
Epichloë gansuensis Increases the Tolerance of Achnatherum inebrians to Low-P Stress by Modulating Amino Acids Metabolism and Phosphorus Utilization Efficiency

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Achnatherum Inebrians Seedlings Growth and the Different P Concentrations Treatment
2.2. The Dry Weight of Leaves and Roots in E+ and E− Achnatherum inebrians and Preparation of GS-MS
2.3. The Determination of C, N, P and Metal ion Contents of E+ and E− Achnatherum inebrians Plants
2.4. GC-MS Analysis
2.5. The Amino Acids Content of Leaves and Roots in E+ and E− Achnatherum inebrians
2.6. Calculation of Phosphorus Utilization Efficiency (PUE) of E+ and E− Achnatherum inebrians Plants
2.7. Statistical Analysis
3. Results
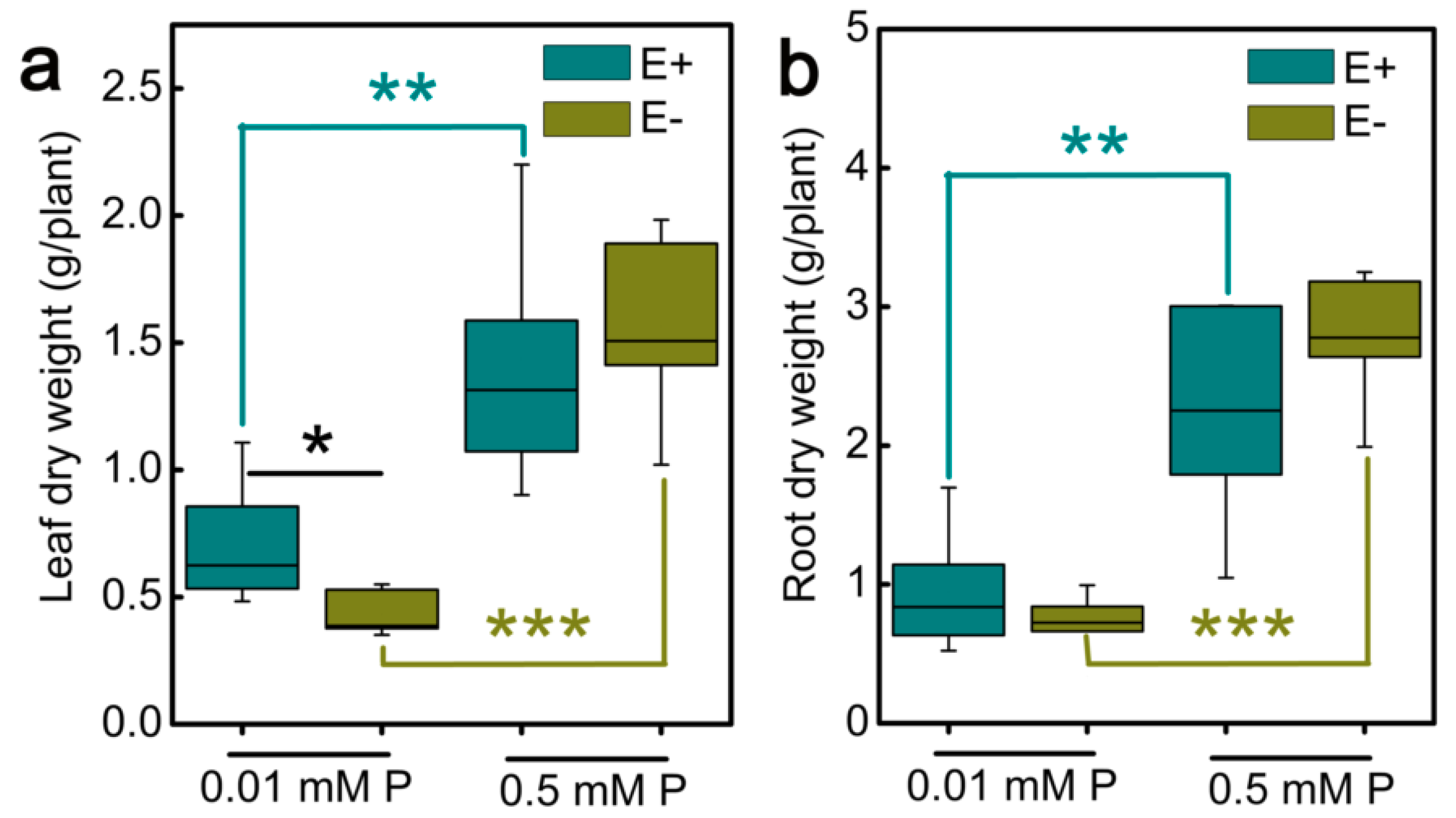
3.1. Effects of Epichloë gansuensis on the Dry Weight of Achnatherum inebrians Seedlings at Various P Concentration
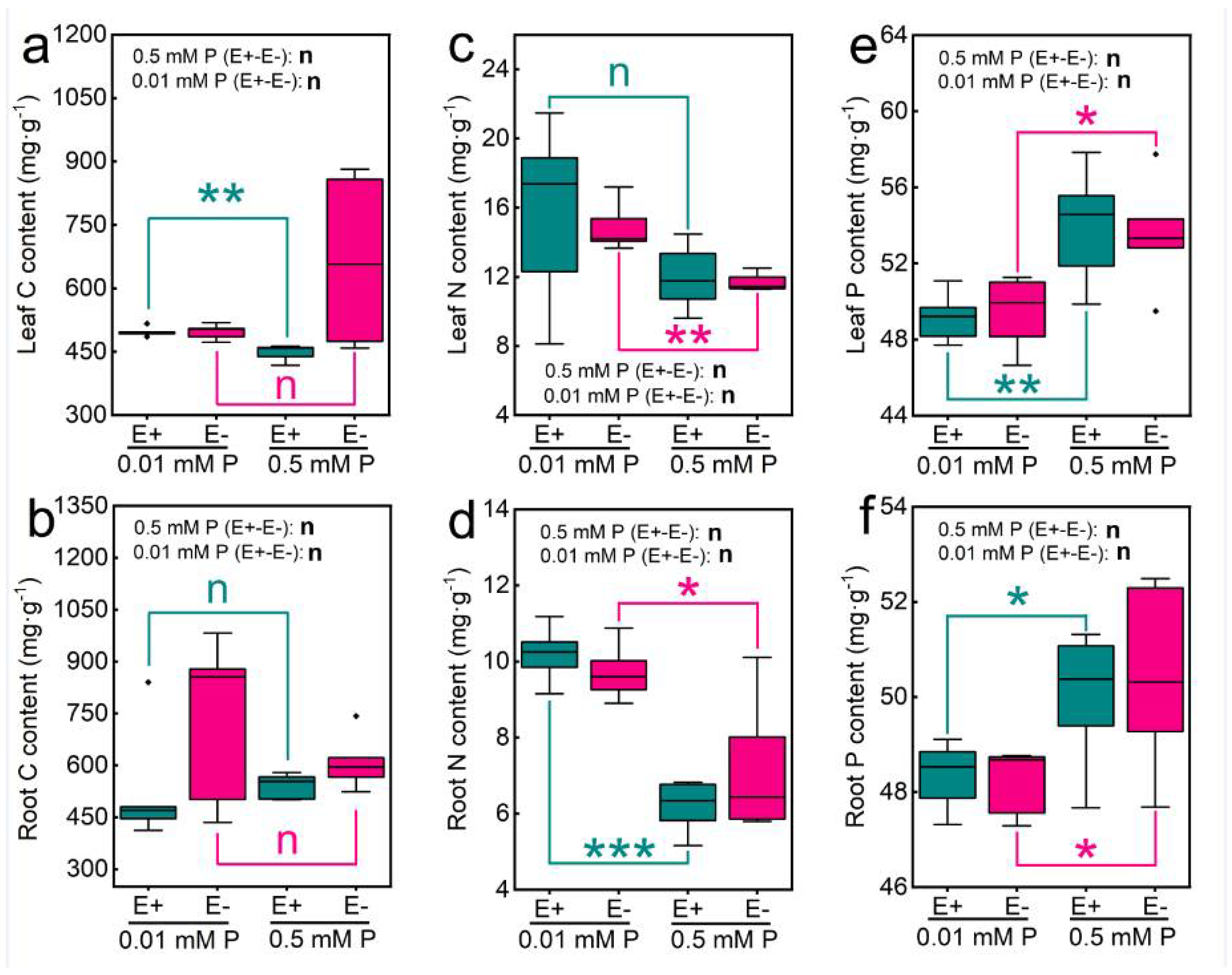
3.2. Effects of Epichloë gansuensis on C, N, and P Content of Achnatherum inebrians at Various P Concentration
3.3. Effects of Epichloë gansuensis on the K, Ca, Na, Mg and Fe Content of Achnatherum inebrians at Low P Stress
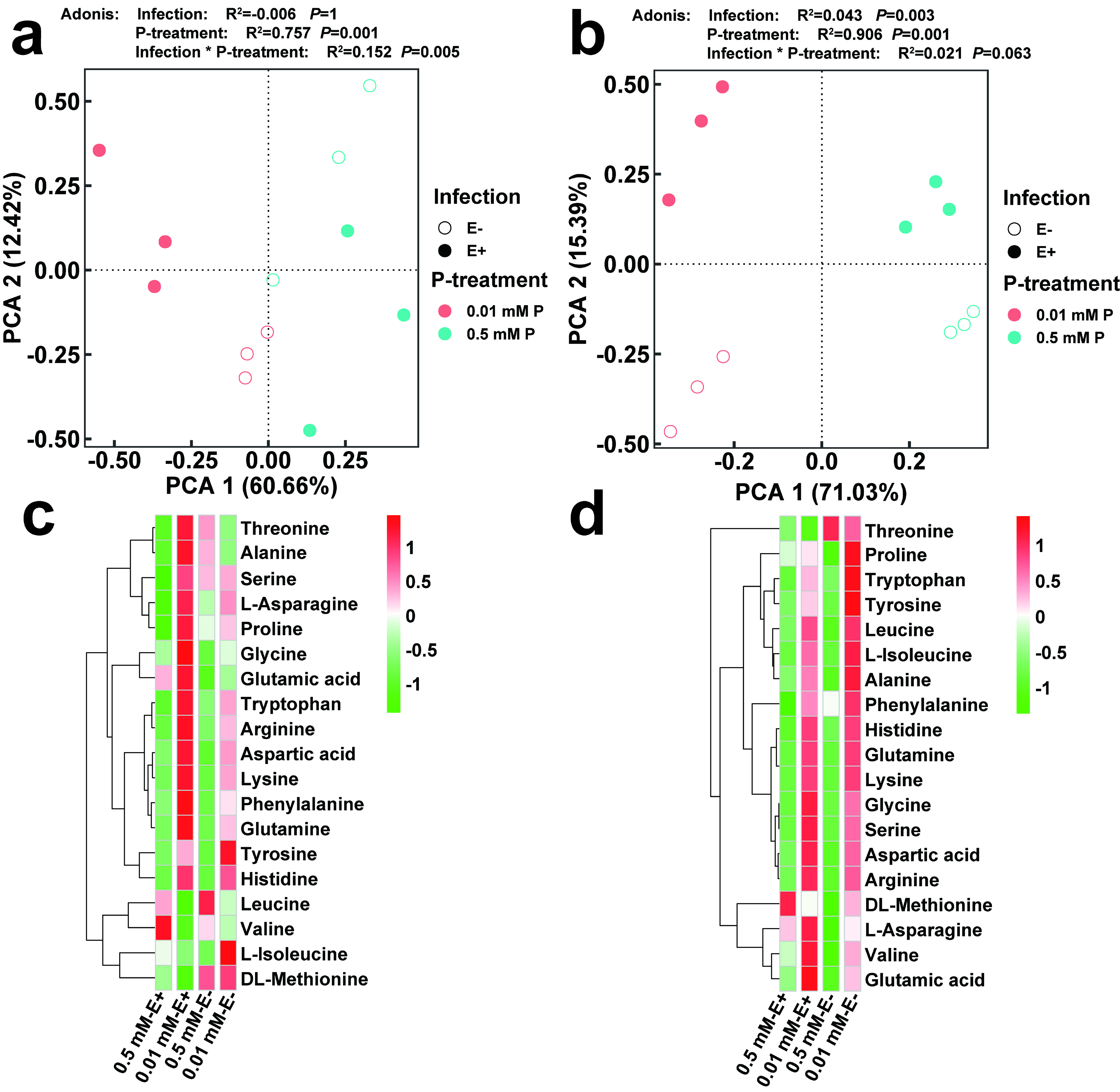
3.4. Metabolic Changes
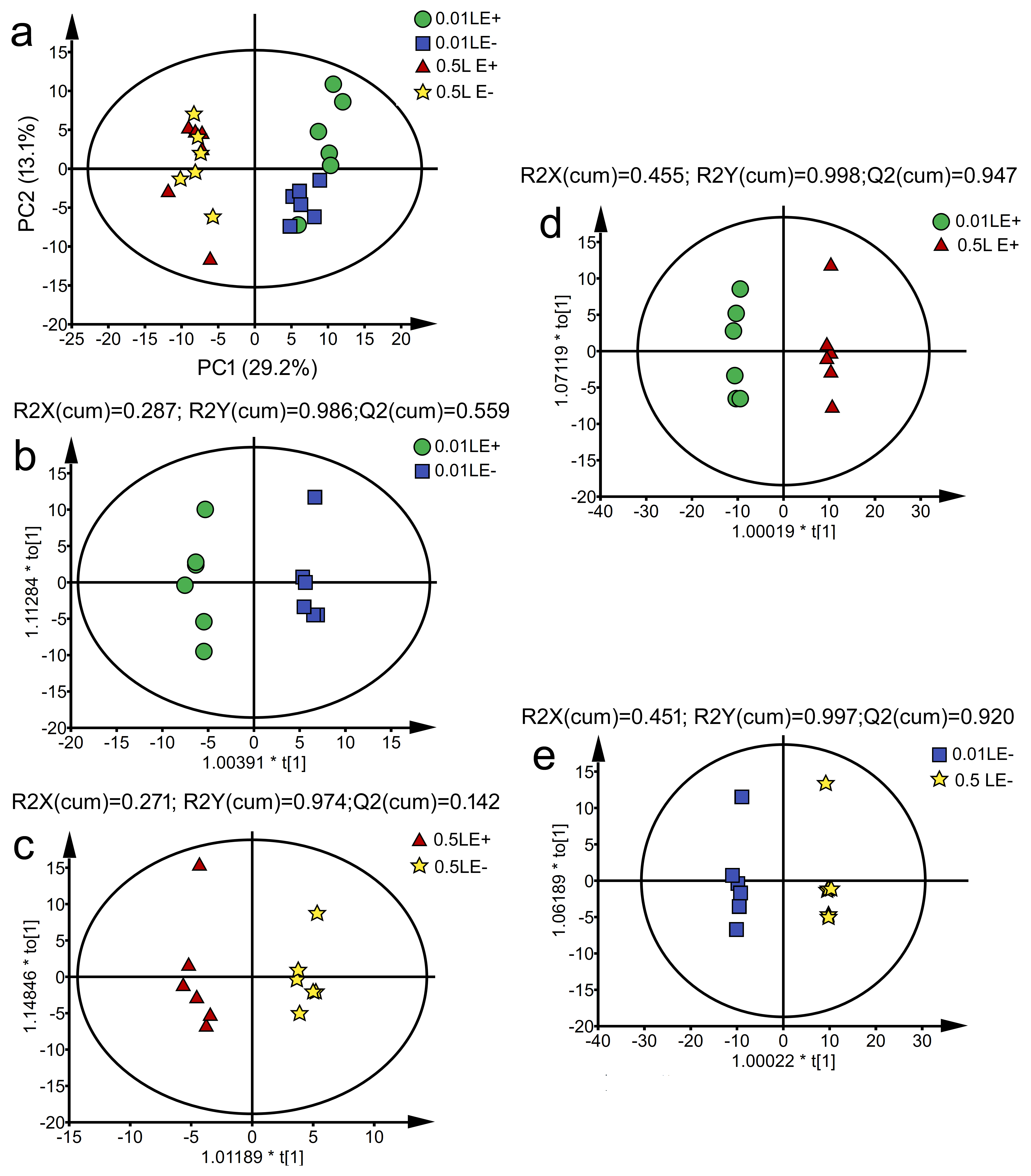
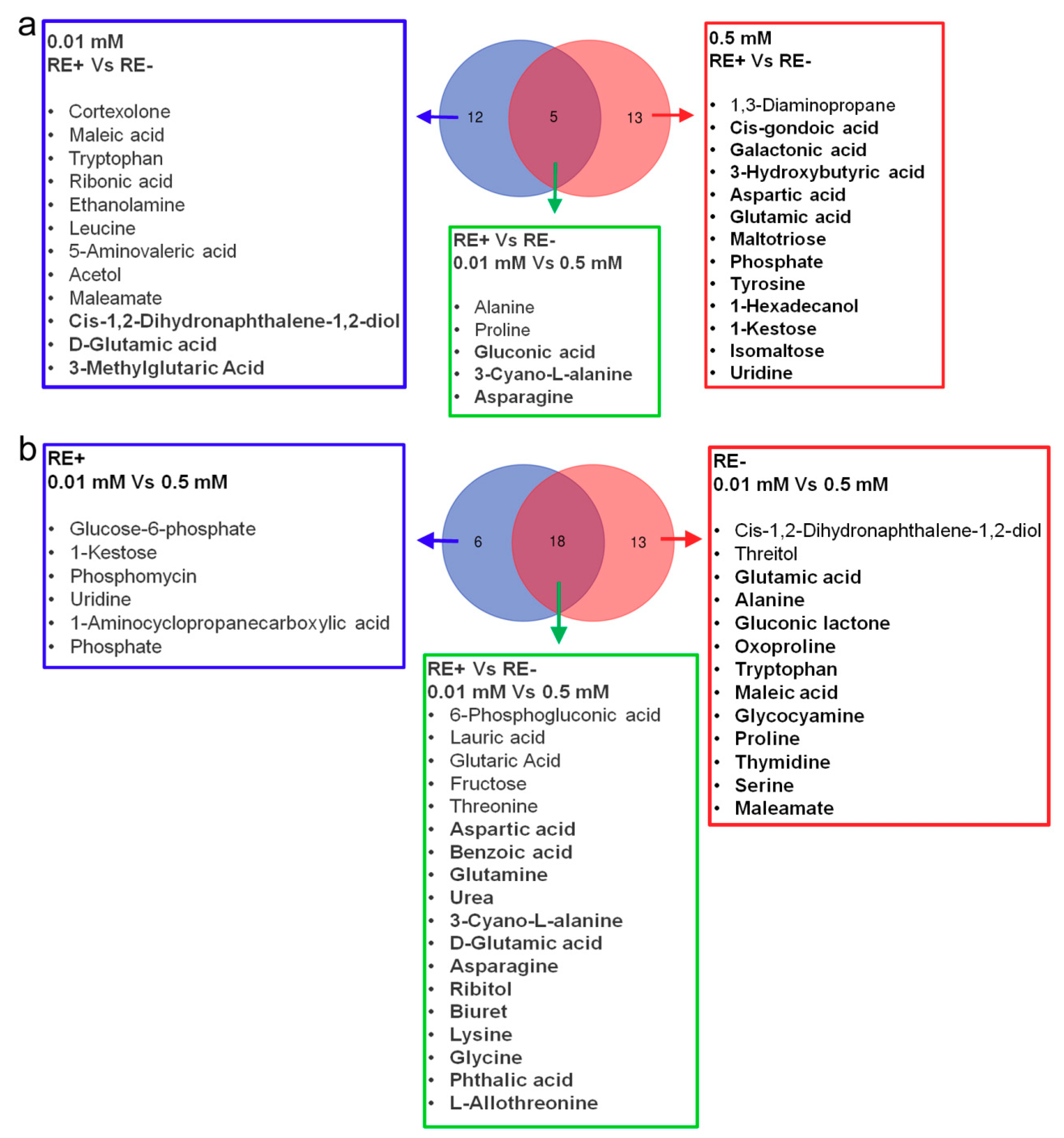
3.5. Effects of Epichloë gansuensis on Metabolic Profiles in the Leaves of Achnatherum inebrians at 0.01 and 0.5 mM P
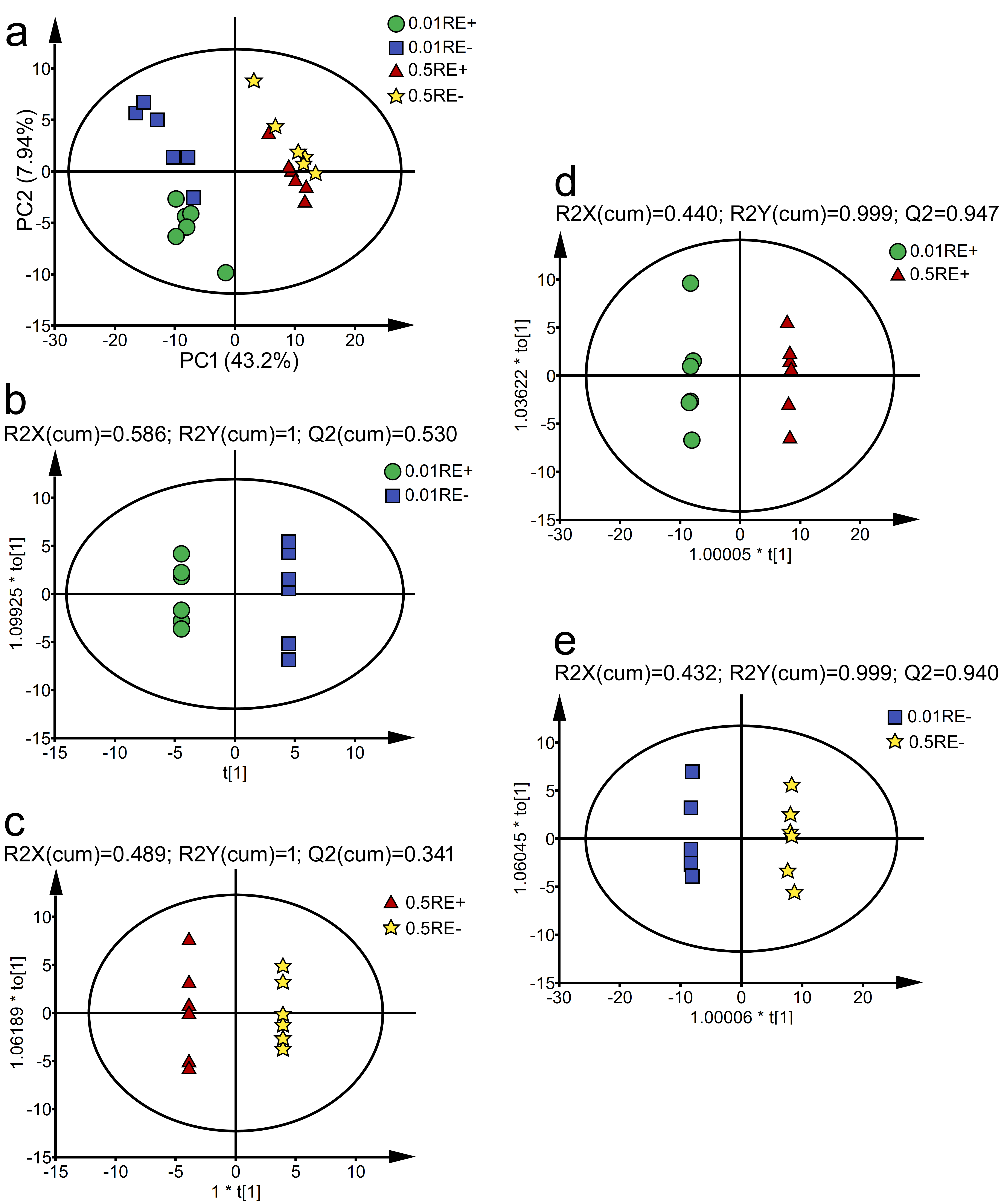
3.6. Effects of Epichloë gansuensis on Metabolic Profiles in the Roots of Achnatherum inebrians at 0.01 and 0.5 mM P
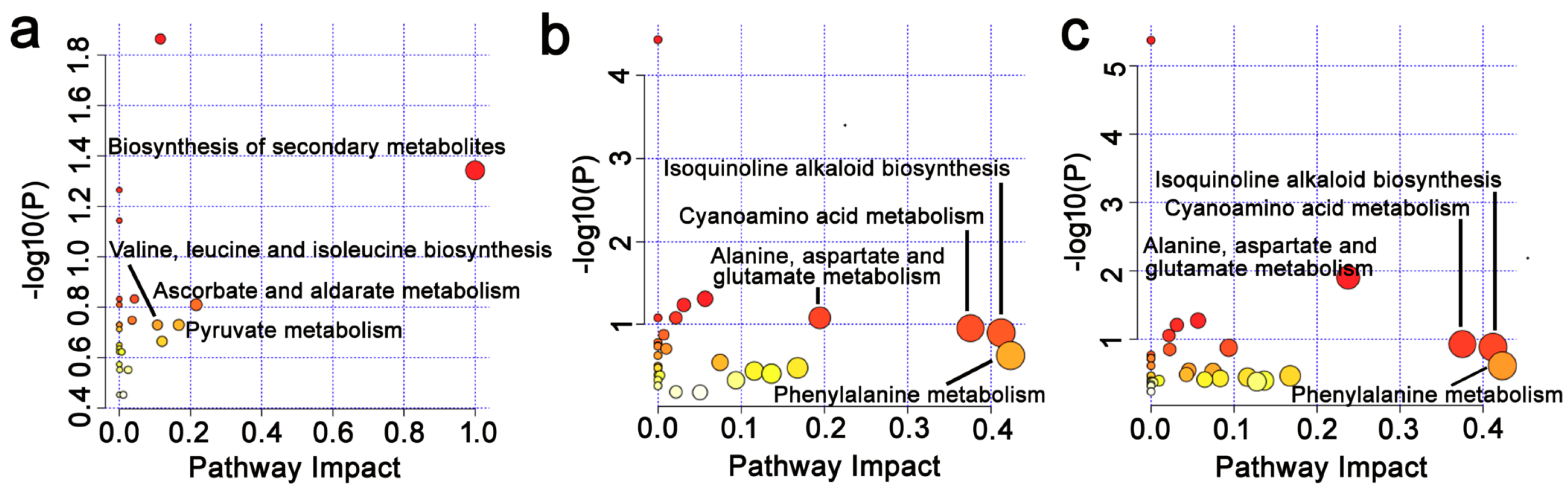
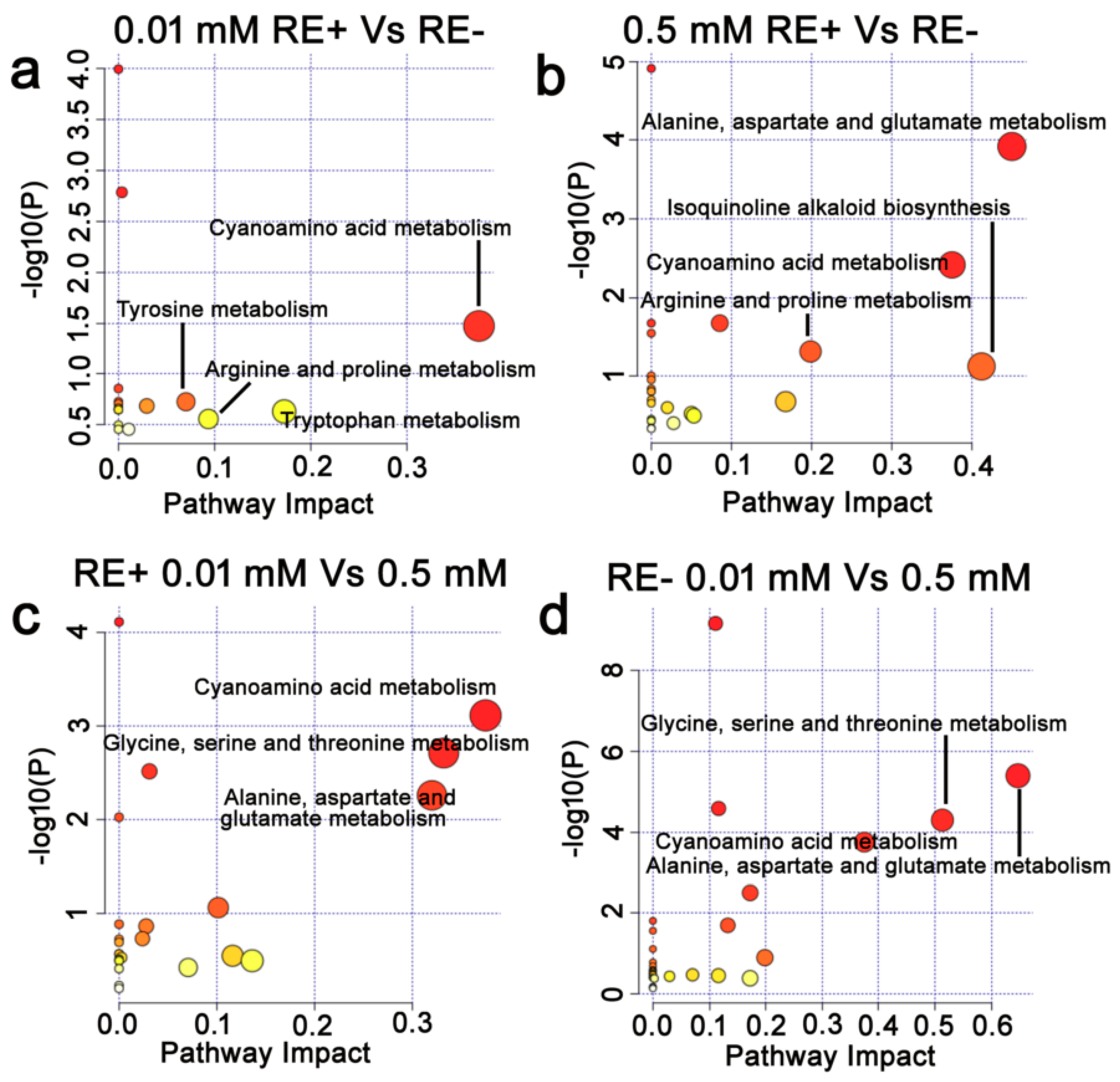
3.7. Effects of Epichloë gansuensis on Metabolic Pathways in the Leaves of Achnatherum inebrians at 0.01 and 0.5 mM P
3.8. Effects of Epichloë gansuensis on Metabolic Pathways in the Roots of Achnatherum inebrians at 0.01 and 0.5 mM P
3.9. The Changes of Amino Acids in Leaves and Roots of E+ and E− Achnatherum inebrians at Low P Stress
3.10. Effects of Epichloë gansuensis on Phosphorus Utilization Efficiency (PUE) of Achnatherum inebrians at Low P Stress
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Müller, J.; Gödde, V.; Niehaus, K.; Zörb, C. Metabolic adaptations of white lupin roots and shoots under phosphorus deficiency. Front. Plant Sci. 2015, 6, 1014. [Google Scholar] [CrossRef]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [PubMed]
- Mórrígan, T. Global Climate Change, Human Security, and Democracy. In Peak Phosphorus: A Potential Food Security Crisis. Global & International Studies; University of California: Oakland, CA, USA, 2010; Available online: http://climateproject.global.ucsb.edu/publications/pdf/Morrigan_2010_Peak%20Phosphorus.pdf (accessed on 14 May 2021).
- Hammond, J.P.; Broadley, M.R.; White, P.J.; King, G.J.; Bowen, H.C.; Hayden, R.; Meacham, M.C.; Mead, A.; Overs, T.; Spracklen, W.P.; et al. Shoot yield drives phosphorus use efficiency in Brassica oleracea and correlates with root architecture traits. J. Exp. Bot. 2009, 60, 1953–1968. [Google Scholar] [CrossRef] [PubMed]
- Koliaei, A.A.; Akbari, G.A.; Armandpisheh, O.; Labbafi, M.R.; Zarghami, R. Effects of phosphate chemical fertilizers and biologic fertilizers in various moisture regimes on some morphological characteristics and seeds performance in maize. Asian J. Agric. Food Sci. 2011, 3, 223–233. [Google Scholar]
- Reddy, M.S.; Kumar, S.; Babita, K. Biosolubilization of poorly soluble rock phosphates by Aspergillus tubingensis and Aspergillus niger. Bioresour. Technol. 2002, 84, 187–189. [Google Scholar] [CrossRef]
- Li, C.J.; Nan, Z.B.; Paul, V.H.; Dapprich, P.D.; Liu, Y. A new Neotyphodium species symbiotic with drunken horse grass (Achnatherum inebrians) in China. Mycotaxon 2004, 90, 141–147. [Google Scholar]
- Wang, J.; Hou, W.; Christensen, M.J.; Xia, C.; Chen, T.; Zhang, Z.; Nan, Z. The fungal endophyte Epichloë gansuensis increases NaCl-tolerance in Achnatherum inebrians through enhancing the activity of plasma membrane H+-ATPase and glucose-6-phosphate dehydrogenase. Sci. China Life Sci. 2020, 63, 1–14. [Google Scholar] [CrossRef]
- Wang, Z.; Li, C.; White, J. Effects of Epichloë endophyte infection on growth, physiological properties and seed germination of wild barley under saline conditions. J. Agron. Crop Sci. 2019, 206, 43–51. [Google Scholar] [CrossRef]
- Wang, J.; Nan, Z.; Christensen, M.J.; Zhang, X.; Tian, P.; Zhang, Z.; Niu, X.; Gao, P.; Chen, T.; Ma, L. Effect of Epichloë gansuensis endophyte on the nitrogen metabolism, nitrogen use efficiency, and stoichiometry of Achnatherum inebrians under nitrogen limitation. J. Agric. Food Chem. 2018, 66, 4022–4031. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.P.; Wang, J.F.; Nan, Z.B.; Michael, J.C.; Xia, C.; Chen, T.; Zhang, Z.X.; Niu, X.L. Epichloë gansuensis endophyte-infection alters soil enzymes activity and soil nutrients at different growth stages of Achnatherum inebrians. Plant Soil 2020, 55, 227–240. [Google Scholar] [CrossRef]
- Hou, W.; Xia, C.; Christensen, M.J.; Wang, J.; Li, X.; Chen, T.; Nan, Z. Effect of Epichloë gansuensis endophyte on rhizosphere bacterial communities and nutrient concentrations and ratios in the perennial grass species Achnatherum inebrians during three growth seasons. Crop Pasture Sci. 2020, 1, 1050–1066. [Google Scholar] [CrossRef]
- Yao, X.; Christensen, M.J.; Bao, G.; Zhang, C.; Li, X.; Li, C.; Nan, Z. A toxic endophyte-infected grass helps reverse degradation and loss of biodiversity of over-grazed grasslands in northwest China. Sci. Rep. 2015, 5, 18527. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, D.; Belesky, D.; Hill, N.; Baligar, V.; Fedders, J.M. Influence of phosphorus on the growth and ergot alkaloid content of Neotyphodium coenophialum-infected tall fescue (Festuca arundinacea Schreb.). Plant Soil 1998, 198, 53–61. [Google Scholar] [CrossRef]
- Chen, Z.; Jin, Y.; Yao, X.; Chen, T.; Wei, X.; Li, C.; White, J.F.; Nan, Z. Fungal endophyte improves survival of Lolium perenne in low fertility soils by increasing root growth, metabolic activity and absorption of nutrients. Plant Soil 2020, 452, 185–206. [Google Scholar] [CrossRef]
- Arbona, V.; Manzi, M.; de Ollas, C.; Gómez-Cadenas, A. Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef]
- Hong, J.; Yang, L.; Zhang, D.; Shi, J. Plant metabolomics: An indispensable system biology tool for plant science. Int. J. Mol. Sci. 2016, 17, 767. [Google Scholar] [CrossRef]
- Bao, S. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000; pp. 178–200. [Google Scholar]
- Han, C.; Liu, Y.J.; Zhang, C.K.; Li, Y.G.; Zhou, T.R.; Salman, K.; Chen, N.; Zhao, C.M. Effects of three plantation coniferous species on plant-soil feedbacks and soil physical and chemical properties in semi-arid mountain ecosystems. For. Ecosyst. 2021, 8, 3. [Google Scholar] [CrossRef]
- Yang, D.S.; Zhang, J.; Li, M.X.; Shi, L.X. Metabolomics analysis reveals the salt-tolerant mechanism in Glycine soja. J. Plant Growth Regul. 2017, 36, 460–471. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 7, 1060–1083. [Google Scholar] [CrossRef]
- Guo, Y.; Kong, F.M.; Xu, Y.F.; Zhao, Y.; Liang, X.; Wang, Y.Y.; An, D.G.; Li, S.S. QTL mapping for seedling traits in wheat grown under varying concentrations of N, P and K nutrients. Theor. Appl. Genet. 2011, 124, 851–865. [Google Scholar] [CrossRef] [PubMed]
- Rojas, X.; Guo, J.; Leff, J.W.; McNear, D.H.; Fierer, N.; McCulley, R.L. Infection with a shoot-specific fungal endophyte (Epichloë) alters tall fescue soil microbial communities. Microb. Ecol. 2016, 72, 197–206. [Google Scholar] [CrossRef]
- Müller, C.B.; Krauss, J. Symbiosis between grasses and asexual fungal endophytes. Curr. Opin. Plant Biol. 2005, 8, 450–456. [Google Scholar] [CrossRef]
- Ganie, A.H.; Ahmad, A.; Pandey, R.; Aref, I.M.; Yousuf, P.Y.; Ahmad, S.; Iqbal, M. Metabolite profiling of low-P tolerant and low-P sensitive maize genotypes under phosphorus starvation and restoration conditions. PLoS ONE 2015, 10, e0129520. [Google Scholar] [CrossRef]
- Malinowski, D.P.; Belesky, D.P. Neotyphodium coenophialum-endophyte infection affects the ability of tall fescue to use sparingly available phosphorus. J. Plant Nutr. 1999, 22, 835–853. [Google Scholar] [CrossRef]
- Chen, T.; Li, C.; White, J.F.; Nan, Z. Effect of the fungal endophyte Epichloë bromicola on polyamines in wild barley (hordeum brevisubulatum) under salt stress. Plant Soil 2018, 436, 29–48. [Google Scholar] [CrossRef]
- Zhou, J.L. Interactions of Nitrogen and Phosphorus Supply and Epichloë bromicola on Growth of Wild Barley. Master’s Thesis, Lanzhou University, Lanzhou, China, 2019. [Google Scholar]
- Vázquez-de-Aldana, B.R.; García-Ciudad, A.; García-Criado, B.; Vicente-Tavera, S.; Zabalgogeazcoa, I. Fungal endophyte (Epichloë festucae) alters the nutrient content of Festuca rubra regardless of water availability. PLoS ONE 2013, 8, e84539. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Li, T.; Shen, M.; Yang, Z.L.; Zhao, Z.W. Evidence for a dark septate endophyte (Exophiala Pisciphila, H93) enhancing phosphorus absorption by maize seedlings. Plant Soil 2020, 452, 249–252. [Google Scholar] [CrossRef]
- Guan, J.; Fan, W. Effect of phosphorus supply on growth, nutrient content and related physiology indexes of Rosa roxburghii seedlings. Acta Bot. Boreal. Occident. Sin. 2018, 38, 1315–1324. [Google Scholar]
- Hammer, E.C.; Jan, P.; Wallander, H.; Olsson Pål, A. Tit for tat? A mycorrhizal fungus accumulates phosphorus under low plant carbon availability. FEMS Microbiol. Ecol. 2011, 76, 236–244. [Google Scholar] [CrossRef]
- Richardson, A.E.; Barea, J.-M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Yildirim, E.; Karlidag, H.; Turan, M.; Dursun, A.; Goktepe, F. Growth, nutrient uptake, and yield promotion of broccoli by plant growth promoting rhizobacteria with manure. Hortscience 2011, 46, 932–936. [Google Scholar] [CrossRef]
- Oosterhuis, D.M.; Loka, D.A.; Kawakami, E.M.; Pettigrew, W.T. The physiology of potassium in crop production. Adv. Agron. 2014, 126, 203–233. [Google Scholar]
- Ouyang, Y.; Hong, T.; Hong, C.; Chen, X.; Li, B.; Chen, J.; Wu, C.; Lin, H. Effects of different endophytic fungi on growth and contents of C, N, P and K of seedlings of Aleurites montana under phosphorus deficiency condition. J. Plant Resour. Environ. 2017, 26, 32–44. [Google Scholar]
- Israr, D.; Mustafa, G.; Khan, K.S.; Shahzad, M.; Ahmad, N.; Masood, S. Interactive effects of phosphorus and Pseudomonas putida on chickpea (Cicer arietinum L.) growth, nutrient uptake, antioxidant enzymes and organic acids exudation. Plant Physiol. Biochem. 2016, 108, 304–312. [Google Scholar] [CrossRef]
- Mostafa, G.; Mohsen, M.D.; Ali, R.Y.; Khosro, P.; Dustmorad, Z. Reduced P fertilization improves Fe and Zn uptake in potato when inoculated with AMF in P, Fe and Zn deficient soil. Rhizosphere 2020, 15, 100239. [Google Scholar]
- Wasaki, J.; Yonetani, R.; Kuroda, S.; Shinano, T.; Yazaki, J.; Fujii, F.; Shimbo, K.; Yamamoto, K.; Sakata, K.; Sasaki, T.; et al. Transcriptomic analysis of metabolic changes by phosphorus stress in rice plant roots. Plant Cell Environ. 2003, 26, 1515–1523. [Google Scholar] [CrossRef]
- Zeh, M.; Casazza, A.P.; Kreft, O.; Roessner, U.; Bieberich, K.; Willmitzer, L.; Hoefgen, R.; Hesse, H. Antisense inhibition of threonine synthase leads to high methionine content in transgenic potato plants. Plant Physiol. 2001, 127, 792–802. [Google Scholar] [CrossRef]
- Alonso-Ramírez, A.; Rodríguez, D.; Reyes, D.; Jiménez, J.A.; Nicolás, G.; López-Climent, M.; Gómez-Cadenas, A.; Nicolás, C. Evidence for a role of gibberellins in salicylic acid-modulated early plant responses to abiotic stress in Arabidopsis seeds. Plant Physiol. 2009, 150, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Brauc, S.; De, V.E.; Claeys, M.; Geuns, J.M.; Höfte, M.; Angenon, G. Overexpression of arginase in Arabidopsis thaliana influences defence responses against Botrytis cinerea. Plant Biol. 2012, 14, 39–45. [Google Scholar] [CrossRef]
- Ding, Z.; Jia, S.; Wang, Y.; Xiao, J.; Zhang, Y. Phosphate stresses affect ionome and metabolome in tea plants. Plant Physiol. Biochem. 2017, 120, 30–39. [Google Scholar] [CrossRef]
- Nguyen, T.T.M.; Kitajima, S.; Izawa, S. Importance of glucose-6-phosphate dehydrogenase (G6PDH) for vanillin tolerance in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2014, 118, 263–269. [Google Scholar] [CrossRef]
- Minemba, D.; Gleeson, D.B.; Veneklaas, E.; Ryan, M.H. Variation in morphological and physiological root traits and organic acid exudation of three sweet potato (Ipomoea batatas) cultivars under seven phosphorus levels. Sci. Hortic. 2019, 256, 108572. [Google Scholar] [CrossRef]
- Bouché, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Fait, A.; Hanhineva, K.; Beleggia, R.; Dai, N.; Rogachev, I.; Nikiforova, V.J.; Fernie, A.R.; Aharoni, A. Reconfiguration of the achene and receptacle metabolic networks during strawberry fruit development. Plant Physiol. 2008, 148, 730–750. [Google Scholar] [CrossRef] [PubMed]
- Imperato, V.; Portillo-Estrada, M.; Mcammond, B.M.; Douwen, Y.; Thijs, S. Genomic diversity of two hydrocarbon-degrading and plant growth-promoting Pseudomonas species isolated from the oil field of Bóbrka (Poland). Genes 2019, 10, 443. [Google Scholar] [CrossRef]
- Gaufichon, L.; Reisdorf-Cren, M.; Rothstein, S.J.; Chardon, F.; Suzuki, A. Biological functions of asparagine synthetase in plants. Plant Sci. 2010, 179, 141–153. [Google Scholar] [CrossRef]
- Kısa, D.; Öztürk, L. Responses of the fatty acid composition of Solanum lycopersicum exposed to heavy metal stress. Karaelmas Sci. Eng. J. 2019, 9, 88–96. [Google Scholar]
- Canellas, N.O.A.; Olivares, F.L.; Canellas, L.P. Metabolite fingerprints of maize and sugarcane seedlings: Searching for markers after inoculation with plant growth-promoting bacteria in humic acids. Chem. Biol. Technol. Agric. 2019, 6, 14. [Google Scholar] [CrossRef]
- Wildermuth, M.C. Variations on a theme: Synthesis and modification of plant benzoic acids. Curr. Opin. Plant Biol. 2006, 9, 288–296. [Google Scholar] [CrossRef]
- Keitaro, T.; Ryota, H.; Saki, S.; Tadao, W.; Kazuki, S.; Akira, O. Metabolite profiling of root exudates of common bean under phosphorus deficiency. Metabolites 2014, 4, 599–611. [Google Scholar]
- Hernández, G.; V aldés-López, O.; Ramírez, M.; Goffard, N.; Weiller, G.; Aparicio-Fabre, R. Global changes in the transcript and metabolic profiles during symbiotic nitrogen fixation in phosphorus-stressed common bean plants. Plant Physiol. 2009, 151, 1221–1238. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.S.; Mao, X.T.; Tian, Q.Y.; Li, L.H.; Zhang, W.H. Phosphorus deficiency-induced reduction in root hydraulic conductivity in Medicago falcate is associated with ethylene production. Env. Exp. Bot. 2009, 67, 172–177. [Google Scholar] [CrossRef]
- Vinci, G.; Cozzolino, V.; Mazzei, P.; Monda, H.; Savy, D.; Drosos, M.; Piccolo, A. Effects of Bacillus amyloliquefaciens and different phosphorus sources on maize plants as revealed by NMR and GC-MS based metabolomics. Plant Soil 2018, 429, 437–450. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid Biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Alhaithloul, H.A.S.; Parvin, K.; Bhuyan, M.b.; Tanveer, M.; Mohsin, S.M.; Nahar, K.; Soliman, M.H.; Al Mahmud, J.; Fujita, M. Polyamine action under metal/metalloid stress: Regulation of biosynthesis, metabolism, and molecular interactions. Int. J. Mol. Sci. 2019, 20, 3215. [Google Scholar] [CrossRef]
- Yang, L.; Wang, X.; Chang, N.; Nan, W.; Wang, S.; Ruan, M.; Sun, L.; Li, S.; Bi, Y. Cytosolic glucose-6- phosphate dehydrogenase is involved in seed germination and root growth under salinity in Arabidopsis. Front. Plant Sci. 2019, 10, 182. [Google Scholar] [CrossRef]
- Gao, Y.; Guo, D.L.; Guo, C.H. Role of trienoic fatty acids in higher plants stress responses. Mol. Plant Breed. 2010, 08, 365–369. [Google Scholar]
- Takahashi, H.; Imamura, T.; Miyagi, A.; Uchimiya, H. Comparative metabolomics of developmental alterations caused by mineral deficiency during in vitro culture of Gentiana triflora. Metabolomics 2012, 8, 154–163. [Google Scholar] [CrossRef]
- Maeda, H.; Dudareva, N. The shikimate pathway and aromatic amino acid biosynthesis in plants. Annu. Rev. Plant Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V.; Lattanzio, V.M.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. Phytochem. Advan. Res. 2006, 661, 23–67. [Google Scholar]
- Weih, M.; Asplund, L.; Bergkvist, G. Assessment of nutrient use in annual and perennial crops: A functional concept for analyzing nitrogen use efficiency. Plant Soil 2010, 339, 513–520. [Google Scholar] [CrossRef]
- Richardson, A.E.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; Smith, S.E.; Harvey, P.R.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; et al. Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil 2011, 349, 121–156. [Google Scholar] [CrossRef]
- Rose, T.J.; Rose, M.T.; Pariasca-Tanaka, J.; Heuer, S.; Wissuwa, M. The frustration with utilization: Why have improvements in internal phosphorus utilization efficiency in crops remained so elusive. Front. Plant Sci. 2011, 10, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Somayeh, E.; Hossein, A.A.; Ahmad, A.P.; Hassan, E.; Babak, M.; Fereydoon, S. Consortium of endophyte and rhizosphere phosphate solubilizing bacteria improves phosphorous use efficiency in wheat cultivars in phosphorus deficient soils. Rhizosphere 2020, 14, 100196. [Google Scholar]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Hou, W.; Jin, J.; Christensen, M.J.; Gu, L.; Cheng, C.; Wang, J. Epichloë gansuensis Increases the Tolerance of Achnatherum inebrians to Low-P Stress by Modulating Amino Acids Metabolism and Phosphorus Utilization Efficiency. J. Fungi 2021, 7, 390. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7050390
Liu Y, Hou W, Jin J, Christensen MJ, Gu L, Cheng C, Wang J. Epichloë gansuensis Increases the Tolerance of Achnatherum inebrians to Low-P Stress by Modulating Amino Acids Metabolism and Phosphorus Utilization Efficiency. Journal of Fungi. 2021; 7(5):390. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7050390
Chicago/Turabian StyleLiu, Yinglong, Wenpeng Hou, Jie Jin, Michael J. Christensen, Lijun Gu, Chen Cheng, and Jianfeng Wang. 2021. "Epichloë gansuensis Increases the Tolerance of Achnatherum inebrians to Low-P Stress by Modulating Amino Acids Metabolism and Phosphorus Utilization Efficiency" Journal of Fungi 7, no. 5: 390. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7050390