
Improved Tolerance of Mycorrhizal Torreya grandis Seedlings to Sulfuric Acid Rain Related to Phosphorus and Zinc Contents in Shoots


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and AMF Preparation
2.2. Experimental Design
2.3. Measurement of Plant Biomass and AMF Colonization
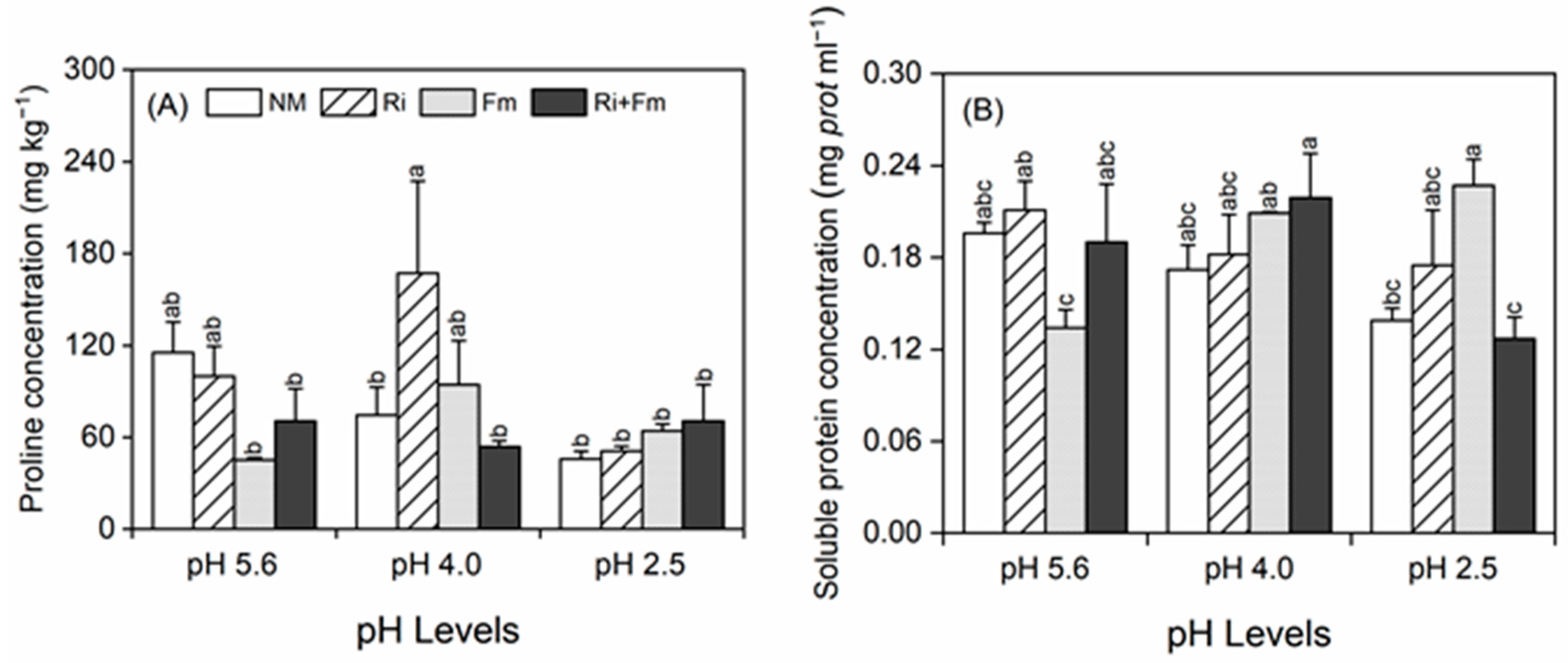
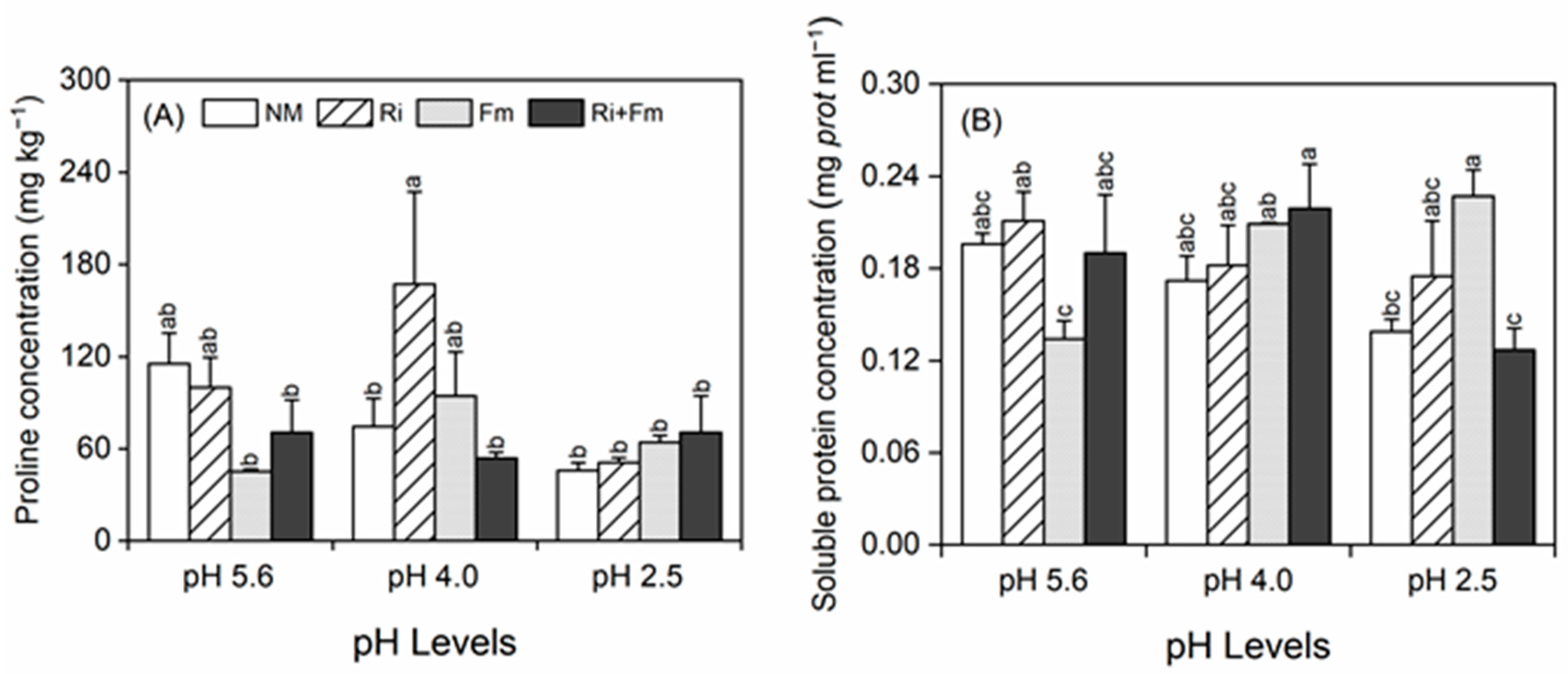
2.4. Measurement of Proline and Soluble Protein Contents
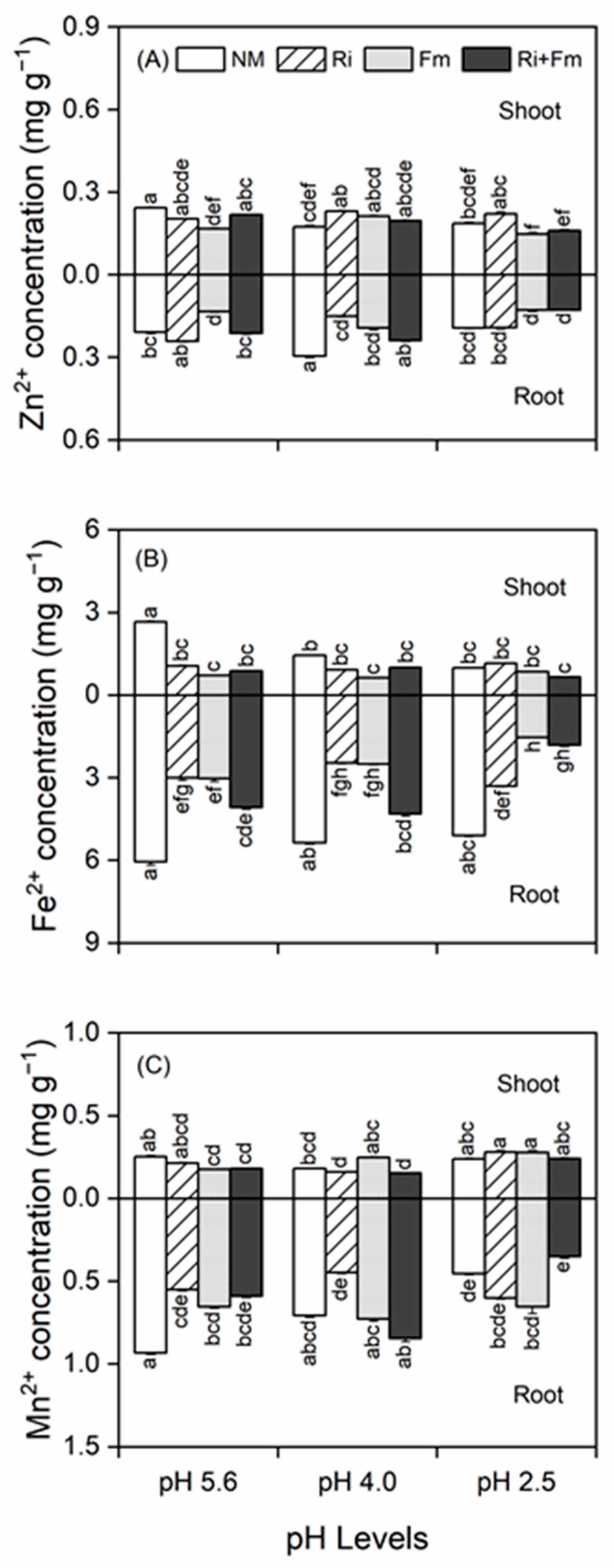
2.5. Measurement of Plant Nutrient Contents
2.6. Data Analysis
3. Results
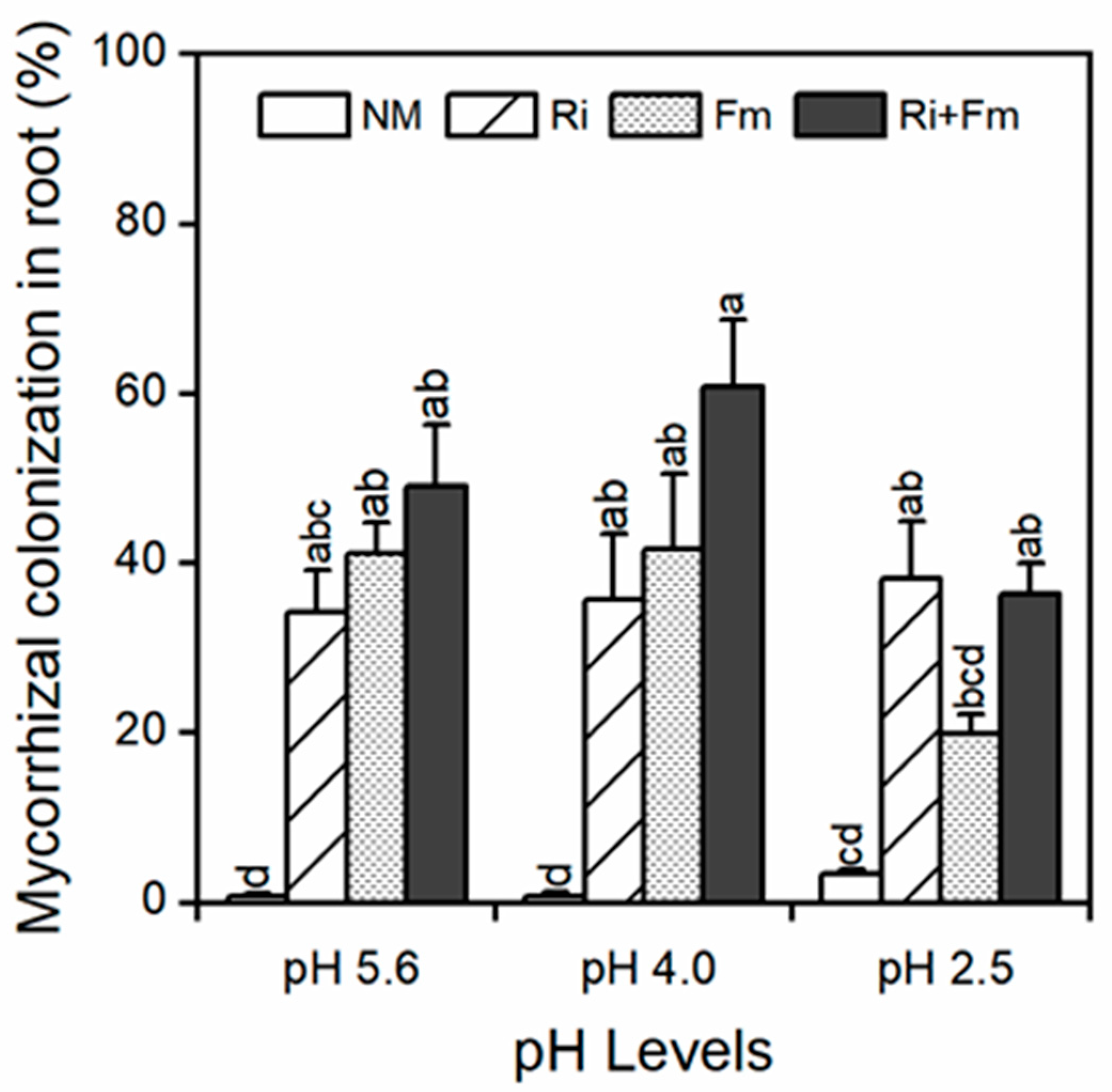
3.1. Mycorrhizal Colonization
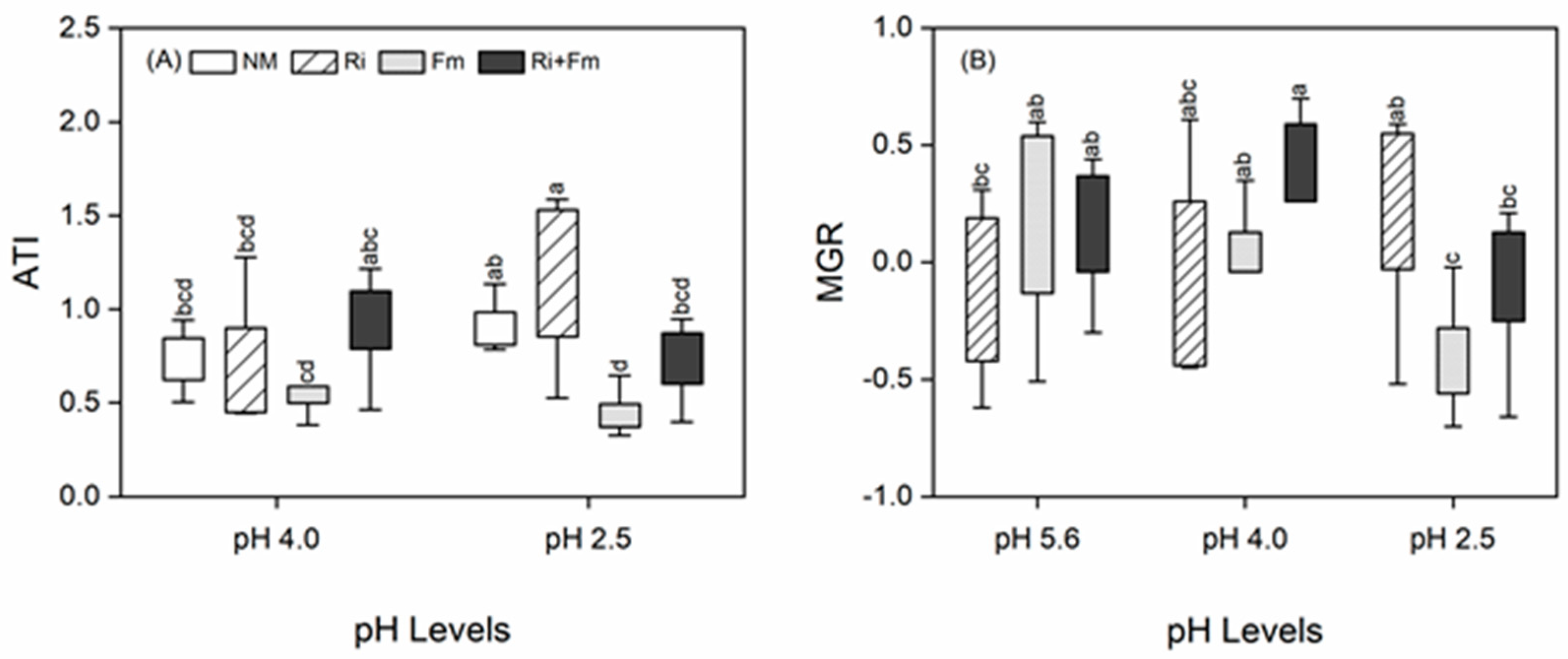
3.2. Plant Growth and Physiochemical Parameters
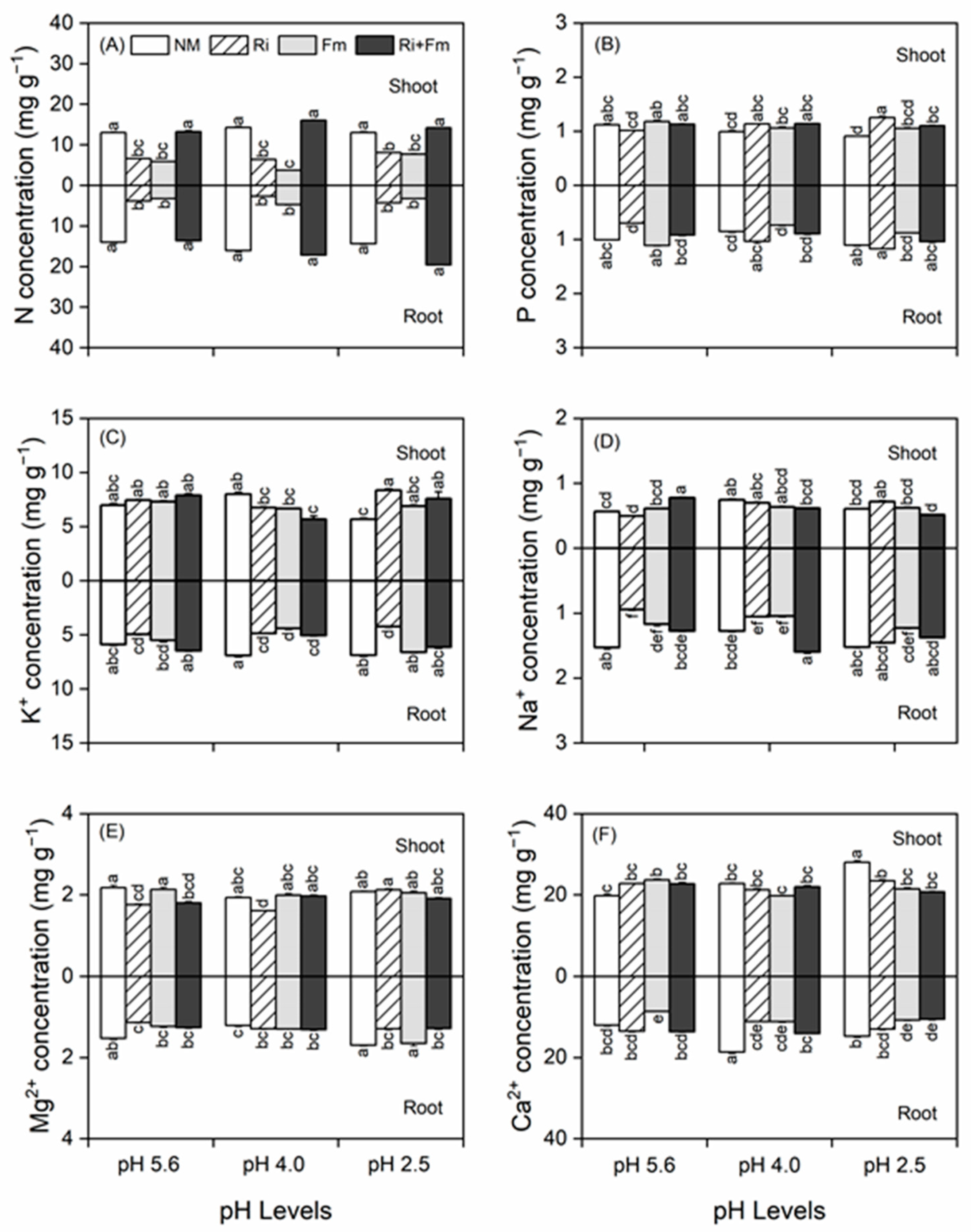
3.3. Concentrations of Shoot and Root Mineral Nutrients
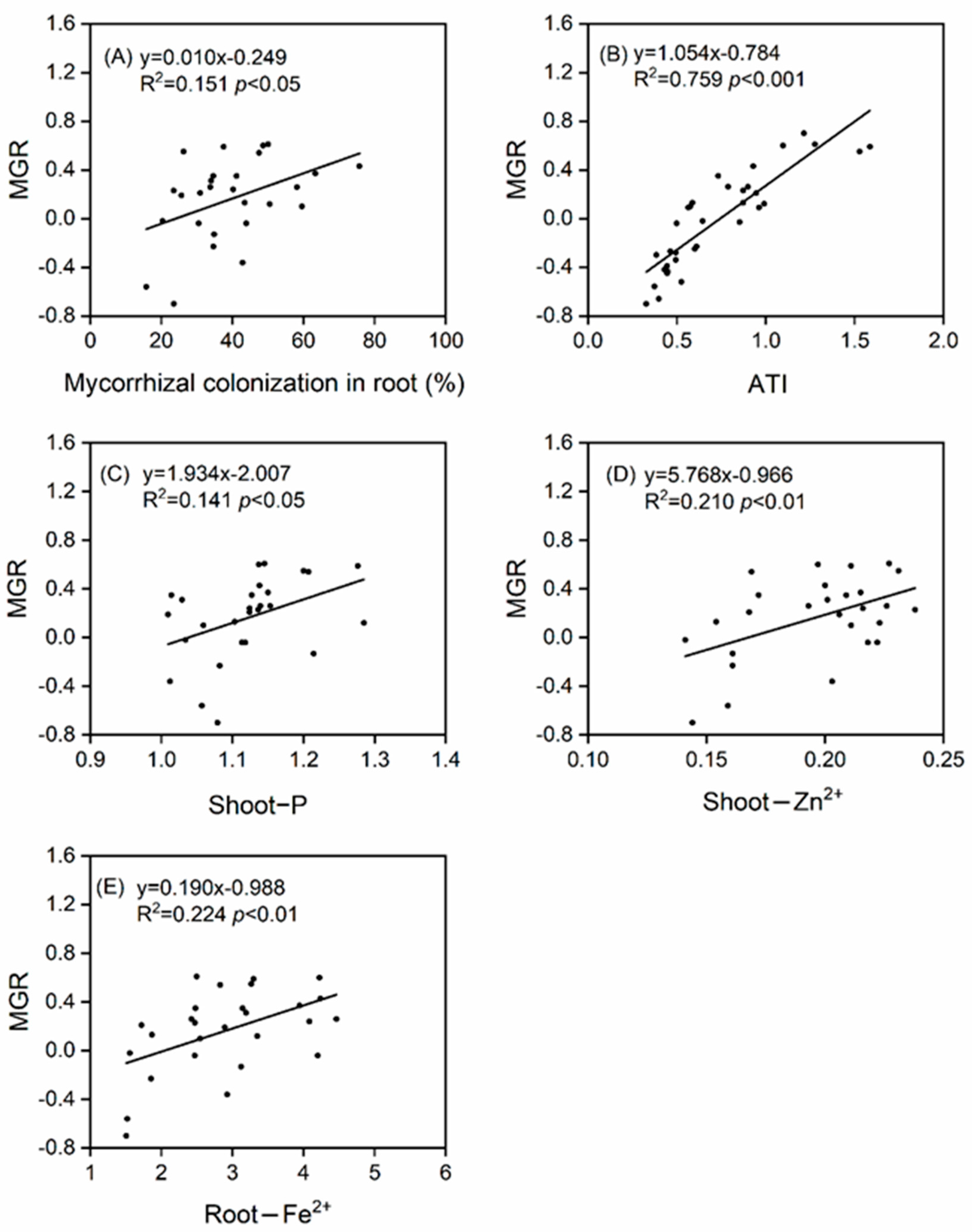
3.4. Mycorrhizal Benefits and the Relationships between Plant Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AR | acid rain |
| SAR | sulfuric acid rain |
| AMF | arbuscular mycorrhizal fungi |
| MGR | mycorrhizal growth response |
| ATI | acid-tolerance index |
| MC | mycorrhizal colonization |
| TDW | total dry weight |
| SP | soluble protein |
References
- Wei, H.; Ma, R.; Zhang, J.E.; Zhou, L.Y.; Liu, Z.Q.; Fan, Z.Y.; Yang, J.Y.; Shan, X.R.; Xiang, H.M. Quality dependence of litter decomposition and its carbon, nitrogen and phosphorus release under simulated acid rain treatments. Environ. Sci. Pollut. Res. 2020, 27, 19858–19868. [Google Scholar] [CrossRef]
- Andrade, G.C.; Castro, L.N.; Silva, L.C.D. Micromorphological alterations induced by simulated acid rain on the leaf surface of Joannesia princeps Vell. (Euphorbiaceae). Ecol. Indic. 2020, 116, 106526. [Google Scholar] [CrossRef]
- Cui, L.L.; Liang, J.H.; Fu, H.B.; Zhang, L.W. The contributions of socioeconomic and natural factors to the acid deposition over China. Chemosphere 2020, 253, 126491. [Google Scholar] [CrossRef]
- Wei, H.; Ma, R.; Zhang, J.E.; Saleem, M.; Liu, Z.Q.; Shan, X.R.; Yang, J.Y.; Xiang, H.M. Crop-litter type determines the structure and function of litter-decomposing microbial communities under acid rain conditions. Sci. Total Environ. 2020, 713, 136600. [Google Scholar] [CrossRef]
- Huang, X.F.; Li, X.; He, L.Y.; Feng, N.; Hu, M.; Niu, Y.W.; Zeng, L.W. 5-Year study of rainwater chemistry in a coastal mega-city in South China. Atmos. Res. 2010, 97, 185–193. [Google Scholar] [CrossRef]
- Larssen, T.; Seip, H.M.; Semb, A.; Mulder, J.; Muniz, I.P.; Vogt, R.D.; Lydersen, E.; Angell, V.; Dagang, T.; Eilertsen, O. Acid deposition and its effects in China: An overview. Environ. Sci. Policy 1999, 2, 9–24. [Google Scholar] [CrossRef]
- Niu, Y.W.; Pu, J.J.; Deng, F.P.; Qi, B. Analysis on spatial and temporal evolution of acid rain and its causes from 1992 to 2012 in Zhejiang. Environ. Monit. China 2017, 33, 55–62. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Lee, X.Q.; Cao, F. Chemical characteristics and sources of organic acids in precipitation at a semi-urban site in Southwest China. Atmos. Environ. 2011, 45, 413–419. [Google Scholar] [CrossRef]
- Pham, H.T.T.; Nguyen, A.T.; Nguyen, T.T.H.; Hens, L. Stakeholder Delphi-perception analysis on impacts and responses of acid rain on agricultural ecosystems in the Vietnamese upland. Environ. Dev. Sustain. 2019, 22, 4467–4493. [Google Scholar] [CrossRef]
- Wei, H.; Liu, W.; Zhang, J.E.; Qin, Z. Effects of simulated acid rain on soil fauna community composition and their ecological niches. Environ. Pollut. 2017, 220, 460–468. [Google Scholar] [CrossRef]
- Bao, G.Z.; Tang, W.Y.; An, Q.R.; Liu, Y.X.; Tian, J.Q.; Zhao, N.; Zhu, S.N. Physiological effects of the combined stresses of freezing-thawing, acid precipitation and deicing salt on alfalfa seedlings. BMC Plant Biol. 2020, 20, 204. [Google Scholar] [CrossRef]
- Da Fonseca, S.S.; Da Silva, B.R.S.; Lobato, A.K.D.S. 24-Epibrassinolide Positively Modulate Leaf Structures, Antioxidant System and Photosynthetic Machinery in Rice Under Simulated Acid Rain. J. Plant Growth Regul. 2020, 39, 1559–1576. [Google Scholar] [CrossRef]
- Liu, X.; Li, C.; Meng, M.J.; Zhai, L.; Zhang, B.; Jia, Z.H.; Gu, Z.Y.; Liu, Q.Q.; Zhang, Y.L.; Zhang, J.C. Comparative effects of the recovery from sulfuric and nitric acid rain on the soil enzyme activities and metabolic functions of soil microbial communities. Sci. Total Environ. 2020, 714, 136788. [Google Scholar] [CrossRef]
- Debnath, B.; Li, M.; Liu, S.; Pan, T.F.; Ma, C.L.; Qiu, D.L. Melatonin-mediate acid rain stress tolerance mechanism through alteration of transcriptional factors and secondary metabolites gene expression in tomato. Ecotox. Environ. Saf. 2020, 200, 110720. [Google Scholar] [CrossRef] [PubMed]
- Ju, S.M.; Wang, L.P.; Chen, J.Y. Effects of Silicon on the Growth, Photosynthesis and Chloroplast Ultrastructure of Oryza sativa L. Seedlings under Acid Rain Stress. Silicon 2019, 12, 655–664. [Google Scholar] [CrossRef]
- Liu, Z.Q.; Yang, J.Y.; Zhang, J.E.; Xiang, H.M.; Wei, H. A bibliometric analysis of research on acid rain. Sustainability 2019, 11, 3077. [Google Scholar] [CrossRef] [Green Version]
- Maltz, M.R.; Chen, Z.; Cao, J.X.; Arogyaswamy, K.; Shulman, H.; Aronson, E.L. Inoculation with Pisolithus tinctorius may ameliorate acid rain impacts on soil microbial communities associated with Pinus massoniana seedlings. Fungal Ecol. 2019, 40, 50–61. [Google Scholar] [CrossRef]
- Aguilera, P.; Cumming, J.R.; Oehl, F.; Cornejo, P.; Borie, F. Diversity of Arbuscular Mycorrhizal Fungi in Acidic Soils and Their Contribution to Aluminum Phytotoxicity Alleviation. In Aluminum Stress Adaptation in Plants: Signaling and Communication in Plants; Panda, S.K., Baluška, F., Eds.; Springer International Publishing: Cham, Switzerland, 2015; Volume 24, pp. 203–228. [Google Scholar] [CrossRef]
- Bunch, W.D.; Cowden, C.C.; Wurzburger, N.; Shefferson, R.P. Geography and soil chemistry drive the distribution of fungal associations in lady’s slipper orchid, Cypripedium acaule. Botany 2013, 91, 850–856. [Google Scholar] [CrossRef]
- Liu, X.D.; Feng, Z.W.; Zhao, Z.Y.; Zhu, H.H.; Yao, Q. Acidic soil inhibits the functionality of arbuscular mycorrhizal fungi by reducing arbuscule formation in tomato roots. Soil Sci. Plant Nutr. 2020, 66, 275–284. [Google Scholar] [CrossRef]
- Sylvia, D.M.; Williams, S.E. Vesicular–arbuscular mycorrhizae and environmental stress. In Mycorrhizae in Sustainable Agriculture; Bethlenfalvay, G.J., Linderman, R.G., Eds.; American Society of Agronomy Special Publications: Madison, WI, USA, 1992; pp. 101–124. [Google Scholar] [CrossRef]
- Genre, A.; Lanfranco, L.; Perotto, S.; Bonfante, P. Unique and common traits in mycorrhizal symbioses. Nat. Rev. Microbiol. 2020, 18, 649–660. [Google Scholar] [CrossRef]
- Pandey, D.; Kehri, H.K.; Zoomi, I.; Akhtar, O.; Singh, A.K. Mycorrhizal fungi: Biodiversity, ecological significance, and industrial applications. In Recent Advancement in White Biotechnology through Fungi: Diversity and Enzymes Perspectives; Yadav, A.N., Mishra, S., Singh, S., Gupta, A., Eds.; Springer: Cham, Switzerland, 2019; Volume 1, pp. 181–199. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.; Martin, F.M.; Selosse, M.A.; Sanders, I.R. Mycorrhizal ecology and evolution: The past, the present, and the future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef]
- Lumini, E.; Pan, J.; Magurno, F.; Huang, C.; Bianciotto, V.; Xue, X.; Balestrini, R.; Tedeschi, A. Native Arbuscular Mycorrhizal Fungi Characterization from Saline Lands in Arid Oases, Northwest China. J. Fungi 2020, 6, 80. [Google Scholar] [CrossRef]
- Clark, R.B. Arbuscular mycorrhizal adaptation, spore germination, root colonization, and host plant growth and mineral acquisition at low pH. Plant Soil 1997, 192, 15–22. [Google Scholar] [CrossRef]
- Vosátka, M.; Batkhuugyin, E.; Albrechtová, J. Response of three arbuscular mycorrhizal fungi to simulated acid rain and aluminium stress. Biol. Plant. 1999, 42, 289–296. [Google Scholar] [CrossRef]
- Anwar, G.; Lilleskov, E.A.; Chimner, R.A. Arbuscular mycorrhizal inoculation has similar benefits to fertilization for Thuja occidentalis L. seedling nutrition and growth on peat soil over a range of pH: Implications for restoration. New For. 2020, 51, 297–311. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Xu, J.; Hu, L.L.; Ren, M.L.; Tang, J.J.; Chen, X. Nurse effects mediated by acid-tolerance of target species and arbuscular mycorrhizal colonization in an acid soil. Plant Soil 2019, 441, 161–172. [Google Scholar] [CrossRef]
- Medeiros, C.A.B.; Clark, R.B.; Ellis, J.R. Growth and nutrient uptake of sorghum cultivated with vesicular-arbuscular mycorrhiza isolates at varying pH. Mycorrhiza 1994, 4, 185–191. [Google Scholar] [CrossRef]
- Vosátka, M.; Dodd, J.C. The role of different arbuscular mycorrhizal fungi in the growth of Calamagrostis villosa and Deschampsia flexuosa, in experiments with simulated acid rain. Plant Soil 1998, 200, 251–263. [Google Scholar] [CrossRef]
- Chen, L.L.; Lei, N.Y. Effect of soil microbe inoculation on Koelreuteria paniculata seedlings growth under simulated acid rain stress. Ecol. Environ. Sci. 2019, 28, 438–445. [Google Scholar] [CrossRef]
- Tang, H.; Hu, Y.Y.; Yu, W.W.; Song, L.L.; Wu, J.S. Growth, photosynthetic and physiological responses of Torreya grandis seedlings to varied light environments. Trees 2015, 29, 1011–1022. [Google Scholar] [CrossRef]
- Zhang, R.; Zhao, Y.X.; Lin, J.H.; Hu, Y.Y.; Hänninen, H.; Wu, J.S. Biochar application alleviates unbalanced nutrient uptake caused by N deposition in Torreya grandis trees and seedlings. For. Ecol. Manag. 2019, 432, 319–326. [Google Scholar] [CrossRef]
- Ye, W.; Li, Y.C.; Yu, W.W.; Ye, X.M.; Qian, Y.T.; Dai, W.S. Microbial biodiversity in rhizospheric soil of Torreya grandis ‘Merrillii’ relative to cultivation history. Chin. J. Appl. Ecol. 2018, 29, 3783–3792. [Google Scholar] [CrossRef]
- Qin, X.C.; Li, J.H. Study on Favourable Environment and Cultivation Techniques of Torreya grandis cv. Merrillii. Hortic. Seed 2012, 3–5. [Google Scholar] [CrossRef]
- Wang, Y.H.; Wang, M.Q.; Li, Y.; Wu, A.P.; Huang, J.Y. Effects of arbuscular mycorrhizal fungi on growth and nitrogen uptake of Chrysanthemum morifolium under salt stress. PLoS ONE 2018, 13, e0196408. [Google Scholar] [CrossRef]
- Porter, W.M. The ‘most probable number’ method for enumerating infective propagules of vesicular arbuscular mycorrhizal fungi in soil. Aust. J. Soil Res. 1979, 17, 515–519. [Google Scholar] [CrossRef]
- McNamara, N.P.; Black, H.I.J.; Beresford, N.A.; Parekh, N.R. Effects of acute gamma irradiation on chemical, physical and biological properties of soils. Appl. Soil Ecol. 2003, 24, 117–132. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Sonmez, O.; Aydemir, S.; Tuna, A.L.; Cullu, M.A. The influence of arbuscular mycorrhizal colonisation on key growth parameters and fruit yield of pepper plants grown at high salinity. Sci. Hortic. 2009, 121, 1–6. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.; Wiemken, A.; Sanders, I.R. Different arbuscular mycorrhizal fungi alter coexistence and resource distribution between co-occurring plant. New Phytol. 2003, 157, 569–578. [Google Scholar] [CrossRef]
- Song, X.Z.; Zhou, G.M.; Gu, H.H.; Qi, L.H. Management practices amplify the effects of N deposition on leaf litter decomposition of the Moso bamboo forest. Plant Soil 2015, 395, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Veiga, R.S.; Faccio, A.; Genre, A.; Pieterse, C.M.; Bonfante, P.; Van der Heijden, M.G. Arbuscular mycorrhizal fungi reduce growth and infect roots of the non-host plant Arabidopsis thaliana. Plant Cell Environ. 2013, 36, 1926–1937. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.; Hayman, D. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Guo, S.W.; Ma, J.X.; Xing, Y.Y.; Xu, Y.Q.; Jin, X.; Yan, S.M.; Shi, B.L. Artemisia annua L. aqueous extract as an alternative to antibiotics improving growth performance and antioxidant function in broilers. Ital. J. Anim. Sci. 2020, 19, 399–409. [Google Scholar] [CrossRef] [Green Version]
- Allen, S.E. Chemical Analysis of Ecological Materials, 2nd ed.; Blackwell Scientific Publications: London, UK, 1989. [Google Scholar]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Tullio, M.; Rivera, C.M.; Rea, E. Alleviation of salt stress by arbuscular mycorrhizal in zucchini plants grown at low and high phosphorus concentration. Biol. Fertil. Soils 2008, 44, 501–509. [Google Scholar] [CrossRef]
- Johnson, N.C.; Wilson, G.W.; Wilson, J.A.; Miller, R.M.; Bowker, M.A. Mycorrhizal phenotypes and the Law of the Minimum. New Phytol. 2015, 205, 1473–1484. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 26 January 2021).
- Macaulay, B.M.; Enahoro, G.E. Effects of simulated acid rain on the morphology, phenology and dry biomass of a local variety of maize (Suwan-1) in Southwestern Nigeria. Environ. Monit. Assess. 2015, 187, 622. [Google Scholar] [CrossRef] [PubMed]
- Saif, S.R. Growth responses of tropical forage plant species to vesicular-arbuscular mycorrhizae. Plant Soil 1987, 97, 23–35. [Google Scholar] [CrossRef]
- Raju, P.S.; Clark, R.B.; Ellis, J.R.; Maranville, J.W. Effects of Va mycorrhizae on growth and mineral uptake of sorghum grown at varied levels of soil acidity. Commun. Soil Sci. Plant Anal. 1988, 19, 919–931. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2015. [Google Scholar]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: London, UK, 2008. [Google Scholar]
- Lambais, M.R.; Cardoso, E.J.B.N. Response of Stylosanthes guianensis to endomycorrhizal fungi inoculation as affected by lime and phosphorus applications. Plant Soil 1993, 150, 109–116. [Google Scholar] [CrossRef]
- Albornoz, F.E.; Dixon, K.W.; Lambers, H. Revisiting mycorrhizal dogmas: Are mycorrhizas really functioning as they are widely believed to do? Soil Ecol. Lett. 2021, 3, 73–82. [Google Scholar] [CrossRef]
- Baum, C.; El-Tohamy, W.; Gruda, N. Increasing the productivity and product quality of vegetable crops using arbuscular mycorrhizal fungi: A review. Sci. Hortic. 2015, 187, 131–141. [Google Scholar] [CrossRef]
- Xue, Y.L.; Li, C.Y.; Wang, C.R.; Wang, Y.; Liu, J.; Chang, S.; Miao, Y.; Dang, T.H. Mechanisms of phosphorus uptake from soils by arbuscular mycorrhizal fungi. J. Soil Water Conserv. 2019, 33, 10–20. [Google Scholar] [CrossRef]
- Evelin, H.; Giri, B.; Kapoor, R. Contribution of Glomus intraradices inoculation to nutrient acquisition and mitigation of ionic imbalance in NaCl-stressed Trigonella foenum-graecum. Mycorrhiza 2012, 22, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.A.; Jakobsen, I.; Smith, S.E. Spatial differences in acquisition of soil phosphate between two arbuscular mycorrhizal fungi in symbiosis with Medicago truncatula. New Phytol. 2000, 147, 357–366. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.; Sanders, I.R. Mycorrhizal Ecology; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar] [CrossRef]
- Bonfante, P.; Genre, A. Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosis. Nat. Commun. 2010, 1, 48. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, M.; Boughattas, S.; Hu, S.; Oh, S.H.; Sa, T. A meta-analysis of arbuscular mycorrhizal effects on plants grown under salt stress. Mycorrhiza 2014, 24, 611–625. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, L.; Shao, C.; Zhang, N.; Wu, A.; Xie, J.; Qiu, Y.; He, X.; Pei, J.; Wang, X.; Wang, Y. Improved Tolerance of Mycorrhizal Torreya grandis Seedlings to Sulfuric Acid Rain Related to Phosphorus and Zinc Contents in Shoots. J. Fungi 2021, 7, 296. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040296
Xia L, Shao C, Zhang N, Wu A, Xie J, Qiu Y, He X, Pei J, Wang X, Wang Y. Improved Tolerance of Mycorrhizal Torreya grandis Seedlings to Sulfuric Acid Rain Related to Phosphorus and Zinc Contents in Shoots. Journal of Fungi. 2021; 7(4):296. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040296
Chicago/Turabian StyleXia, Lina, Changliang Shao, Naili Zhang, Aiping Wu, Jiangbo Xie, Yajing Qiu, Xiaobin He, Jia Pei, Xudong Wang, and Yanhong Wang. 2021. "Improved Tolerance of Mycorrhizal Torreya grandis Seedlings to Sulfuric Acid Rain Related to Phosphorus and Zinc Contents in Shoots" Journal of Fungi 7, no. 4: 296. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040296