
Search for Cryptococcus neoformans/gattii Complexes and Related Genera (Filobasidium, Holtermanniella, Naganishia, Papiliotrema, Solicoccozyma, Vishniacozyma) spp. Biotope: Two Years Surveillance of Wild Avian Fauna in Southern France

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Centre
2.2. Animal Population Sampling
2.3. Method of Sampling
2.4. Treatment of Samples
2.5. Genomic Amplification
2.5.1. DNA Extraction
2.5.2. Amplification
2.5.3. Sequence Analyses
2.6. Statistical Analyses
3. Results
3.1. Cryptococcal Species Distribution in the Bird Population
3.1.1. Total Cryptococcal Species Distribution
3.1.2. Assessment of Highly Pathogenic Cryptococcus Species
3.1.3. Rare Cryptococcus and Former Cryptococcus Species Diversity
3.2. Distribution of RCFC in Birds in Regard of Method of Sampling, Age, Illnesses, Medications and Diets
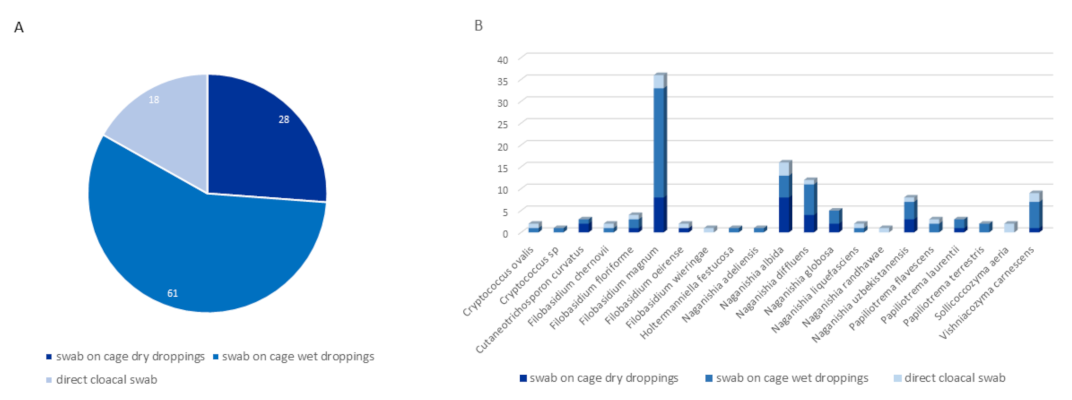
3.2.1. Distribution by Type of Sampling
3.2.2. Distribution by Age
3.2.3. Distribution by Health Issue
3.2.4. Distribution by Medical Treatment
3.2.5. Distribution of RCFC Species according to Their Carrier Bird Feeding Habits
4. Discussion
4.1. RCFC Diversity
4.2. Distribution of RCFC According to Bird Orders, Species, Diet, Clinical Data, Age and Type of Sampling
4.3. The Particular Case of Direct Cloacal Swabs
4.4. Risk Assessment for Humans
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hagen, F.; Jensen, R.H.; Meis, J.F.; Arendrup, M.C. Molecular epidemiology and in vitro antifungal susceptibility testing of 108 clinical Cryptococcus neoformans sensu lato and Cryptococcus gattii sensu lato isolates from Denmark. Mycoses 2016, 59, 576–584. [Google Scholar] [CrossRef]
- Hagen, F.; Lumbsch, H.T.; Arsenijevic, V.A.; Badali, H.; Bertout, S.; Billmyre, R.B.; Bragulat, M.R.; Cabañes, F.J.; Carbia, M.; Chakrabarti, A.; et al. Importance of Resolving Fungal Nomenclature: The Case of Multiple Pathogenic Species in the Cryptococcus Genus. mSphere 2017, 2, e00238-17. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Z.; Wang, G.M.; Göker, M.; Groenewald, M.; Kachalkin, A.V.; Lumbsch, H.T.; Millanes, A.M.; Wedin, M.; Yurkov, A.M.; Bai, F.Y.; et al. Towards an integrated phylogenetic classification of the Tremellomycetes. Stud. Mycol. 2015, 81, 85–147. [Google Scholar] [CrossRef] [Green Version]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.; Loyse, A.; Boulware, D. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Morales-López, S.; Garcia-Effron, G. Infections due to Rare Cryptococcus Species. A Literature Review. J. Fungi 2021, 7, 279. [Google Scholar] [CrossRef]
- Canónico-González, Y.; Adame-Rodríguez, J.M.; Mercado-Hernández, R.; Aréchiga-Carvajal, E.T. Cryptococcus spp. isolation from excreta of pigeons (Columba livia) in and around Monterrey, Mexico. SpringerPlus 2013, 2, 632. [Google Scholar] [CrossRef] [Green Version]
- Cogliati, M.; D’Amicis, R.; Zani, A.; Montagna, M.T.; Caggiano, G.; De Giglio, O.; Balbino, S.; De Donno, A.; Serio, F.; Susever, S.; et al. Environmental distribution of Cryptococcus neoformans and C. gattii around the Mediterranean basin. FEMS Yeast Res. 2016, 16, fow045. [Google Scholar] [CrossRef] [Green Version]
- Soltani, M.; Bayat, M.; Hashemi, S.J.; Zia, M.; Pestechian, N. Isolation of Cryptococcus neoformans and other opportunistic fungi from pigeon droppings. J. Res. Med. Sci. 2013, 18, 56–60. [Google Scholar]
- Cogliati, M.; Patrizia, P.; Vincenzo, C.; Esposto, M.C.; Prigitano, A.; Romanò, L.; Puccianti, E. Cryptococcus neoformans species complex isolates living in a tree micro-ecosystem. Fungal Ecol. 2020, 44, 100889. [Google Scholar] [CrossRef]
- De Jesus, M.S.; Rodrigues, W.; Barbosa, G.; Trilles, L.; Wanke, B.; Lazéra, M.D.S.; da Silva, M. Cryptococcus neoformans carried by Odontomachus bauri ants. Mem. Inst. Oswaldo Cruz 2012, 107, 466–469. [Google Scholar] [CrossRef]
- Mbata, T.I. Isolation of Cryptococcus neoformans from bats (Molossus major) droppings in Awka, Nigeria. Sudan. J. Dermatol. 2007, 4, 81–85. [Google Scholar] [CrossRef]
- Dongmo, W.; Kechia, F.; Tchuenguem, R.; Nangwat, C.; Yves, I.; Kuiate, J.-R.; Dzoyem, J.P. In Vitro Antifungal Susceptibility of Environmental Isolates of Cryptococcus spp. from the West Region of Cameroon. Ethiop. J. Health Sci. 2016, 26, 555–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro e Silva, D.M.; Santos, D.C.S.; Martins, M.A.; Oliveira, L.; Szeszs, M.W.; Melhem, M.S.C. First isolation of Cryptococcus neoformans genotype VNI MAT-alpha from wood inside hollow trunks of Hymenaea courbaril. Med. Mycol. 2015, 54, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Ergin, Ç.; Şengül, M.; Aksoy, L.; Döğen, A.; Sun, S.; Averette, A.F.; Cuomo, C.A.; Seyedmousavi, S.; Heitman, J.; Ilkit, M. Cryptococcus neoformans Recovered $rom Olive Trees (Olea europaea) in Turkey Reveal Allopatry with African and South American Lineages. Front. Cell. Infect. Microbiol. 2019, 9, 384. [Google Scholar] [CrossRef] [Green Version]
- Misra, V.C.; Randhawa, H.S. Occurrence and significance of Cryptococcus neoformans in vegetables and fruits. Indian J. Chest Dis. Allied Sci. 2004, 42, 317–321. [Google Scholar]
- Pal, M.; Onda, C.; Hasegawa, A. Isolation of saprophytic Cryptococcus neoformans. Jpn. J. Vet. Sci. 1990, 52, 1171–1174. [Google Scholar] [CrossRef]
- Botes, A.; Boekhout, T.; Hagen, F.; Vismer, H.; Swart, J.; Botha, A. Growth and Mating of Cryptococcus neoformans var. grubii on Woody Debris . Microb. Ecol. 2008, 57, 757–765. [Google Scholar] [CrossRef] [Green Version]
- Springer, D.J.; Mohan, R.; Heitman, J. Plants promote mating and dispersal of the human pathogenic fungus Cryptococcus. PLoS ONE 2017, 12, e0171695. [Google Scholar] [CrossRef]
- Ellis, D.H.; Pfeiffer, T.J. Natural habitat of Cryptococcus neoformans var. gattii. J. Clin. Microbiol. 1990, 28, 1642–1644. [Google Scholar] [CrossRef] [Green Version]
- Kamari, A.; Sepahvand, A.; Mohammadi, R. Isolation and molecular characterization of Cryptococcus species isolated from pigeon nests and Eucalyptus trees. Curr. Med. Mycol. 2017, 3, 20–25. [Google Scholar] [CrossRef]
- Linares, C.; Colom, M.F.; Torreblanca, M.; Esteban, V.; Romera, Á.; Hagen, F. Environmental sampling of Ceratonia siliqua (carob) trees in Spain reveals the presence of the rare Cryptococcus gattii genotype AFLP7/VGIV. Rev. Iberoam. Micol. 2015, 32, 269–272. [Google Scholar] [CrossRef]
- Hurst, S.; Lysen, C.; Cooksey, G.; Vugia, D.J.; Litvintseva, A.P.; Lockhart, S.R. Molecular typing of clinical and environmental isolates of Cryptococcus gattii species complex from southern California, United States. Mycoses 2019, 62, 1029–1034. [Google Scholar] [CrossRef]
- Khayhan, K.; Hagen, F.; Norkaew, T.; Puengchan, T.; Boekhout, T.; Sriburee, P. Isolation of Cryptococcus gattii from a Castanopsis argyrophylla tree hollow (Mai-Kaw), Chiang Mai, Thailand. Mycopathologia 2017, 182, 365–370. [Google Scholar] [CrossRef]
- Lazera, M.S.; Wanke, B.; Nishikawa, M.M. Isolation of both varieties of Cryptococcus neoformans from saprophytic sources in the city of Rio de Janeiro, Brazil. J. Med. Vet. Mycol. 1993, 31, 449–454. [Google Scholar] [CrossRef]
- Gesuele, E.; Calegar, L.; Sanabria, D.; Davel, G.; Civila, E. Isolation in Uruguay of Cryptococcus neoformans var. gattii from a nest of the wasp Polybia occidentalis. Rev. Iberoam. Micol. 1993, 10, 5–6. [Google Scholar]
- Kidd, S.; Sorrell, T.C.; Meyer, W. Isolation of two molecular types of Cryptococcus neoformans var. gattii from insect frass. J. Med. Vet. Mycol. 2003, 41, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Abulreesh, H.H.; Organji, S.R.; Elbanna, K.; Osman, G.E.; Almalki, M.H.; Abdel-Malek, A.Y.; Ghyathuddin, A.A.; Ahmad, I. Diversity, Virulence Factors, and Antifungal Susceptibility Patterns of Pathogenic and Opportunistic Yeast Species in Rock Pigeon (Columba livia) Fecal Droppings in Western Saudi Arabia. Pol. J. Microbiol. 2019, 68, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Siqueira, N.P.; Favalessa, O.C.; Maruyama, F.H.; Dutra, V.; Nakazato, L.; Hagen, F.; Hahn, R.C. Domestic Birds as Source of Cryptococcus deuterogattii (AFLP6/VGII): Potential Risk for Cryptococcosis. Mycopathologia 2021, 187, 103–111. [Google Scholar] [CrossRef]
- Farrer, R.A.; Chang, M.; Davis, M.J.; van Dorp, L.; Yang, D.-H.; Shea, T.; Sewell, T.R.; Meyer, W.; Balloux, F.; Edwards, H.M.; et al. A New Lineage of Cryptococcus gattii (VGV) Discovered in the Central Zambezian Miombo Woodlands. mBio 2019, 10, e02306-19. [Google Scholar] [CrossRef] [Green Version]
- Duncan, C.; Schwantje, H.; Stephen, C.; Campbell, J.; Bartlett, K. Cryptococcus gattii in Wildlife of Vancouver Island, British Columbia, Canada. J. Wildl. Dis. 2006, 42, 175–178. [Google Scholar] [CrossRef] [Green Version]
- Stephen, C.; Lester, S.; Black, W.; Fyfe, M.; Raverty, S. Multispecies outbreak of cryptococcosis on southern Vancouver Island, British Columbia. Can. Vet. J. 2002, 43, 792–794. [Google Scholar]
- Pavlova, K.; Grigorova, D.; Hristozova, T.; Angelov, A. Yeast strains from Livingston Island, Antarctica. Folia Microbiol. 2001, 46, 397–401. [Google Scholar] [CrossRef]
- Sláviková, E.; Vadkertiová, R. The occurrence of yeasts in the forest soils. J. Basic Microbiol. 2000, 40, 207–212. [Google Scholar] [CrossRef]
- Sláviková, E.; Vadkertiová, R. The diversity of yeasts in the agricultural soil. J. Basic Microbiol. 2003, 43, 430–436. [Google Scholar] [CrossRef]
- Leite, D.P., Jr.; Amadio, J.V.R.S.; Martins, E.R.; Simões, S.A.A.; Yamamoto, A.C.A.; Leal-Santos, F.A.; Takahara, D.T.; Hahn, R.C. Cryptococcus spp. isolated from dust microhabitat in Brazilian libraries. J. Occup. Med. Toxicol. 2012, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Chand-Goyal, T.; Spotts, R.A. Enumeration of bacterial and yeast colonists of apple fruits and identification of epiphytic yeasts on pear fruits in the Pacific Northwest United States. Microbiol. Res. 1996, 151, 427–432. [Google Scholar] [CrossRef]
- Prakash, A.; Randhawa, H.S.; Khan, Z.U.; Ahmad, S.; Hagen, F.; Meis, J.F.; Chowdhary, A. Environmental distribution of Cryptococcus species and some other yeast-like fungi in India. Mycoses 2018, 61, 305–313. [Google Scholar] [CrossRef]
- Silva-Bedoya, L.M.; Ramirez-Castrillon, M.; Osorio-Cadavid, E. Yeast diversity associated to sediments and water from two Colombian artificial lakes. Braz. J. Microbiol. 2014, 45, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Rosario, I.; Soro, G.; Déniz, S.; Ferrer, O.; Acosta, F.; Padilla, D.; Acosta, B. Presence of C. albidus, C. laurentii and C. uniguttulatus in Crop and Droppings of Pigeon Lofts (Columba livia). Mycopathologia 2009, 169, 315–319. [Google Scholar] [CrossRef]
- Danesi, P.; Furnari, C.; Granato, A.; Schivo, A.; Otranto, D.; Capelli, G.; Cafarchia, C. Molecular identity and prevalence of Cryptococcus spp. nasal carriage in asymptomatic feral cats in Italy. Med. Mycol. 2014, 52, 667–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scorzetti, G.; Petrescu, I.; Yarrow, D.; Fell, J.W. Cryptococcus adeliensis sp. nov., a xylanase producing basidiomycetous yeast from Antarctica. Antonie Van Leeuwenhoek 2000, 77, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Yalçın, H.T.; Çorbacı, C.; Uçar, F.B. Molecular characterization and lipase profiling of the yeasts isolated from environments contaminated with petroleum. J. Basic Microbiol. 2014, 54, S85–S92. [Google Scholar] [CrossRef] [PubMed]
- Crestani, J.; Landell, M.; Faganello, J.; Vainstein, M.H.; Vishniac, H.S.; Valente, P. Cryptococcus terrestris sp. nov., a tremellaceous, anamorphic yeast phylogenetically related to Cryptococcus flavescens. Int. J. Syst. Evol. Microbiol. 2009, 59, 631–636. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Ge, C.; Zhou, J.; Li, J. Diversity of soil fungi in the vineyards of Changli region in China. Can. J. Microbiol. 2022, 2022, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Al-Yasiri, M.H.; Normand, A.-C.; L’Ollivier, C.; Lachaud, L.; Bourgeois, N.; Rebaudet, S.; Piarroux, R.; Mauffrey, J.-F.; Ranque, S. Opportunistic fungal pathogen Candida glabrata circulates between humans and yellow-legged gulls. Sci. Rep. 2016, 6, 36157. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, T.G.; Castañeda, E.; Nielsen, K.; Wanke, B.; Lazéra, M.S. Environmental Niches for Cryptococcus neoformans and Cryptococcus gattii. In Cryptococcus; Heitman, J., Kozel, T.R., Kwon-Chung, K.J., Perfect, J.R., Casadevall, A., Eds.; 2010. [Google Scholar] [CrossRef]
- Kassi, F.K.; Drakulovski, P.; Bellet, V.; Krasteva, D.; Gatchitch, F.; Doumbia, A.; Kouakou, G.A.; Delaporte, E.; Reynes, J.; Mallié, M.; et al. Molecular epidemiology reveals genetic diversity among 363 isolates of the Cryptococcus neoformans and Cryptococcus gattii species complex in 61 Ivorian HIV-positive patients. Mycoses 2016, 59, 811–817. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar] [CrossRef]
- O’Donnell, K. Fusarium and its near relatives. In The Fungal Holomorph: Mitotic, Meiotic and Pleomorphic Speciation in Fungal Systematics; Reynolds, D.R., Taylor, J.W., Eds.; CAB International: Wallingford, UK, 1993; pp. 225–233. [Google Scholar]
- Cogliati, M.; Puccianti, E.; Montagna, M.T.; De Donno, A.; Susever, S.; Ergin, C.; Velegraki, A.; Ellabib, M.S.; Nardoni, S.; Macci, C.; et al. Fundamental niche prediction of the pathogenic yeasts Cryptococcus neoformans and Cryptococcus gattii in Europe. Environ. Microbiol. 2017, 19, 4318–4325. [Google Scholar] [CrossRef]
- Alaniz, A.J.; Carvajal, J.G.; Carvajal, M.A.; Cogliati, M.; Vergara, P.M. Spatial Quantification of the Population Exposed to Cryptococcus neoformans and Cryptococcus gattii Species Complexes in Europe: Estimating the Immunocompetent and HIV/AIDS Patients under Risk. Risk Anal. 2019, 40, 524–533. [Google Scholar] [CrossRef]
- Amirrajab, N.; Haghani, I.; Rasuli, M.; Shokohi, T. MigratoryBirds as a Potential Reservoirs of Cryptococcus neoformans. Int. J. Environ. Res. 2016, 10, 459–464. [Google Scholar]
- Daniel, A.J.; Thachil, A.; Savory-Davis, M. Severe cryptococcal granulomas of the nasopharynx in a horse treated with standing endoscopic laser surgery and oral fluconazole. Equine Vet. Educ. 2020, 32, e230–e234. [Google Scholar] [CrossRef]
- Kano, R.; Ishida, R.; Nakane, S.; Sekiguchi, M.; Hasegawa, A.; Kamata, H. The First Reported Case of Canine Subcutaneous Cryptococcus flavescens Infection. Mycopathologia 2011, 173, 179–182. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Brito, M.; de Souza Bessa, M.A.; de Paula Menezes, R.; de Brito Röder, D.V.D.; Penatti, M.P.A.; Pimenta, J.P.; De Aguiar, P.A.D.F.; dos Santos Pedroso, R. Isolation of Cryptococcus species from the external environments of hospital and academic areas. J. Infect. Dev. Ctries. 2019, 13, 545–553. [Google Scholar] [CrossRef]
- Syakalima, M.; Ramatla, T.; Lubanza, N. Opportunistic pathogenic fungi isolated from feces of feral pigeons in Mafikeng, North West Province of South Africa. Vet. World 2019, 12, 1066–1069. [Google Scholar] [CrossRef] [PubMed]
- Tintelnot, K.; Losert, H. Isolation of Cryptococcus adeliensis from Clinical Samples and the Environment in Germany. J. Clin. Microbiol. 2005, 43, 1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirag, B.; Khidir, E.-S.; Dumyati, M.; Sindi, B.; Alsinnari, M.; Faidah, H.; Ahmed, A. Cryptococcus neoformans and Other Opportunistic Cryptococcus Species in Pigeon Dropping in Saudi Arabia: Identification and Characterization by DNA Sequencing. Front. Microbiol. 2021, 12, 726203. [Google Scholar] [CrossRef]
- Ferreira-Paim, K.; Andrade-Silva, L.; Mora, D.J.; Pedrosa, A.L.; Rodrigues, V.; Silva-Vergara, M.L. Genotyping of Cryptococcus neoformans isolated from captive birds in Uberaba, Minas Gerais, Brazil. Mycoses 2010, 54, e294–e300. [Google Scholar] [CrossRef]
- Decostere, A.; Hermans, K.; De Baere, T.; Pasmans, F.; Haesebrouck, F. First report on Cryptococcus laurentii associated with feather loss in a glossy starling (Lamprotornis chalybaeus). Avian Pathol. 2003, 32, 309–311. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, D.C.; Paula, C.R.; da Silva Ruiz, L.; Domaneschi, C.; Navarro, B.S.; de Assis Baroni, F.; Orsi, R.B.; de Souza Carvalho Melhem, M.; Leite, D.P., Jr. Cryptococcus albidus var. albidus Isolated from Turquoise-Fronted Parrots (Amazona aestiva: Psittacidae) Kept in Captivity: A Probable Reservoir Ecological of Fungal Specimen. J. Vet. Sci. Technol. 2017, 8, 1–5. [Google Scholar] [CrossRef]
- Kielstein, P.; Hotzel, H.; Schmalreck, A.; Khaschabi, D.; Glawischnig, W. Occurrence of Cryptococcus spp. in excreta of pigeons and pet birds. Mycoses 2000, 43, 7–15. [Google Scholar] [CrossRef]
- Danesi, P.; Firacative, C.; Cogliati, M.; Otranto, D.; Capelli, G.; Meyer, W. Multilocus sequence typing (MLST) and M13 PCR fingerprinting revealed heterogeneity amongst Cryptococcus species obtained from Italian veterinary isolates. FEMS Yeast Res. 2014, 14, 897–909. [Google Scholar] [CrossRef]
- Cafarchia, C.; Romito, D.; Iatta, R.; Camarda, A.; Montagna, M.T.; Otranto, D. Role of birds of prey as carriers and spreaders of Cryptococcus neoformans and other zoonotic yeasts. J. Med. Vet. Mycol. 2006, 44, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filion, T.; Kidd, S.; Aguirre, K. Isolation of Cryptococcus laurentii from Canada Goose guano in rural upstate New York. Mycopathologia 2006, 162, 363–368. [Google Scholar] [CrossRef]
- Dulisz, B.; Dynowska, M.; Nowakowski, J.J. Body condition and colonization by fungi of House Sparrows Passer domesticus in the urban and rural environment. Eur. Zool. J. 2021, 88, 152–164. [Google Scholar] [CrossRef]
- Dynowska, M.; Biedunkiewicz, A.; Kisicka, I.; Ejdys, E.; Kubiak, D.; Sucharzewska, E. Epidemiological Importance of Yeasts Isolated from the Beak and Cloaca of Healthy Charadriiformes. Bull. Vet. Inst. Pulawy 2015, 59, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Chryssanthou, E.; Wennberg, H.; Bonnedahl, J.; Olsen, B. Occurrence of yeasts in faecal samples from Antarctic and South American seabirds. Mycoses 2011, 54, e811–e815. [Google Scholar] [CrossRef]
- Schreiber, T.; Kamphausen, L.; Haag-Wackernagel, D. Umwelteinflüsse und Gesundheitszustand bei Strassentauben (Columba livia) [Effects of the environment on health of feral pigeons (Columba livia)]. Berl. Munch. Tierarztl. Wochenschr. 2015, 128, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Marietto-Gonçalves, G.A.; Grandi, F. Are all psittacine birds carriers of Cryptococcus neoformans? Mem. Inst. Oswaldo Cruz 2011, 106, 781–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, F.; Leal, D.; Raso, T.F.; Souza, B.; Cunha, R.; Martinez, V.H.R.; Barrouin-Melo, S.M.; Franke, C.R. Risk factors associated with Chlamydia psittaci infection in psittacine birds. J. Med. Microbiol. 2014, 63, 458–463. [Google Scholar] [CrossRef]
- Yurkov, A.M. Yeasts of the soil—Obscure but precious. Yeast 2018, 35, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Iatta, R.; Immediato, D.; Puttilli, M.R.; Danesi, P.; Passantino, G.; Parisi, A.; Mallia, E.; Otranto, D.; Cafarchia, C. Cryptococcus neoformans in the respiratory tract of squirrels, Callosciurus finlaysonii (Rodentia, Sciuridae). Med. Mycol. 2015, 53, 666–673. [Google Scholar] [CrossRef] [Green Version]
- Cadete, R.M.; Lopes, M.R.; Rosa, C.A. Yeasts Associated with Decomposing Plant Material and Rotting Wood. In Yeasts in Natural Ecosystems: Diversity; Buzzini, P., Lachance, M.-A., Yurkov, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 265–292. [Google Scholar] [CrossRef]
- Schmertmann, L.J.; Kan, A.; Mella, V.; Fernandez, C.M.; Crowther, M.; Madani, G.; Malik, R.; Meyer, W.; Krockenberger, M.B. Prevalence of cryptococcal antigenemia and nasal colonization in a free-ranging koala population. Med. Mycol. 2019, 57, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J.; Bennet, J.E. Medical Mycology, 2nd ed.; Lea & Febiger: Philadelphia, PA, USA, 1992. [Google Scholar]
- Pollock, C. Fungal diseases of columbiformes and anseriformes. Vet. Clin. N. Am. Exot. Anim. Pract. 2003, 6, 351–361. [Google Scholar] [CrossRef]
- Gonzalez-Hein, G.; González-Hein, J.; Díaz Jarabrán, M.C. Isolation of Cryptococcus neoformans in Dry Droppings of Captive Birds in Santiago, Chile. J. Avian Med. Surg. 2010, 24, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.K.; Vimercati, L.; Darcy, J.L.; Arán, P.; Gendron, E.M.; Solon, A.J.; Porazinska, D.; Dorador, C. A Naganishiain high places: Functioning populations or dormant cells from the atmosphere? Mycology 2017, 8, 153–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swinne-Desgain, D. Cryptococcus neoformans in the crops of pigeons following its experimental administration. J. Med. Vet. Mycol. 1976, 14, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Pal, M. Cryptococcus neoformans var. neoformans and Munia Birds Cryptococcus neofovmans var. neoformans bei Prachtfinken. Mycoses 2009, 32, 250–252. [Google Scholar] [CrossRef]
- Rosario, I.; de Mendoza, M.H.; Deniz, S.; Soro, G.; Alamo, I.; Acosta, B. Isolation of Cryptococcus species including C. neoformans from cloaca of pigeons. Mycoses 2005, 48, 421–424. [Google Scholar] [CrossRef]
- Johnston, S.A.; Voelz, K.; May, R. Cryptococcus neoformans Thermotolerance to Avian Body Temperature Is Sufficient for Extracellular Growth but not Intracellular Survival in Macrophages. Sci. Rep. 2016, 6, 20977. [Google Scholar] [CrossRef] [Green Version]
- Foti, M.; Rinaldo, D.; Guercio, A.; Giacopello, C.; Aleo, A.; De Leo, F.; Fisichella, V.; Mammina, C. Pathogenic microorganisms carried by migratory birds passing through the territory of the island of Ustica, Sicily (Italy). Avian Pathol. 2011, 40, 405–409. [Google Scholar] [CrossRef] [Green Version]
- Konicek, C.; Vodrážka, P.; Barták, P.; Knotek, Z.; Hess, C.; Racka, K.; Hess, M.; Troxler, S. Detection of Zoonotic Pathogens in Wild Birds in the Cross-Border Region Austria—Czech Republic. J. Wildl. Dis. 2016, 52, 850–861. [Google Scholar] [CrossRef] [Green Version]
- Cano, E.J.; Yetmar, Z.A.; Razonable, R.R. Cryptococcus Species Other Than Cryptococcus neoformans and Cryptococcus gattii: Are They Clinically Significant? Open Forum Infect. Dis. 2020, 7, ofaa527. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Bird Order | Total Birds Sampled within the Order | Number of Birds Positive to RCFC within the Order | Individual Species Names for Positive Birds | Common Names for Positive Birds | Total Number of Individuals Sampled within the Species | Number of Positive Individuals within the Species |
|---|---|---|---|---|---|---|
| Accipitriformes | 75 | 8 (10.6%) | Buteo buteo | Common buzzard | 43 | 4 (9.3%) |
| Accipiter nisus | European sparrowhawk | 20 | 1 (5%) | |||
| Aegypius monachus | Cinereous vulture | 1 | 1 (100%) | |||
| Circaetus gallicus | Short toed snake eagle | 2 | 1 (50%) | |||
| Pernis apivorus | European honey buzzard | 1 | 1 (100%) | |||
| Anseriformes | 17 | 1 (5.9%) | Cygnus olor | Mute swan | 2 | 1 (50%) |
| Apodiformes | 14 | 3 (21.4%) | Apus apus | Common swift | 12 | 3 (25%) |
| Bucerotiformes | 8 | 1 (12.5%) | Upupa epops | Eurasian hoopoe | 8 | 1 (12.5%) |
| Charadriiformes | 113 | 15 (13.3%) | Larus michaelis | Yellow-legged gull | 100 | 13 (13%) |
| Chroicocephalus ridibundus | Black Headed gull | 10 | 1 (10%) | |||
| Scolopex rusticola | Eurasian woodcock | 2 | 1 (50%) | |||
| Ciconiformes | 5 | 1 (20%) | Ciconia ciconia | White stork | 3 | 1 (33.3%) |
| Columbiformes | 180 | 33 (18.3%) | Streptopelia decaocto | Eurasian collared dove | 127 | 20 (15.7%) |
| Columba palumbus | Common wood pigeon | 27 | 9 (33.3%) | |||
| Columba livia | Common pigeon/rock pigeon | 23 | 4 (17.4%) | |||
| Falconiformes | 32 | 9 (28.1%) | Falco tinnunculus | Common kestrel | 25 | 7 (28%) |
| Falco naumanni | Lesser kestrel | 4 | 2 (50%) | |||
| Galliformes | 8 | 1 (12.5%) | Phasianus colchicus | Common pheasant | 1 | 1 (100%) |
| Otidiformes | 1 | 1 (100%) | Tetrax tetrax | Little bustard | 1 | 1 (100%) |
| Passeriformes | 153 | 27 (17.6%) | Sturnus vulgaris | Common starling | 22 | 5 (22.7%) |
| Turdus philomelos | Song thrush | 10 | 4 (40%) | |||
| Passer domesticus | House sparrow | 19 | 3 (15.8%) | |||
| Coeloeus monedula | Eurasian jackdaw | 16 | 2 (12.5%) | |||
| Fringilla coelobs | Common chaffinch | 6 | 2 (33.3%) | |||
| Phoenicurus ochruros | Black redstart | 4 | 2 (50%) | |||
| Pica pica | Eurasian magpie | 24 | 2 (8.3%) | |||
| Sylvia melanocephala | Sardinian warbler | 3 | 2 (66.6%) | |||
| Carduelis carduelis | European goldfinch | 4 | 1 (25%) | |||
| Spinus spinus | Eurasian siskin | 2 | 1 (50%) | |||
| Sylvia atricapilla | Eurasian blackcap | 5 | 1 (20%) | |||
| Troglodytes troglodytes | Eurasian wren | 1 | 1 (100%) | |||
| Turdus merula | Common blackbird | 2 | 1 (50%) | |||
| Pelecaniformes | 7 | 3 (42.8%) | Ardea cinerea | Grey heron | 5 | 3 (60%) |
| Phoenicopteriformes | 6 | 1 (16.6%) | Phoenicopterus roseus | Greater flamingo | 6 | 1 (16.6%) |
| Strigiformes | 33 | 2 (6%) | Otus scops | European scops owl | 6 | 1 (16.6%) |
| Tyto alba | Barn owl | 3 | 1 (33.3%) | |||
| Suliformes | 6 | 1 (16.6%) | Phalacrocorax carbo | Great cormoran | 3 | 1 (33.3%) |
| RCFC Species | Numbers Isolated | Birds from Which RCFC Were Isolated (Order) | Birds from Which RCFC Were Isolated (Species) |
|---|---|---|---|
| Cryptococcus ovalis | 2 | Columbiforme (1) Chaaradriforme (1) | Columba livia (1) Larus michaelis (1) |
| Cryptococcus sp. | 1 | Columbiforme (1) | Streptopelia decaocto (1) |
| Cutaneotrichosporon curvatus | 3 | Falconiforme (1) Apodiforme (1) Charadriiforme (1) | Falco tinnunculus (1) Apus apus (1) Larus michaelis (1) |
| Filobasidium chernovii | 2 | Phoenicoptiforme (1) Passeriforme (1) | Phoenicopterus roseus (1) Phoenicurus ochruros (1) |
| Filobasidium floriforme | 4 | Columbiforme (2) Charadriiforme (2) Strigiforme (1) | Chroicocephalus ridibundus (1) Streptopelia decaocto (2) Tyto alba (1) |
| Filobasidium magnum | 36 | Accipitriforme (1) Apodiforme (2) Columbiforme (15) Falconiforme (2) Passeriforme (11) Charadriiforme (3) Suliforme (1) Galliforme (1) | Accipiter nisus (1) Apus apus (2) Columba livia (1) Columba palumbus (5) Falco naumanni (1) Falco tinnunculus (1) Fringilla coelebs (2) Larus michaelis (3) Passer domesticus (2) Phalocrocorax carbo (1) Phasianus colchicus (1) Phoenicurus ochruros (1) Spinus spinus (1) Streptopelia decaocto (9) Sturnus vulgaris (3) Turdus philomelos (2) |
| Filobasidium wieringae | 1 | Accipitriforme (1) | Buteo buteo (1) |
| Holtermanniella festucosa | 1 | Passeriforme (1) | Carduelis carduelis (1) |
| Naganishia adeliensis | 1 | Columbiforme (1) | Columba palumbus (1) |
| Naganishia albida | 20 | Accipitriforme (2) Columbiforme (6) Charadriiforme (1) Falconiforme (3) Passeriforme (4) | Aegypius monachus (1) Columba livia (1) Streptopelia decaocto (5) Buteo buteo (1) Falco naumanni (1) Falco tinnunculus (2) Larus michaelis (1) Pica pica (1) Sylvia melanocephala (2) Turdus philomelos (1) |
| Naganishia diffluens | 12 | Apodiforme (1) Pelecaniforme (1) Accipitriforme (1) Passeriforme (2) Falconiforme (2) Charadriiforme (1) Strigiforme (1) Columbiform (2) Bucerotiforme (1) | Apus apus (1) Ardea cinerea (1) Circaetus gallicus (1) Coloeus monedula (1) Columba palumbus (1) Falco tinnunculus (2) Larus michaelis (1) Otus scops (1) Pica pica (1) Streptopelia decaocto (1) Upupa epops (1) |
| Naganishia globosa | 5 | Charadriiforme (1) Columbiforme (2) Passeriforme (1) Otidiforme (1) | Larus michaelis (1) Streptopelia decaocto (2) Sylvia atracapilla (1) Tetrax tetrax (1) |
| Naganishia liquefasciens | 2 | Ciconiiforme (1) Passeriforme (1) | Ciconia ciconia (1) Passer domesticus (1) |
| Naganishia oeirense | 2 | Accipitriforme (1) Falconiforme (1) | Buteo buteo (1) Falco tinnunculus (1) |
| Naganishia randhawae | 1 | Charadriiforme (1) | Larus michaelis (1) |
| Naganishia uzbekistanensis | 8 | Pelecaniforme (1) Accipitriforme (1) Columbiforme (2) Falconiforme (1) Charadriiforme (2) Passeriforme (1) | Ardea cinerea (1) Buteo buteo (1) Columba livia (1) Falco tinnunculus (1) Larus michaelis (1) Scolopax rusicola (1) Streptopelia decaocto (1) Coelus monedula (1) |
| Papiliotrema flavescens | 3 | Passeriforme (2) Columbiforme (1) | Sylvia melanocephala (1) Sturnus vulgaris (1) Streptopelia decaocto (1) |
| Papiliotrema laurentii | 3 | Columbiforme (1) Charadriiforme (1) Passeriforme (1) | Columba palumbus (1) Larus michaelis (1) Sturnus vulgaris (1) |
| Papiliotrema terrestris | 2 | Passeriformes (2) | Turdus philomelos (1) Turdus merula (1) |
| Solicoccozyma aeria | 2 | Columbiforme (1) Passeriforme (1) | Streptopelia decaocto (1) Turdus philomelos (1) |
| Vishniacozyma carnescens | 9 | Passeriforme (2) Columbiforme (2) Pelecaniforme (1) Charadriiforme (2) Anseniforme (1) Accipitriforme (1) | Coelus monedula (1) Ardea cinerea (1) Columba palumbus (1) Cygnus olor (1) Larus michaelis (2) Pernus apivorus (1) Troglodytes troglodytes (1) Streptopelia decaocto (1) |
| Total Birds (669) | Positive Birds to RFCF (107, 16%) | |
|---|---|---|
| Age | ||
| Aged birds Adults Juveniles Immatures | 10 486 132 41 | 0 94 (19.3%) 8 (6%) 5 (12.2%) |
| Birds with documented clinical data | 69 | 11 (16%) |
| Infectious diseases Trichomonosis Botulism Bot fly Coccidiosis Avian pox Newcastle disease Ocular infection Digestive infection/parasites | 29 6 8 1 3 6 2 1 2 | 5 (17.2%) 0 1 (12.5%) 0 1 (33.3%) 1 (16.6%) 2 (100%) 0 0 |
| Trauma or physiological trouble Wounds by lead pellets Electrocution Neurological shock Internal bleeding Hematoma Poisoning Trauma Oiled birds Cachexia | 40 14 7 7 3 2 1 1 2 3 | 6 (15%) 5 (35.7%) 1 (14.3%) 0 0 0 0 0 0 0 |
| Birds with documented treatments | 90 | 14 (15.5%) |
| Amoxicillin + clavulanate Amoxicillin + clavulanate +Rapidexon Amoxicillin + clavulanate + Meloxicam Rapidexon Meloxicam Thiamine-Pyridoxine + Vincamine-papaverine Thiamine-Pyridoxine + Vincamine-papaverine + Rapidexon Thiamine-Pyridoxine + Vincamine-papaverine + Meloxicam + vitamin K Rapidexon + Etamsylate Ocular oilment with chloramphenicol Activated coal Fortol (enriched recovery liquid) Activated coal + Etamsylate Rapidexon + Meloxicam + Etamsylate Ocular oilment with oxytetracyline and dexamethasone Carnidazole + Meloxicam Etamsylate + Meloxicam Glucose Rapidexon + Meloxicam Thiamine-Pyridoxine + Vincamine-papaverine + Meloxicam + Rapidexon Etamsylate | 8 3 19 11 17 3 2 1 3 2 4 2 4 1 1 1 2 2 2 1 1 | 1 (12.5%) 0 5 (26.3%) 1 (9%) 4 (23.5%) 0 1 (50%) 0 0 0 1 (25%) 0 0 1 (100%) 0 0 0 0 0 0 0 |
| Main Diet Category at Adult Stage | Total Number of Birds in the Diet Category | Birds Positive to RCFC in the Diet Category |
|---|---|---|
| Grain eaters | 208 | 37 (17.8%) |
| Fruits eaters | 71 | 17 (23.9%) |
| Small vertebrates eaters | 109 | 15 (13.7%) |
| Insectivores | 88 | 17 (19.3%) |
| Omnivorous | 201 | 27 (13.4%) |
| Opportunistic feeders | 169 | 23 (13.6%) |
| Others | 69 | 11 (15.9%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertout, S.; Gouveia, T.; Krasteva, D.; Pierru, J.; Pottier, C.; Bellet, V.; Arianiello, E.; Salipante, F.; Roger, F.; Drakulovski, P. Search for Cryptococcus neoformans/gattii Complexes and Related Genera (Filobasidium, Holtermanniella, Naganishia, Papiliotrema, Solicoccozyma, Vishniacozyma) spp. Biotope: Two Years Surveillance of Wild Avian Fauna in Southern France. J. Fungi 2022, 8, 227. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8030227
Bertout S, Gouveia T, Krasteva D, Pierru J, Pottier C, Bellet V, Arianiello E, Salipante F, Roger F, Drakulovski P. Search for Cryptococcus neoformans/gattii Complexes and Related Genera (Filobasidium, Holtermanniella, Naganishia, Papiliotrema, Solicoccozyma, Vishniacozyma) spp. Biotope: Two Years Surveillance of Wild Avian Fauna in Southern France. Journal of Fungi. 2022; 8(3):227. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8030227
Chicago/Turabian StyleBertout, Sébastien, Tiphany Gouveia, Donika Krasteva, Julie Pierru, Cyrille Pottier, Virginie Bellet, Emilie Arianiello, Florian Salipante, Frédéric Roger, and Pascal Drakulovski. 2022. "Search for Cryptococcus neoformans/gattii Complexes and Related Genera (Filobasidium, Holtermanniella, Naganishia, Papiliotrema, Solicoccozyma, Vishniacozyma) spp. Biotope: Two Years Surveillance of Wild Avian Fauna in Southern France" Journal of Fungi 8, no. 3: 227. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8030227