, E.P.; Kramer, G.; et al. Profiling Ssb-Nascent Chain Interactions Reveals Principles of Hsp70-Assisted Folding. Cell 2017, 170, 298–311.e20. [Google Scholar] [CrossRef] [Green Version]
- Lotz, S.K.; Knighton, L.E.; Nitika; Jones, G.W.; Truman, A.W. Not quite the SSAme: Unique roles for the yeast cytosolic Hsp70s. Curr. Genet. 2019, 65, 1127–1134. [Google Scholar] [CrossRef]
- Gómez, B.L.; Porta, A.; Maresca, B. Heat Shock Response in Pathogenic Fungi. In Human Fungal Pathogens; Springer: Berlin/Heidelberg, Germany, 2004; pp. 113–132. [Google Scholar]
- Cleare, L.G.; Zamith-Miranda, D.; Nosanchuk, J.D. Heat shock proteins in Histoplasma and Paracoccidioides. Clin. Vaccine Immunol. 2017, 24, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, S.; Shankar, J. Hsp70 in Fungi: Evolution, Function and Vaccine Candidate. In HSP70 in Human Diseases and Disorders; Springer: Cham, Switzerland, 2018; pp. 381–400. [Google Scholar]
- Eroles, P.; Sentandreu, M.; Elorza, M.V.; Sentandreu, R. The highly immunogenic enolase and Hsp70p are adventitious Candida albicans cell wall proteins. Microbiology 1997, 143, 313–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira, C.P.; Piffer, A.C.; Kmetzsch, L.; Fonseca, F.L.; Soares, D.A.; Staats, C.C.; Rodrigues, M.L.; Schrank, A.; Vainstein, M.H. The heat shock protein (Hsp) 70 of Cryptococcus neoformans is associated with the fungal cell surface and influences the interaction between yeast and host cells. Fungal Genet. Biol. 2013, 60, 53–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubitschek-Barreira, P.H.; Curty, N.; Neves, G.W.P.; Gil, C.; Lopes-Bezerra, L.M. Differential proteomic analysis of Aspergillus fumigatus morphotypes reveals putative drug targets. J. Proteomics 2013. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.-J.; Krüger, T.; Blango, M.G.; von Eggeling, F.; Kniemeyer, O.; Brakhage, A.A. Biotinylated Surfome Profiling Identifies Potential Biomarkers for Diagnosis and Therapy of Aspergillus fumigatus Infection. mSphere 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Kakeya, H.; Udono, H.; Ikuno, N.; Yamamoto, Y.; Mitsutake, K.; Miyazaki, T.; Tomono, K.; Koga, H.; Tashiro, T.; Nakayama, E.; et al. A 77-kilodalton protein of Cryptococcus neoformans, a member of the heat shock protein 70 family, is a major antigen detected in the sera of mice with pulmonary cryptococcosis. Infect. Immun. 1997, 65, 1653–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakeya, H.; Udono, H.; Maesaki, S.; Sasaki, E.; Kawamura, S.; Hossain, M.A.; Yamamoto, Y.; Sawai, T.; Fukuda, M.; Mitsutake, K.; et al. Heat shock protein 70 (hsp70) as a major target of the antibody response in patients with pulmonary cryptococcosis. Clin. Exp. Immunol. 1999, 115, 485–490. [Google Scholar] [CrossRef]
- Bromuro, C.; La Valle, R.; Sandini, S.; Urbani, F.; Ausiello, C.M.; Morelli, L.; D’Ostiani, C.F.; Romani, L.; Cassone, A. A 70-kilodalton recombinant heat shock protein of Candida albicans is highly immunogenic and enhances systemic murine candidiasis. Infect. Immun. 1998, 66, 2154–2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.N.; Solis, N.V.; Phan, Q.T.; Bajwa, J.S.; Kashleva, H.; Thompson, A.; Liu, Y.; Dongari-Bagtzoglou, A.; Edgerton, M.; Filler, S.G. Host Cell Invasion and Virulence Mediated by Candida albicans Ssa1. PLoS Pathog. 2010, 6, e1001181. [Google Scholar] [CrossRef] [Green Version]
- Weissman, Z.; Pinsky, M.; Wolfgeher, D.J.; Kron, S.J.; Truman, A.W.; Kornitzer, D. Genetic analysis of Hsp70 phosphorylation sites reveals a role in Candida albicans cell and colony morphogenesis. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140135. [Google Scholar] [CrossRef]
- Zhang, S.; Hacham, M.; Panepinto, J.; Hu, G.; Shin, S.; Zhu, X.; Williamson, P.R. The Hsp70 member, Ssa1, acts as a DNA-binding transcriptional co-activator of laccase in Cryptococcus Neoformans. Mol. Microbiol. 2006, 62, 1090–1101. [Google Scholar] [CrossRef]
- Eastman, A.J.; He, X.; Qiu, Y.; Davis, M.J.; Vedula, P.; Lyons, D.M.; Park, Y.-D.; Hardison, S.E.; Malachowski, A.N.; Osterholzer, J.J.; et al. Cryptococcal Heat Shock Protein 70 Homolog Ssa1 Contributes to Pulmonary Expansion of Cryptococcus neoformans during the Afferent Phase of the Immune Response by Promoting Macrophage M2 Polarization. J. Immunol. 2015, 194, 5999–6010. [Google Scholar] [CrossRef]
- Nagao, J.I.; Cho, T.; Uno, J.; Ueno, K.; Imayoshi, R.; Nakayama, H.; Chibana, H.; Kaminishi, H. Candida albicans Msi3p, a homolog of the Saccharomyces cerevisiae Sse1p of the Hsp70 family, is involved in cell growth and fluconazole tolerance. Fems Yeast Res. 2012, 12, 728–737. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.-W.; Kang, H.A.; Bahn, Y.-S. Essential Roles of the Kar2/BiP Molecular Chaperone Downstream of the UPR Pathway in Cryptococcus neoformans. PLoS ONE 2013, 8, 58956. [Google Scholar] [CrossRef] [Green Version]
- Hartl, F.U. Molecular chaperones in cellular protein folding. Nature 1996, 381, 571–580. [Google Scholar] [CrossRef]
- Walsh, P.; Bursać, D.; Law, Y.C.; Cyr, D.; Lithgow, T. The J-protein family: Modulating protein assembly, disassembly and translocation. Embo Rep. 2004, 5, 567–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennessy, F.; Nicoll, W.S.; Zimmermann, R.; Cheetham, M.E.; Blatch, G.L. Not all J domains are created equal: Implications for the specificity of Hsp40-Hsp70 interactions. Protein Sci. 2005, 14, 1697–1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampinga, H.H.; Andreasson, C.; Barducci, A.; Cheetham, M.E.; Cyr, D.; Emanuelsson, C.; Genevaux, P.; Gestwicki, J.E.; Goloubinoff, P.; Huerta-Cepas, J.; et al. Function, evolution, and structure of J-domain proteins. Cell Stress Chaperones 2019, 24, 7–15. [Google Scholar] [CrossRef]
- Saito, H.; Uchida, H. Organization and expression of the dnaJ and dnaK genes of Escherichia coli K12. MGG Mol. Gen. Genet. 1978, 164, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yochem, J.; Uchida, H.; Sunshine, M.; Saito, H.; Georgopoulos, C.P.; Feiss, M. Genetic analysis of two genes, dnaJ and dnaK, necessary for Escherichia coli and bacteriophage lambda DNA replication. MGG Mol. Gen. Genet. 1978, 164, 9–14. [Google Scholar] [CrossRef]
- Qiu, X.B.; Shao, Y.M.; Miao, S.; Wang, L. The diversity of the DnaJ/Hsp40 family, the crucial partners for Hsp70 chaperones. Cell. Mol. Life Sci. 2006, 63, 2560–2570. [Google Scholar] [CrossRef] [PubMed]
- Musskopf, M.K.; de Mattos, E.P.; Bergink, S.; Kampinga, H.H. HSP40/DNAJ Chaperones. eLS 2018, 1–11. [Google Scholar] [CrossRef]
- Craig, E.A.; Marszalek, J. How Do J-Proteins Get Hsp70 to Do Therefore, Many Different Things? Trends Biochem. Sci. 2017, 42, 355–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahi, C.; Craig, E.A. Network of general and specialty J protein chaperones of the yeast cytosol. Proc. Natl. Acad. Sci. USA 2007, 104, 7163–7168. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.L.; Bohovych, I.; Wong, E.O.Y.; Lambert, J.-P.; Gingras, A.-C.; Khalimonchuk, O.; Cowen, L.E.; Leach, M.D. Ydj1 governs fungal morphogenesis and stress response, and facilitates mitochondrial protein import via Mas1 and Mas2. Microb. Cell 2017, 4, 342–361. [Google Scholar] [CrossRef] [PubMed]
- Horianopoulos, L.C.; Hu, G.; Caza, M.; Schmitt, K.; Overby, P.; Johnson, J.D.; Valerius, O.; Braus, G.H.; Kronstad, J.W. The novel J-domain protein Mrj1 is required for mitochondrial respiration and virulence in Cryptococcus neoformans. MBio 2020, 11. [Google Scholar] [CrossRef]
- Wang, J.; Ying, S.H.; Hu, Y.; Feng, M.G. Mas5, a homologue of bacterial DnaJ, is indispensable for the host infection and environmental adaptation of a filamentous fungal insect pathogen. Environ. Microbiol. 2016, 18, 1037–1047. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Ying, S.H.; Hu, Y.; Feng, M.G. Vital role for the J-domain protein Mdj1 in asexual development, multiple stress tolerance, and virulence of Beauveria bassiana. Appl. Microbiol. Biotechnol. 2017, 101, 185–195. [Google Scholar] [CrossRef]
- Son, Y.E.; Cho, H.J.; Chen, W.; Son, S.H.; Lee, M.K.; Yu, J.H.; Park, H.S. The role of the VosA-repressed dnjA gene in development and metabolism in Aspergillus species. Curr. Genet. 2020, 66, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Lo Presti, L.; López Díaz, C.; Turrà, D.; Di Pietro, A.; Hampel, M.; Heimel, K.; Kahmann, R. A conserved co-chaperone is required for virulence in fungal plant pathogens. New Phytol. 2016, 209, 1135–1148. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; Yang, J.; Shi, Y.; Wang, X.; Wang, G.-L. The DnaJ protein OsDjA6 negatively regulates rice innate immunity to the blast fungus Magnaporthe oryzae. Mol. Plant. Pathol. 2018, 19, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Yi, M.; Lee, Y.-H. Identification of genes encoding heat shock protein 40 family and the functional characterization of teo Hsp40s, MHF15 and MHF21, in Magnaporthe oryzae. Plant. Pathol. J. 2008, 24, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.-G.; Lee, J.-G.; Kim, J.-M.; Park, J.-A.; Park, S.-M.; Yang, M.-S.; Kim, D.-H. A DnaJ-like Homolog from Cryphonectria parasitica Is Not Responsive to Hypoviral Infection but Is Important for Fungal Growth in Both Wild-Type and Hypovirulent Strains. Mol. Cells 2010, 30, 235–243. [Google Scholar] [CrossRef]
- Mosser, D.D.; Ho, S.; Glover, J.R. Saccharomyces cerevisiae Hsp104 enhances the chaperone capacity of human cells and inhibits heat stress-induced proapoptotic signaling. Biochemistry 2004, 43, 8107–8115. [Google Scholar] [CrossRef]
- Grimminger-Marquardt, V.; Lashuel, H.A. Structure and function of the molecular chaperone Hsp104 from yeast. Biopolymers 2010, 93, 252–276. [Google Scholar] [CrossRef]
- Tessarz, P.; Mogk, A.; Bukau, B. Substrate threading through the central pore of the Hsp104 chaperone as a common mechanism for protein disaggregation and prion propagation. Mol. Microbiol. 2008, 68, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, Y.; Lindquist, S.L. HSP104 required for induced thermotolerance. Science (80-.) 1990, 248, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Heuck, A.; Schitter-Sollner, S.; Józef Suskiewicz, M.; Kurzbauer, R.; Kley, J.; Schleiffer, A.; Rombaut, P.; Herzog, F.; Clausen, T. Structural basis for the disaggregase activity and regulation of Hsp104. Elife 2016, 5, e21516. [Google Scholar] [CrossRef] [PubMed]
- Fiori, A.; Kucharíková, S.; Govaert, G.; Cammue, B.P.A.; Thevissen, K.; Van Dijck, P. The heat-induced molecular disaggregase Hsp104 of Candida albicans plays a role in biofilm formation and pathogenicity in a worm infection model. Eukaryot. Cell 2012, 11, 1012–1020. [Google Scholar] [CrossRef] [Green Version]
- Leach, M.D.; Stead, D.A.; Argo, E.; Brown, A.J.P. Identification of sumoylation targets, combined with inactivation of SMT3, reveals the impact of sumoylation upon growth, morphology, and stress resistance in the pathogen Candida albicans. Mol. Biol. Cell 2011, 22, 687–702. [Google Scholar] [CrossRef]
- Cuéllar, J.; Ludlam, W.G.; Tensmeyer, N.C.; Aoba, T.; Dhavale, M.; Santiago, C.; Bueno-Carrasco, M.T.; Mann, M.J.; Plimpton, R.L.; Makaju, A.; et al. Structural and functional analysis of the role of the chaperonin CCT in mTOR complex assembly. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Stoldt, V.; Rademacher, F.; Kehren, V.; Ernst, J.F.; Pearce, D.A.; Sherman, F. Review: The Cct Eukaryotic Chaperonin Subunits of Saccharomyces cerevisiae and other Yeasts. Yeast 1996, 12, 523–529. [Google Scholar] [CrossRef]
- Archibald, J.M.; Logsdon, J.M., Jr.; Doolittle, W.F. Origin and Evolution of Eukaryotic Chaperonins: Phylogenetic Evidence for Ancient Duplications in CCT Genes. Mol. Biol. Evol. 2000, 17, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Houry, W.A.; Frishman, D.; Eckerskorn, C.; Lottspeich, F.; Hartl, F.U. Identification of in vivo substrates of the chaperonin GroEL. Nature 1999, 402, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Thomas, J.O.; Chow, R.L.; Lee, G.H.; Cowan, N.J. A cytoplasmic chaperonin that catalyzes β-actin folding. Cell 1992, 69, 1043–1050. [Google Scholar] [CrossRef]
- Dekker, C.; Stirling, P.C.; McCormack, E.A.; Filmore, H.; Paul, A.; Brost, R.L.; Costanzo, M.; Boone, C.; Leroux, M.R.; Willison, K.R. The interaction network of the chaperonin CCT. Embo J. 2008, 27, 1827–1839. [Google Scholar] [CrossRef] [Green Version]
- Willison, K.R. The substrate specificity of eukaryotic cytosolic chaperonin CCT. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170192. [Google Scholar] [CrossRef]
- Yébenes, H.; Mesa, P.; Muñoz, I.G.; Montoya, G.; Valpuesta, J.M. Chaperonins: Two rings for folding. Trends Biochem. Sci. 2011, 36, 424–432. [Google Scholar] [CrossRef]
- Rademacher, F.; Kehren, V.; Stoldt, V.R.; Ernst, J.F. A Candida albicans chaperonin subunit (CaCct8p) as a suppressor of morphogenesis and Ras phenotypes in C. albicans and Saccharomyces cerevisiae. Microbiology 1998, 144, 2951–2960. [Google Scholar] [CrossRef] [Green Version]
- Caplan, T.; Polvi, E.J.; Xie, J.L.; Buckhalter, S.; Leach, M.D.; Robbins, N.; Cowen, L.E. Functional Genomic Screening Reveals Core Modulators of Echinocandin Stress Responses in Candida albicans. Cell Rep. 2018, 23, 2292–2298. [Google Scholar] [CrossRef] [Green Version]
- Deepe, G.S.; Gibbons, R.S. Cellular and molecular regulation of vaccination with heat shock protein 60 from Histoplasma capsulatum. Infect. Immun. 2002, 70, 3759–3767. [Google Scholar] [CrossRef] [Green Version]
- De Bastos Ascenço Soares, R.; Gomez, F.J.; De Almeida Soares, C.M.; Deepe, G.S. Vaccination with heat shock protein 60 induces a protective immune response against experimental Paracoccidioides brasiliensis pulmonary infection. Infect. Immun. 2008, 76, 4214–4221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mchaourab, H.S.; Godar, J.A.; Stewart, P.L. Structure and mechanism of protein stability sensors: Chaperone activity of small heat shock proteins. Biochemistry 2009, 48, 3828–3837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, K.L.; Giese, K.C.; Buan, N.R.; Vierling, E. Interactions between Small Heat Shock Protein Subunits and Substrate in Small Heat Shock Protein-Substrate Complexes. J. Biol. Chem. 2004, 279, 1080–1089. [Google Scholar] [CrossRef] [Green Version]
- Nisamedtinov, I.; Lindsey, G.G.; Karreman, R.; Orumets, K.; Koplimaa, M.; Kevvai, K.; Paalme, T. The response of the yeast Saccharomyces cerevisiae to sudden vs. gradual changes in environmental stress monitored by expression of the stress response protein Hsp12p. Fems Yeast Res. 2008, 8, 829–838. [Google Scholar] [CrossRef] [Green Version]
- Karreman, R.J.; Dague, E.; Gaboriaud, F.; Quilès, F.; Duval, J.F.L.; Lindsey, G.G. The stress response protein Hsp12p increases the flexibility of the yeast Saccharomyces cerevisiae cell wall. Biochim. Biophys. Acta Proteins Proteomics 2007, 1774, 131–137. [Google Scholar] [CrossRef]
- Mayer, F.L.; Wilson, D.; Jacobsen, I.D.; Miramón, P.; Slesiona, S.; Bohovych, I.M.; Brown, A.J.P.; Hube, B. Small but crucial: The novel small heat shock protein Hsp21 mediates stress adaptation and virulence in Candida albicans. PLoS ONE 2012, 7, e038584. [Google Scholar] [CrossRef] [Green Version]
- Mayer, F.L.; Wilson, D.; Hube, B. Hsp21 Potentiates Antifungal Drug Tolerance in Candida albicans. PLoS ONE 2013, 8, e60417. [Google Scholar] [CrossRef] [Green Version]
- Fu, M.S.; de Sordi, L.; Mühlschlegel, F.A. Functional characterization of the small heat shock protein Hsp12p from Candida albicans. PLoS ONE 2012, 7, e42894. [Google Scholar] [CrossRef] [PubMed]
- Maeng, S.; Ko, Y.J.; Kim, G.B.; Jung, K.W.; Floyd, A.; Heitman, J.; Bahn, Y.S. Comparative transcriptome analysis reveals novel roles of the ras and cyclic AMP signaling pathways in environmental stress response and antifungal drug sensitivity in Cryptococcus neoformans. Eukaryot. Cell 2010, 9, 360–378. [Google Scholar] [CrossRef] [Green Version]
- Kravats, A.N.; Hoskins, J.R.; Reidy, M.; Johnson, J.L.; Doyle, S.M.; Genest, O.; Masison, D.C.; Wickner, S. Functional and physical interaction between yeast Hsp90 and Hsp70. Proc. Natl. Acad. Sci. USA 2018, 115, E2210–E2219. [Google Scholar] [CrossRef] [Green Version]
- Doyle, S.M.; Hoskins, J.R.; Kravats, A.N.; Heffner, A.L.; Garikapati, S.; Wickner, S. Intermolecular Interactions between Hsp90 and Hsp70. J. Mol. Biol. 2019, 431, 2729–2746. [Google Scholar] [CrossRef]
- Patricia Hernández, M.; Sullivan, W.P.; Toft, D.O. The assembly and intermolecular properties of the Hsp70-Hop-Hsp90 molecular chaperone complex. J. Biol. Chem. 2002, 277, 38294–38304. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Smith, D.F. Hop as an adaptor in the heat shock protein 70 (Hsp70) and Hsp90 chaperone machinery. J. Biol. Chem. 1998, 273, 35194–35200. [Google Scholar] [CrossRef] [Green Version]
- Biebl, M.M.; Riedl, M.; Buchner, J. Hsp90 Co-chaperones Form Plastic Genetic Networks Adapted to Client Maturation. Cell Rep. 2020, 32, 108063. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Kakihara, Y.; Krogan, N.; Greenblatt, J.; Emili, A.; Zhang, Z.; Houry, W.A. An atlas of chaperone-protein interactions in Saccharomyces cerevisiae: Implications to protein folding pathways in the cell. Mol. Syst. Biol. 2009, 5, 1–14. [Google Scholar] [CrossRef]
- Schilke, B.A.; Ciesielski, S.J.; Ziegelhoffer, T.; Kamiya, E.; Tonelli, M.; Lee, W.; Cornilescu, G.; Hines, J.K.; Markley, J.L.; Craig, E.A. Broadening the functionality of a J-protein/Hsp70 molecular chaperone system. PLoS Genet. 2017, 13, e1007084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glover, J.R.; Lindquist, S. Hsp104, Hsp70, and Hsp40: A novel chaperone system that rescues previously aggregated proteins. Cell 1998, 94, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Nillegoda, N.B.; Stank, A.; Malinverni, D.; Alberts, N.; Szlachcic, A.; Barducci, A.; De Los Rios, P.; Wade, R.C.; Bukau, B. Evolution of an intricate J-protein network driving protein disaggregation in eukaryotes. Elife 2017, 6, e24560. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, G.; Andréasson, C. Hsp70-Hsp110 chaperones deliver ubiquitin-dependent and -independent substrates to the 26S proteasome for proteolysis in yeast. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [Green Version]
- Taipale, M.; Tucker, G.; Peng, J.; Krykbaeva, I.; Lin, Z.Y.; Larsen, B.; Choi, H.; Berger, B.; Gingras, A.C.; Lindquist, S. A quantitative chaperone interaction network reveals the architecture of cellular protein homeostasis pathways. Cell 2014, 158, 434–448. [Google Scholar] [CrossRef] [Green Version]
- Lamoth, F.; Juvvadi, P.R.; Soderblom, E.J.; Moseley, M.A.; Steinbach, W.J. Hsp70 and the cochaperone StiA (hop) orchestrate Hsp90-mediated caspofungin tolerance in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2015, 59, 4727–4733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Y.; Li, T.; Yu, C.; Sun, S. Candida albicans heat shock proteins and Hsps-associated signaling pathways as potential antifungal targets. Front. Cell. Infect. Microbiol. 2017, 7, 520. [Google Scholar] [CrossRef]
- Refos, J.M.; Vonk, A.G.; ten Kate, M.T.; Eadie, K.; Verbrugh, H.A.; Bakker-Woudenberg, I.A.J.M.; van de Sande, W.W.J. Addition of 17-(allylamino)-17-demethoxygeldanamycin to a suboptimal caspofungin treatment regimen in neutropenic rats with invasive pulmonary aspergillosis delays the time to death but does not enhance the overall therapeutic efficacy. PLoS ONE 2017, 12, e0180961. [Google Scholar] [CrossRef] [Green Version]
- Whitesell, L.; Robbins, N.; Huang, D.S.; McLellan, C.A.; Shekhar-Guturja, T.; LeBlanc, E.V.; Nation, C.S.; Hui, R.; Hutchinson, A.; Collins, C.; et al. Structural basis for species-selective targeting of Hsp90 in a pathogenic fungus. Nat. Commun. 2019, 10, 402. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.S.; Leblanc, E.V.; Shekhar-Guturja, T.; Robbins, N.; Krysan, D.J.; Pizarro, J.; Whitesell, L.; Cowen, L.E.; Brown, L.E. Design and Synthesis of Fungal-Selective Resorcylate Aminopyrazole Hsp90 Inhibitors. J. Med. Chem. 2020, 63, 2139–2180. [Google Scholar] [CrossRef]
- Gestwicki, J.E.; Shao, H. Inhibitors and chemical probes for molecular chaperone networks. J. Biol. Chem. 2019, 294, 2151–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freilich, R.; Arhar, T.; Abrams, J.L.; Gestwicki, J.E. Protein-Protein Interactions in the Molecular Chaperone Network. Acc. Chem. Res. 2018, 51, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Wisén, S.; Bertelsen, E.B.; Thompson, A.D.; Patury, S.; Ung, P.; Chang, L.; Evans, C.G.; Walter, G.M.; Wipf, P.; Carlson, H.A.; et al. Binding of a small molecule at a protein-protein interface regulates the chaperone activity of Hsp70-Hsp40. Acs Chem. Biol. 2010, 5, 611–622. [Google Scholar] [CrossRef]
Figure 1.
The major roles of heat shock proteins (HSPs) in fungal pathogens. The HSPs facilitate the acquisition of thermotolerance and allow human fungal pathogens to grow at human body temperature as well as to survive after heat shock at elevated temperatures. Several of these chaperones are required for morphological changes including the yeast to hyphal transition and conidiation. The HSPs, and in particular the Hsp90s, also facilitate antifungal drug tolerance in the major fungal pathogens suggesting that inhibitors would potentiate the activities of existing antifungals. Finally, in mammalian and insect models of infection when HSPs were pharmacologically inhibited or upon inoculation with deletion mutants lacking a gene for an HSP, they were often attenuated for virulence (indicated by the blue line in the hypothetical survival curve) compared to wild type or untreated strains (red line).
Figure 1.
The major roles of heat shock proteins (HSPs) in fungal pathogens. The HSPs facilitate the acquisition of thermotolerance and allow human fungal pathogens to grow at human body temperature as well as to survive after heat shock at elevated temperatures. Several of these chaperones are required for morphological changes including the yeast to hyphal transition and conidiation. The HSPs, and in particular the Hsp90s, also facilitate antifungal drug tolerance in the major fungal pathogens suggesting that inhibitors would potentiate the activities of existing antifungals. Finally, in mammalian and insect models of infection when HSPs were pharmacologically inhibited or upon inoculation with deletion mutants lacking a gene for an HSP, they were often attenuated for virulence (indicated by the blue line in the hypothetical survival curve) compared to wild type or untreated strains (red line).

Figure 2.
Schematic diagrams of the major classes of chaperones. (A) An Hsp90 dimer schematic indicating the ATP binding N-terminal domain (NTD) and the C-terminal domain (CTD) which is required for homodimerization. (B) An Hsp70 schematic indicating the nucleotide binding domain (NBD) and the substrate binding domain (SBD) including the lid of the SBD in an open conformation. (C) A schematic of an Hsp100/ClpB chaperone showing the stacked hexameric rings through which peptide strands are pulled to unfold them. (D) A schematic of a CCT/TRiC complex showing the stacked oligomeric rings which form a protected environment for protein folding in the lumen when ATP is bound and the chaperonin adopts a closed conformation.
Figure 2.
Schematic diagrams of the major classes of chaperones. (A) An Hsp90 dimer schematic indicating the ATP binding N-terminal domain (NTD) and the C-terminal domain (CTD) which is required for homodimerization. (B) An Hsp70 schematic indicating the nucleotide binding domain (NBD) and the substrate binding domain (SBD) including the lid of the SBD in an open conformation. (C) A schematic of an Hsp100/ClpB chaperone showing the stacked hexameric rings through which peptide strands are pulled to unfold them. (D) A schematic of a CCT/TRiC complex showing the stacked oligomeric rings which form a protected environment for protein folding in the lumen when ATP is bound and the chaperonin adopts a closed conformation.

Figure 3.
Network analyses of known and predicted interactions between chaperones in (A) S. cerevisiae, (B) C. albicans, (C) C. neoformans, and (D) A. fumigatus. STRING network analyses (https://string-db.org/, accessed on 5 January 2021) displaying the interactions between members of the Hsp100, Hsp90, Hsp70, JDP, sHsp, and chaperonin (Hsp60 and T-complex) families in S. cerevisiae and three of the major human fungal pathogens. Only experimentally determined interactions are shown including those inferred from interactions between putative homologs in other species. There are few known interactions with Hsp100s, JDPs, and sHsps even in S. cerevisiae. Furthermore, there are fewer characterized interactions within the chaperone networks of the human fungal pathogens compared to S. cerevisiae. Gene names are given where known and gene identification numbers from FungiDB (https://fungidb.org/fungidb/app, accessed on 5 January 2021) are provided otherwise (from Candida albicans SC5314, Cryptococcus neoformans var. neoformans JEC21, and Aspergillus fumigatus Af293).
Figure 3.
Network analyses of known and predicted interactions between chaperones in (A) S. cerevisiae, (B) C. albicans, (C) C. neoformans, and (D) A. fumigatus. STRING network analyses (https://string-db.org/, accessed on 5 January 2021) displaying the interactions between members of the Hsp100, Hsp90, Hsp70, JDP, sHsp, and chaperonin (Hsp60 and T-complex) families in S. cerevisiae and three of the major human fungal pathogens. Only experimentally determined interactions are shown including those inferred from interactions between putative homologs in other species. There are few known interactions with Hsp100s, JDPs, and sHsps even in S. cerevisiae. Furthermore, there are fewer characterized interactions within the chaperone networks of the human fungal pathogens compared to S. cerevisiae. Gene names are given where known and gene identification numbers from FungiDB (https://fungidb.org/fungidb/app, accessed on 5 January 2021) are provided otherwise (from Candida albicans SC5314, Cryptococcus neoformans var. neoformans JEC21, and Aspergillus fumigatus Af293).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Horianopoulos, L.C.; Kronstad, J.W. Chaperone Networks in Fungal Pathogens of Humans. J. Fungi 2021, 7, 209. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030209
AMA Style
Horianopoulos LC, Kronstad JW. Chaperone Networks in Fungal Pathogens of Humans. Journal of Fungi. 2021; 7(3):209. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030209
Chicago/Turabian StyleHorianopoulos, Linda C., and James W. Kronstad. 2021. "Chaperone Networks in Fungal Pathogens of Humans" Journal of Fungi 7, no. 3: 209. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030209
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.