
Gram-Level Production of Balanol through Regulatory Pathway and Medium Optimization in Herb Fungus Tolypocladium ophioglossoides


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Strain, Media and Culture Conditions
2.3. Heterologous Expression of blnR in E. coli and Its Purification
2.4. Affinity Analysis by Electrophoretic Mobility Shift Assay (EMSA)
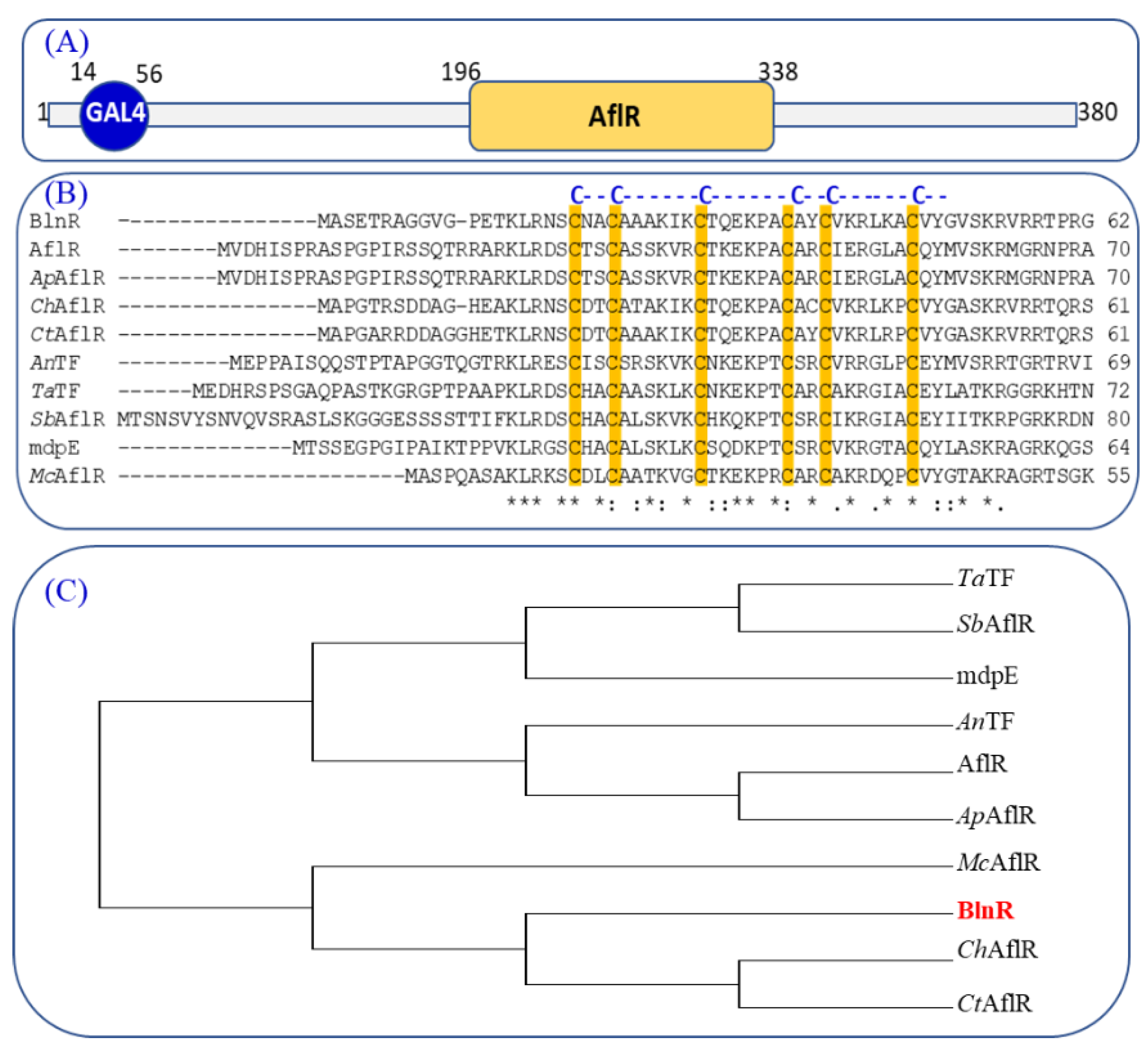
2.5. Phylogenetic Analysis
2.6. mRNA-Seq Analysis and Differential Gene Expression Analysis
2.7. Optimization of Medium Components for Balanol Production by Response Surface Methodology (RSM)
2.8. Analysis of Balanol Production by HPLC
3. Results
3.1. Characterization of Regulator BlnR within the Gene Cluster bln in T. ophioglossoides
3.2. BlnR Positively Regulates Balanol Biosynthesis by Binding All the Promoters of bln Gene Members
3.3. BlnR Is a Positive Regulator toward Balanol Biosynthesis in T. ophioglossoides
3.4. BlnR Is Involved in the Crosstalk between Primary and Secondary Metabolism
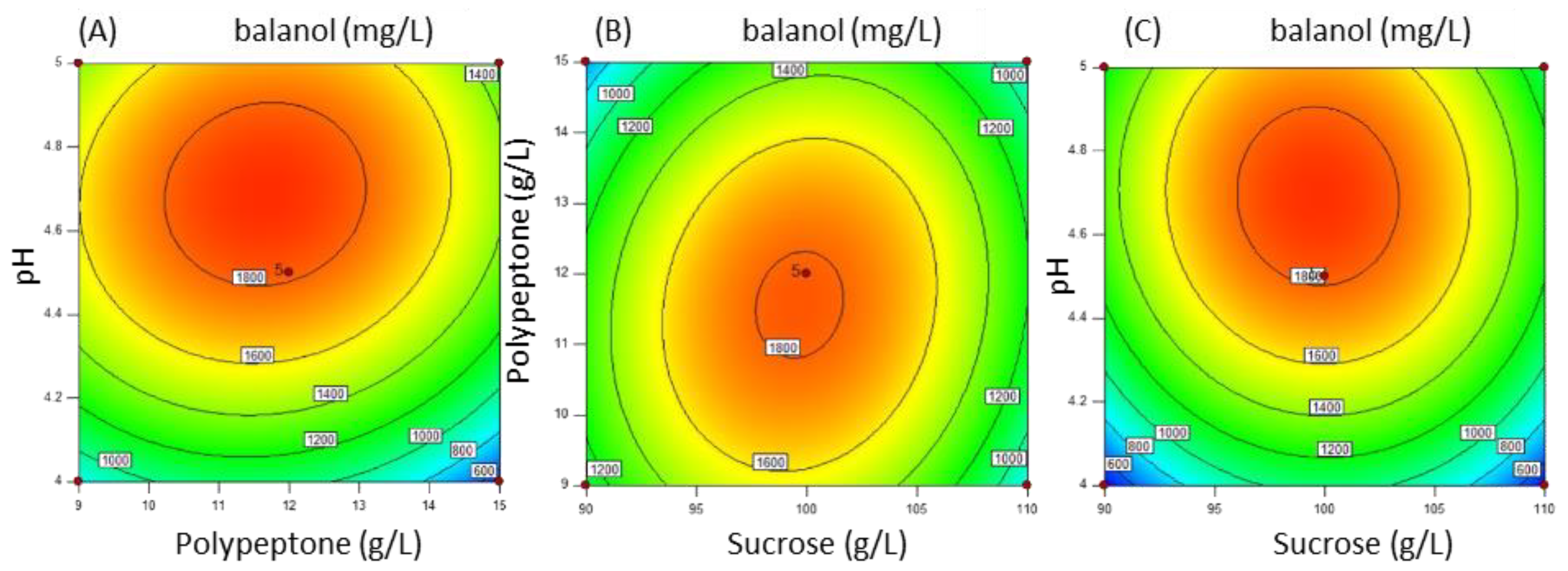
3.5. High Production of Balanol at the Gram Level through Medium Optimization via Response Surface Methodology (RSM)
3.6. Batch Fermentation for Balanol Production in a Scaled-Up 15 L Tank
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kulanthaivel, P.; Hallock, Y.F.; Boros, C.; Hamilton, S.M.; Janzen, W.P.; Ballas, L.M.; Loomis, C.R.; Jiang, J.B. Balanol: A novel and potent inbibitor of protein kinase C from the fungus Verticum balanoides. J. Am. Chem. Soc. 1993, 115, 6452–6453. [Google Scholar] [CrossRef]
- Boros, C.; Hamilton, S.M.; Katz, B.; Kulanthaivel, P. Comparison of balanol from Verticillium balanoides and ophiocordin from Cordyceps ophioglossoides. J. Antibiot. 1994, 9, 1010–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohshima, S.; Yanagisawa, M.; Katoh, A.; Fujii, T.; Sano, T.; Matsukuma, S.; Furumai, T.; Fujiu, M.; Watanabe, K.; Yokose, K. Fusarium merismoides Corda NR 6356, the source of the protein kinase C inhibitor, azepinostatin. Taxonomy, yield improvement, fermentation and biological activity. J. Antibiot. 1994, 47, 639–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kneifel, H.; König, W.A.; Loeffler, W.; Müller, R. Ophiocordin, an antifungal antibiotic of Cordyceps ophioglossoides. Arch. Microbiol. 1977, 113, 121–130. [Google Scholar] [CrossRef]
- Nishizuka, Y. The molecular heterogeneity of protein kinase C and its implications for cellular regulation. Nature 1988, 334, 661–665. [Google Scholar] [CrossRef]
- Castagna, M.; Takai, Y.; Kaibuchi, K.; Sano, K.; Kikkawa, U.; Nishizuka, Y. Direct activation of calcium-activated, phospholipid-dependent protein kinase by tumor-promoting phorbol esters. J. Biol. Chem. 1982, 257, 7847–7851. [Google Scholar] [CrossRef]
- Bradshaw, D.; Hill, C.H.; Nixon, J.S.; Wilkinson, S.E. Therapeutic potential of protein kinase C inhibitors. Agents Actions 1993, 38, 135–147. [Google Scholar] [CrossRef]
- Hirokazu, K.; Kentaro, O.; Fumio, K.; Noriko, K. Estrogenic substances from the mycelia of medicinal fungus Cordyceps ophioglossoides (Ehrh.) Fr. (Ascomycetes). Int. J. Med. Mushrooms 2004, 6, 249–251. [Google Scholar]
- Sun, Y.; Zhao, Z.; Feng, Q.; Xu, Q.; Lv, L.; Liu, J.K.; Zhang, L.; Wu, B.; Li, Y.Q. Unusual spirodecane sesquiterpenes and a sumagillol analogue from Cordyceps ophioglossoides. Helv. Chim. Acta 2013, 96, 76–84. [Google Scholar] [CrossRef]
- He, X.; Zhang, M.; Guo, Y.Y.; Mao, X.M.; Chen, X.A.; Li, Y.Q. Revelation of the balanol biosynthetic pathway in Tolypocladium ophioglossoides. Org. Lett. 2018, 20, 6323–6326. [Google Scholar] [CrossRef]
- Schroeckh, V.; Scherlach, K.; Nützmann, H.W.; Shelest, E.; Schmidt-Heck, W.; Schuemann, J.; Martin, K.; Hertweck, C.; Brakhage, A.A. Intimate bacterial-fungal interaction triggers biosynthesis of archetypal polyketides in Aspergillus nidulans. Proc. Natl. Acad. Sci. USA 2009, 106, 14558–14563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moody, S.C. Microbial co-culture: Harnessing intermicrobial signaling for the production of novel antimicrobials. Future Microbiol. 2014, 9, 575–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, L.Q.; Jin, W.R.; Ma, Z.C.; Shen, Q.; Cai, X.; Liu, Z.Q.; Zheng, Y.G. Promoter engineering strategies for the overproduction of valuable metabolites in microbes. Appl. Microbiol. Biotechnol. 2019, 103, 8725–8736. [Google Scholar] [CrossRef] [PubMed]
- Lopatniuk, M.; Myronovskyi, M.; Nottebrock, A.; Busche, T.; Kalinowski, J.; Ostash, B.; Fedorenko, V.; Luzhetskyy, A. Effect of “ribosome engineering” on the transcription level and production of S. albus indigenous secondary metabolites. Appl. Microbiol. Biotechnol. 2019, 103, 7097–7110. [Google Scholar] [CrossRef] [PubMed]
- Shwab, E.K.; Bok, J.W.; Tribus, M.; Galehr, J.; Graessle, S.; Keller, N.P. Histone deacetylase activity regulates chemical diversity in Aspergillus. Eukaryot. Cell 2007, 6, 1656–1664. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.M.; Xu, W.; Li, D.; Yin, W.B.; Chooi, Y.H.; Li, Y.Q.; Tang, Y.; Hu, Y. Epigenetic genome mining of an endophytic fungus leads to the pleiotropic biosynthesis of natural products. Angew. Chem. Int. Ed. Engl. 2015, 54, 7592–7596. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.M.; Szewczyk, E.; Davidson, A.D.; Entwistle, R.; Keller, N.P.; Wang, C.C.; Oakley, B.R. Characterization of the Aspergillus nidulans monodictyphenone gene cluster. Appl. Environ. Microbiol. 2010, 76, 2067–2074. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.H.; Lin, D.W.; Eames, A.; Chandrasekaran, S. Next-generation genome-scale metabolic modeling through integration of regulatory mechanisms. Metabolites 2021, 11, 606. [Google Scholar] [CrossRef]
- Martín, J.F.; Liras, P. Engineering of regulatory cascades and networks controlling antibiotic biosynthesis in Streptomyces. Curr. Opin. Microbiol. 2010, 13, 263–273. [Google Scholar] [CrossRef]
- Liu, K.; Hu, X.R.; Zhao, L.X.; Wang, Y.; Deng, Z.; Tao, M. Enhancing ristomycin a production by overexpression of ParB-like StrR family regulators controlling the biosynthesis genes. Appl. Environ. Microbiol. 2021, 87, e0106621. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Hewage, R.T.; Aree, T.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. One strain-many compounds (OSMAC) method for production of polyketides, azaphilones, and an isochromanone using the endophytic fungus Dothideomycete sp. Phytochemistry 2014, 108, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) Principle to Marine Microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supratman, U.; Suzuki, T.; Nakamura, T.; Yokoyama, Y.; Harneti, D.; Maharani, R.; Salam, S.; Abdullah, F.F.; Koseki, T.; Shiono, Y. New metabolites produced by endophyte Clonostachys rosea B5-2. Nat. Prod. Res. 2021, 35, 1525–1531. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.A.; He, X.; Zhang, M.; Mao, X.M.; Li, Y.Q. An efficient genetic transformation system for Chinese medicine fungus Tolypocladium ophioglossoides. J. Microbiol. Methods 2020, 176, 106032. [Google Scholar] [CrossRef] [PubMed]
- Hellman, L.M.; Fried, M.G. Electrophoretic mobility shift assay (EMSA) for detecting protein-nucleic acid interactions. Nat. Protoc. 2007, 2, 1849–1861. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.K.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Bennett, J.W.; Linz, J.E.; Woloshuk, C.P.; Payne, G.A. Cloning of the Aspergillus parasiticus apa-2 gene associated with the regulation of aflatoxin biosynthesis. Appl. Environ. Microbiol. 1993, 59, 3273–3279. [Google Scholar] [CrossRef] [Green Version]
- Payne, G.A.; Nystrom, G.J.; Bhatnagar, D.; Cleveland, T.E.; Woloshuk, C.P. Cloning of the afl-2 gene involved in aflatoxin biosynthesis from Aspergillus flavus. Appl. Environ. Microbiol. 1993, 59, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Harrison, S.C. A structural taxonomy of DNA-binding domains. Nature 1991, 353, 715–719. [Google Scholar] [CrossRef]
- Anderson, S.F.; Steber, C.M.; Esposito, R.E.; Coleman, J.E. UME6, a negative regulator of meiosis in Saccharomyces cerevisiae, contains a C-terminal Zn2Cys6 binuclear cluster that binds the URS1 DNA sequence in a zinc-dependent manner. Protein Sci. 1995, 4, 1832–1843. [Google Scholar] [CrossRef] [Green Version]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism from biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, H.; Zhu, Q.Q.; Hao, S.; Chai, S.Y.; Li, Y.H.; Jiao, Z.; Shi, J.C.; Sun, B.G.; Wang, C.T. Overexpression of global regulator LaeA increases secondary metabolite production in Monascus purpureus. Appl. Microbiol. Biotechnol. 2000, 104, 3049–3060. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Guo, F.; Dong, S.H.; Zhao, H. Activation of silent biosynthetic gene clusters using transcription factor decoys. Nat. Chem. Biol. 2019, 15, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Rerngsamran, P.; Murphy, M.B.; Doyle, S.A.; Ebbole, D.J. Fluffy, the major regulator of conidiation in Neurospora crassa, directly activates a developmentally regulated hydrophobin gene. Mol. Microbiol. 2005, 56, 282–297. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Yu, J.H. Genetic control of asexual sporulation in filamentous fungi. Curr. Opin. Microbiol. 2012, 15, 669–677. [Google Scholar] [CrossRef]
- Price, M.S.; Yu, J.; Nierman, W.C.; Kim, H.S.; Pritchard, B.; Jacobus, C.A.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A. The aflatoxin pathway regulator AflR induces gene transcription inside and outside of the aflatoxin biosynthetic cluster. FEMS Microbiol. Lett. 2006, 255, 275–279. [Google Scholar] [CrossRef] [Green Version]
- Kong, Q.; Chang, P.K.; Li, C.; Hu, Z.; Zheng, M.; Sun, Q.; Shan, S. Identification of AflR binding sites in the genome of Aspergillus flavus by ChIP-Seq. J. Fungi 2020, 6, 52. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | A | B | C | D | E | F | G | Balanol (mg/L) |
|---|---|---|---|---|---|---|---|---|
| 1 | 1 | −1 | −1 | −1 | 1 | −1 | 1 | 1218.48 |
| 2 | 1 | −1 | 1 | 1 | −1 | 1 | 1 | 586.54 |
| 3 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | 832.88 |
| 4 | −1 | −1 | −1 | 1 | −1 | 1 | 1 | 633.36 |
| 5 | −1 | 1 | −1 | 1 | 1 | −1 | 1 | 400.22 |
| 6 | −1 | 1 | 1 | 1 | −1 | −1 | −1 | 318.68 |
| 7 | 1 | 1 | −1 | −1 | −1 | 1 | −1 | 638.92 |
| 8 | 1 | 1 | 1 | −1 | −1 | −1 | 1 | 1197.94 |
| 9 | 1 | 1 | −1 | 1 | 1 | 1 | −1 | 579.02 |
| 10 | −1 | −1 | 1 | −1 | 1 | 1 | −1 | 405.68 |
| 11 | 1 | −1 | 1 | 1 | 1 | −1 | −1 | 1633.4 |
| 12 | −1 | 1 | 1 | −1 | 1 | 1 | 1 | 143.46 |
| Gene ID | Protein Name | log2FC |
|---|---|---|
| Starch and sucrose metabolism | ||
| g1571 | putative betaglucosidase I | 1.797 |
| g2561 | putative sucrose utilization protein SUC1 | 1.574 |
| g2754 | Hexokinase-1 | 1.715 |
| g6339 | Alphaamylase A type-3 | 1.338 |
| g6369 | Alphaglucosidase | 1.387 |
| g6411 | Endoglucanase EG-II | −1.405 |
| g6822 | Probable betaglucosidase A | 1.301 |
| g7696 | alphatrehalose-phosphate synthase | 1.388 |
| g857 | endo-1,3-betaglucosidase eglC | 1.360 |
| g9295 | Glucose-6-phosphate isomerase | −1.422 |
| Fatty acid metabolism | ||
| g507 | Cytochrome P450 | −2.278 |
| g672 | putative aldehyde dehydrogenase | −1.201 |
| g2909 | 3-ketoacyl-CoA thiolase | −1.931 |
| g3641 | Acetyl-CoA acetyltransferase | −1.198 |
| g3915 | Enoyl-CoA isomerase/hydratase | −1.153 |
| g5070 | Enoyl-(Acyl carrier protein) reductase | 2.751 |
| g5094 | Short-chain-type dehydrogenase/reductase | 11.281 |
| g5215 | Acyl-CoA dehydrogenase family member 10 | 1.212 |
| g6087 | Phosphotransferase | −1.056 |
| g6743 | Aldehyde dehydrogenase | 1.447 |
| g7131 | Acetoacetyl-CoA reductase | 1.059 |
| g7814 | Isotrichodermin C-15 hydroxylase | −1.498 |
| g9557 | Short/branched-chain-specific acyl-CoA dehydrogenase | −1.433 |
| Citrate cycle (TCA cycle) | ||
| g3392 | 2-methylcitrate synthase | −1.435 |
| g8077 | Succinyl-CoA ligase | 1.935 |
| g9576 | putative succinate dehydrogenase | −1.168 |
| Lysine biosynthesis | ||
| g3241 | Homoaconitase, mitochondrial | 1.077 |
| g6260 | Homocitrate synthase, mitochondrial | 1.129 |
| Phenylalanine, tyrosine and tryptophan biosynthesis | ||
| g6745 | Fungal-specific transcription factor | 2.344 |
| MAPK signaling pathway | ||
| g7635 | Catalase | −1.13221 |
| g9623 | Catalase | −3.62842 |
| Spore development | ||
| g6286 | Spore development regulator vosA | −1.22895 |
| g655 | Outer spore wall protein RRT8 | −0.94555 |
| Source | SS | DF | MS | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 4.336 × 105 | 7 | 6.821 × 105 | 6.82 | 0.0413 * |
| A | 2.028 × 105 | 1 | 2.028 × 105 | 20.27 | 0.0108 * |
| B | 8.603 × 104 | 1 | 8.603 × 105 | 8.60 | 0.04727 |
| C | 4.005 × 105 | 1 | 6.15 | 6.145 × 10−4 | 0.9814 |
| D | 1.706 × 103 | 1 | 1.706 × 103 | 0.12 | 0.7009 |
| E | 5.495 × 104 | 1 | 6.159 × 102 | 0.044 | 0.8163 |
| F | 6.16 × 102 | 1 | 1.424 × 105 | 10.18 | 0.0188 * |
| G | 1.424 × 105 | 1 | 4.390 × 105 | 1.55 | 0.2677 |
| Residual | 4.003 × 105 | 4 | 1.001 × 105 | ||
| Cor Total | 5.175 × 105 | 11 |
| Source | SS | DF | MS | F-Value | Pr > F |
|---|---|---|---|---|---|
| Model | 4.148 × 106 | 9 | 4.609 × 105 | 32.35 | <0.0001 ** |
| A | 3.715 × 104 | 1 | 3.715 × 104 | 0.26 | 0.6253 |
| B | 8.506 × 105 | 1 | 8.506 × 105 | 5.97 | 0.0445 * |
| C | 8.869 × 105 | 1 | 8.869 × 105 | 62.25 | <0.0001 ** |
| AB | 4.618 × 105 | 1 | 4.618 × 105 | 3.24 | 0.1148 |
| AC | 4.575 × 103 | 1 | 4.575 × 103 | 0.32 | 0.5886 |
| BC | 1.174 × 105 | 1 | 1.174 × 105 | 0.82 | 0.3942 |
| A2 | 1.441 × 106 | 1 | 1.441 × 106 | 101.15 | <0.0001 ** |
| B2 | 5.614 × 105 | 1 | 5.614 × 105 | 39.40 | 0.0004 ** |
| C2 | 7.957 × 105 | 1 | 7.957 × 105 | 55.85 | 0.0001 ** |
| Lack of Fit | 2.485 × 105 | 3 | 8.283 × 103 | 0.44 | 0.7355 |
| Pure Error | 7.488 × 105 | 4 | 1.872 × 105 | ||
| Cor Total | 4.248 × 106 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, R.-Q.; Liu, X.; Zhang, M.; Xu, W.-Q.; Li, Y.-Q.; Chen, X.-A. Gram-Level Production of Balanol through Regulatory Pathway and Medium Optimization in Herb Fungus Tolypocladium ophioglossoides. J. Fungi 2022, 8, 510. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050510
Li R-Q, Liu X, Zhang M, Xu W-Q, Li Y-Q, Chen X-A. Gram-Level Production of Balanol through Regulatory Pathway and Medium Optimization in Herb Fungus Tolypocladium ophioglossoides. Journal of Fungi. 2022; 8(5):510. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050510
Chicago/Turabian StyleLi, Rui-Qi, Xiang Liu, Min Zhang, Wei-Qun Xu, Yong-Quan Li, and Xin-Ai Chen. 2022. "Gram-Level Production of Balanol through Regulatory Pathway and Medium Optimization in Herb Fungus Tolypocladium ophioglossoides" Journal of Fungi 8, no. 5: 510. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050510