
Bioprospecting of Rhizosphere-Resident Fungi: Their Role and Importance in Sustainable Agriculture

 , , , , ,
, , , , ,
Abstract
:1. Introduction
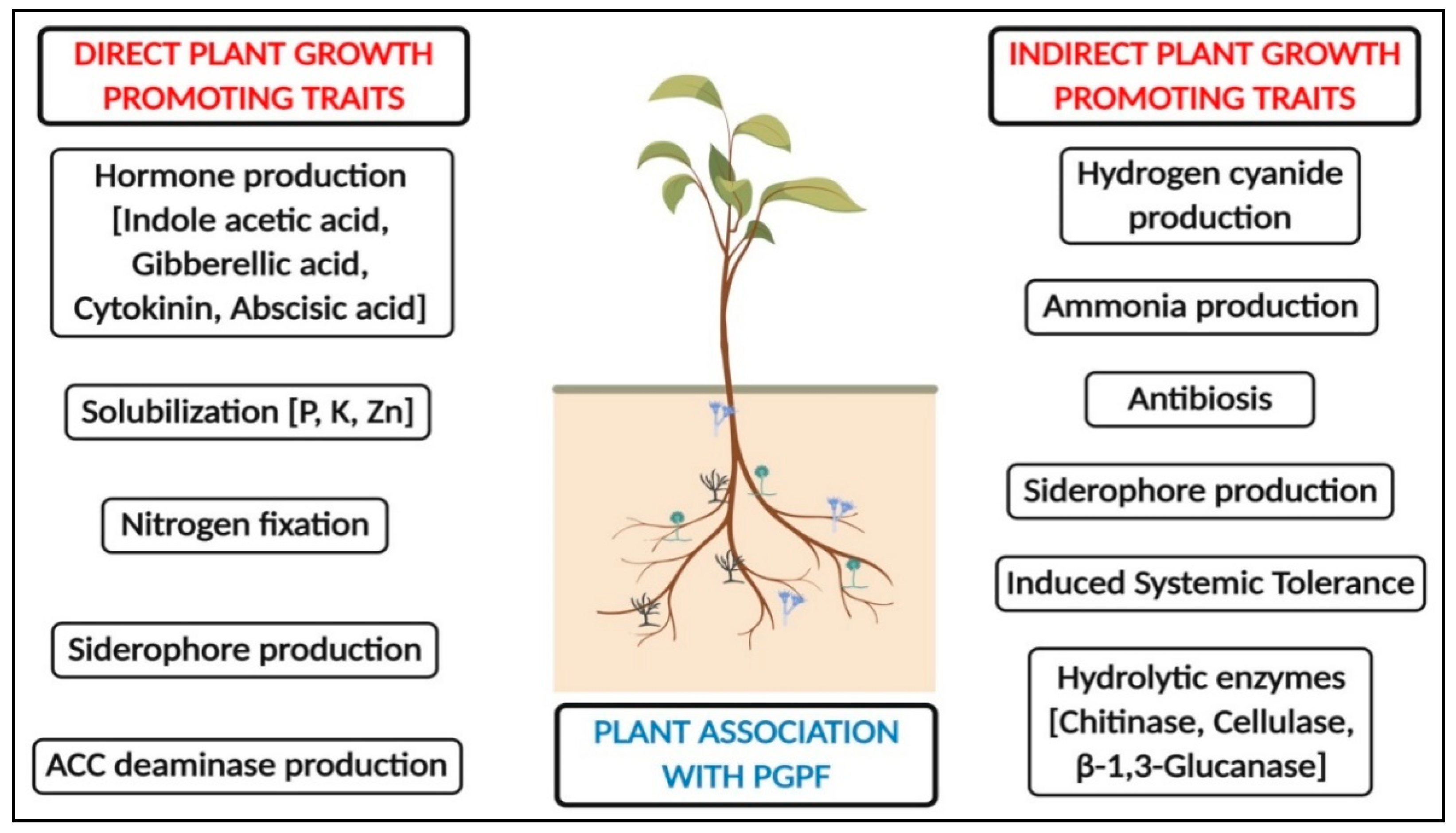
2. Beneficial Aspects of Rhizosphere-Resident PGPF
2.1. Production of Phytohormones
2.2. Plant Growth Promotion
2.3. Mineralization of Soil Nutrients
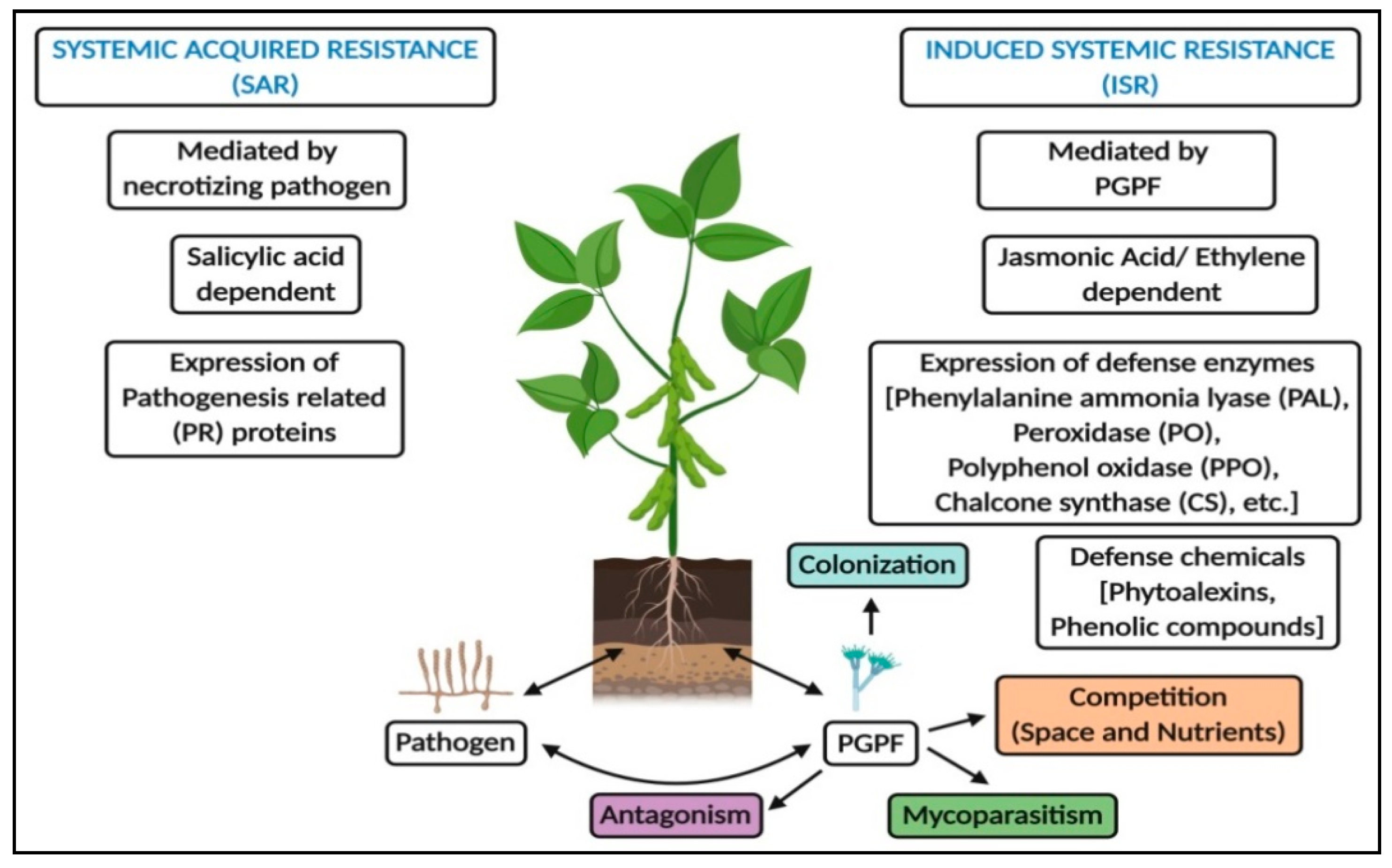
2.4. Resistance against Stressors
2.4.1. Antagonism
2.4.2. Induction of Resistance
2.4.3. Morphological and Histochemical Defense
2.4.4. Biochemical Defense
2.4.5. Defense Signaling
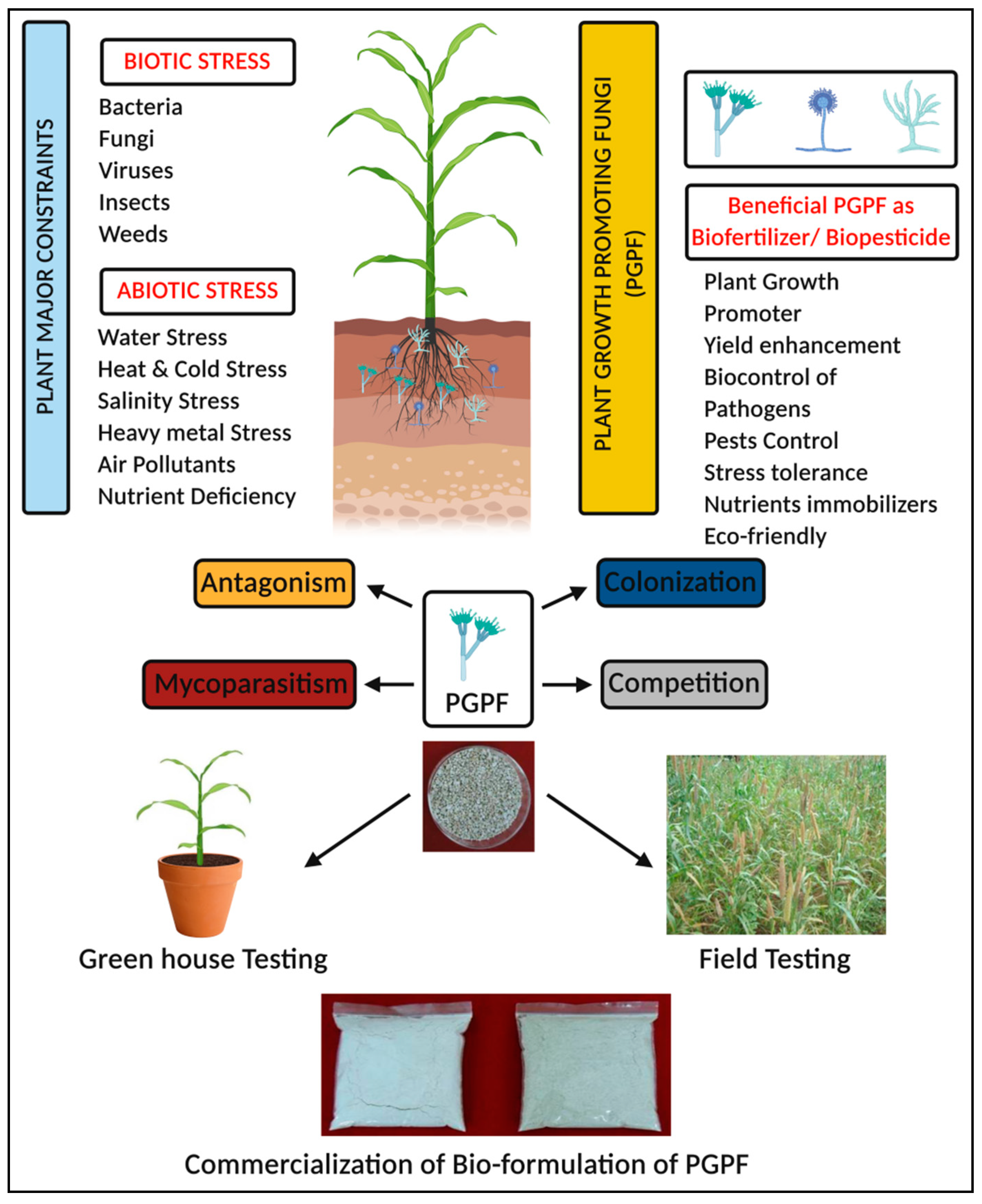
2.4.6. PGPF in Abiotic Stress Improvement
2.5. PGPF as a Source of Alternatives
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PGPF | Plant Growth Promoting Fungi |
| ISR | Induction of Systemic Resistance |
| SAR | Systemic Acquired Resistance |
| JA | Jasmonic acid |
| ET | Ethylene |
| GLU | Glucanase |
| CHI | Chitinase |
| IAA | Indole acetic acid |
| GA | Gibberellic acid |
| TCP | Tri-calcium phosphate |
| ROS | Reactive oxygen species |
| H2O2 | Hydrogen peroxide |
| PAL | Phenylalanine ammonia lyase |
| PPO | Polyphenol oxidase |
| POD | Peroxidase |
| CAT | Catalase |
| SOD | Superoxide Dismutase |
| APX | Ascorbate Peroxidase |
| POX | Peroxidase |
| LOX | Lipoxygenase |
| SA | Salicylic acid |
| GAB | Gibberellin related molecule |
| RNO | Reactive Nitrogen Species |
| HR | Hypersensitive Reaction |
References
- Desa, U.N. World Population Prospects 2019: Highlights; United Nations Department for Economic and Social Affairs: New York, NY, USA, 2019. [Google Scholar]
- Wimalawansa, S.A.; Wimalawansa, S.J. Agrochemical-related environmental pollution: Effects on human health. GJBAHS 2014, 3, 72–83. [Google Scholar]
- Hyakumachi, M. Plant-growth-promoting fungi from turfgrass rhizosphere with potential for disease suppression. Soil Microorg. 1994, 44, 53–68. [Google Scholar]
- Hyakumachi, M.; Kubota, M. Fungi as plant growth promoter and disease suppressor. Fungal Biotechnol. Agric. Food Environ. Appl. 2004, 21, 101–110. [Google Scholar]
- Murali, M.; Amruthesh, K.N.; Sudisha, J.; Niranjana, S.R.; Shetty, H.S. Screening for plant growth promoting fungi and their ability for growth promotion and induction of resistance in pearl millet against downy mildew disease. J. Phytol. 2012, 4, 30–36. [Google Scholar]
- Hossain, M.M.; Sultana, F.; Islam, S. Plant growth-promoting fungi (PGPF): Phytostimulation and induced systemic resistance. In Plant-Microbe Interactions in Agro-Ecological Perspectives; Springer: Singapore, 2017; pp. 135–191. [Google Scholar]
- Naziya, B.; Murali, M.; Amruthesh, K.N. Plant Growth-Promoting Fungi (PGPF) Instigate Plant Growth and Induce Disease Resistance in Capsicum annuum L. upon Infection with Colletotrichum capsici (Syd.). Butl. Bisby Biomol. 2020, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Meera, M.S.; Shivanna, M.B.; Kageyama, K.; Hyakumachi, M. Plant growth promoting fungi from zoysiagrass rhizosphere as potential inducers of systemic resistance in cucumbers. Phytopathology 1994, 84, 1399–1406. [Google Scholar] [CrossRef]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef]
- Shivanna, M.B.; Meera, M.S.; Kubota, M.; Hyakumachi, M. Promotion of growth and yield in cucumber by zoysia grass rhizosphere fungi. Microbes Environ. 2005, 20, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Shoresh, M.; Yedidia, I.; Chet, I. Involvement of jasmonic acid/ethylene signalling pathway in the systemic resistance induced in cucumber by Trichoderma asperellum T203. Phytopathology 2005, 95, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Tucci, M.; Ruocco, M.; De Masi, L.; De Palma, M.; Lorito, M. The beneficial effect of Trichoderma spp. on tomato is modulated by plant genotype. Mol. Plant Pathol. 2011, 12, 341–354.sss. [Google Scholar] [CrossRef]
- Nawrocka, J.; Małolepsza, U.; Szymczak, K.; Szczech, M. Involvement of metabolic components, volatile compounds, PR proteins, and mechanical strengthening in multilayer protection of cucumber plants against Rhizoctonia solani activated by Trichoderma atroviride TRS25. Protoplasma 2018, 255, 359–373. [Google Scholar] [CrossRef] [Green Version]
- Madi, L.; Katan, J. Penicillium janczewskii and its metabolites, applied to leaves, elicit systemic acquired resistance to stem rot caused by Rhizoctonia solani. Physiol. Mol. Plant Pathol. 1998, 53, 163–175. [Google Scholar] [CrossRef]
- Aamir, M.; Kashyap, S.P.; Singh, V.K.; Dubey, M.K.; Ansari, W.A.; Upadhyay, R.S.; Singh, S. Trichoderma erinaceum Bio-priming Modulates the WRKYs Defense Programming in tomato against the Fusarium oxysporum f. sp. lycopersici (Fol) Challenged Condition. Front. Plant Sci. 2019, 10, 911. [Google Scholar] [CrossRef] [Green Version]
- Yadav, J.; Verma, J.P.; Tiwari, K.N. Plant growth promoting activities of fungi and their effect on chickpea plant growth. Asian J. Biol. Sci. 2011, 4, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Abri, T.K.; Sengin, E.L.; Sjahrir, R. Production of indole acetic acid (IAA) hormone from fungal isolates collected from rhizosphere of aromatic rice in Tana Toraja. Int. J. Curr. Res. Biosci. Plant Biol. 2015, 2, 198–201. [Google Scholar]
- Kumar, N.V.; Rajam, K.S.; Rani, M.E. Plant growth promotion efficacy of indole acetic acid (IAA) produced by a mangrove associated fungi- Trichoderma viride VKF3. Int. J. Curr. Microbiol. App. Sci. 2017, 6, 2692–2701. [Google Scholar] [CrossRef]
- Ozimek, E.; Jaroszuk-Sciseł, J.; Bohacz, J.; Korniłłowicz-Kowalska, T.; Tyskiewicz, R.; Słomka, A.; Nowak, A.; Hanaka, A. Synthesis of indole acetic acid, gibberellic acid and ACC-deaminase by Mortierella strains promote winter wheat seedlings growth under different conditions. Int. J. Mol. Sci. 2018, 19, 3218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillespie-Sasse, L.; Alamassi, J.F.; Ghisalberti, E.L.; Sivasithamparam, K. Use of clean seedling assay to test plant growth promotion by exudates from a sterile red fungus. Soil Biol. Biochem. 1991, 23, 95–97. [Google Scholar] [CrossRef]
- Hamayun, M.; Khan, S.A.; Khan, A.L.; Rehman, G.; Kim, Y.H.; Iqbal, I.; Hussain, J.; Sohn, E.Y.; Lee, I.J. Gibberellin production and plant growth promotion from pure cultures of Cladosporium sp. MH-6 isolated from cucumber (Cucumis sativus L.). Mycologia 2010, 102, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Vadassery, J.; Ritter, C.; Venus, Y.; Camehl, I.; Varma, A.; Shahollari, B.; Novák, O.; Strnad, M.; Ludwig-Müller, J.; Oelmüller, R. The role of auxins and cytokinins in the mutualistic interaction between Arabidopsis and Piriformospora indica. Mol. Plant -Microbe Interact. 2008, 21, 1371–1383. [Google Scholar] [CrossRef] [Green Version]
- Speakman, J.B.; Kruger, W. Control of Gaeumannomyces graminis var. tritici by a sterile, black mycelial fungus. J. Plant Dis. Prot. 1984, 91, 391–395. [Google Scholar]
- Martinez-Medina, A.; Alguacil, M.D.M.; Pascual, J.A.; Van Wees, S.C. Phytohormone profiles induced by Trichoderma isolates correspond with their biocontrol and plant growth-promoting activity on melon plants. J. Chem. Ecol. 2014, 40, 804–815. [Google Scholar] [CrossRef]
- Murali, M.; Sudisha, J.; Amruthesh, K.N.; Shetty, H.S. Rhizosphere fungus Penicillium chrysogenum promotes growth and induces defence-related genes and downy mildew disease resistance in pearl millet. Plant Biol. 2013, 15, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Dewan, M.; Sivasithamparam, K. Growth promotion of rotation crop species by sterile fungus from wheat and effect of soil temperature and water potential on its suppression of take-all. Mycol. Res. 1989, 93, 156–160. [Google Scholar] [CrossRef]
- Shivanna, M.B.; Meera, M.S.; Hyakumachi, M. Sterile fungi from zoysiagrass rhizosphere as plant growth promoters in spring wheat. Can. J. Microbiol. 1994, 40, 637–644. [Google Scholar] [CrossRef]
- Muslim, A.; Horinouchi, H.; Hyakumachi, M. Biological control of Fusarium wilt of tomato with hypovirulent binucleate Rhizoctonia in greenhouse conditions. Mycoscience 2003, 44, 77–84. [Google Scholar] [CrossRef]
- Yamagiwa, Y.; Toyoda, K.; Inagaki, Y.; Ichinose, Y.; Hyakumachi, M.; Shiraishi, T. Talaromyces wortmannii FS2 emits β-caryophyllene, which promotes plant growth and induces resistance. J. Gen. Plant Pathol. 2011, 77, 336–341. [Google Scholar] [CrossRef]
- Mushtaq, S.; Nasim, G.; Khokhar, I.; Mukhtar, I. Effects of Penicillium extracts on germination vigour in subsequent seedling growth of tomato (Solanum lycopersicum L.). Arch. Phytopathol. Plant Prot. 2012, 45, 932–937. [Google Scholar] [CrossRef]
- Nagaraju, A.; Sudisha, J.; Mahadeva Murthy, S.; Ito, S.I. Seed priming with Trichoderma harzianum isolates enhances plant growth and induces resistance against Plasmopara halstedii, an incitant of sunflower downy mildew disease. Australas. Plant Pathol. 2012, 41, 609–620. [Google Scholar] [CrossRef]
- Jogaiah, S.; Abdelrahman, M.; Tran, L.S.P.; Shin-ichi, I. Characterization of rhizosphere fungi that mediate resistance in tomato against bacterial wilt disease. J. Exp. Bot. 2013, 64, 3829–3842. [Google Scholar] [CrossRef] [Green Version]
- Sindhu, G.M.; Murali, M.; Thriveni, M.C.; Anupama, N.; Amruthesh, K.N. Growth promotion and disease resistance in muskmelon induced by crude proteins of Penicillium verruculosum against gummy stem blight disease. J. Crop. Sci. 2018, 10, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Basavaraj, G.L.; Murali, M.; Lavanya, S.N.; Amruthesh, K.N. Seed priming with biotic agents invokes defense response and enhances plant growth in pearl millet upon infection with Magnaporthe grisea. Biocatal. Agric. Biotechnol. 2019, 21, 101279. [Google Scholar] [CrossRef]
- Mastouri, F.; Björkman, T.; Harman, G.E. Seed treatment with Trichoderma harzianum alleviates biotic, abiotic, and physiological stresses in germinating seeds and seedlings. Phytopathology 2010, 100, 1213–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.M.; Sultana, F.; Miyazawa, M.; Hyakumachi, M. The plant growth-promoting fungus Penicillium spp. GP15-1 enhances growth and confers protection against damping-off and anthracnose in the cucumber. J. Oleo Sci. 2014, 63, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Islam, S.; Akanda, A.M.; Sultana, F.; Hossain, M.M. Chilli rhizosphere fungus Aspergillus spp. PPA1 promotes vegetative growth of cucumber (Cucumis sativus) plants upon root colonisation. Arch. Phytopathol. Plant Prot. 2014, 47, 1231–1238. [Google Scholar] [CrossRef]
- Khan, A.L.; Waqas, M.; Lee, I.J. Resilience of Penicillium resedanum LK6 and exogenous gibberellin in improving Capsicum annuum growth under abiotic stresses. J. Plant Res. 2015, 128, 259–268. [Google Scholar] [CrossRef]
- Shivanna, M.B.; Meera, M.S.; Hyakumachi, M. Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat. Crop. Prot. 1996, 15, 497–504. [Google Scholar] [CrossRef]
- Rabeendran, N.; Moot, D.J.; Jones, E.E.; Stewart, A. Inconsistent growth promotion of cabbage and lettuce from Trichoderma isolates. N. Z. Plant Prot. 2000, 53, 143–146. [Google Scholar] [CrossRef] [Green Version]
- Salas-Marina, M.A.; Silva-Flores, M.A.; Cervantes-Badillo, M.G.; Rosales-Saavedra, M.T.; Islas-Osuna, M.A.; Casas-Flores, S. The plant growth-promoting fungus Aspergillus ustus promotes growth and induces resistance against different lifestyle pathogens in Arabidopsis thaliana. J. Microbiol. Biotechnol. 2011, 21, 686–696. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Gan, Y.; Xu, B. Application of plant-growth-promoting fungi Trichoderma longibrachiatumT6 enhances tolerance of wheat to salt stress through improvement of antioxidative defense system and gene expression. Front. Plant Sci. 2016, 7, 1405. [Google Scholar]
- Halifu, S.; Deng, X.; Song, X.; Song, R. Effects of two Trichoderma strains on plant growth, rhizosphere soil nutrients, and fungal community of Pinus sylvestris var. mongolica annual seedlings. Forests 2019, 10, 758. [Google Scholar] [CrossRef] [Green Version]
- Zavala-Gonzalez, E.A.; Rodríguez-Cazorla, E.; Escudero, N.; Aranda-Martinez, A.; Martinez-Laborda, A.; Ramírez-Lepe, M.; Vera, A.; Lopez-Llorca, L.V. Arabidopsis thaliana root colonization by the nematophagous fungus Pochoniachlamydosporia is modulated by jasmonate signaling and leads to accelerated flowering and improved yield. New Phytol. 2017, 213, 351–364. [Google Scholar] [CrossRef] [Green Version]
- Poveda, J.; Hermosa, R.; Monte, E.; Nicolas, C. Trichoderma harzianum favours the access of arbuscular mycorrhizal fungi to non-host Brassicaceae roots and increases plant productivity. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual, C.B.; Raymundo, A.D.; Hayakumachi, M. Efficacy of hypovirulent binucleate Rhizoctonia sp. to control banded leaf and sheath blight in corn. J. Gen. Plant Pathol. 2000, 66, 95–102. [Google Scholar] [CrossRef]
- Desender, S.; Andrivon, D.; Florence, V. Activation of defence reactions in Solanaceae: Where is the specificity? -Microreview. Cell. Microbiol. 2007, 9, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, P.K.; Shenoy, B.D.; Gupta, M.; Vaish, A.; Mannan, S.; Singh, N.; Tewari, S.K.; Tripathi, R.D. Stimulatory effects of arsenic-tolerant soil fungi on plant growth promotion and soil properties. Microbes Environ. 2012, 27, 477–482, ME11316. [Google Scholar] [CrossRef] [Green Version]
- Akhter, W.; Bhuiyan, M.K.A.; Sultana, F.; Hossain, M.M. Integrated effect of microbial antagonist, organic amendment and fungicide in controlling seedling mortality (Rhizoctonia solani) and improving yield in pea (Pisum sativum L.). Comptes Rendus Biol. 2015, 338, 21–28. [Google Scholar] [CrossRef]
- Hammad, R.; Elbagory, M. Using plant growth-promoting fungi (PGPF), as a biofertilizer and biocontrol agents against Tetranychuscucurbitacearum on Nubian watermelon (Citrullus lanatus L.). J. Adv. Microbiol. 2019, 16, 1–15. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N.; Malnoy, M.; Maffei, M.E. Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 2015, 6, 151. [Google Scholar] [CrossRef] [Green Version]
- Hung, R.; Lee, S.; Bennett, J.W. Arabidopsis thaliana as a model system for testing the effect of Trichoderma volatile organic compounds. Fungal Ecol. 2013, 6, 19–26. [Google Scholar] [CrossRef]
- Naznin, H.A.; Kiyohara, D.; Kimura, M.; Miyazawa, M.; Shimizu, M.; Hyakumachi, M. Systemic resistance induced by volatile organic compounds emitted by plant growth-promoting fungi in Arabidopsis thaliana. PLoS ONE 2014, 9, e86882. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Sarma, B.K.; Upadhyay, R.S.; Singh, H.B. Compatible rhizosphere microbes mediated alleviation of biotic stress in chickpea through enhanced antioxidant and phenylpropanoid activities. Microbiol. Res. 2013, 168, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Doni, F.; Anizan, I.; Radziah, C.C.; Salman, A.H.; Rodzihan, M.H.; Yusoff, W.M.W. Enhancement of rice seed germination and vigour by “Trichoderma” spp. Res. J. Appl. Sci. Eng. Technol. 2014, 7, 4547–4552. [Google Scholar] [CrossRef]
- Zhang, F.; Huo, Y.; Cobb, A.B.; Luo, G.; Zhou, J.; Yang, G.; Wilson, G.W.; Zhang, Y. Trichoderma biofertilizer links to altered soil chemistry, altered microbial communities, and improved grassland biomass. Front. Microbiol. 2018, 9, 848. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, D.L.; Baker, R. Effect of certain fungi on dwarf tomatoes grown under gnotobiotic conditions. Phytopathol. 1967, 57, 1262–1263. [Google Scholar]
- Windham, M.T.; Elad, Y.; Baker, R. A mechanism for increased plant induced by Trichoderma spp. Phytopathology 1986, 76, 518–521. [Google Scholar] [CrossRef]
- Chang, Y.C.; Baker, R.; Kleifeld, O.; Chet, I. Increased growth of plants in the presence of the biological control agent Trichoderma harzianum. Plant Dis. 1986, 70, 145–148. [Google Scholar] [CrossRef]
- Narita, Y.; Suzuki, T. Influence of a sterile dark mycelial fungus on take-all of wheat. Annu. Soc. Jpn. 1991, 57, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.E.; Petzoldt, R.; Comis, A.; Chen, J. Interactions between Trichoderma harzianum strain T22 and maize inbred line Mo17 and effects of this interaction on diseases caused by Pythium ultimum and Colletotrichum graminicola. Phytopathology 2004, 94, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Chuang, C.C.; Kuo, Y.L.; Chao, C.C.; Chao, W.L. Solubilization of inorganic phosphates and plant growth promotion by Aspergillus niger. Biol. Fertil. Soils. 2007, 43, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Wakelin, S.A.; Gupta, V.V.S.R.; Harvey, P.R.; Ryder, M.H. The effect of Penicillium fungi on plant growth and phosphorus mobilization in neutral to alkaline soils from southern Australia. Can. J. Microbiol. 2007, 53, 106–115. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sultana, F.; Kubota, M.; Koyama, H.; Hyakumachi, M. The plant growth-promoting fungus Penicillium simplicissimum GP17-2 induces resistance in Arabidopsis thaliana by activation of multiple defense signals. Plant Cell Physiol. 2007, 48, 1724–1736. [Google Scholar] [CrossRef] [Green Version]
- Horinouchi, H.; Katsuyama, N.; Taguchi, Y.; Hyakumachi, M. Control of Fusarium crown and root rot of tomato in a soil system by combination of a plant growth-promoting fungus, Fusarium equiseti, and biodegradable pots. Crop. Prot. 2008, 27, 859–864. [Google Scholar] [CrossRef]
- Khan, S.A.; Hamayun, M.; Yoon, H.; Kim, H.Y.; Suh, S.J.; Hwang, S.K.; Kim, J.M.; Lee, I.J.; Choo, Y.S.; Yoon, U.H.; et al. Plant growth promotion and Penicillium citrinum. BMC Microbiol. 2008, 8, 231. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Cornejo, H.A.; Macias-Rodriguez, L.I.; Cortes-Penagos, C.; Lopez-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin dependent mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef] [Green Version]
- Chandanie, W.A.; Kubota, M.; Hyakumachi, M. Interactions between the arbuscular mycorrhizal fungus Glomus mosseae and plant growth promoting fungi and their significance for enhancing plant growth and suppressing damping-off of cucumber (Cucumis sativus L.). Appl. Soil Ecol. 2009, 41, 336–341. [Google Scholar] [CrossRef]
- Hamayun, M.; Khan, S.A.; Khan, M.A.; Khan, A.L.; Kang, S.M.; Kim, S.K.; Joo, G.J.; Lee, I.J. Gibberellin production by pure cultures of a new strain of Aspergillus fumigates. World J. Microbiol. Biotechnol. 2009, 25, 1785–1792. [Google Scholar] [CrossRef]
- Yadav, R.L.; Shukla, S.K.; Suman, A.; Singh, P.N. Trichoderma inoculation and trash management effects on soil microbial biomass, soil respiration, nutrient uptake and yield of ratoon sugarcane under subtropical conditions. Biol. Fertil. Soils 2009, 45, 461–468. [Google Scholar] [CrossRef]
- Horinouchi, H.; Muslim, A.; Hyakumachi, M. Biocontrol of Fusarium wilt of spinach by the plant growth promoting fungus Fusarium equiseti GF183. J. Plant Pathol. 2010, 92, 249–254. [Google Scholar]
- Shanmugaiah, V.; Mathivanan, N.; Varghese, B. Purification, crystal structure and antimicrobial activity of phenazine-1-carboxamide produced by a growth-promoting biocontrol bacterium, Pseudomonas aeruginosa MML2212. J. Appl. Microbiol. 2010, 108, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Saldajeno, M.G.B.; Hyakumachi, M. The plant growth-promoting fungus Fusarium equiseti and the arbuscular mycorrhizal fungus Glomus mosseae stimulate plant growth and reduce severity of anthracnose and damping off diseases in cucumber (Cucumis sativus) seedlings. Ann. Appl. Biol. 2011, 159, 28–40. [Google Scholar] [CrossRef]
- Kaveh, H.; Vatandoost, S.; Aroiee, H.; Mazhabi, M. Would Trichoderma affect seed germination and seedling quality of two muskmelon cultivars, Khatooni and Qasri and increase their transplanting success? J. Biol. Environ. Sci. 2011, 5, 169–175. [Google Scholar]
- Minerdi, D.; Bossi, S.; Maffei, M.E.; Gullino, M.L.; Garibaldi, A. Fusarium oxysporum and its bacterial consortium promote lettuce growth and expansin A5 gene expression through microbial volatile organic compound (MVOC) emission. FEMS Microbiol. Ecol. 2011, 76, 342–351. [Google Scholar] [CrossRef] [Green Version]
- Elsharkawy, M.M.; Shimizu, M.; Takahashi, H.; Hyakumachi, M. The plant growth-promoting fungus Fusarium equiseti and the arbuscular mycorrhizal fungus Glomus mosseae induce systemic resistance against Cucumber mosaic virus in cucumber plants. Plant Soil 2012, 361, 397–409. [Google Scholar] [CrossRef]
- Fontenelle, A.D.B.; Guzzo, S.D.; Lucon, C.M.M.; Harakava, R. Growth promotion and induction of resistance in tomato plant against Xanthomonas euvesicatoria and Alternaria solani by Trichoderma spp. Crop. Prot. 2011, 30, 1492–1500. [Google Scholar] [CrossRef]
- Sofo, A.; Tataranni, G.; Xiloyannis, C.; Dichio, B.; Scopa, A. Direct effects of Trichoderma harzianum strain T-22 on micropropagated shoots of GiSeLa6® Prunus cerasus × Prunus canescens) rootstock. Environ. Exp Bot. 2012, 76, 33–38. [Google Scholar] [CrossRef]
- Vujanovic, V.; Goh, Y.K. qPCR quantification of Sphaerodesmycoparasitica biotrophic mycoparasite interaction with Fusarium graminearum: In vitro and in planta assays. Arch. Microbiol. 2012, 194, 707–717. [Google Scholar] [CrossRef]
- Hossain, M.M.; Hossain, N.; Sultana, F.; Islam, S.M.N.; Islam, S.; Bhuiyan, M.K.A. Integrated management of Fusarium wilt of chickpea (Cicer arietinum L.) caused by Fusarium oxysporumf.sp. ciceris with microbial antagonist, botanical extract and fungicide. Afr. J. Biotechnol. 2013, 12, 4699–4706. [Google Scholar]
- Zhou, Z.; Zhang, C.; Zhou, W.; Li, W.; Chu, L.; Yan, J.; Li, H. Diversity and plant growth-promoting ability of endophytic fungi from the five flower plant species collected from Yunnan, Southwest China. J. Plant Interact. 2014, 9, 585–591. [Google Scholar] [CrossRef]
- Dixit, R.; Bahadur Singh, R.; Bahadur Singh, H. Screening of antagonistic potential and plant growth promotion activities of Trichoderma spp. and fluorescent Pseudomonas spp. isolates against Sclerotinia sclerotiorum causing stem rot of French bean. Legume Res. 2015, 38, 375–381. [Google Scholar] [CrossRef]
- Zhang, F.; Meng, X.; Feng, C.; Ran, W.; Yu, G.; Zhang, Y.; Shen, Q. Hydrolytic amino acids employed as a novel organic nitrogen source for the preparation of PGPF-containing bio-organic fertilizer for plant growth promotion and characterization of substance transformation during BOF production. PLoS ONE 2016, 11, e0149447. [Google Scholar] [CrossRef] [Green Version]
- Prasad, M.R.; Vidyasagar, B.; Umadevi, G.; Triveni, S.; Koteswar Rao, S.R.; Damodara Chari, S.R. Isolation and Screening of Bacterial and Fungal Isolates for Plant Growth Promoting Properties from Tomato (Lycopersicon esculentum Mill.). Int. J. Curr. Microbiol. App. Sci. 2017, 6, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Elsharkawy, M.M.; Abass, J.M.; Kamel, S.M.; Hyakumachi, M. The plant growth promoting fungus Penicillium sp. GP16-2 enhances the growth and confers protection against Cucumber mosaic virus in tobacco. Virol. Sci. 2017, 1, 145–154. [Google Scholar]
- Yoo, S.J.; Shin, D.J.; Won, H.Y.; Song, J.; Sang, M.K. Aspergillus terreus JF27 promotes the growth of tomato plants and induces resistance against Pseudomonas syringae pv. tomato. Mycobiology 2018, 46, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.S.; Tang, K.; Guo, S.X. The plant growth-promoting fungus (PGPF) Alternaria sp. A13 markedly enhances Salvia miltiorrhiza root growth and active ingredient accumulation under greenhouse and field conditions. Int. J. Mol. Sci. 2018, 19, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdappa, P.; Kumar, S.M.; Lakshmi, M.J.; Upreti, K.K. Growth stimulation and induction of systemic resistance in tomato against early and late blight by Bacillus subtilis OTPB1 or Trichoderma harzianum OTPB3. Biol. Control 2013, 65, 109–117. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; El-Khateeb, N.M.M. Antifungal activity and resistance induction against Sclerotium cepivorum by plant growth-promoting fungi in onion plants. Egypt. J. Biol. Pest Control 2019, 29, 68. [Google Scholar] [CrossRef]
- Al-Ghazali, M.R.; Korshed, M.S.H.; Al-Azawi, S.K. Some observations on phosphorus solubilisation by aerobic microorganisms isolated from sediments of Al-Khair River, (Iraq). J. Biol. Sci. Res. 1986, 17, 157–171. [Google Scholar]
- Illmer, P.; Schinner, F. Solubilization of insoluble phosphates by microrganisms isolated from forest soils. Soil Biol. Biochem. 1992, 24, 389–395. [Google Scholar] [CrossRef]
- Altomare, C.; Norvell, W.A.; Bjorkman, T.; Harman, G.E. Solubilization of phosphates and micronutrients by the plant-growth-promoting and biocontrol fungus Trichoderma harzianum Rifai 1295-22. Appl. Environ. Microbiol. 1999, 65, 2926–2933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.; Das, N.; Kumar, B.; Rinu, K.; Trivedi, P. Phosphate solubilization by Penicillium spp. isolated from soil samples of Indian Himalayan region. World J. Microbiol. Biotechnol. 2008, 24, 97–102. [Google Scholar] [CrossRef]
- Murali, M.; Thriveni, M.C.; Manjula, S.; Mythrashree, S.R.; Amruthesh, K.N. Isolation of phosphate solubilizing fungi from rhizosphere soil and its effect on seed growth parameters of different crop plants. J Appl. Biol. Biotechnol. 2016, 4, 022–026. [Google Scholar]
- Khan, M.S.; Zaidi, A.; Ahemad, M.; Oves, M.; Wani, P.A. Plant growth promotion by phosphate solubilizing fungi–current perspective. Arch. Agron. Soil Sci. 2010, 56, 73–98. [Google Scholar] [CrossRef]
- Shivanna, M.B.; Meera, M.S.; Kageyama, K.; Hyakumachi, M. Influence of zoysiagrass rhizosphere fungal isolates on growth and yield of soybean plants. Mycoscience 1995, 36, 25–30. [Google Scholar] [CrossRef]
- Reddy, M.S.; Kumar, S.K.; Babita, M.S.R. Bio-solubilization of poorly soluble rock phosphates by Aspergillus tubingensis and Aspergillus niger. Bioresour. Technol. 2002, 84, 187–189. [Google Scholar] [CrossRef]
- Li, Z.; Bai, T.; Dai, L.; Wang, F.; Tao, J.; Meng, S.; Hu, Y.; Wang, S.; Hu, S. A study of organic acid production in contrasts between two phosphate solubilizing fungi: Penicillium oxalicum and Aspergillus niger. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef]
- Murali, M.; Amruthesh, K.N. Plant Growth-promoting Fungus Penicillium oxalicum enhances plant growth and induces resistance in pearl millet against downy mildew disease. J. Phytopathol. 2015, 163, 743–754. [Google Scholar] [CrossRef]
- Fiorentino, N.; Ventorino, V.; Woo, S.L.; Pepe, O.; De Rosa, A.; Gioia, L.; Romano, I.; Lombardi, N.; Napolitano, M.; Colla, G.; et al. Trichoderma-based biostimulants modulate rhizosphere microbial populations and improve N uptake efficiency, yield, and nutritional quality of leafy vegetables. Front. Plant Sci. 2018, 9, 743. [Google Scholar] [CrossRef] [Green Version]
- Yedidia, I.; Srivastva, A.K.; Kapulnik, Y.; Chet, I. Effect of Trichoderma harzianum on microelement concentrations and increased growth of cucumber plants. Plant Soil 2001, 235, 235–242. [Google Scholar] [CrossRef]
- Singh, V.; Singh, P.N.; Yadav, R.L.; Awasthi, S.K.; Joshi, B.B.; Singh, R.K.; Lal, R.J.; Duttamajumder, S.K. Increasing the efficacy of Trichoderma harzianum for nutrient uptake and control of red rot in sugarcane. J. Hortic. For. 2010, 2, 66–71. [Google Scholar]
- Gizaw, B.; Tsegay, Z.; Tefera, G.; Aynalem, E.; Abatneh, E. Bio-Priming effect of three phosphate solubilizing fungi on seed germination and seedling development of Faba Bean (Vicia faba). J. Pest Biofertil. 2018, 1, 1–6. [Google Scholar]
- Rijavec, T.; Lapanje, A. Hydrogen cyanide in the rhizosphere: Not suppressing plant pathogens, but rather regulating availability of phosphate. Front. Microbiol. 2016, 7, 1785. [Google Scholar] [CrossRef] [Green Version]
- Berg, G. Plant-microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Crowley, D.E. Microbial siderophores in the plant rhizosphere. In Iron Nutrition in Plants and Rhizospheric Microorganisms; Springer: Dordrecht, The Netherlands, 2006; pp. 169–198. [Google Scholar]
- Wolfgang, H.; Volker, R.; Guenther, M. Fusarinines and dimerum acid, mono- and dihydroxamate siderophores from Penicillium chrysogenum, improve iron utilization by strategy I and strategy II plants. Biometals 2000, 13, 37–46. [Google Scholar]
- Machuca, A.; Milgres, A.M.F. Use of CAS agar plate modified to study the effects of different variables on the siderophore production by Aspergillus. Lett. Appl. Microbiol. 2003, 36, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Dhara, P.; Stuti, P.; Pooja, T.; Meenu, S. Siderophore producing Aspergillus spp. as bioinoculant for enhanced growth of Mung Bean. Int. J. Adv. Agric. Sci. Technol. 2017, 6, 111–120. [Google Scholar]
- Mahmoud, A.; Abd-Alla, M. Siderophore production by some microorganisms and their effect on Bradyrhizobium-Mung Bean symbiosis. Int. J Agri. Biol. 2001, 3, 157–162. [Google Scholar]
- Nenwani, V.; Doshi, P.; Saha, T.; Rajkumar, S. Isolation and characterization of a fungal isolate for phosphate solubilization and plant growth promoting activity. J. Yeast Fungal Res. 2010, 1, 009–014. [Google Scholar]
- Ghosh, S.K.; Banerjee, S.; Sengupta, C. Bioassay, characterization and estimation of siderophores from some important antagonistic Fungi. J. Biofertil. Biopestic. 2017, 10, 105–112. [Google Scholar]
- Pandya, N.D.; Desai, P.V.; Jadhav, H.P.; Sayyed, R.Z. Plant growth promoting potential of Aspergillus sp. NPF7, isolated from wheat rhizosphere in South Gujarat, India. Environ. Sustain. 2018, 1, 245–252. [Google Scholar] [CrossRef]
- Knowles, C.J.; Bunch, A.W. Microbial cyanide metabolism. Adv. Microb. Physiol. 1986, 27, 73–111. [Google Scholar]
- Curl, E.A.; Truelove, B. The Rhizosphere; Springer: Berlin, Germany, 1986. [Google Scholar]
- Samuel, S.; Muthukkaruppan, S.M. Characterization of plant growth promoting rhizobacteria and fungi associated with rice, mangrove and effluent contaminated soil. Curr. Bot. 2011, 2, 22–25. [Google Scholar]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; del-Val, E.; Larsen, J. Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere: Interactions with plants. FEMS Microbiol. Ecol. 2016, 92, fiw036. [Google Scholar] [CrossRef] [Green Version]
- Mohiddin, F.A.; Bashir, I.; Padder, S.A.; Hamid, B. Evaluation of different substrates for mass multiplication of Trichoderma species. J. Pharm. Phytochem. 2017, 6, 563–569. [Google Scholar]
- Benhamou, N.; Chet, I. Cellular and Molecular Mechanisms Involved in the Interaction between Trichoderma harzianum and Pythium ultimum. Appl. Environ. Microbiol. 1997, 63, 2095–2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajera, H.P.; Vakharia, D.N. Production of lytic enzymes by Trichoderma isolates during in vitro antagonism with Aspergillus niger, the causal agent of collar rot of peanut. Braz. J. Microbiol. 2012, 43, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Howell, C.R.; Stipanovic, R.D. Gliovirin, a new antibiotic from Gliocladium virens and its role in the biological control of Pythium ultimum. Can. J. Microbiol. 1983, 29, 321–324. [Google Scholar] [CrossRef]
- Kaur, R.; Kaur, J.; Singh, R.S. Non-pathogenic Fusarium as a biological control agent. Plant Pathol. J. 2011, 9, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Moreno, A.B.; Martinez del Pozo, A.; Borja, M.; Segundo, B.S. Activity of the antifungal protein from Aspergillus giganteus against Botrytis cinerea. Phytopathology 2003, 93, 1344–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toth, L.; Varadi, G.; Boros, E.; Borics, A.; Ficze, H.; Nagy, I.; Toth, G.K.; Rakhely, G.; Marx, F.; Galgoczy, L. Biofungicidal potential of Neosartorya (Aspergillus) fischeri antifungal protein NFAP and novel synthetic γ-core peptides. Front. Microbiol. 2020, 11, 820. [Google Scholar] [CrossRef]
- Bhatia, P.; Harsh, N.S.K.; Dubey, R.C.; Kaushik, P.K. Antagonistic activity of rhizosphere fungi against Fusarium solani causing wilt in Dalbergia sissoo. J. Sci. Res. Adv. 2016, 3, 352–357. [Google Scholar]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Barbetti, M.J.; Li, H.; Woo, S.L.; Lorito, M. A noel role for Trichoderma secondary metabolites in the interactions with plants. Physiol. Mol. Plant Pathol. 2008, 72, 80–86. [Google Scholar] [CrossRef]
- Hammerschmidt, R.; Kuc, J. Induced Resistance to Disease in Plants; Kluwer: Dordrecht, The Netherlands, 1995; p. 182. [Google Scholar]
- Choudhary, D.K.; Prakash, A.; Johri, B.N. Induced systemic resistance (ISR) in plants: Mechanism of action. Indian J. Microbiol. 2007, 47, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Montesinos, B.; Dianez, F.; Moreno-Gavira, A.; Gea, F.J.; Santos, M. Role of Trichoderma aggressivum f. europaeum as plant-growth promoter in horticulture. Agronomy 2020, 10, 1004. [Google Scholar]
- Houssien, A.A.; Ahmed, S.M.; Ismail, A.A. Activation of tomato plant defense response against Fusarium wilt disease using Trichoderma harzianum and salicylic acid under greenhouse conditions. Res. J. Agric. Biol. Sci. 2010, 6, 328–338. [Google Scholar]
- Heil, M. Induced systemic resistance (ISR) against pathogens- a promising field for ecological research. Perspect. Plant Ecol. Evol. SysteYedidia 2001, 4, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Koike, N.; Hyakumachi, M.; Kageyama, K.; Doke, N. Induction of systemic resistance in cucumber against several diseases by plant growth promoting fungi: Lignifications and superoxide generation. Eur. J. Plant Pathol. 2001, 107, 523–533. [Google Scholar] [CrossRef]
- Dong, H.Z.; Cohen, Y. Dry mycelium of Penicillium chrysogenum induces resistance against Verticillium wilt and enhances growth of cotton plants. Phytoparasitica 2002, 30, 147–157. [Google Scholar] [CrossRef]
- Hanson, L.E.; Howell, C.R. Elicitors of plant defense responses from biocontrol strains of Trichoderma virens. Phytopathology 2004, 94, 171–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segarra, G.; Casanova, E.; Bellido, D.; Odena, M.A.; Oliveira, E.; Trillas, I. Proteome, salicylic acid and jasmonic acid changes in cucumber plants inoculated with Trichoderma asperellum strain T34. Proteomics 2007, 7, 3943–3952. [Google Scholar] [CrossRef] [PubMed]
- Reglinski, T.; Rodenburg, N.; Taylor, J.T.; Northcott, G.L.; Chee, A.A.; Spiers, T.M.; Hill, R.A. Trichoderma atroviride promotes growth and enhances systemic resistance to Diplodiapinea in radiata pine (Pinus radiata) seedlings. For. Pathol. 2012, 42, 75–78. [Google Scholar] [CrossRef]
- Perazzolli, M.; Roatti, B.; Bozza, E.; Pertot, I. Trichoderma harzianum T39 induces resistance against downy mildew by priming for defense without costs for grapevine. Biol. Cont. 2011, 58, 74–82. [Google Scholar] [CrossRef]
- Yoshioka, Y.; Ichikawa, H.; Naznin, H.A.; Kogure, A.; Hyakumachi, M. Systemic resistance induced in Arabidopsis thaliana by Trichoderma asperellum SKT-1, a microbial pesticide of seedborne diseases of rice. Pest Manag. Sci. 2011, 68, 60–66. [Google Scholar] [CrossRef]
- Kakvan, N.; Heydari, A.; Zamanizadeh, H.R.; Rezaee, S.; Naraghi, L. Development of new bioformulations using Trichoderma and Talaromyces fungal antagonists for biological control of sugar beet damping-off disease. Crop. Prot. 2013, 53, 80–84. [Google Scholar] [CrossRef]
- Loganathan, M.; Sible, G.V.; Maruthasalam, S.; Saravanakumar, D.; Raguchander, T.; Sivakumar, M.; Samiyappan, R. Trichoderma and chitin mixture based bioformulation for the management of head rot (Sclerotinia sclerotiorum (Lib.) deBary)–root-knot (Meloidogyne incognitaKofoid and White; Chitwood) complex diseases of cabbage. Arch. Phytopathol. Plant Prot. 2010, 43, 1011–1024. [Google Scholar] [CrossRef]
- Konappa, N.; Krishnamurthy, S.; Siddaiah, C.N.; Ramachandrappa, N.S.; Chowdappa, S. Evaluation of biological efficacy of Trichoderma asperellum against tomato bacterial wilt caused by Ralstonia solanacearum. Egypt. J. Biol. Pest Control 2018, 28, 63. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, M.; Ryals, J. Systemic acquired resistance signal transduction. Crit. Rev. Plant Sci. 1996, 15, 583–606. [Google Scholar] [CrossRef]
- Neuenschwander, U.; Friedrich, L.; Delaney, T.; Vernooij, B.; Kessmann, H.; Ryals, J. Activation of plant disease resistance. Asp. Appl. Biol. 1995, 42, 217–225. [Google Scholar]
- Vernooij, B.; Friedrich, L.; Morse, A.; Reist, R.; Kolditz-Jawhar, R.; Ward, E.; Uknes, S.; Kessmann, H.; Ryals, J. Salicylic acid is not the translocated signal responsible for inducing systemic acquired resistance but is required in signal transduction. Plant Cell 1994, 6, 959–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezziyyani, M.; Hamdache, A.; Egea-Gilabert, C.; Requena, M.E.; Candela, M.E.; Mater, J. Production of Pathogenesis-Related proteins during the induction of resistance to Phytophthora capsici in pepper plants treated with Burkholderiacepacia and Trichoderma harzianum in combination compatible. J. Mater. Environ. Sci. 2017, 8, 4785–4795. [Google Scholar]
- Saxena, A.; Mishra, S.; Ray, S.; Raghuwanshi, R.; Singh, H.B. Differential reprogramming of defense network in Capsicum annum L. plants against Colletotrichum truncatum infection by phyllospheric and rhizosphericTrichoderma strains. J. Plant Growth Regul. 2019, 39, 751–763. [Google Scholar] [CrossRef]
- Wany, A.; Pathak, P.K.; Fernie, A.R.; Gupta, K.J. Trichoderma asperelloidesenhances local (LAR) and systemic acquired resistance (SAR) response under low nitrate nutrition in Arabidopsis. bioRxiv 2020. bioRxiv:502492. [Google Scholar]
- Alkooranee, J.T.; Yin, Y.; Aledan, T.R.; Jiang, Y.; Lu, G.; Wu, J.; Li, M. Systemic resistance to powdery mildew in Brassica napus (AACC) and Raphanus alboglabra (RRCC) by Trichoderma harzianum TH12. PLoS ONE 2015, 10, e0142177. [Google Scholar]
- Gomes, E.V.; Ulhoa, C.J.; Cardoza, R.E.; Silva, R.N.; Gutierrez, S. Involvement of Trichoderma harzianum Epl-1 protein in the regulation of Botrytis virulence-and tomato defense-related genes. Front. Plant Sci. 2017, 8, 880. [Google Scholar] [CrossRef] [PubMed]
- Patel, Z.M.; Mahapatra, R.; Jampala, S.S.M. Role of fungal elicitors in plant defense mechanism. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Academic Press: Cambridge, MA, USA, 2020; pp. 143–158. [Google Scholar]
- Bartnicki-Garcia, S. Chemistry of hyphal walls of Phytophthora. Microbiology 1966, 42, 57–69. [Google Scholar] [CrossRef] [Green Version]
- De Wit, P.J.G.M.; Hofman, A.E.; Velthuis, G.C.M.; Kuc, J.A. Isolation and characterization of an elicitor of necrosis isolated from intercellular fluids of compatible interactions of Cladosporium fulvum (syn. Fulvia fulva) and tomato. Plant Physiol. 1985, 77, 642–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, M.J.; Mainzer, J.; Dickerson, A.G. Characterization of a fungal glycoprotein that elicits a defense response in French bean. Physiol. Mol. Plant Pathol. 1992, 40, 333–351. [Google Scholar] [CrossRef]
- Velazhahan, R.; Samiyappan, R.; Vidhyasekaran, P. Purification of an elicitor-inducible antifungal chitinase from suspesion-cultured rice cells. Phytoparasitica 2000, 28, 131–139. [Google Scholar] [CrossRef]
- Osman, H.; Vauthrin, S.; Mikes, V.; Milat, M.L.; Panabières, F.; Marais, A.; Brunie, S.; Maume, B.; Ponchet, M.; Blein, J.P. Mediation of elicitin activity on tobacco is assumed by elicitin-sterol complexes. Mol. Biol. Cell 2001, 12, 2825–2834. [Google Scholar] [CrossRef] [Green Version]
- Djonovic, S.; Vargas, W.A.; Kolomiets, M.V.; Horndeski, M.; Wiest, A.; Kenerley, C.M.A. Proteinaceous elicitor Sm1 from the beneficial fungus Trichoderma virens is required for induced systemic resistance in maize. Plant Physiol. 2007, 145, 875–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Wang, W.Q.; Zhang, G.L.; Kaminek, M.; Dobrev, P.; Xu, J.; Gruissem, W. Senescence-inducible expression of isopentenyl transferase extends leaf life, increases drought stress resistance and alters cytokinin metabolism in cassava. J. Integr. Plant Biol. 2010, 52, 653–669. [Google Scholar] [CrossRef] [PubMed]
- Miao, F.; Yang, R.; Chen, D.; Wang, Y.; Qin, B.; Yang, X.; Zhou, L. Isolation, identification and antimicrobial activities of two secondary metabolites of Talaromycesverruculosus. Molecules 2012, 17, 14091–14098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishimoto, K.; Kouzai, Y.; Kaku, H.; Shibuya, N.; Minami, E.; Nishizawa, Y. Perception of the chitin oligosaccharides contributes to disease resistance to blast fungus Magnaportheoryzae in rice. Plant J. 2010, 64, 343–354. [Google Scholar] [CrossRef]
- Al-Waily, D.S.; Hassan, S.M. Effect of two bioformulation Trichoderma harzianum and Pseudomonas fluorescens with Manure in controlling Fusarium wilt disease in Pumpkin. J. Univ. Babylon Pure Appl. Sci. 2019, 27, 446–456. [Google Scholar]
- Darvill, A.G.; Albersheim, P. Phytoalexins and their elicitors: A defense against microbial infection in plants. Ann. Rev. Plant Physiol. 1984, 35, 243–275. [Google Scholar] [CrossRef]
- Sequeira, L. Mechanisms of induced resistance in plants. Ann. Rev. Microbiol. 1983, 37, 51–79. [Google Scholar] [CrossRef]
- Heath, M.C. Hypersensitive response-related death. Plant Mol. Biol. 2000, 44, 321–334. [Google Scholar] [CrossRef]
- Lamb, C.; Dixon, R.A. The oxidative burst in plant disease resistance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 251–275. [Google Scholar] [CrossRef]
- Kogel, K.H.; Beckhove, U.; Dreschers, J.; Munch, S.; Romme, Y. Acquired-resistance in barley- the resistance mechanism induced by 2,6-dichloroisonicotinic acid is a phenocopy of a genetically based mechanism governing race-specific powdery mildew resistance. Plant Physiol. 1994, 106, 1269–1277. [Google Scholar] [CrossRef] [Green Version]
- Soylu, E.M.; Soylu, S.; Mansfield, J.W. Ultrastructural characterisation of pathogen development and host responses during compatible and incompatible interactions between Arabidopsis thaliana and Peronospora parasitica. Physiol. Mol. Plant Pathol. 2004, 65, 67–78. [Google Scholar] [CrossRef]
- Yedidia, I.; Benhamou, N.; Chet, I. Induction of defense responses in cucumber plants (Cucumis sativus L.) by the biocontrol agent Trichoderma harzianum. Appl. Environ. Microbiol. 1999, 65, 1061–1070. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Ramirez, A.; Poveda, J.; Martin, I.; Hermosa, R.; Monte, E.; Nicolas, C. Trichoderma harzianum root colonization in Arabidopsis. Bio-Protocol 2015, 5, e1512. [Google Scholar]
- Siddaiah, C.N.; Satyanarayana, N.R.; Mudili, V.; Gupta, V.K.; Gurunathan, S.; Rangappa, S.; Huntrike, S.S.; Srivastava, R.K. Elicitation of resistance and associated defense responses in Trichoderma hamatum induced protection against pearl millet downy mildew pathogen. Sci. Rep. 2017, 7, 43991. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, R.L.; Hammerschmidt, R. Phenolic compounds and their role in disease resistance. Annu. Rev. Phytopathol. 1992, 30, 369–389. [Google Scholar] [CrossRef]
- Shimizu, K.; Hossain, M.M.; Kato, K.; Kubota, M.; Hyakumachi, M. Induction of defense responses in cucumber plants by using the cell-free filtrate of the plant growth-promoting fungus Penicillium simplicissimumGP17-2. J. Oleo Sci. 2013, 62, 613–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsharkawy, M.M.; Shivanna, M.B.; Meera, M.S. and Hyakumachi, M. Mechanism of induced systemic resistance against anthracnose disease in cucumber by plant growth-promoting fungi. Acta Agric. Scand. B Soil Plant Sci. 2015, 65, 287–299. [Google Scholar]
- Muslim, A.; Hyakumachi, M.; Kageyama, K.; Suwandi, S. Induction of systemic resistance in cucumber by hypovirulent binucleate Rhizoctonia against anthracnose caused by Colletotrichum orbiculare. Trop. Life Sci. Res. 2019, 30, 109. [Google Scholar] [CrossRef]
- Wojtaszek, P. Oxidative burst: An early plant response to pathogen infection. Biochem. J. 1997, 322, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Saxena, A.; Raghuwanshi, R.; Singh, H.B. Elevation of defense network in chilli against Colletotrichum capsici by phyllosphericTrichoderma strain. J. Plant Growth Regul. 2016, 35, 377–389. [Google Scholar] [CrossRef]
- Thakker, J.N.; Patel, S.; Dhandhukia, P.C. Induction of Defense-Related Enzymes in Banana Plants: Effect of Live and Dead Pathogenic Strain of Fusarium oxysporum f. sp. cubense. Isrn Biotechnol. 2013, 2013, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, G.S.; Reddy, N.N.R.; Surekha, C. Induction of plant systemic resistance in Legumes Cajanus cajan, Vigna radiata, Vigna mungo against plant pathogens Fusarium oxysporum and Alternaria alternataTrichodermaviride mediated reprogramming of plant defense mechanism. Int. J. Recent Sci. Res. 2015, 6, 4270–4280. [Google Scholar]
- Ronseaux, S.; Clement, C.; Barka, E. Interaction of Ulocladiumatrum, a potential biological control agent with Botrytis cinerea and grapevine plantlets. Agron J. 2013, 3, 632–647. [Google Scholar]
- Alizadeh, H.; Behboudi, K.; Ahmadzadeh, M.; Javan-Nikkhah, M.; Zamioudis, C.; Pieterse, C.M.J.; Bakker, P.A.H.M. Induced systemic resistance in cucumber and Arabidopsis thaliana by the combination of Trichoderma harzianum Tr6 and Pseudomonas sp. Ps14. Biol. Control 2013, 65, 14–23. [Google Scholar] [CrossRef]
- Hassan, N.; Elsharkawy, M.M.; Shivanna, M.B.; Meera, M.S.; Hyakumachi, M. Elevated expression of hydrolases, oxidase, and lyase in susceptible and resistant cucumber cultivars systemically induced with plant growth-promoting fungi against anthracnose. Acta Agric. Scand. B Soil Plant Sci. 2014, 64, 155–164. [Google Scholar] [CrossRef]
- Abhayashree, M.S.; Murali, M.; Thriveni, M.C.; Sindhu, G.M.; Amruthesh, K.N. Crude oligosaccharides mediated resistance and Histo- chemical changes in Capsicum annuum against anthracnose disease caused by Colletotrichum capsici. Plant Biosys. 2016, 151, 221–233. [Google Scholar] [CrossRef]
- Wu, L.; Wu, H.; Chen, L.I.; Zhang, H.; Gao, X. Induction of systemic disease resistance in Nicotiana benthamiana by the cyclodipeptides cyclo (LPro- L-Pro) and cyclo (D-Pro-D-Pro). Mol. Plant Pathol. 2017, 18, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Bowles, D.J. Defense-related proteins in higher plants. Annu. Rev. Biochem. 1990, 59, 873–907. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting mechanisms of defence against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Koorneef, A.; Pieterse, C.M.J. Cross talk in defense signaling. Plant Physiol. 2008, 146, 839–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfano, G.; Ivey, M.L.; Cakir, C.; Bos, J.I.; Miller, S.A.; Madden, L.V.; Kamoun, S.; Hoitink, H.A. Systemic modulation of gene expression in tomato by Trichoderma hamatum. Phytopathology 2007, 97, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Shoresh, M.; Harman, G.E. The molecular basis of maize responses to Trichoderma harzianum T22 inoculation: A proteomic approach. Plant Physiol. 2008, 147, 2147–2163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, C.A.; Castillo, F.; Gonzalez, A.; Bernal, D.; Jaimes, Y.; Chaparro, M.; Gonzalez, C.; Rodriguez, F.; Restrepo, S.; Cotes, A.M. Biological and molecular characterization of the response of tomato plants treated with Trichoderma koningiopsis. Physiol. Mol. Plant Pathol. 2009, 74, 111–120. [Google Scholar] [CrossRef]
- Gusakov, A.V. Alternatives to Trichoderma reesei in biofuel production. Trends Biotechnol. 2011, 29, 419–425. [Google Scholar] [CrossRef]
- Hermosa, R.; Rubio, M.B.; Cardoza, R.E.; Nicolas, C.; Monte, E.; Gutierrez, S. The contribution of Trichoderma to balancing the costs of plant growth and defense. Int. Microbiol. 2013, 16, 69–80. [Google Scholar]
- Saravanakumar, K.; Fan, L.; Fu, K.; Yu, C.; Wang, M.; Xia, H.; Sun, J.; Li, Y.; Chen, J. Cellulase from Trichoderma harzianum interacts with roots and triggers induced systemic resistance to foliar disease in maize. Sci. Rep. 2016, 10, 35543. [Google Scholar] [CrossRef]
- Al-Tuwaijri, M.M. Role of the biocontrol agents Trichoderma viride and Bacillus subtilis in elimination of the deteriorative effects of the root-rot pathogens Fusarium oxysporum and F. solani on some metabolic and enzyme activities of cucumber plants. Egypt J. Exp. Biol. 2009, 5, 29–35. [Google Scholar]
- Wang, J.; Li, J.; Li, J.; Li, J.; Liu, S.; Gao, W. LSP1, a responsive protein from Meyerozymaguilliermondii, elicits defence response and improves glycyrrhizic acid biosynthesis in Glycyrrhiza uralensis Fisch adventitious roots. J. Cell Physiol. 2017, 232, 3510–3519. [Google Scholar] [CrossRef] [PubMed]
- Hammerschmidt, R. Rapid deposition of lignin in potato tuber tissue as a response to fungi non-pathogenic on potato. Physiol. Plant Pathol. 1984, 24, 33–42. [Google Scholar] [CrossRef]
- Larkin, R.P.; Fravel, D.R. Mechanisms of action and dose-response relationships governing biological control of Fusarium wilt of tomato by non-pathogenic Fusarium spp. Phytopathology 1999, 89, 1152–1161. [Google Scholar] [CrossRef] [Green Version]
- He, C.Y.; Hsiang, T.; Wolyn, D.J. Induction of systemic disease resistance and pathogen defence responses in Asparagus officinalis inoculated with non-pathogenic strains of Fusarium oxysporum. Plant Pathol. 2002, 51, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Kroner, A.; Hamelin, G.; Andrivon, D.; Val, F. Quantitative resistance of potato to Pectobacteriumatrosepticum and Phytophthora infestans: Integrating PAMP-triggered response and pathogen growth. PLoS ONE 2011, 6, e23331. [Google Scholar] [CrossRef] [PubMed]
- Mathys, J.; De Cremer, K.; Timmermans, P.; Van Kerkhove, S.; Lievens, B.; Vanhaecke, M.; Cammue, B.; De Coninck, B. Genome-wide characterization of ISR induced in Arabidopsis thaliana by Trichoderma hamatum T382 against Botrytis cinerea infection. Front. Plant Sci. 2012, 3, 108. [Google Scholar] [CrossRef] [Green Version]
- Roy, C.K.; Akter, N.; Sarkar, M.K.; Pk, M.U.; Begum, N.; Zenat, E.A.; Jahan, M.A. Control of early blight of tomato caused by and screening of tomato varieties against the pathogen. Open Microbiol. J. 2019, 13, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Jin, N.; Liu, S.M.; Peng, H.; Huang, W.K.; Kong, L.A.; Wu, Y.H.; Chen, Y.P.; Ge, F.Y.; Jian, H.; Peng, D.L. Isolation and characterization of Aspergillus niger NBC001 underlying suppression against Heteroderaglycines. Sci. Rep. 2019, 9, 591. [Google Scholar] [CrossRef] [Green Version]
- He, M.; He, C.Q.; Ding, N.Z. Abiotic stresses: General defenses of land plants and chances for engineering multi stress tolerance. Front. Plant Sci. 2018, 9, 1771. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Montesinos, B.; Dianez, F.; Moreno-Gavira, A.; Gea, F.J.; Santos, M. Plant growth promotion and biocontrol of Pythium ultimum by saline tolerant Trichoderma isolates under salinity stress. Int. J. Environ. Res. Public Health 2019, 16, 2053. [Google Scholar] [CrossRef] [Green Version]
- Mantri, N.L.; Ford, R.; Coram, T.E.; Pang, E.C.K. Transcriptional profiling of chickpea genes differentially regulated in response to high-salinity, cold and drought. BMC Genom. 2007, 8, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tigchelaar, M.; Battisti, D.S.; Naylor, R.L.; Ray, D.K. Future warming increases probability of globally synchronized maize production shocks. Proc. Natl. Acad. Sci. USA 2018, 115, 6644–6649. [Google Scholar] [CrossRef] [Green Version]
- Qadir, M.; Quillerou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J.; Drechsel, P.; Noble, A.D. Economics of salt-induced land degradation and restoration. Nat. Res. Forum. 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Kang, S.; Baek, I.; Lee, I. Characterization of plant growth promoting traits of Penicillium species against the effects of high soil salinity and root disease. J. Plant Interact. 2014, 9, 754–762. [Google Scholar] [CrossRef]
- Qi, W.; Zhao, L. Study of the siderophore-producing Trichoderma asperellum Q1 on cucumber growth promotion under salt stress. J. Basic Microbiol. 2013, 53, 355–364. [Google Scholar] [CrossRef]
- Masunaka, A.; Hyakumachi, M.; Takenaka, S. Plant growth-promoting fungus, Trichoderma koningi suppresses isoflavonoid phytoalexin vestitol production for colonization on/in the roots of Lotus japonicus. Microbes Environ. 2011, 26, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, A.; Raghuwanshi, R.; Singh, H.B. Trichoderma species mediated differential tolerance against biotic stress of phytopathogens in Cicer arietinum L. J. Basic Microbiol. 2015, 55, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Guler, N.S.; Pehlivan, N.; Karaoglu, S.A.; Guzel, S.; Bozdeveci, A. Trichoderma atroviride ID20G inoculation ameliorates drought stress-induced damages by improving antioxidant defence in maize seedlings. Acta Physiol. Plant. 2016, 38, 132. [Google Scholar] [CrossRef]
- Ahmad, P.; Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; John, R.; Egamberdieva, D.; Gucel, S. Role of Trichoderma harzianum in mitigating NaCl stress in Indian mustard (Brassica juncea L) through antioxidative defense system. Front. Plant Sci. 2015, 6, 868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Li, Q.; Xiong, L.; Kronzucker, H.J.; Kramer, U.; Shi, W. Arabidopsis plastid AMOS1/EGY1 integrates abscisic acid signalling to regulate global gene expression response to ammonium stress. Plant Physiol. 2012, 160, 2040–2051. [Google Scholar] [CrossRef] [Green Version]
- Montero-Barrientos, M.; Hermosa, R.; Cardoza, R.E.; Gutierrez, S.; Nicolas, C.; Monte, E. Transgenic expression of the Trichoderma harzianum hsp70 gene increases Arabidopsis resistance to heat and other abiotic stresses. J. Plant Physiol. 2010, 167, 659–665. [Google Scholar] [CrossRef]
- Dixit, P.; Mukherjee, P.K.; Ramachandran, V.; Eapen, S. Glutathione transferase from Trichoderma virens enhances cadmium tolerance without enhancing its accumulation in transgenic Nicotiana tabacum. PLoS ONE 2011, 6, e16360. [Google Scholar] [CrossRef] [Green Version]
- Vieira, P.M.; Santos, M.P.; Andrade, C.M.; Souza-Neto, O.A.; Ulhoa, C.J.; Aragao, F.J.L. Overexpression of an aquaglyceroporin gene from Trichoderma harzianum improves water-use efficiency and drought tolerance in Nicotiana tabacum. Plant Physiol. Biochem. 2017, 121, 38–47. [Google Scholar] [CrossRef]
- Benitez, T.; Rincon, A.M.; Limon, M.C.; Codon, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar]
- Plemenitas, A.; Vaupotic, T.; Lenassi, M.; Kogej, T.; Gunde-Cimerman, N. Adaptation of extremely halotolerant black yeast Hortaeawerneckii to increased osmolarity: A molecular perspective at a glance. Stud. Mycol. 2008, 61, 67–75. [Google Scholar] [CrossRef]
- Nishimura, M.T.; Stein, M.; Hou, B.H.; Vogel, J.P.; Edwards, H.; Somerville, S.C. Loss of a callose synthase results in salicylic acid-dependent disease resistance. Science 2003, 301, 969–972. [Google Scholar] [CrossRef]
- Moura, J.C.M.S.; Bonine, C.A.V.; Viana, J.D.O.F.; Dornelas, M.C.; Mazzafera, P. Abiotic and biotic stresses and changes in the lignin content and composition in plants. J. Integr. Plant Biol. 2010, 52, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Turk, M.; Mejanelle, L.; Sentjurc, M.; Grimalt, J.O.; Gunde-Cimerman, N.; Plemenitas, A. Salt-induced changes in lipid composition and membrane fluidity of halophilic yeast-like melanized fungi. Extremophiles 2004, 8, 53–61. [Google Scholar] [CrossRef]
- Gostincar, C.; Turk, M.; Plemenitas, A.; Gunde-Cimerman, N. The expressions of Δ9-, Δ12-desaturases and an elongase by the extremely halotolerant black yeast Hortaeawerneckii are salt dependent. FEMS Yeast Res. 2009, 9, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Ruppel, S.; Franken, P.; Witzel, K. Properties of the halophyte microbiome and their implications for plant salt tolerance. Funct. Plant Biol. 2013, 40, 940–951. [Google Scholar] [CrossRef] [Green Version]
- Macia-Vicente, J.G.; Ferraro, V.; Burruano, S.; Lopez-Llorca, L.V. Fungal assemblages associated with roots of halophytic and non-halophytic plant species vary differentially along a salinity gradient. Microb. Ecol. 2012, 64, 668–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunde-Cimerman, N.; Zalar, P.; de Hoog, S.; Plemenitas, A. Hypersaline waters in salterns–natural ecological niches for halophilic black yeasts. FEMS Microbiol. Ecol. 2000, 32, 235–240. [Google Scholar]
- Tresner, H.D.; Hayes, J.A. Sodium chloride tolerance of terrestrial fungi. Appl. Microbiol. 1971, 22, 210–213. [Google Scholar] [CrossRef]
- Rodriguez, R.; Redman, R. More than 400 million years of evolution and some plants still can’t make it on their own: Plant stress tolerance via fungal symbiosis. J. Exp. Bot. 2008, 59, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Chibuike, G.U.; Obiora, S.C. Heavy metal polluted soils: Effect on plants and bioremediation methods. Appl. Environ. Soil Sci. 2014, 2014, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kacprzak, M.J.; Rosikon, K.; Fijalkowski, K.; Grobelak, A. The effect of Trichoderma on heavy metal mobility and uptake by Miscanthus giganteus, Salix sp., Phalaris arundinacea, and Panicum virgatum. Appl. Environ. Soil Sci. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ayad, F.; Matallah-Boutiba, A.; Rouane-Hacene, O.; Bouderbala, M.; Boutiba, Z. Tolerance of Trichoderma sp. to heavy metals and its antifungal activity in algerian marine environment. J. Mar. Biol. Oceanogr. 2018, 7, 2. [Google Scholar] [CrossRef]
- Rasool, S.; Ahmad, A.; Siddiqi, T.O.; Ahmad, P. Changes in growth, lipid peroxidation and some key antioxidant enzymes in chickpea genotypes under salt stress. Acta Physiol. Plant. 2013, 35, 1039–1050. [Google Scholar] [CrossRef]
- Ortiz-Castro, R.; Martinez-Trujillo, M.; Lopez-Bucio, J. N-acyl-L-homoserine lactones: A class of bacterial quorum-sensing signals alter post-embryonic root development in Arabidopsis thaliana. Plant Cell Environ. 2008, 31, 1497–1509. [Google Scholar] [CrossRef]
- Zhang, S.W.; Gan, Y.T.; Xu, B.L. Mechanisms of the IAA and ACC deaminase producing strain of Trichoderma longibrachiatum T6 in enhancing wheat seedling tolerance to NaCl stress. BMC Plant Biol. 2019, 19, 22. [Google Scholar] [CrossRef] [Green Version]
- Bjorkman, T.; Blanchard, L.M.; Harman, G.E. Growth enhancement of shrunken-2 (sh2) sweet corn by Trichoderma harzianum 1295-22: Effect of environmental stress. J. Am. Soc. Hortic. Sci. 1998, 123, 35–40. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Kim, Y.H.; Kang, S.M.; Lee, J.H.; Lee, I.N. Gibberellins producing endophytic Aspergillus fumigatus sp. LH02 influenced endogenous phytohormonal levels, isoflavonoids production and plant growth in salinity stress. Process Biochem. 2011, 46, 440–447. [Google Scholar] [CrossRef]
- Mastouri, F.; Bjorkman, T.; Harman, G.E. Trichoderma harzianum enhances antioxidant defense of tomato seedlings and resistance to water deficit. Mol. Plant Microbe 2012, 25, 1264–1271. [Google Scholar] [CrossRef] [Green Version]
- Gusain, Y.S.; Singh, U.S.; Sharma, A.K. Enhance activity of stress related enzymes in rice (Oryza sativa L.) induced by plant growth promoting fungi under drought stress. Afr. J. Agr. Res. 2014, 9, 1430–1434. [Google Scholar]
- Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; Al Huqail, A.A.; Egamberdieva, D. Alleviation of abiotic salt stress in Ochradenusbaccatus (Del.) by Trichoderma hamatum (Bonord.) Bainier. J. Plant Interact. 2014, 9, 857–868. [Google Scholar] [CrossRef]
- Chen, S.C.; Ren, J.J.; Zhao, H.J.; Wang, X.L.; Wang, T.H.; Jin, S.D.; Wang, Z.H.; Li, C.Y.; Liu, A.R.; Lin, X.M.; et al. Trichoderma harzianum improves defense against Fusarium oxysporum by Regulating ROS and RNS Metabolism, redox balance and energy flow in cucumber roots. Phytopathology 2019, 109, 972–982. [Google Scholar] [CrossRef]
- Pang, Z.; Zhao, Y.; Xu, P.; Yu, D. Microbial Diversity of Upland Rice Roots and Their Influence on Rice Growth and Drought Tolerance. Microorganisms 2020, 8, 1329. [Google Scholar] [CrossRef]
- Savci, S. An agricultural pollutant: Chemical fertilizer. Int. J. Environ. Sci. Dev. 2012, 3, 73. [Google Scholar] [CrossRef] [Green Version]
- Gowtham, H.G.; Singh, B.; Murali, M.; Shilpa, N.; Prasad, M.; Aiyaz, M.; Amruthesh, K.N.; Niranjana, S.R. Induction of drought tolerance in tomato upon the application of ACC deaminase producing plant growth promoting rhizobacterium Bacillus subtilis Rhizo SF 48. Microbiol. Res. 2020, 234, 126422. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| PGPF | Plant | Effect | Reference |
|---|---|---|---|
| R. nigricans, F. roseum | Solanum lycopersicum | Enhanced shoot dry weight | [56] |
| T. harzianum, T. koningii | Solanum lycopersicum and Nicotiana tabacum | Enhanced dry weight and improved seed germination | [57] |
| T. ciride | Solanum lycopersicum | Maximized plant height | |
| T. harzianum | Vinca minor | Improved flowering, weight and height of the plant | [58] |
| Sterile dark fungus (SDF) | T. aestivum | Increased shoot dry weight | [59] |
| Penicillium simplicissimum (GP17-2) | Cucumis sativus | Increased root and shoot growth | [4] |
| T. harzianum (T22) | Zea mays | Increased shoot growth, root area and root size | [60] |
| A. niger | B. chinensis | Increased plant dry weight and N, P content | [61] |
| Penicillium sp. | Triticum aestivum | Solubilized phosphate | [62] |
| Penicillium janthinellum and Penicillium simplicissimum | Arabidopsis thaliana | Elevated shoot biomass and leaf number | [63] |
| Fusarium equiseti (GF19-1) | Lycopersicon lycopersicum | Increased plant biomass with more root and shoot growth | [64] |
| Penicillium citrinum | Suaeda japonica | Increased root and shoot length | [65] |
| Trichoderma virens | Arabidopsis thaliana | Improved biomass and lateral roots development with the production of IAA | [66] |
| Trichoderma harzianum (GT3-2) | Cucumis sativus | Increased root and shoot growth | [67] |
| Phoma herbarum and Aspergillus fumigatus | Glycine max | Increased plant height and plant biomass. Maximized shoot growth, leaf area and chlorophyll content | [68] |
| Trichoderma viride | Saccharum officinarum | Improved crop yield | [69] |
| Fusarium equiseti | Spinacia oleracea | Improved overall plant biomass by maximizing root and shoot growth | [70] |
| Trichoderma viride | Gossypium arboreum | Increased root, shoot length and plant dry weight | [71] |
| Fusarium equiseti (GF19-1) | Cucumis sativus | Increased root and shoot growth | [72] |
| Trichoderma harzianum | Cucumis melo | Induced early seed germination and increased seedling vigor | [73] |
| Fusarium oxysporum(MSA-35) | Lactuca sativus | Increased root and shoot growth with high chlorophyll content | [74] |
| Penicillium simplicissimum | A. thaliana and N. tabacum | Maximized shoot fresh weight, shoot dry weight and leaf number | [75] |
| Trichoderma spp. | Lycopersicon lycopersicum | Increased dry matter biomass and improved overall plant growth | [76] |
| Trichoderma harzianum(T-22) | Prunus cerasus × P. canescens | Increased root growth and development | [77] |
| Sphaerodes mycoparasitica | Triticum aestivum | Increased seed germination and seedling vigour | [78] |
| T. harzianum (T-75) | Cicer arietinum | Increased crop yield | [79] |
| Alternaria sp., Phomopsis sp., Cladosporium sp., Colletotrichum sp., Phoma sp., Aspergillus sp. and | Nicotiana tabacum | Improved overall plant growth biomass by maximizing the root and shoot growth and high chlorophyll, soluble sugar content | [80] |
| Trichoderma sp. | Phaseolus vulgaris | Positive for plant growth promoting traits, i.e., phosphate, siderophore, HCN and Ammonia | [81] |
| Trichoderma longibrachiatum | Triticum aestivum | Increased plant height, root length, shoot fresh and dry weights. Increased chlorophyll a, b and total chlorophyll content. | [82] |
| Trichoderma sp. | Lycopersicon esculentum | Produced IAA, siderophore, HCN, ammonia and solubilized phosphate | [83] |
| Penicillium spp. GP16-2 | N. tabacum | Enhanced shoot fresh, dry weight and increased the leaf number | [84] |
| Aspergillus terreus JF27 | Lycopersicon esculentum | Enhanced fresh weight and shoot length | [85] |
| Alternaria sp. | Salvia miltiorrhiza | Enhanced fresh weight and dry weight | [86] |
| T. harzianum | Lycopersicon esculentum | Increased root and shoot growth. Maximized leaf area and vigour of tomato seedlings. Elicited the production of IAA and GA. | [87] |
| T. viride and T. harzianum | Citrullus lanatus | Increased the number of leaves, leaf dry weight, stem length and the number of branches. Enhanced chlorophyll content and N, P, K uptake. Also enhanced the fruit number, seeds number, fruit weight and dry weight. | [49] |
| Phoma sp.GS 8-3, Trichoderma asperellum SKT-1, Fusarium equiseti GF18-3 and Penicillium simplicissmum GP17-2 | Allium cepa | Enhanced the plant height, root length, bulb perimeter and plant dry weight. | [88] |
| PGPF | Plant | Effect | Reference |
|---|---|---|---|
| C. cucumerinum and F. roseum | S. tuberosum | Increased lignin deposition | [194] |
| Trichoderma harzianum (T-203) | Cucumis sativus | Enhanced callose deposition | [153] |
| F. oxysporum | Lycopersicon esculentum | Induced resistance by reducing the incidence of Fusarium wilt disease | [195] |
| F. oxysporum | Asparagus officinalis Lycopersicon esculentum | Exhibited HR response, increased defense related enzymes activity PO, PAL, lignin content and reduced the disease severity upon pathogen infection | [196] |
| Pectobacterium atrosepticum and P. infestans | Solanum tuberosum | Elicitors obtained from the culture filtrates induced high levels of phenolic compound and PAL enzyme activity | [197] |
| Trichoderma sp., | Cucumis sativus, Arabidopsis thaliana. | Triggered SA and JA/ET pathways in eliciting defense in plants | [198] |
| Penicillium spp. | P. glaucaum | Enhanced protection against downy mildew pathogen | [25] |
| Trichoderma harzianum | Cucumis sativus and Arabidopsis thaliana | Induced systemic resistance in cucumber and A. thaliana with defense related genes expression | [164] |
| Phoma sp., Penicillium sp., Trichoderma sp., Aspergillus sp., Fusarium sp. | Lycopersicon esculentum | Enhanced protection by reduction of disease against R. solanacearum with increasing the activity of defense related enzymes PAL, POX and GLU activities | [32] |
| Penicillium simplicissimum | Cucumis sativus | Induced protection against anthracnose disease showing a reduction in the lesion number and lesion diameter. Elevated the activity of exo-glucanase, exo-chitinase, PO and PPO | [171] |
| Phoma spp. | Cucumis sativus | Induced resistance by increasing the chitinase activity upon infection with Colletotrichum orbiculare | [180] |
| T. harzianum | Arabidopsis thaliana | Elicited callose deposition in the roots of seedlings | [168] |
| Trichoderma viride | Cajanus cajan, Vigna radiata and Vigna mungo | Elevated the levels of ROS and increased the levels of PO, PPO and PAL activities. Maximized SOD, CAT, AOX activities and total phenolics. Reduce the severity of diseases against Fusarium oxysporum and Alternaria alternata | [177] |
| Phoma sp. and non-sporulating fungus | Cucumis sativus | Induced protection against anthracnose disease under pot and field conditions. Inhibited the germination of C. orbiculare pathogen. Elevated the formation of lignin and expression of PAL, PO, PPO, GLU and CHI activities upon pathogen infection | [172] |
| Penicillium spp. GP16-2 | N. tabacum | Decreased disease severity of Cucumber Mosaic Virus (CMV) in tobacco plants | [75] |
| Trichoderma spp., T. viride, T. harzianum | Lycopersicon esculentum | Antagonize the growth of A. solani (early blight pathogen) with a reduction in the percent disease index | [199] |
| Rhizoctonia sp. | Cucumis sativus | Induced systemic resistance against C. orbiculare by reducing the total lesion number and total lesion diameter along with increasing the activity of POX enzyme | [173] |
| A. niger | Glycine max | Triggered callose deposition in leaves and roots upon infection with Heterodera glycines | [200] |
| PGPF | Plant | Effect | Reference |
|---|---|---|---|
| Trichoderma harzianum (1295-22) | Zea mays | Mitigated oxidative stress caused by sodium hypochlorite and lipid peroxidation in sweet corn | [233] |
| Trichoderma harzianum (T-22) | Lycopersicon lycopersicum | Improved seed germination under stress | [35] |
| Exophiala sp. LHL08 | Cucumis sativus | Improved growth under salinity and drought stresses | [234] |
| Trichoderma harzianum | Lycopersicon esculentum | Increased plant growth biomass and Induced resistance to water deficit. Enhanced APX, CAT and SOD activity. Improved the ability of plants damage caused by ROS | [235] |
| Trichoderma harrzianum and Fusarium pallidoroseum | Oryza sativa | Higher biomass production and increased induction of SOD, CAT and POD | [236] |
| Trichoderma hamatum | Ochradenus baccatus | Alleviated abiotic salt stress by improving plant growth and antioxidant defense enzyme activity | [237] |
| Penicillium sp. | Sesamum indicum | Increased root and shoot length, maximized fresh and dry weight of seedlings under salt stress. Increased amino acid, chlorophyll a,b and total chlorophyll content. Also enhanced protein and nitrogen content. Induced protection against Fusarium wilt disease | [206] |
| Trichoderma harzianum | Brassica juncea | Mitigated NaCl stress by enhancement of antioxidant defense machinery. Improved shoot, root length and plant dry weight | [211] |
| Trichoderma longibrachiatum | Triticum aestivum | Increased the tolerance of plants to salt stress by SOD, POD, CAT gene expression | [82] |
| Trichoderma atroviride | Zea mays | Ameliorated drought stress by enhancement of antioxidant defense in plant seedlings | [210] |
| Trichoderma harzianum | Cucumis sativus | Improved defense by alleviated oxidative and nitro-stative stress by minimizing ROS production and RNO species production upon infection with F. oxysporum by enhancing the antioxidant potential | [238] |
| Talaromyces sp., Penicillium sp., Mucor sp., Fusarium sp., Pestalotiopsis sp., Aspergillus sp., etc. | Oryza sativa | Improved the growth of plants and antioxidant capability, also, to increase in proline and soluble sugar content | [239] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murali, M.; Naziya, B.; Ansari, M.A.; Alomary, M.N.; AlYahya, S.; Almatroudi, A.; Thriveni, M.C.; Gowtham, H.G.; Singh, S.B.; Aiyaz, M.; et al. Bioprospecting of Rhizosphere-Resident Fungi: Their Role and Importance in Sustainable Agriculture. J. Fungi 2021, 7, 314. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040314
Murali M, Naziya B, Ansari MA, Alomary MN, AlYahya S, Almatroudi A, Thriveni MC, Gowtham HG, Singh SB, Aiyaz M, et al. Bioprospecting of Rhizosphere-Resident Fungi: Their Role and Importance in Sustainable Agriculture. Journal of Fungi. 2021; 7(4):314. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040314
Chicago/Turabian StyleMurali, Mahadevamurthy, Banu Naziya, Mohammad Azam Ansari, Mohammad N. Alomary, Sami AlYahya, Ahmad Almatroudi, M. C. Thriveni, Hittanahallikoppal Gajendramurthy Gowtham, Sudarshana Brijesh Singh, Mohammed Aiyaz, and et al. 2021. "Bioprospecting of Rhizosphere-Resident Fungi: Their Role and Importance in Sustainable Agriculture" Journal of Fungi 7, no. 4: 314. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040314