
Evaluation of Sex Differences in Murine Diabetic Ketoacidosis and Neutropenic Models of Invasive Mucormycosis


 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Organisms and Culture Conditions
2.2. Antifungal Agents
2.3. Mouse Strains and Immunosuppression
2.4. Infection and Treatment
3. Results
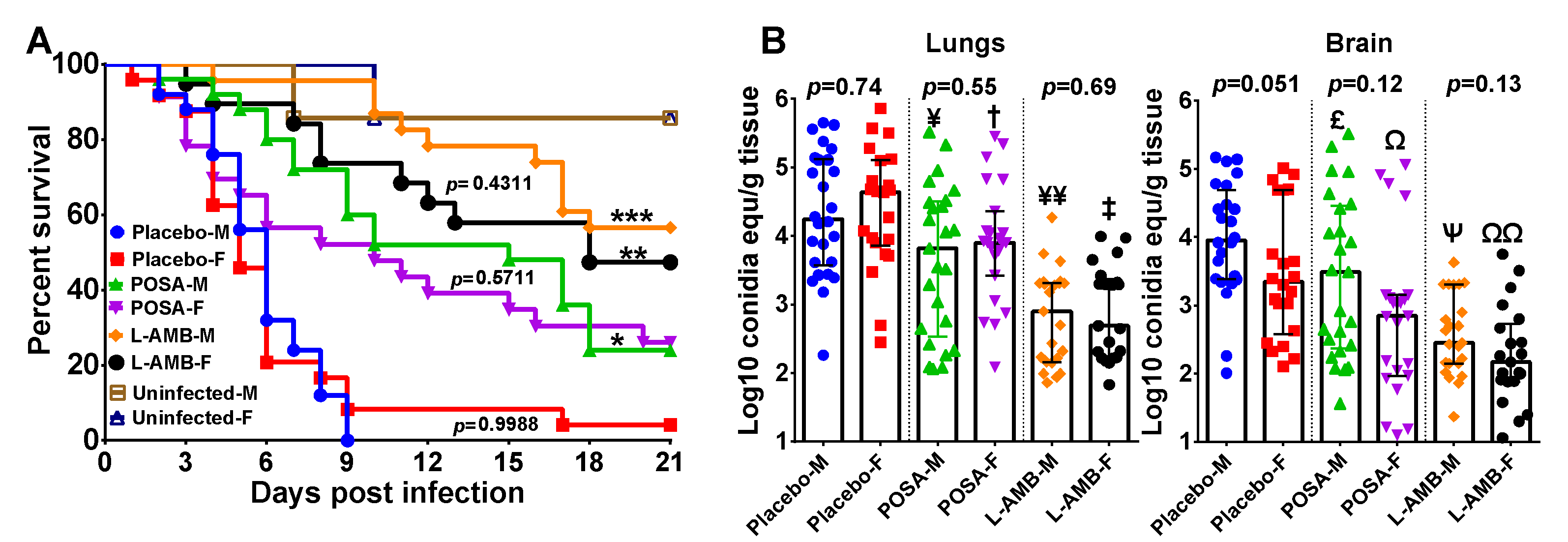
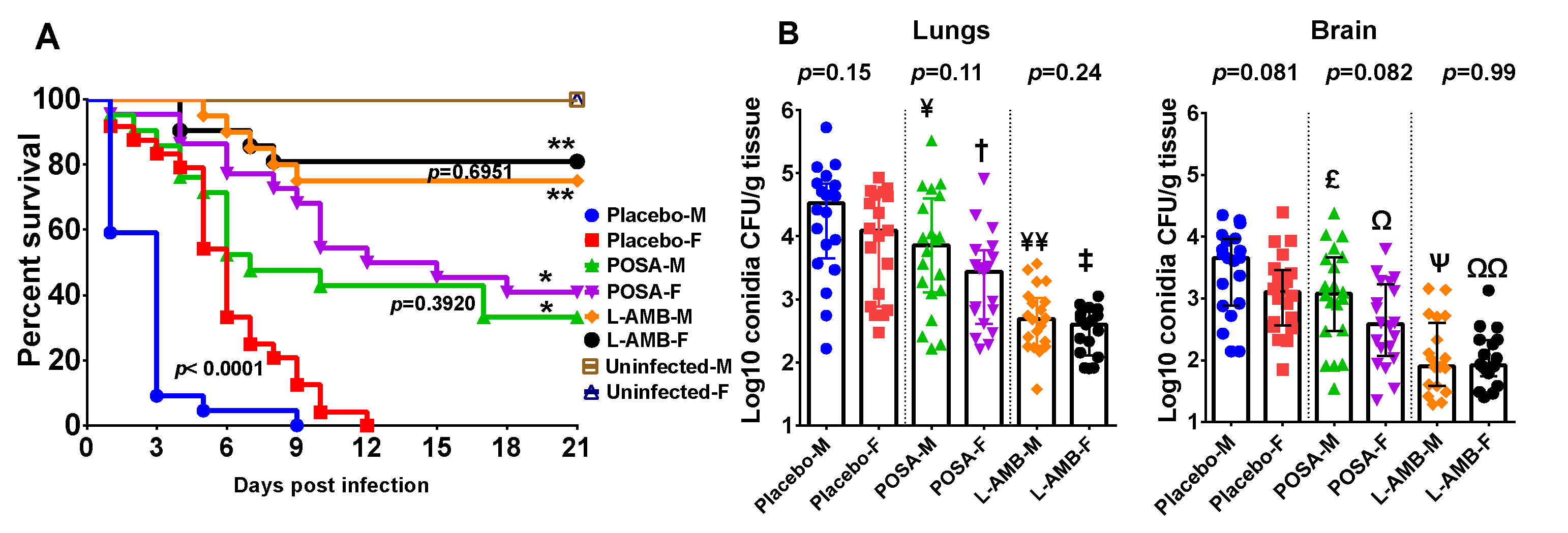
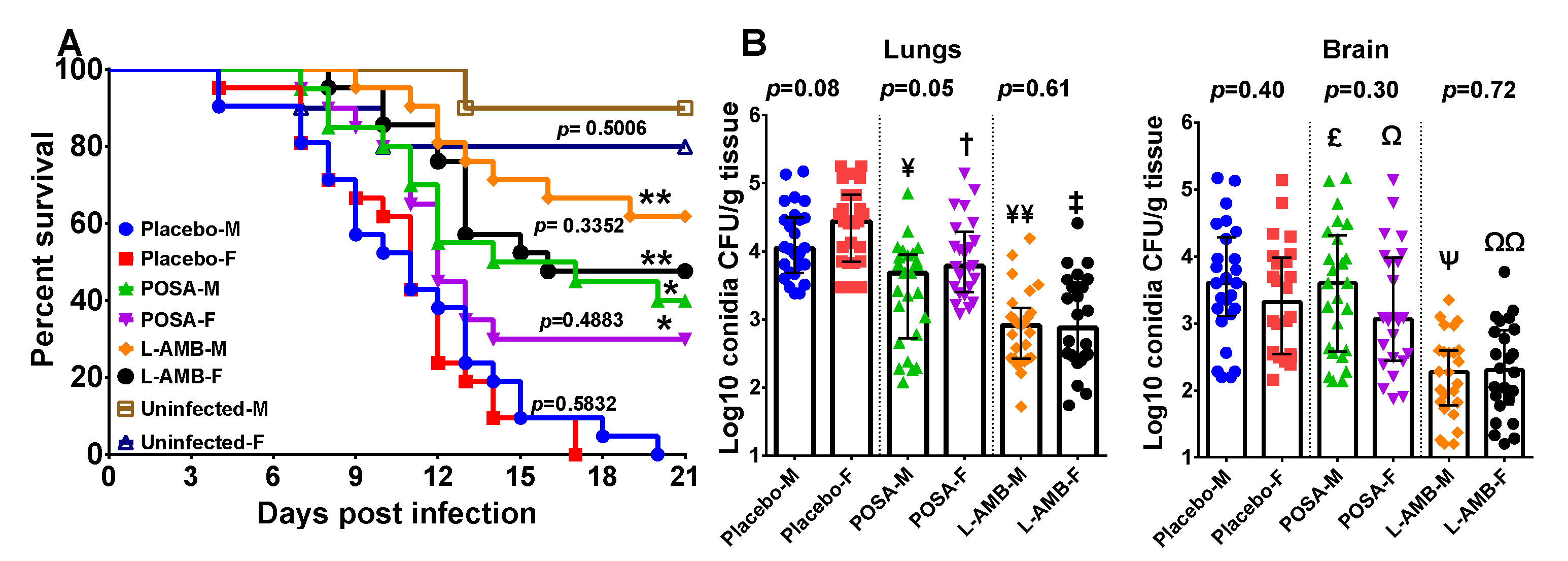
3.1. DKA Mice Infected with R. delemar
3.2. DKA Mice Infected with M. circinelloides
3.3. Neutropenic Mice Infected with M. circinelloides
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clayton, J.A.; Collins, F.S. Policy: NIH to balance sex in cell and animal studies. Nature 2014, 509, 282–283. [Google Scholar] [CrossRef]
- Collins, F.S.; Tabak, L.A. Policy: NIH plans to enhance reproducibility. Nature 2014, 505, 612–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NIH. Considerations of Sex as a Biological Variable in NIH-Funded Research. 2015. Available online: https://grants.nih.gov/grants/guide/notice-files/not-od-15-102.html (accessed on 1 April 2021).
- Spellberg, B.; Edwards, J., Jr.; Ibrahim, A. Novel perspectives on mucormycosis: Pathophysiology, presentation, and management. Clin. Microbiol. Rev. 2005, 18, 556–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugar, A.M. Agents of Mucormycosis and Related Species. In Principles and Practice of Infectious Diseases, 6th ed.; Mandell, G.L., Bennett, J.E., Dolin, R., Eds.; Elsevier: Philadelphia, PA, USA; Churchill Livingstone: London, UK, 2005; pp. 2973–2984. [Google Scholar]
- Cornely, O.A.; Alastruey-Izquierdo, A.; Arenz, D.; Chen, S.C.A.; Dannaoui, E.; Hochhegger, B.; Hoenigl, M.; Jensen, H.E.; Lagrou, K.; Lewis, R.E.; et al. Global guideline for the diagnosis and management of mucormycosis: An initiative of the European Confederation of Medical Mycology in cooperation with the Mycoses Study Group Education and Research Consortium. Lancet Infect. Dis. 2019, 19, e405–e421. [Google Scholar] [CrossRef]
- Roden, M.M.; Zaoutis, T.E.; Buchanan, W.L.; Knudsen, T.A.; Sarkisova, T.A.; Schaufele, R.L.; Sein, M.; Sein, T.; Chiou, C.C.; Chu, J.H.; et al. Epidemiology and Outcome of Zygomycosis: A Review of 929 Reported Cases. Clin. Infect. Dis. 2005, 41, 634–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleissner, B.; Schilling, A.; Anagnostopolous, I.; Siehl, I.; Thiel, E. Improved Outcome of Zygomycosis in Patients with Hematological Diseases? Leuk. Lymphoma 2004, 45, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Gebremariam, T.; Lee, H.; French, S.W.; Wiederhold, N.P.; Patterson, T.F.; Filler, S.G.; Ibrahim, A.S. Efficacy of Liposomal Amphotericin B and Posaconazole in Intratracheal Models of Murine Mucormycosis. Antimicrob. Agents Chemother. 2013, 57, 3340–3347. [Google Scholar] [CrossRef] [Green Version]
- Gebremariam, T.; Wiederhold, N.P.; Alqarihi, A.; Uppuluri, P.; Azie, N.; Edwards, J.E.; Ibrahim, A.S. Monotherapy or combination therapy of isavuconazole and micafungin for treating murine mucormycosis. J. Antimicrob. Chemother. 2016, 72, 462–466. [Google Scholar] [CrossRef] [Green Version]
- Gebremariam, T.; Alkhazraji, S.; Baldin, C.; Kovanda, L.; Wiederhold, N.P.; Ibrahim, A.S. Prophylaxis with Isavuconazole or Posaconazole Protects Immunosuppressed Mice from Pulmonary Mucormycosis. Antimicrob. Agents Chemother. 2017, 61, e02589-16. [Google Scholar] [CrossRef] [Green Version]
- Gebremariam, T.; Lin, L.; Liu, M.; Kontoyiannis, D.P.; French, S.; Edwards, J.E., Jr.; Filler, S.G.; Ibrahim, A.S. Bicarbonate Correction of Ketoacidosis Alleviates Mucormycosis Through Host, Pathogen. Interact. J. Clin. Investig. 2016, in press. [Google Scholar] [CrossRef] [Green Version]
- López-Fernández, L.; Sanchis, M.; Navarro-Rodríguez, P.; Nicolás, F.E.; Silva-Franco, F.; Guarro, J.; Garre, V.; Navarro-Mendoza, M.I.; Pérez-Arques, C.; Capilla, J. Understanding Mucor circinelloides pathogenesis by comparative genomics and phenotypical studies. Virulence 2018, 9, 707–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.N.; Najvar, L.K.; Bocanegra, R.; Loebenberg, D.; Graybill, J.R. In Vivo Activity of Posaconazole against Mucor spp. in an Immunosuppressed-Mouse Model. Antimicrob. Agents Chemother. 2002, 46, 2310–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, M.M.; Serena, C.; Mariné, M.; Pastor, F.J.; Guarro, J. Posaconazole Combined with Amphotericin B, an Effective Therapy for a Murine Disseminated Infection Caused by Rhizopus oryzae. Antimicrob. Agents Chemother. 2008, 52, 3786–3788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skiada, A.; Pagano, L.; Groll, A.; Zimmerli, S.; Dupont, B.; Lagrou, K.; Lass-Florl, C.; Bouza, E.; Klimko, N.; Gaustad, P.; et al. Zygomycosis in Europe: Analysis of 230 cases accrued by the registry of the European Confederation of Medical Mycology (ECMM) Working Group on Zygomycosis between 2005 and 2007. Clin. Microbiol. Infect. 2011, 17, 1859–1867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebremariam, T.; Liu, M.; Luo, G.; Bruno, V.; Phan, Q.T.; Waring, A.J.; Edwards, J.E.; Filler, S.G.; Yeaman, M.R.; Ibrahim, A.S. CotH3 mediates fungal invasion of host cells during mucormycosis. J. Clin. Investig. 2014, 124, 237–250. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.S.; Bowman, J.C.; Avanessian, V.; Brown, K.; Spellberg, B.; Edwards, J.E., Jr.; Douglas, C.M. Caspofungin Inhibits Rhizopus oryzae 1,3-β-d-Glucan Synthase, Lowers Burden in Brain Measured by Quantitative PCR, and Improves Survival at a Low but Not a High Dose during Murine Disseminated Zygomycosis. Antimicrob. Agents Chemother. 2005, 49, 721–727. [Google Scholar] [CrossRef] [Green Version]
- Castillo, P.; Wright, K.E.; Kontoyiannis, D.P.; Walsh, T.; Patel, S.; Chorvinsky, E.; Bose, S.; Hazrat, Y.; Omer, B.; Albert, N.; et al. A New Method for Reactivating and Expanding T Cells Specific for Rhizopus oryzae. Mol. Ther. Methods Clin. Dev. 2018, 9, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Gil-Lamaignere, C.; Simitsopoulou, M.; Roilides, E.; Maloukou, A.; Winn, R.M.; Walsh, T.J. Interferon- gamma and Granulocyte-Macrophage Colony-Stimulating Factor Augment the Activity of Polymorphonuclear Leukocytes against Medically Important Zygomycetes. J. Infect. Dis. 2005, 191, 1180–1187. [Google Scholar] [CrossRef] [Green Version]
- Montaño, D.E.; Voigt, K. Host Immune Defense upon Fungal Infections with Mucorales: Pathogen-Immune Cell Interactions as Drivers of Inflammatory Responses. J. Fungi 2020, 6, 173. [Google Scholar] [CrossRef]
- Bennouna, S.; Bliss, S.K.; Curiel, T.J.; Denkers, E.Y. Cross-Talk in the Innate Immune System: Neutrophils Instruct Recruitment and Activation of Dendritic Cells during Microbial Infection. J. Immunol. 2003, 171, 6052–6058. [Google Scholar] [CrossRef] [Green Version]
- Sawant, K.V.; Poluri, K.M.; Dutta, A.K.; Sepuru, K.M.; Troshkina, A.; Garofalo, R.P.; Rajarathnam, K. Chemokine CXCL1 mediated neutrophil recruitment: Role of glycosaminoglycan interactions. Sci. Rep. 2016, 6, 33123. [Google Scholar] [CrossRef] [Green Version]
- Murooka, T.T.; Rahbar, R.; Platanias, L.C.; Fish, E.N. CCL5-mediated T-cell chemotaxis involves the initiation of mRNA translation through mTOR/4E-BP1. Blood 2008, 111, 4892–4901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, A.E.; Berlin, A.A.; Lukacs, N.W. Respiratory syncytial virus-induced CCL5/RANTES contributes to exacerbation of allergic airway inflammation. Eur. J. Immunol. 2003, 33, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Lapteva, N.; Huang, X.F. CCL5 as an adjuvant for cancer immunotherapy. Expert Opin. Biol. Ther. 2010, 10, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Arango, M.; Yarzábal, L. T-cell dysfunction and hyperimmunoglobulinemia E in paracoccidioidomycosis. Mycopathologia 1982, 79, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Mota, N.; Rezkallah-Iwasso, M.; Peraçoli, M.; Audi, R.; Mendes, R.; Marcondes, J.; Marques, S.; Dillon, N.; Franco, M. Correlation between cell-mediated immunity and clinical forms of paracoccidioidomycosis. Trans. R. Soc. Trop. Med. Hyg. 1985, 79, 765–772. [Google Scholar] [CrossRef]
- Singer-Vermes, L.M.; Caldeira, C.B.; Burger, E.; Calich, V.L.G. Experimental murine paracoccidioidomycosis: Relationship among the dissemination of the infection, humoral and cellular immune responses. Clin. Exp. Immunol. 2008, 94, 75–79. [Google Scholar] [CrossRef]
- Pinzan, C.F.; Ruas, L.P.; Casabona-Fortunato, A.S.; Carvalho, F.C.; Roque-Barreira, M.-C. Immunological Basis for the Gender Differences in Murine Paracoccidioides brasiliensis Infection. PLoS ONE 2010, 5, e10757. [Google Scholar] [CrossRef] [Green Version]
- Restrepo, A.; Salazar, M.E.; Cano, L.E.; Stover, E.P.; Feldman, D.; Stevens, D.A. Estrogens inhibit mycelium-to-yeast transformation in the fungus Paracoccidioides brasiliensis: Implications for resistance of females to paracoccidioidomycosis. Infect. Immun. 1984, 46, 346–353. [Google Scholar] [CrossRef] [Green Version]
- Salazar, M.E.; Restrepo, A.; Stevens, D.A. Inhibition by estrogens of conidium-to-yeast conversion in the fungus Paracoccidioides brasiliensis. Infect. Immun. 1988, 56, 711–713. [Google Scholar] [CrossRef] [Green Version]
- Sano, A.; Nishimura, K.; Miyaji, M. Effects of Sex Hormones on Sexual Difference of Experimental Paracoccidioidomycosis. Nippon. Ishinkin Gakkai Zasshi 1999, 40, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Muchmore, H.G.; McKown, B.A.; Mohr, J.A. Effects of the steroid hormones on the proliferation of Paracoccidioidomicosis braziliensis. Boletin de la Sanit. Panam. Pan Am. Sanit. Bur. 1974, 77, 55–70. [Google Scholar]
- Schmidt, S.; Tramsen, L.; Perkhofer, S.; Lass-Flörl, C.; Röger, F.; Schubert, R.; Lehrnbecher, T. Characterization of the Cellular Immune Responses to Rhizopus oryzae With Potential Impact on Immunotherapeutic Strategies in Hematopoietic Stem Cell Transplantation. J. Infect. Dis. 2012, 206, 135–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbrecht, R.; Denning, D.W.; Patterson, T.F.; Bennett, J.E.; Greene, R.E.; Oestmann, J.-W.; Kern, W.V.; Marr, K.A.; Ribaud, P.; Lortholary, O.; et al. Voriconazole versus Amphotericin B for Primary Therapy of Invasive Aspergillosis. N. Engl. J. Med. 2002, 347, 408–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maertens, J.A.; Raad, I.I.; Marr, K.A.; Patterson, T.F.; Kontoyiannis, D.P.; Cornely, O.A.; Bow, E.J.; Rahav, G.; Neofytos, D.; Aoun, M.; et al. Isavuconazole versus voriconazole for primary treatment of invasive mould disease caused by Aspergillus and other filamentous fungi (SECURE): A phase 3, randomised-controlled, non-inferiority trial. Lancet 2016, 387, 760–769. [Google Scholar] [CrossRef]
- Cornely, O.A.; Maertens, J.; Winston, D.J.; Perfect, J.; Ullmann, A.J.; Walsh, T.J.; Helfgott, D.; Holowiecki, J.; Stockelberg, D.; Goh, Y.-T.; et al. Posaconazole vs. Fluconazole or Itraconazole Prophylaxis in Patients with Neutropenia. N. Engl. J. Med. 2007, 356, 348–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullmann, A.J.; Lipton, J.H.; Vesole, D.H.; Chandrasekar, P.; Langston, A.; Tarantolo, S.R.; Greinix, H.; De Azevedo, W.M.; Reddy, V.; Boparai, N.; et al. Posaconazole or Fluconazole for Prophylaxis in Severe Graft-versus-Host Disease. N. Engl. J. Med. 2007, 356, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Marty, F.M.; Ostrosky-Zeichner, L.; Cornely, O.A.; Mullane, K.M.; Perfect, J.R.; Thompson, G.R.; Alangaden, G.J.; Brown, J.M.; Fredricks, D.N.; Heinz, W.J.; et al. Isavuconazole treatment for mucormycosis: A single-arm open-label trial and case-control analysis. Lancet Infect. Dis. 2016, 16, 828–837. [Google Scholar] [CrossRef]
- Skiada, A.; Lass-Floerl, C.; Klimko, N.; Ibrahim, A.; Roilides, E.; Petrikkos, G. Challenges in the diagnosis and treatment of mucormycosis. Med Mycol. 2018, 56, S93–S101. [Google Scholar] [CrossRef] [Green Version]
- Xu-Vanpala, S.; Deerhake, M.E.; Wheaton, J.D.; Parker, M.E.; Juvvadi, P.R.; Maciver, N.; Ciofani, M.; Shinohara, M.L. Functional heterogeneity of alveolar macrophage population based on expression of CXCL2. Sci. Immunol. 2020, 5, eaba7350. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Gebremariam, T.; Schwartz, J.A.; Edwards, J.E.; Spellberg, B. Posaconazole Mono- or Combination Therapy for Treatment of Murine Zygomycosis. Antimicrob. Agents Chemother. 2008, 53, 772–775. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gebremariam, T.; Alkhazraji, S.; Alqarihi, A.; Wiederhold, N.P.; Najvar, L.K.; Patterson, T.F.; Filler, S.G.; Ibrahim, A.S. Evaluation of Sex Differences in Murine Diabetic Ketoacidosis and Neutropenic Models of Invasive Mucormycosis. J. Fungi 2021, 7, 313. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040313
Gebremariam T, Alkhazraji S, Alqarihi A, Wiederhold NP, Najvar LK, Patterson TF, Filler SG, Ibrahim AS. Evaluation of Sex Differences in Murine Diabetic Ketoacidosis and Neutropenic Models of Invasive Mucormycosis. Journal of Fungi. 2021; 7(4):313. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040313
Chicago/Turabian StyleGebremariam, Teclegiorgis, Sondus Alkhazraji, Abdullah Alqarihi, Nathan P. Wiederhold, Laura K. Najvar, Thomas F. Patterson, Scott G. Filler, and Ashraf S. Ibrahim. 2021. "Evaluation of Sex Differences in Murine Diabetic Ketoacidosis and Neutropenic Models of Invasive Mucormycosis" Journal of Fungi 7, no. 4: 313. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040313