
Establishment of an Efficient Polyethylene Glycol (PEG)-Mediated Transformation System in Pleurotus eryngii var. ferulae Using Comprehensive Optimization and Multiple Endogenous Promoters


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Medium
2.2. Resistance Gene Selection and Fungal Sensitivity
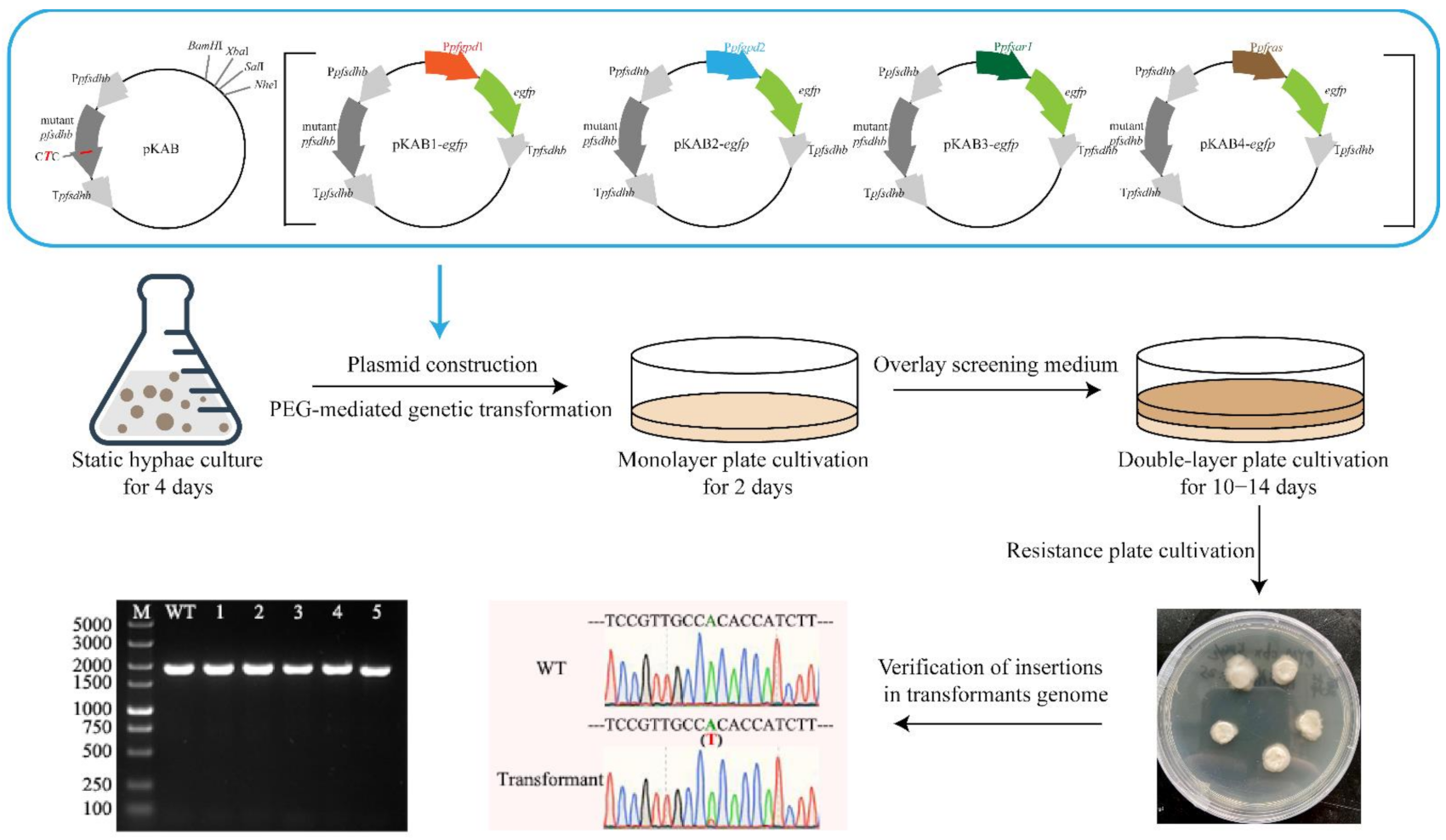
2.3. Plasmid Construction
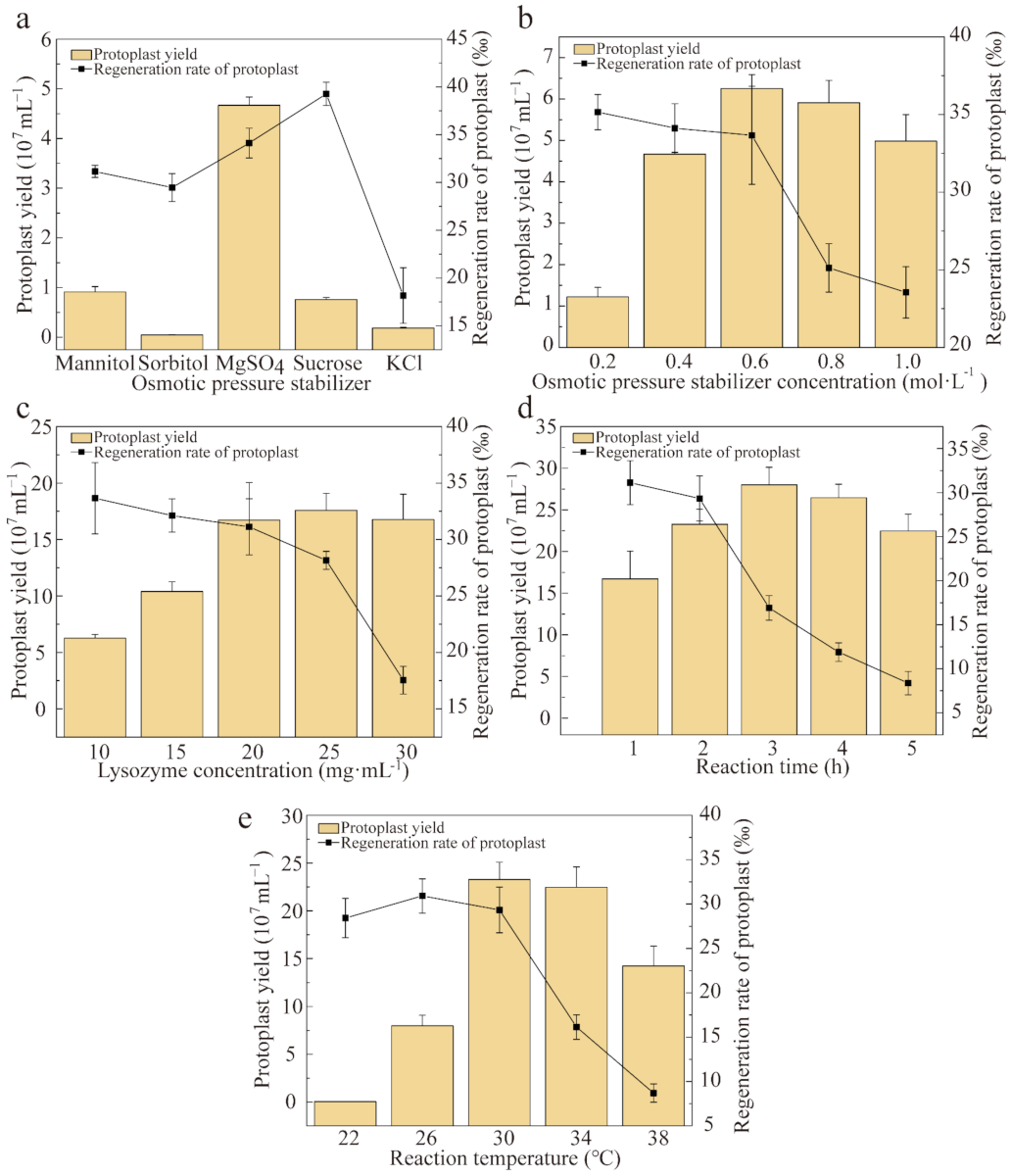
2.4. Preparation of Protoplasts
2.5. PEG-Mediated Protoplast Transformation
2.6. Transformant Verification
2.7. Fluorescence Intensity Analysis
2.8. RT-qPCR
2.9. Statistical Analysis
3. Results
3.1. Resistance Gene Selection and Fungal Sensitivity
3.2. Protoplast Preparation and Regeneration of P. eryngii var. ferulae
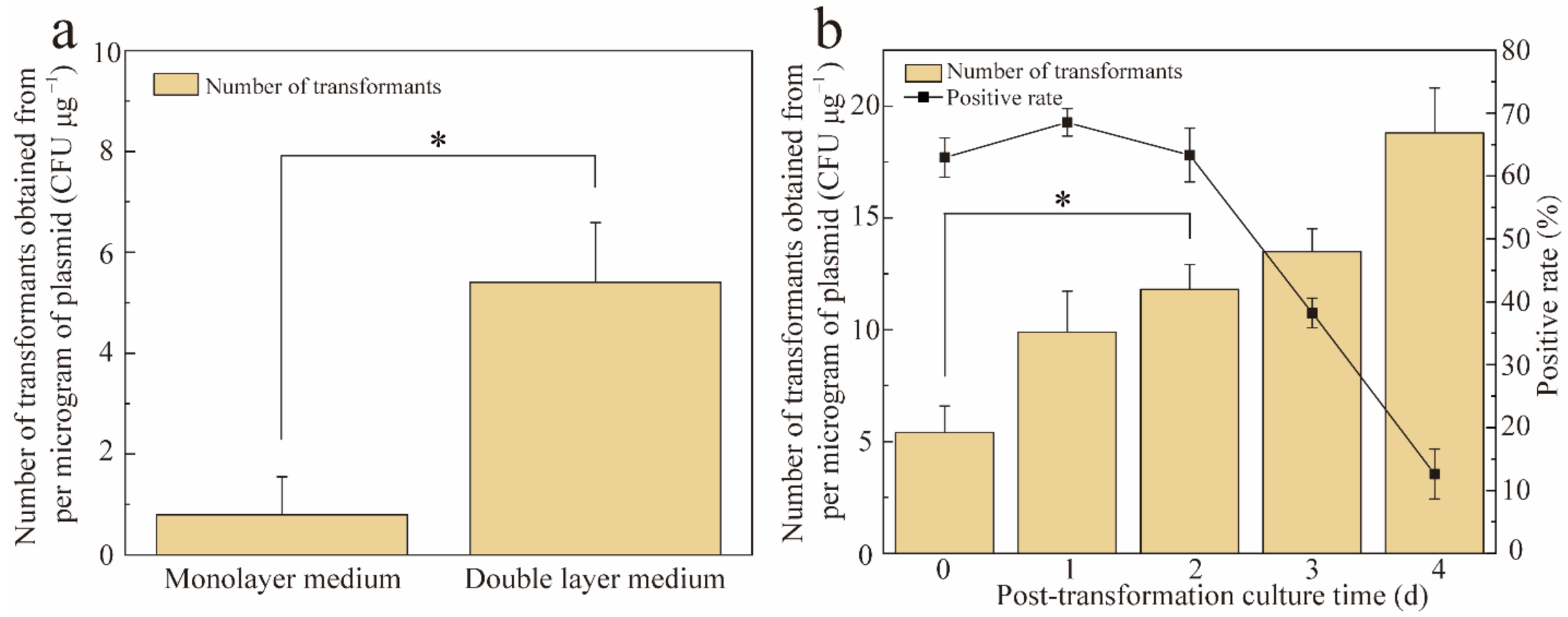
3.3. Transformant Screening Methods
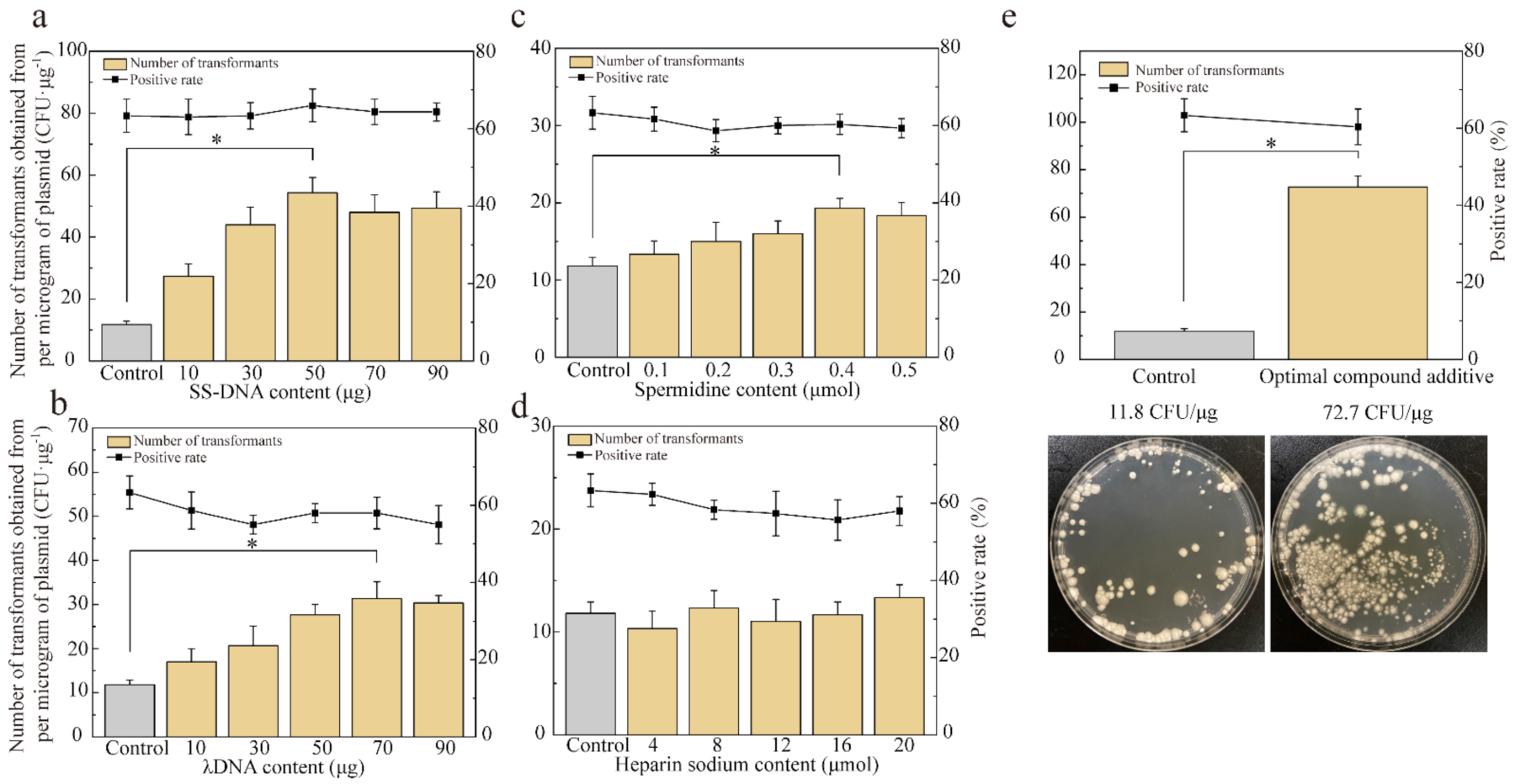
3.4. Effect of Promoting Factors on Enhancing Transformation Efficiency
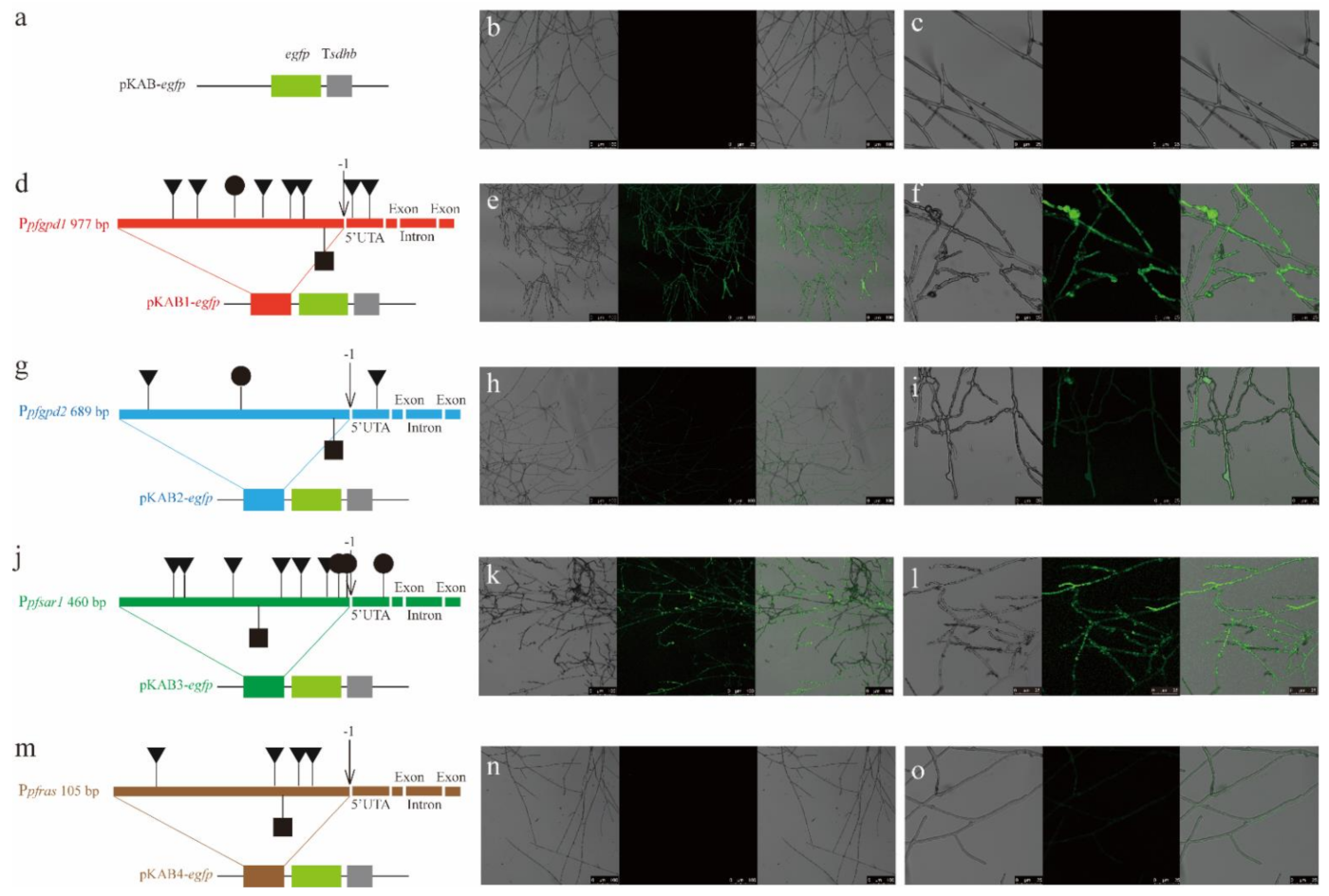
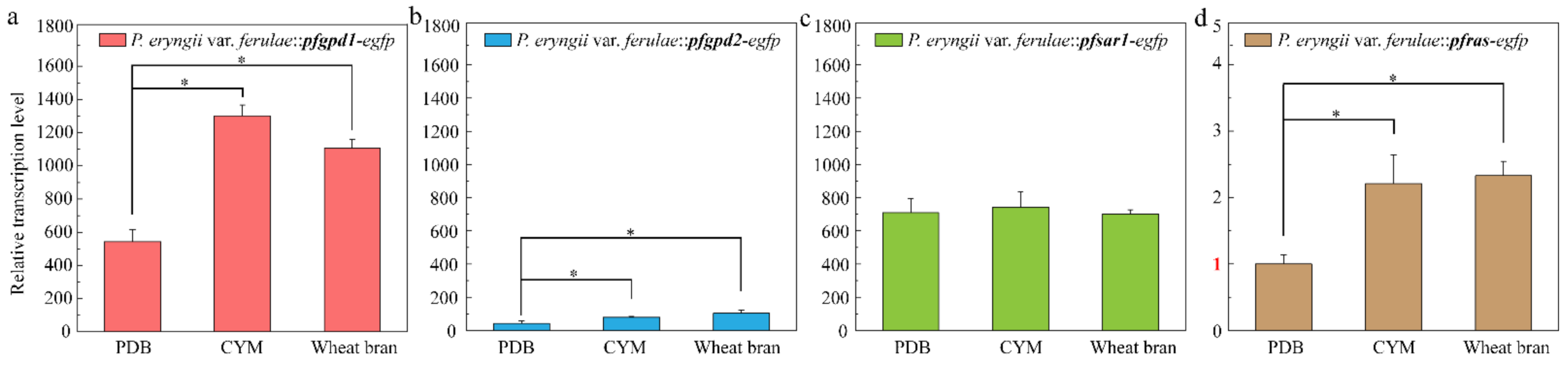
3.5. Effect of Four Endogenous Promoters on EGFP Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fu, Y.; Dai, Y.; Yang, C.; Wei, P.; Song, B.; Yang, Y.; Sun, L.; Zhang, Z.W.; Li, Y. Comparative transcriptome analysis identified candidate genes related to bailinggu mushroom formation and genetic markers for genetic analyses and breeding. Sci. Rep. 2017, 7, 9266. [Google Scholar] [CrossRef] [Green Version]
- La Rosa, A.; Cornara, L.; Saitta, A.; Salam, A.M.; Grammatico, S.; Caputo, M.; La Mantia, T.; Quave, C.L. Ethnobotany of the Aegadian Islands: Safeguarding biocultural refugia in the Mediterranean. J. Ethnobiol. Ethnomed. 2021, 17, 47. [Google Scholar] [CrossRef]
- Zervakis, G.I.; Venturella, G.; Papadopoulou, K. Genetic polymorphism and taxonomic infrastructure of the Pleurotus eryngii speciescomplex as determined by RAPD analysis, isozyme profiles and ecomorphological characters. Microbiology 2001, 147, 3183–3194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Li, X.; Chen, K.; Yang, H.; Jialengbieke, B.; Hu, X. Extraction optimization, characterization and the antioxidant activities in vitro and in vivo of polysaccharide from Pleurotus ferulae. Int. J. Biol. Macromol. 2020, 160, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Mahabati, M.; Aipire, A.; Yuan, P.; Liu, X.; Cai, S.; Aimaier, A.; Ziyayiding, D.; Yasheng, M.; Abudujilile, D.; Li, J. Comparison of structural characteristics and immunoregulatory activities of polysaccharides from four natural plants. Food Agric. Immunol. 2020, 31, 545–562. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Park, Y.H.; Lee, J.S.; Jeong, H.I.; Lee, K.W.; Kang, T.H. Anti-obesity effect of DKB-117 through the inhibition of pancreatic lipase and alpha-amylase activity. Nutrients 2020, 12, 3053. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Aipire, A.; Zhao, H.; Yuan, P.; Li, J. Pleurotus ferulae polysaccharides improve the antitumor efficacy of therapeutic human papillomavirus dendritic cell-based vaccine. Hum. Vaccines Immunother. 2019, 15, 611–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Xu, L.; Zhao, L.; Ding, Z.; Ma, H.; Terry, N. Fungal laccase production from lignocellulosic agricultural wastes by solid-state fermentation: A review. Microorganisms 2019, 7, 665. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Jain, K.K.; Jain, A.; Kidwai, M.; Kuhad, R.C. Bifunctional in vivo role of laccase exploited in multiple biotechnological applications. Appl. Microbiol. Biotechnol. 2018, 102, 10327–10343. [Google Scholar] [CrossRef]
- Guo, C.; Zhao, L.; Wang, F.; Lu, J.; Ding, Z.; Shi, G. Beta-Carotene from yeasts enhances laccase production of Pleurotus eryngii var. ferulae in co-culture. Front. Microbiol. 2017, 8, 1101. [Google Scholar] [CrossRef]
- Wang, H.; Peng, L.; Ding, Z.; Wu, J.; Shi, G. Stimulated laccase production of Pleurotus ferulae JM301 fungus by Rhodotorula mucilaginosa yeast in co-culture. Process Biochem. 2015, 50, 901–905. [Google Scholar] [CrossRef]
- Li, D.; Tang, Y.; Lin, J.; Cai, W. Methods for genetic transformation of filamentous fungi. Microb. Cell Factories 2017, 16, 168. [Google Scholar] [CrossRef] [Green Version]
- Chai, R.; Zhang, G.; Sun, Q.; Zhang, M.; Zhao, S.; Qiu, L. Liposome-mediated mycelial transformation of filamentous fungi. Fungal Biol. 2013, 117, 577–583. [Google Scholar] [CrossRef]
- Lim, F.H.; Rasid, O.A.; Idris, A.S.; As’wad, A.W.M.; Vadamalai, G.; Parveez, G.K.A.; Wong, M.Y. Enhanced polyethylene glycol (PEG)-mediated protoplast transformation system for the phytopathogenic fungus, Ganoderma boninense. Folia Microbiol. 2021, 66, 677–688. [Google Scholar] [CrossRef]
- Kemppainen, M.; Chowdhury, J.; Lundberg-Felten, J.; Pardo, A. Fluorescent protein expression in the ectomycorrhizal fungus Laccaria bicolor: A plasmid toolkit for easy use of fluorescent markers in basidiomycetes. Curr. Genet. 2020, 66, 791–811. [Google Scholar] [CrossRef]
- Yan, L.; Xu, R.; Zhou, Y.; Gong, Y.; Dai, S.; Liu, H.; Bian, Y. Effects of medium composition and genetic background on Agrobacterium-mediated transformation efficiency of Lentinula edodes. Genes 2019, 10, 467. [Google Scholar] [CrossRef] [Green Version]
- Lou, H.-W.; Ye, Z.-W.; Yu, Y.-H.; Lin, J.-F.; Guo, L.-Q.; Chen, B.-X.; Tang, H.-B.; Wei, T.; Chen, L.-T.; Yun, F. The efficient genetic transformation of Cordyceps militaris by using mononuclear protoplasts. Sci. Hortic. 2019, 243, 307–313. [Google Scholar] [CrossRef]
- Zhou, J.; Bai, Y.; Dai, R.; Guo, X.; Liu, Z.H.; Yuan, S. Improved polysaccharide production by homologous co-overexpression of phosphoglucomutase and UDP glucose pyrophosphorylase genes in the mushroom Coprinopsis cinerea. J. Agric. Food Chem. 2018, 66, 4702–4709. [Google Scholar] [CrossRef]
- Shi, L.; Chen, D.; Xu, C.; Ren, A.; Yu, H.; Zhao, M. Highly-efficient liposome-mediated transformation system for the basidiomycetous fungus Flammulina velutipes. J. Gen. Appl. Microbiol. 2017, 63, 179–185. [Google Scholar] [CrossRef] [Green Version]
- You, H.; Sun, B.; Li, N.; Xu, J.W. Efficient expression of heterologous genes by the introduction of the endogenous glyceraldehyde-3-phosphate dehydrogenase gene intron 1 in Ganoderma lucidum. Microb. Cell Factories 2021, 20, 164. [Google Scholar] [CrossRef]
- Geoghegan, I.; Steinberg, G.; Gurr, S. The role of the fungal cell wall in the infection of plants. Trends Microbiol. 2017, 25, 957–967. [Google Scholar] [CrossRef]
- Sato, M.; Kurahashi, A.; Nishibori, K.; Fujimori, F. Development of a transformation system for the edible mushroom Grifola frondosa: Demonstrating heterologous gene expression and RNAi-mediated gene silencing. Mycoscience 2015, 56, 364–372. [Google Scholar] [CrossRef]
- Shi, L.; Fang, X.; Li, M.; Mu, D.; Ren, A.; Tan, Q.; Zhao, M. Development of a simple and efficient transformation system for the basidiomycetous medicinal fungus Ganoderma lucidum. World J. Microbiol. Biotechnol. 2012, 28, 283–291. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, L.; Li, Y.; Wang, F.; Li, S.; Shi, G.; Ding, Z. Comparative transcriptomics and transcriptional regulation analysis of enhanced laccase production induced by co-culture of Pleurotus eryngii var. ferulae with Rhodotorula mucilaginosa. Appl. Microbiol. Biotechnol. 2020, 104, 241–255. [Google Scholar] [CrossRef]
- Shang, J.; Li, Y.; Yang, R.; Wang, Y.; Mao, W.; Tang, L.; Wu, Y.; Nakazawa, T.; Honda, Y.; Li, Y.; et al. Efficient transformation of Pleurotus eryngii with a safe selective marker mutated from the Pesdi1 gene. J. Microbiol. Methods 2018, 152, 7–9. [Google Scholar] [CrossRef]
- Xu, J.-W.; Xu, Y.-N.; Zhong, J.-J. Enhancement of ganoderic acid accumulation by overexpression of an N-terminally truncated 3-hydroxy-3-methylglutaryl coenzyme A reductase gene in the basidiomycete Ganoderma lucidum. Appl. Environ. Microbiol. 2012, 78, 7968–7976. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Yuan, C.; Wang, F.; Xu, S.; Li, Y.; Shi, G.; Ding, Z. Roles of small subunits of laccase (ssPOXA3a/b) in laccase production by Pleurotus eryngii var. ferulae. J. Agric. Food Chem. 2021, 69, 13113–13124. [Google Scholar] [CrossRef]
- Castanera, R.; Lopez-Varas, L.; Pisabarro, A.G.; Ramirez, L. Validation of reference genes for transcriptional analyses in Pleurotus ostreatus by using reverse transcription-quantitative PCR. Appl. Environ. Microbiol. 2015, 81, 4120–4129. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Ji, S.-L.; He, Y.-L.; Ren, M.-F.; Xu, J.-W. Development of an expression plasmid and its use in genetic manipulation of Lingzhi or Reishi medicinal mushroom, Ganoderma lucidum (higher basidiomycetes). Int. J. Med. Mushrooms 2014, 16, 161–168. [Google Scholar] [CrossRef]
- Amore, A.; Honda, Y.; Faraco, V. Enhanced green fluorescent protein expression in Pleurotus ostreatus for in vivo analysis of fungal laccase promoters. Appl. Biochem. Biotechnol. 2012, 168, 761–769. [Google Scholar] [CrossRef]
- Yin, Y.; Liu, Y.; Jin, H.; Wang, S.; Zhao, S.; Geng, X.; Li, M.; Xu, F. Polyethylene glycol-mediated transformation of fused egfp-hph gene under the control of gpd promoter in Pleurotus eryngii. Biotechnol. Lett. 2012, 34, 1895–1900. [Google Scholar] [CrossRef] [PubMed]
- Rajib, S.A.; Sharif Siam, M.K. Characterization and analysis of mammalian AKR7A gene promoters: Implications for transcriptional regulation. Biochem. Genet. 2020, 58, 171–188. [Google Scholar] [CrossRef] [PubMed]
- Porto, M.S.; Pinheiro, M.P.; Batista, V.G.; dos Santos, R.C.; Filho Pde, A.; de Lima, L.M. Plant promoters: An approach of structure and function. Mol. Biotechnol. 2014, 56, 38–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, M.; Wu, X.; Zhang, J.; Wang, H.; Huang, C. Establishment of an efficient transformation system for Pleurotus ostreatus. World J. Microbiol. Biotechnol. 2017, 33, 214. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, R.; Mao, W.J.; Gong, M.; Gao, Y.N.; Tang, L.H.; Yang, R.F.; Li, Y.; Zhou, C.L.; Bao, D.P. A simple and efficient transformation system for the edible mushroom Pleurotus eryngii. Mycoscience 2016, 57, 356–360. [Google Scholar] [CrossRef]
- Irie, T.; Sato, T.; Saito, K.; Honda, Y.; Watanabe, T.; Kuwahara, M.; Enei, H. Construction of a homologous selectable marker gene for Lentinula edodes transformation. Biosci. Biotechnol. Biochem. 2003, 67, 2006–2009. [Google Scholar] [CrossRef] [Green Version]
- Liang, M.; Li, W.; Qi, L.; Chen, G.; Cai, L.; Yin, W.B. Establishment of a genetic transformation system in guanophilic fungus Amphichorda guana. J. Fungi 2021, 7, 138. [Google Scholar] [CrossRef]
- Lin, J.; Zheng, M.; Wang, J.; Shu, W.; Guo, L. Efficient transformation and expression of gfp gene in the edible mushroom Pleurotus nebrodensis. Prog. Nat. Sci. 2008, 18, 819–824. [Google Scholar] [CrossRef]
- Noh, W.; Kim, S.W.; Dong-Won, B.; Kim, J.Y.; Ro, H.S. Genetic introduction of foreign genes to Pleurotus eryngii by restriction enzyme-mediated integration. J. Microbiol. 2010, 48, 253–256. [Google Scholar] [CrossRef]
- Matsunaga, Y.; Ando, M.; Izumitsu, K.; Suzuki, K.; Honda, Y.; Irie, T. A development and an improvement of selectable markers in Pleurotus ostreatus transformation. J. Microbiol. Methods 2017, 134, 27–29. [Google Scholar] [CrossRef]
- Dalya, P.; Slagheka, G.G.; Lópeza, S.C.; Wiebengaa, A.; Hildenb, K.S.; de Vriesa, R.P.; Mäkelä, M.R. Genetic transformation of the white-rot fungus Dichomitus squalens using a new commercial protoplasting cocktail. J. Microbiol. Methods 2017, 143, 38–43. [Google Scholar] [CrossRef]
- Sun, Q.; Wei, W.; Zhao, J.; Song, J.; Peng, F.; Zhang, S.; Zheng, Y.; Chen, P.; Zhu, W. An efficient PEG/CaCl2-mediated transformation approach for the medicinal fungus Wolfiporia cocos. J. Microbiol. Biotechnol. 2015, 25, 1528–1531. [Google Scholar] [CrossRef]
- Irie, T.; Honda, Y.; Watanabe, T.; Kuwahara, M. Efficient transformation of filamentous fungus Pleurotus ostreatus using single-strand carrier DNA. Appl. Microbiol. Biotechnol. 2001, 55, 563–565. [Google Scholar] [CrossRef]
- Wang, J.; Ye, X.; Ni, H.; Zhang, J.; Ju, S.; Ding, W. Transfection efficiency evaluation and endocytosis exploration of different polymer condensed agents. DNA Cell Biol. 2019, 38, 1048–1055. [Google Scholar] [CrossRef]
- Wang, P.A.; Xiao, H.; Zhong, J.J. CRISPR-Cas9 assisted functional gene editing in the mushroom Ganoderma lucidum. Appl. Microbiol. Biotechnol. 2020, 104, 1661–1671. [Google Scholar] [CrossRef]
- Zhang, T.J.; Shi, L.; Chen, D.D.; Liu, R.; Shi, D.K.; Wu, C.G.; Sun, Z.H.; Ren, A.; Zhao, M.W. 14-3-3 proteins are involved in growth, hyphal branching, ganoderic acid biosynthesis, and response to abiotic stress in Ganoderma lucidum. Appl. Microbiol. Biotechnol. 2018, 102, 1769–1782. [Google Scholar] [CrossRef]
- Mu, D.; Shi, L.; Ren, A.; Li, M.; Wu, F.; Jiang, A.; Zhao, M. The development and application of a multiple gene co-silencing system using endogenous URA3 as a reporter gene in Ganoderma lucidum. PLoS ONE 2012, 7, e43737. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Transformation Methods | Transformation Efficiency (Transformants/μg DNA) | Reference |
|---|---|---|---|
| P. eryngii var. ferulae | PMT * | 72.7 | This study |
| P. nebrodensis | PMT | 9 | [38] |
| P. eryngii | REMI * | 10–40 | [39] |
| P. ostreatus | PMT | 26.7(±11.5) | [40] |
| Dichomitus squalens | PMT | 0.8 (±0.3) | [41] |
| Grifola frondosa | PMT | 5.6–11.2 | [22] |
| L. edodes | REMI | 3.6 | [36] |
| Wolfiporia cocos | PMT | 3 | [42] |
| G. lucidum | PMT | 15–20 | [29] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Zhao, L.; Shen, M.; Liu, J.; Li, Y.; Xu, S.; Chen, L.; Shi, G.; Ding, Z. Establishment of an Efficient Polyethylene Glycol (PEG)-Mediated Transformation System in Pleurotus eryngii var. ferulae Using Comprehensive Optimization and Multiple Endogenous Promoters. J. Fungi 2022, 8, 186. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8020186
Zhang Q, Zhao L, Shen M, Liu J, Li Y, Xu S, Chen L, Shi G, Ding Z. Establishment of an Efficient Polyethylene Glycol (PEG)-Mediated Transformation System in Pleurotus eryngii var. ferulae Using Comprehensive Optimization and Multiple Endogenous Promoters. Journal of Fungi. 2022; 8(2):186. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8020186
Chicago/Turabian StyleZhang, Qi, Liting Zhao, Mengye Shen, Jingyun Liu, Youran Li, Sha Xu, Lei Chen, Guiyang Shi, and Zhongyang Ding. 2022. "Establishment of an Efficient Polyethylene Glycol (PEG)-Mediated Transformation System in Pleurotus eryngii var. ferulae Using Comprehensive Optimization and Multiple Endogenous Promoters" Journal of Fungi 8, no. 2: 186. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8020186