
Metrics of Antifungal Effects of Ciprofloxacin on Aspergillus fumigatus Planktonic Growth and Biofilm Metabolism; Effects of Iron and Siderophores

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
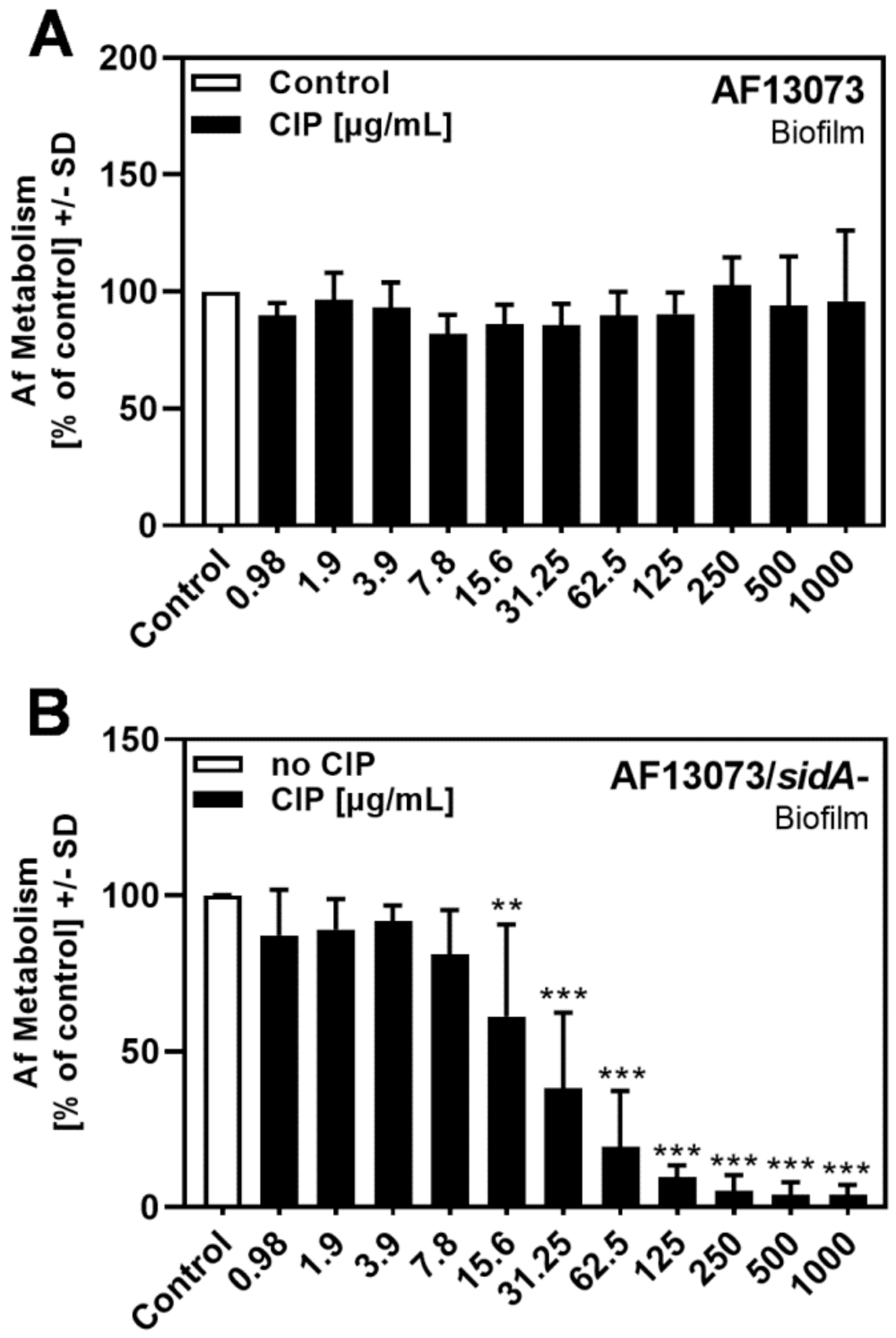
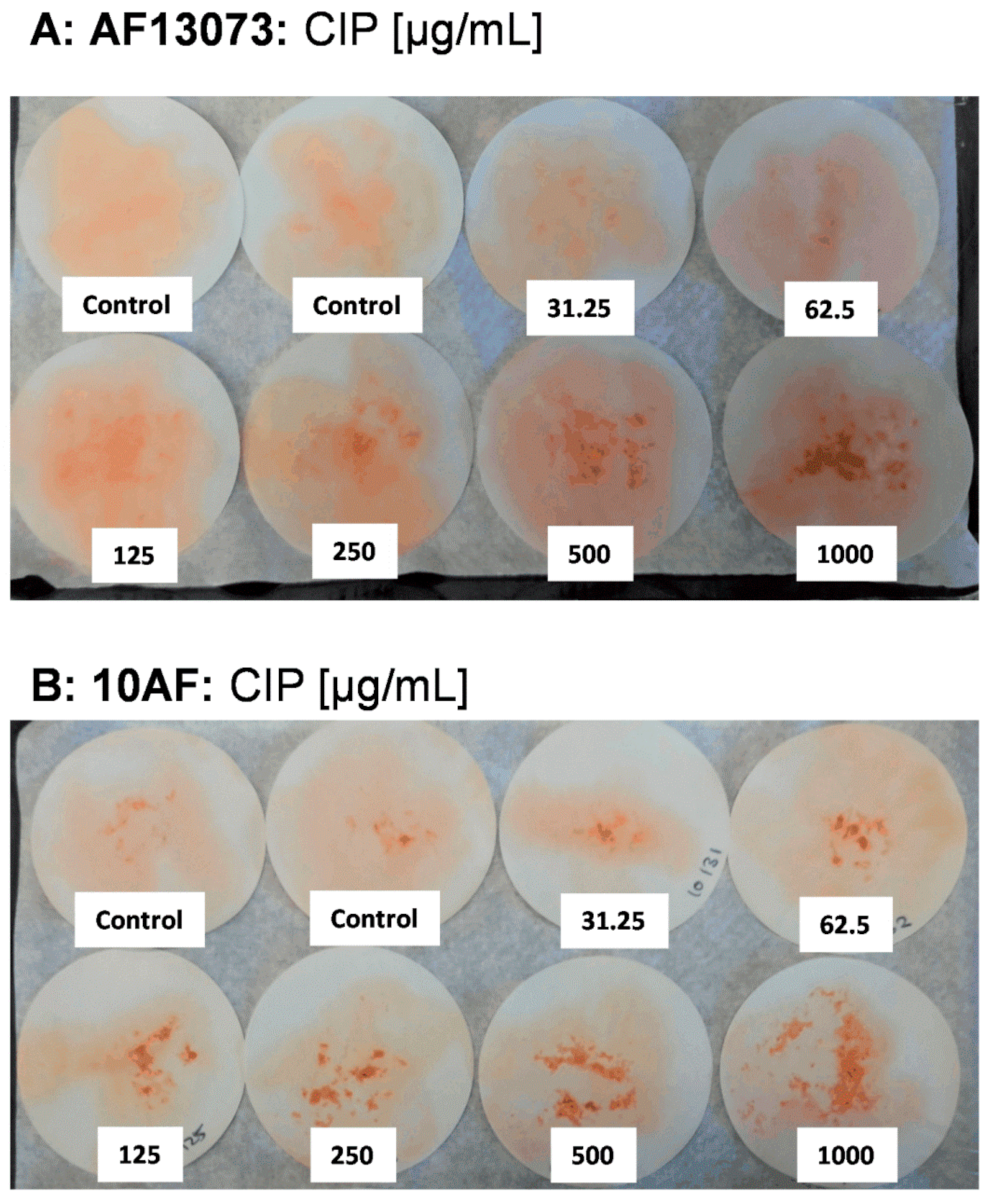
3.1. CIP Reduces A. fumigatus Biofilm Metabolism in the Absence of Siderophores
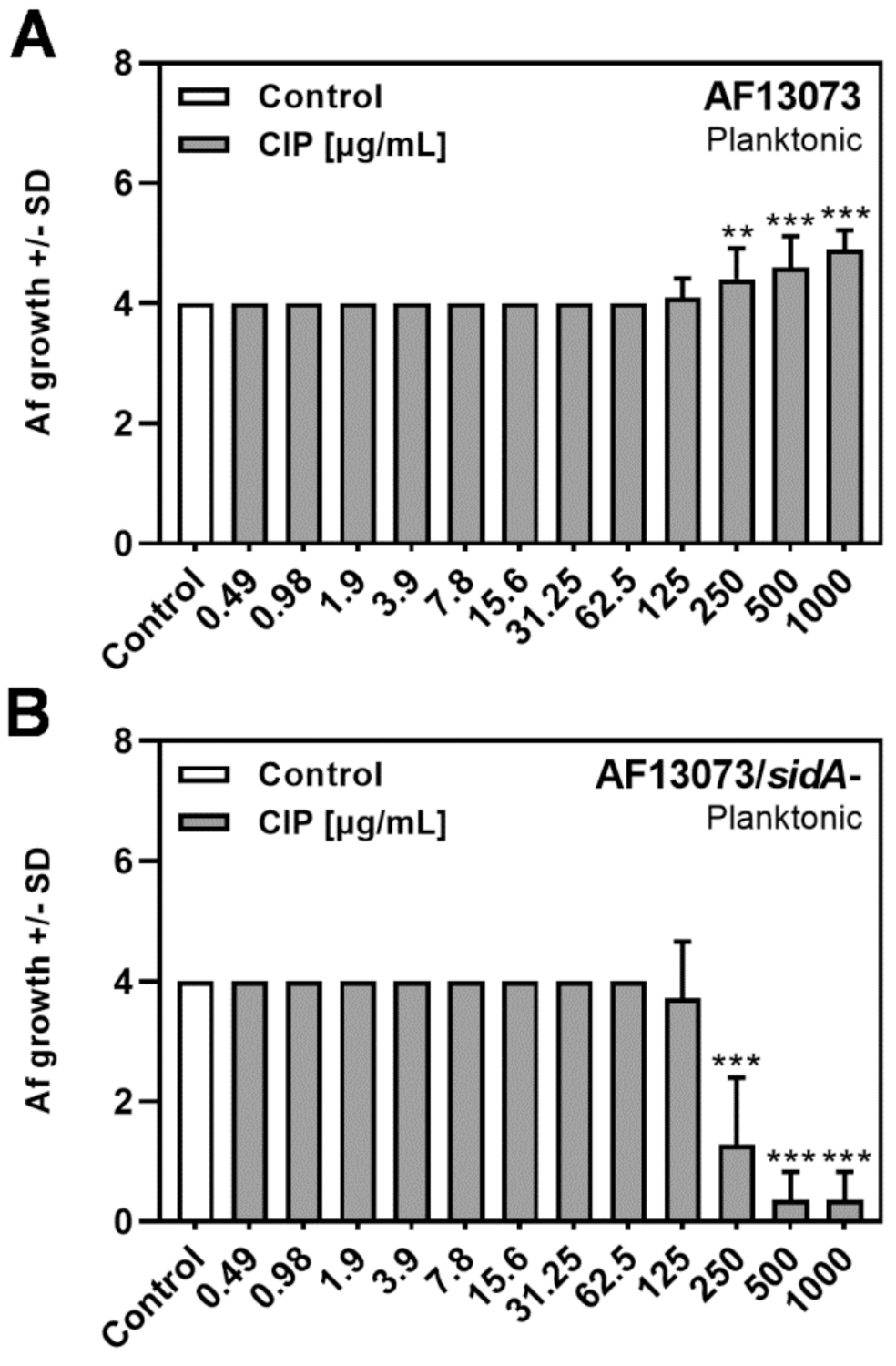
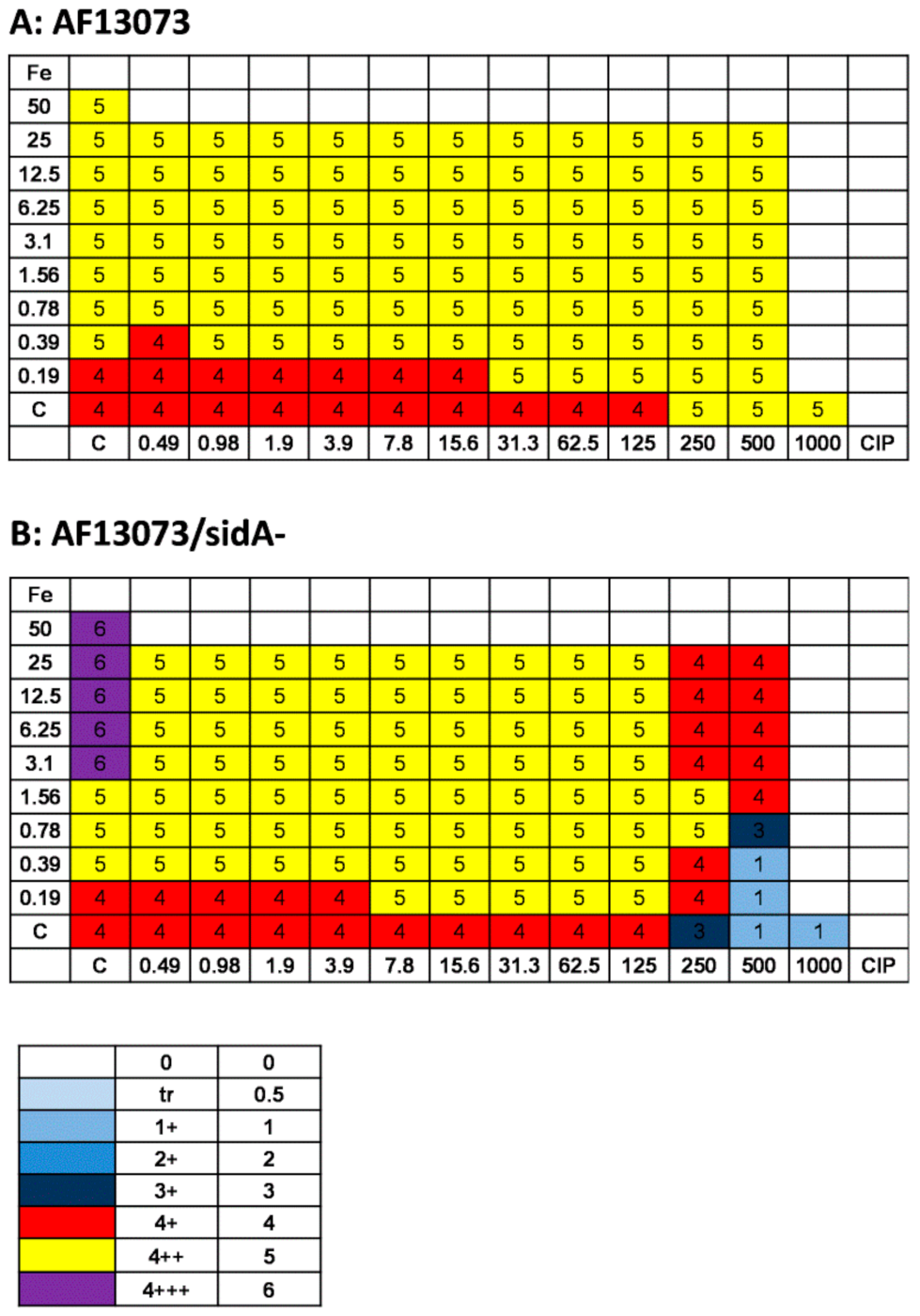
3.2. CIP Induces A. fumigatus Planktonic Wildtype Growth, but Reduces Planktonic Growth of a Siderophore-Deficient Mutant
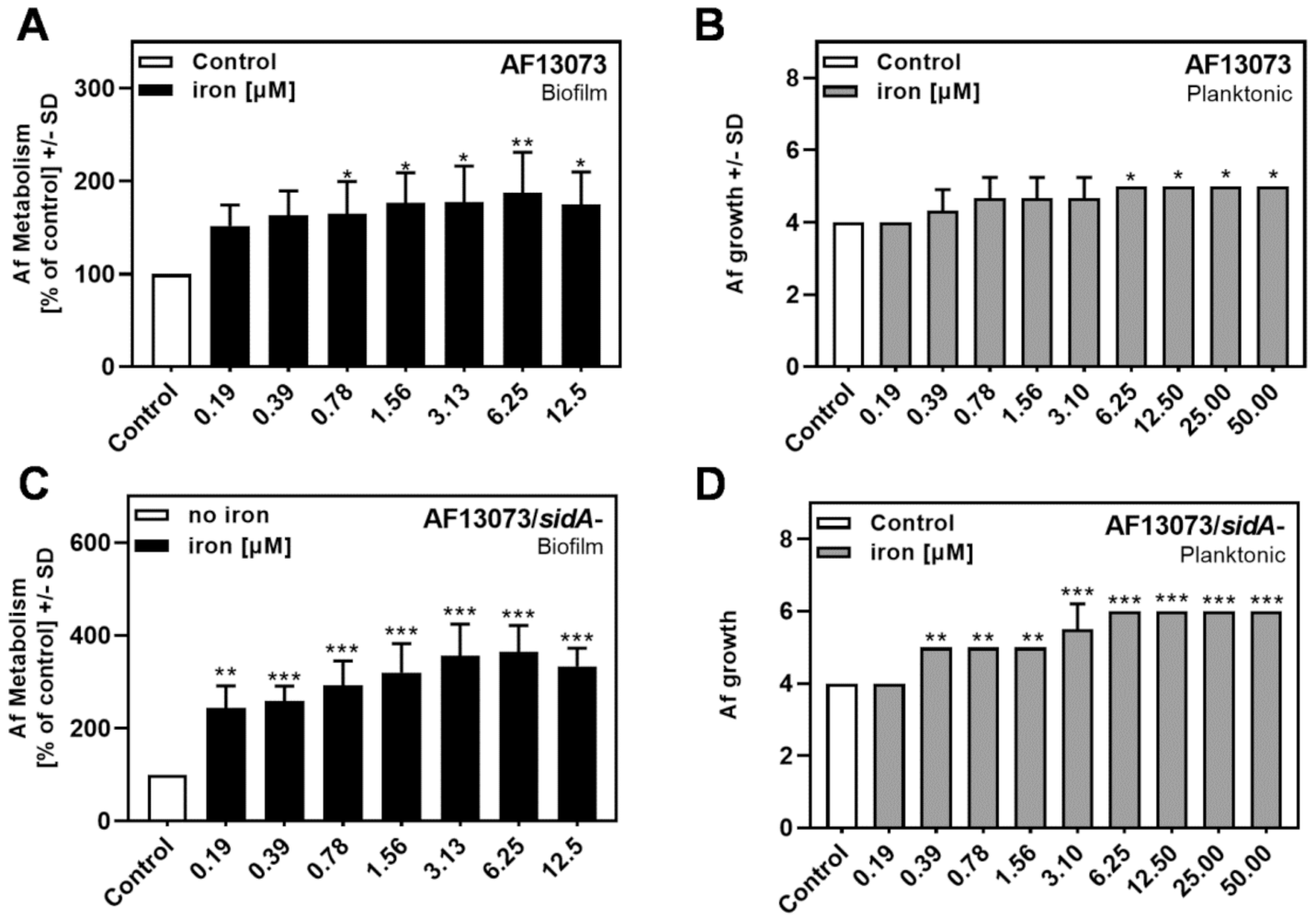
3.3. Iron Induces the Metabolism and Growth of A. fumigatus
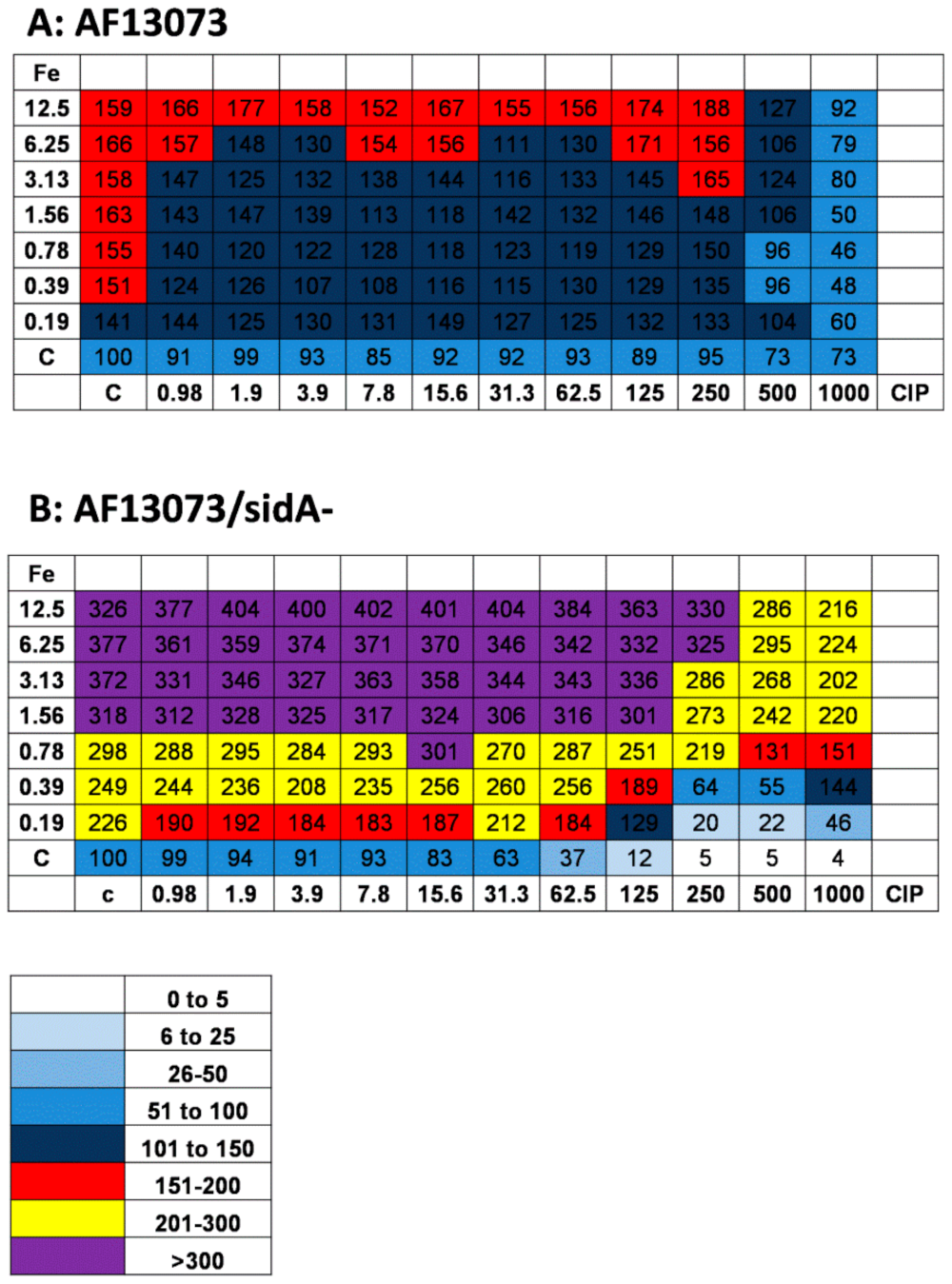
3.4. Iron and CIP Interact and Affect A. fumigatus Biofilm Metabolism
3.5. Iron Does Not Further Increase Profungal Effects of CIP on A. fumigatus Planktonic Growth
4. Discussion
5. Impact
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- King, J.; Brunel, S.F.; Warris, A. Aspergillus infections in cystic fibrosis. J. Infect. 2016, 72, S50–S55. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.; Fothergill, J.L. The role of multispecies social interactions in shaping Pseudomonas aeruginosa pathogenicity in the cystic fibrosis lung. FEMS Microbiol. Lett. 2017, 364, fnx128. [Google Scholar] [CrossRef] [PubMed]
- Wambaugh, M.A.; Denham, S.T.; Brammer, B.; Stonhill, M.; Brown, J.C. Synergistic and antagonistic drug interactions in the treatment of systemic fungal infections. Elife 2020, 9, e54160. [Google Scholar] [CrossRef] [PubMed]
- Elborn, J.S. Cystic fibrosis. Lancet 2016, 388, 2519–2531. [Google Scholar] [CrossRef]
- Ferreira, J.A.; Penner, J.C.; Moss, R.B.; Haagensen, J.A.; Clemons, K.V.; Spormann, A.M.; Nazik, H.; Cohen, K.; Banaei, N.; Carolino, E.; et al. Inhibition of Aspergillus fumigatus and its biofilm by Pseudomonas aeruginosa is dependent on the source, phenotype and growth conditions of the bacterium. PLoS ONE 2015, 10, e0134692. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, C.; Hartl, D.; Eickmeier, O.; Hector, A.; Benden, C.; Durieu, I.; Sole, A.; Gartner, S.; Milla, C.E.; Barry, P.J. Progress in definition, prevention and treatment of fungal infections in cystic fibrosis. Mycopathologia 2018, 183, 21–32. [Google Scholar] [CrossRef]
- Sass, G.; Nazik, H.; Penner, J.; Shah, H.; Ansari, S.R.; Clemons, K.V.; Groleau, M.C.; Dietl, A.M.; Visca, P.; Haas, H.; et al. Studies of Pseudomonas aeruginosa mutants indicate pyoverdine as the central factor in inhibition of Aspergillus fumigatus biofilm. J. Bacteriol. 2017, 200, e00345-17. [Google Scholar] [CrossRef] [Green Version]
- Nazik, H.; Sass, G.; Ansari, S.R.; Ertekin, R.; Haas, H.; Déziel, E.; Stevens, D.A. Novel intermicrobial molecular interaction: Pseudomonas aeruginosa quinolone signal (PQS) modulates Aspergillus fumigatus response to iron. Microbiology 2019, 166, 44–55. [Google Scholar] [CrossRef]
- Sass, G.; Ansari, S.R.; Dietl, A.M.; Déziel, E.; Haas, H.; Stevens, D.A. Intermicrobial interaction: Aspergillus fumigatus siderophores protect against competition by Pseudomonas aeruginosa. PLoS ONE 2019, 14, e0216085. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, P.; Sass, G.; Swietnicki, W.; Stevens, D.A. Review of potential Pseudomonas weaponry, relevant to the Pseudomonas-Aspergillus interplay, for the Mycology community. J. Fungi 2020, 6, 81. [Google Scholar] [CrossRef]
- Heymann, P.; Gerads, M.; Schaller, M.; Dromer, F.; Winkelmann, G.; Ernst, J. The siderophore iron transporter of Candida albicans (Sit1p/Arn1p) mediates uptake of ferrichrome-type siderophores and is required for epithelial invasion. Infect. Immun. 2002, 70, 5246–5255. [Google Scholar] [CrossRef] [Green Version]
- Matthaiou, E.I.; Sass, G.; Stevens, D.A.; Hsu, J.L. Iron: An essential nutrient for Aspergillus fumigatus and a fulcrum for pathogenesis. Curr. Opin. Infect. Dis. 2018, 31, 506–511. [Google Scholar] [CrossRef]
- Reid, D.W.; Withers, N.J.; Francis, L.; Wilson, J.W.; Kotsimbos, T.C. Iron deficiency in cystic fibrosis: Relationship to lung disease severity and chronic Pseudomonas aeruginosa infection. Chest 2002, 121, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Reid, D.W.; Carroll, V.; O’May, C.; Champion, A.; Kirov, S.M. Increased airway iron as a potential factor in the persistence of Pseudomonas aeruginosa infection in cystic fibrosis. Eur. Respir. J. 2007, 30, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.D.M.; Ziora, Z.M.; Blaskovich, M.A.T. Quinolone antibiotics. MedChemComm 2019, 10, 1719–1739. [Google Scholar] [CrossRef]
- Bosso, J.A. Use of ciprofloxacin in cystic fibrosis patients. Am. J. Med. 1989, 87, 123S–127S. [Google Scholar] [CrossRef]
- Scully, B.E.; Nakatomi, M.; Ores, C.; Davidson, S.; Neu, H.C. Ciprofloxacin therapy in cystic fibrosis. Am. J. Med. 1987, 82, 196–201. [Google Scholar]
- Heeb, S.; Fletcher, M.P.; Chhabra, S.R.; Diggle, S.P.; Williams, P.; Cámara, M. Quinolones: From antibiotics to autoinducers. FEMS Microbiol. Rev. 2011, 35, 247–274. [Google Scholar] [CrossRef] [Green Version]
- Ghelani, R. Ciprofloxacin (Ciproxin): An antibiotic for bacterial infections. Netdoctor. 2019. Available online: https://www.netdoctor.co.uk/medicines/infection/a28009/ciprofloxacin-uses-actions/ (accessed on 9 December 2021).
- Schrettl, M.; Bignell, E.; Kragl, C.; Joechl, C.; Rogers, T.; Arst, H.N., Jr.; Haynes, K.; Haas, H. Siderophore biosynthesis but not reductive iron assimilation is essential for Aspergillus fumigatus virulence. J. Exp. Med. 2004, 200, 1213–1219. [Google Scholar] [CrossRef] [Green Version]
- Denning, D.W.; Clemons, K.V.; Hanson, L.H.; Stevens, D.A. Restriction endonuclease analysis of total cellular DNA of Aspergillus fumigatus isolates of geographically and epidemiologically diverse origin. J. Infect. Dis. 1990, 162, 1151–1158. [Google Scholar] [CrossRef]
- Denning, D.W.; Stevens, D.A. Efficacy of cilofungin alone and in combination with amphotericin B in a murine model of disseminated aspergillosis. Antimicrob. Agents Chemother. 1991, 35, 1329–1333. [Google Scholar] [CrossRef] [Green Version]
- Scudiero, D.A.; Shoemaker, R.H.; Paull, K.D.; Monks, A.; Tierney, S.; Nofziger, T.H.; Currens, M.J.; Seniff, D.; Boyd, M.R. Evaluation of a soluble tetrazolium/formazan assay for cell growth and drug sensitivity in culture using human and other tumor cell lines. Cancer Res. 1988, 48, 4827–4833. [Google Scholar]
- Sass, G.; Shrestha, P.; Stevens, D.A. Pseudomonas aeruginosa virulence factors support voriconazole effects on Aspergillus fumigatus. Pathogens 2021, 10, 519. [Google Scholar] [CrossRef]
- Nazik, H.; Penner, J.C.; Ferreira, J.A.; Haagensen, J.A.; Cohen, K.; Spormann, A.M.; Martinez, M.; Chen, V.; Hsu, J.L.; Clemons, K.V.; et al. Effects of iron chelators on the formation and development of Aspergillus fumigatus biofilm. Antimicrob. Agents Chemother. 2015, 59, 6514–6520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Yoon, S.S. Pseudomonas aeruginosa biofilm, a programmed bacterial life for fitness. J. Microbiol. Biotechnol. 2017, 27, 1053–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Singh, S. Biofilm formation by Aspergillus fumigatus. Med. Mycol. 2014, 52, 2–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naoi, T.; Shibuya, N.; Inoue, H.; Mita, S.; Kobayashi, S.; Watanabe, K.; Orino, K. The effect of tert-butylhydroquinone-induced oxidative stress in MDBK cells using XTT assay: Implication of tert-butylhydroquinone-induced NADPH generating enzymes. J. Vet. Med. Sci. 2009, 72, 321–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGaw, L.J.; Elgorashi, E.E.; Eloff, J.N. Cytotoxicity of African medicinal plants against normal animal and human cells. Toxicol. Surv. Afr. Med. Plants 2014, 181–233. [Google Scholar] [CrossRef]
- Hangas, A.; Aasumets, K.; Kekäläinen, N.J.; Paloheinä, M.; Pohjoismäki, J.L.; Gerhold, J.M.; Goffart, S. Ciprofloxacin impairs mitochondrial DNA replication initiation through inhibition of Topoisomerase 2. Nucleic Acids Res. 2018, 46, 9625–9636. [Google Scholar] [CrossRef]
- LeBel, M. Ciprofloxacin: Chemistry, mechanism of action, resistance, antimicrobial spectrum, pharmacokinetics, clinical trials, and adverse reactions. Pharmacotherapy 1988, 8, 3–33. [Google Scholar] [CrossRef]
- Stergiopoulou, T.; Meletiadis, J.; Sein, T.; Papaioannidou, P.; Tsiouris, I.; Roilides, E.; Walsh, T.J. Isobolographic analysis of pharmacodynamic interactions between antifungal agents and ciprofloxacin against Candida albicans and Aspergillus fumigatus. Antimicrob. Agents Chemother. 2008, 52, 2196–2204. [Google Scholar] [CrossRef] [Green Version]
- Brilhante, R.S.; Caetano, E.P.; Sidrim, J.J.; Cordeiro, R.A.; Camargo, Z.P.; Fechine, M.A.; Lima, R.A.; Castelo Branco, D.S.; Marques, F.J.; Mesquita, J.R.; et al. Ciprofloxacin shows synergism with classical antifungals against Histoplasma capsulatum var. capsulatum and Coccidioides posadasii. Mycoses 2012, 56, 397–401. [Google Scholar] [CrossRef] [Green Version]
- Ben Yaakov, D.; Shadkchan, Y.; Albert, N.; Kontoyiannis, D.P.; Osherov, N. The quinolone bromoquinol exhibits broad-spectrum antifungal activity and induces oxidative stress and apoptosis in Aspergillus fumigatus. J. Antimicrob. Chemother. 2017, 72, 2263–2272. [Google Scholar] [CrossRef]
- D’Espine, M.; Bellido, F.; Pechère, J.C.; Auckenthaler, R.; Rohner, P.; Lew, D.; Hirschel, B. Serum levels of ciprofloxacin after single oral doses in patients with septicemia. Eur. J. Clin. Microbiol. Infect. Dis. 1989, 8, 1019–1023. [Google Scholar] [CrossRef]
- Kontou, P.; Chatzika, K.; Pitsiou, G.; Stanopoulos, I.; Argyropoulou-Pataka, P.; Kioumis, I. Pharmacokinetics of ciprofloxacin and its penetration into bronchial secretions of mechanically ventilated patients with chronic obstructive pulmonary disease. Antimicrob. Agents Chemother. 2011, 55, 4149–4153. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.U.; Hong, S.W.; Ko, J.H. Efficacy of inhaled ciprofloxacin agents for the treatment of bronchiectasis: A systematic review and meta-analysis of randomized controlled trials. Ther. Adv. Respir. Dis. 2019, 13, 1753466619875930. [Google Scholar] [CrossRef]
- Dalhoff, A. Pharmacokinetics and pharmacodynamics of aerosolized antibacterial agents in chronically infected cystic fibrosis patients. Clin. Microbiol. Rev. 2014, 27, 753–782. [Google Scholar] [CrossRef] [Green Version]
- Polk, R.E.; Healy, D.P.; Sahai, J.; Drwal, L.; Racht, E. Effect of ferrous sulfate and multivitamins with zinc on absorption of ciprofloxacin in normal volunteers. Antimicrob. Agents Chemother. 1989, 33, 1841–1844. [Google Scholar] [CrossRef] [Green Version]
- Andriole, V.T. The quinolones history and overview. In The Quinolones; Academic Press: San Diego, CA, USA, 2000; p. 22. [Google Scholar]
- Kara, M.; Hasinoff, B.B.; McKay, D.W.; Campbell, N.R. Clinical and chemical interactions between iron preparations and ciprofloxacin. Br. J. Clin. Pharmacol. 1991, 31, 257–261. [Google Scholar] [CrossRef]
- Wang, J.; Lory, S.; Ramphal, R.; Jin, S. Isolation and characterization of Pseudomonas aeruginosa genes inducible by respiratory mucus derived from cystic fibrosis patients. Mol. Microbiol. 1996, 22, 1005–1012. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sass, G.; Scherpe, L.; Martinez, M.; Marsh, J.J.; Stevens, D.A. Metrics of Antifungal Effects of Ciprofloxacin on Aspergillus fumigatus Planktonic Growth and Biofilm Metabolism; Effects of Iron and Siderophores. J. Fungi 2022, 8, 240. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8030240
Sass G, Scherpe L, Martinez M, Marsh JJ, Stevens DA. Metrics of Antifungal Effects of Ciprofloxacin on Aspergillus fumigatus Planktonic Growth and Biofilm Metabolism; Effects of Iron and Siderophores. Journal of Fungi. 2022; 8(3):240. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8030240
Chicago/Turabian StyleSass, Gabriele, Lynn Scherpe, Marife Martinez, Julianne J. Marsh, and David A. Stevens. 2022. "Metrics of Antifungal Effects of Ciprofloxacin on Aspergillus fumigatus Planktonic Growth and Biofilm Metabolism; Effects of Iron and Siderophores" Journal of Fungi 8, no. 3: 240. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8030240