
Encephalartos villosus Lem. Displays a Strong In Vivo and In Vitro Antifungal Potential against Candida glabrata Clinical Isolates

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Materials
2.3. Animals
2.4. LC–MS/MS for Metabolite Profiling
2.5. In Vitro Antifungal Activity of EVME
2.5.1. Fungi
2.5.2. Antifungal Susceptibility
2.5.3. Determination of Minimum Inhibitory Concentrations (MICs)
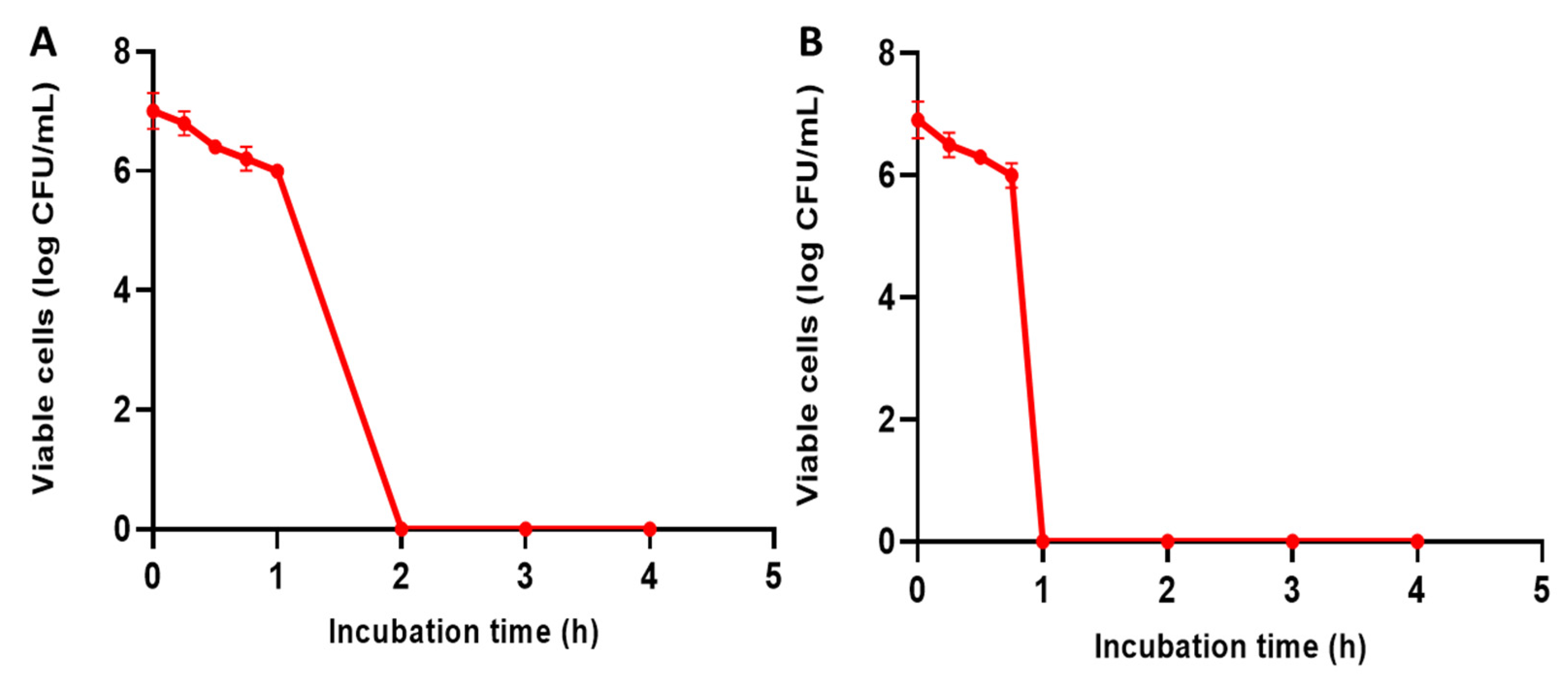
2.5.4. Time–Kill Curve
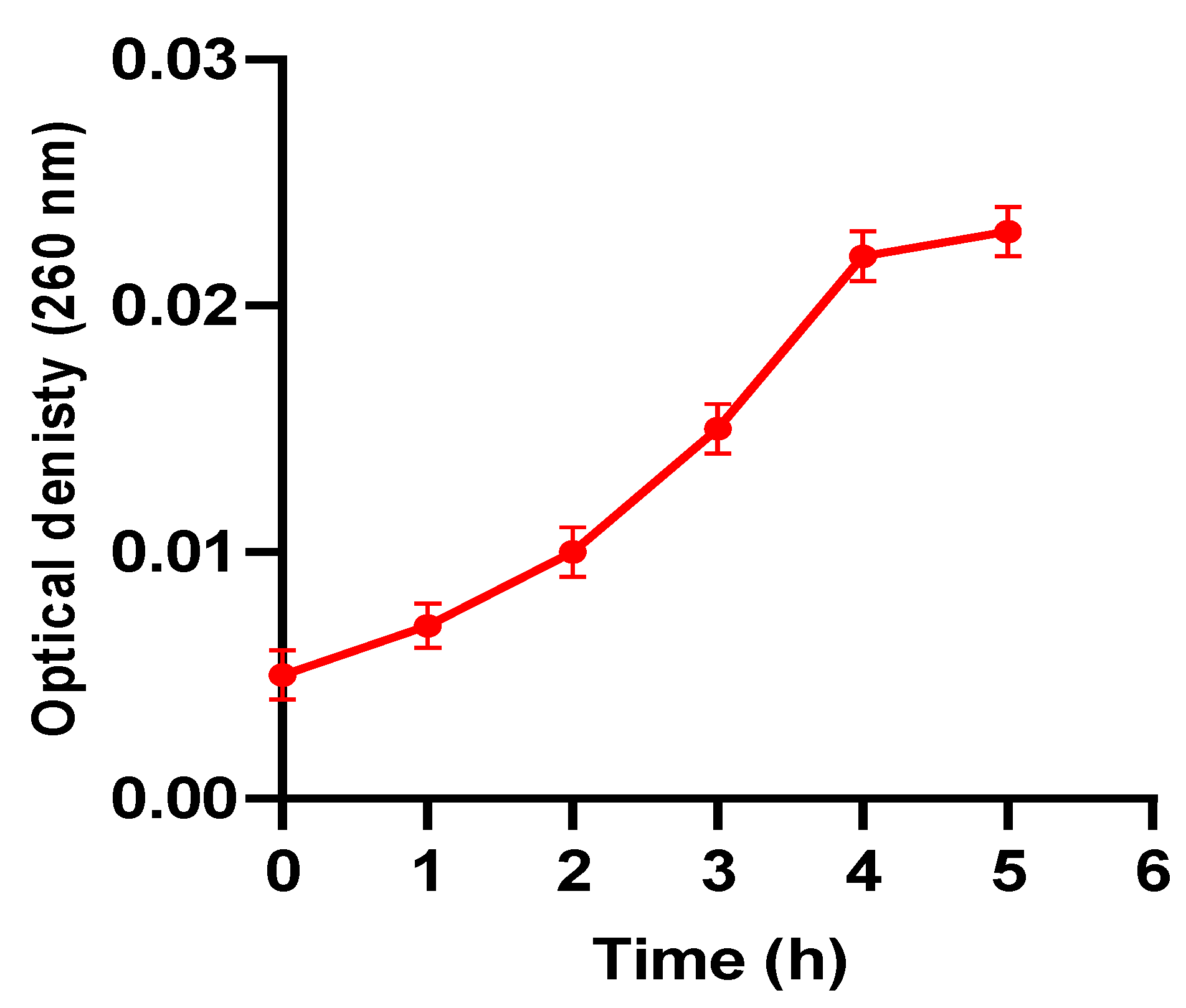
2.5.5. Nucleotide Leakage
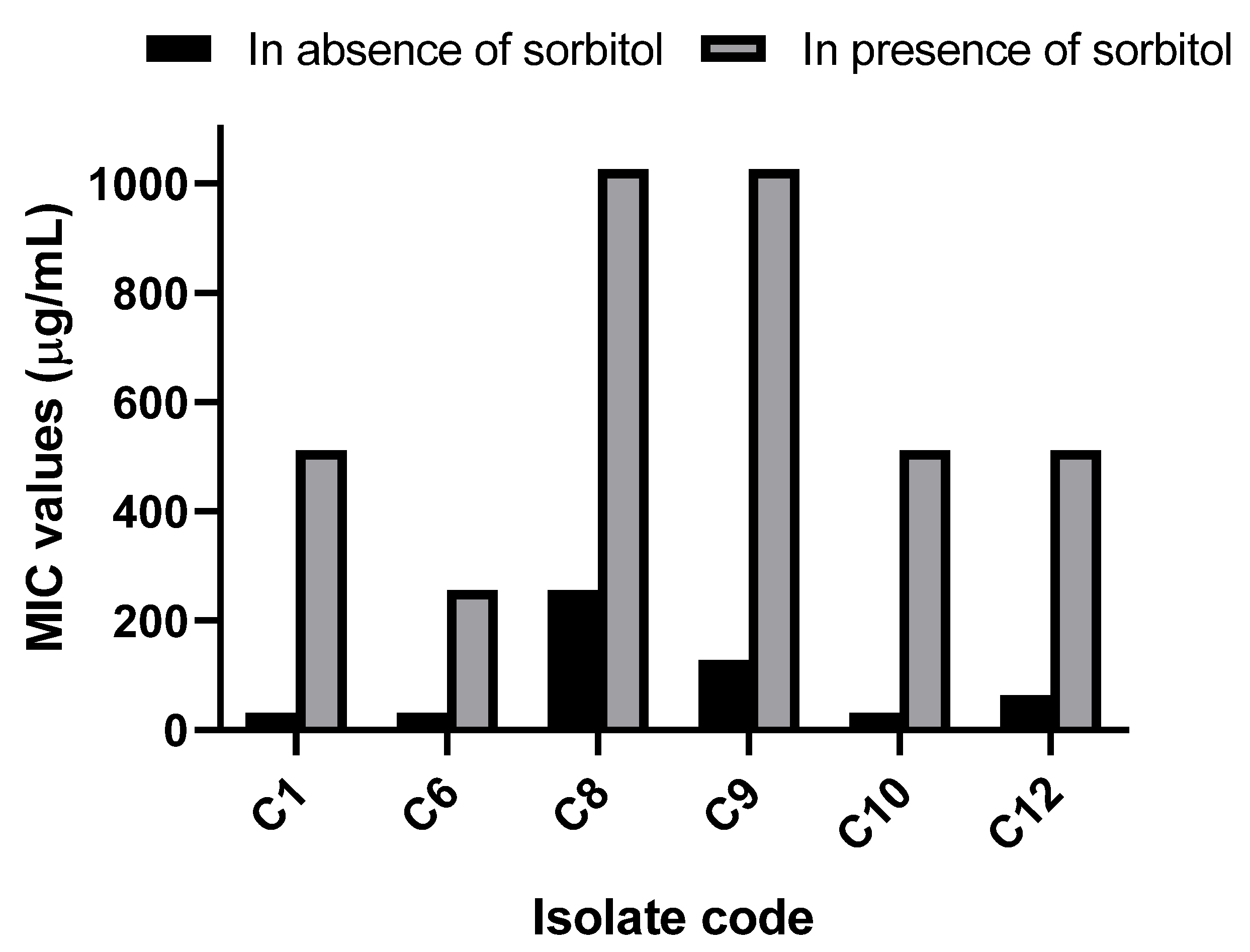
2.5.6. Sorbitol Protection Assay
2.5.7. Phenotypic Detection of Efflux Pump Inhibition
2.5.8. Quantitative Real-Time PCR (qRT-PCR) for Detection of the Expression of the Efflux Pump Genes
2.6. In Vivo Antifungal Activity of EVME
2.6.1. Experimental Protocol
2.6.2. Kidney Function Tests
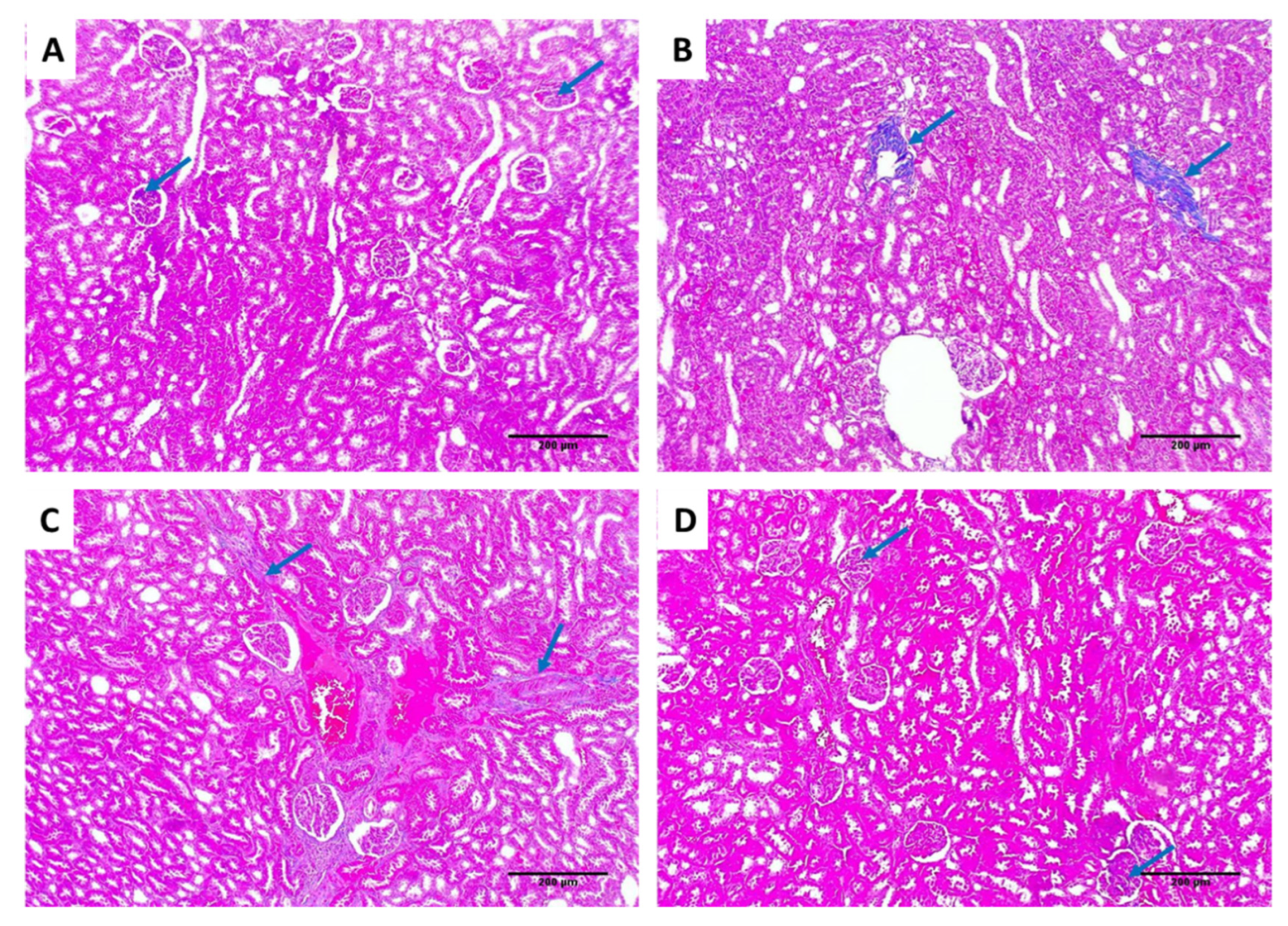
2.6.3. Histological Assessment
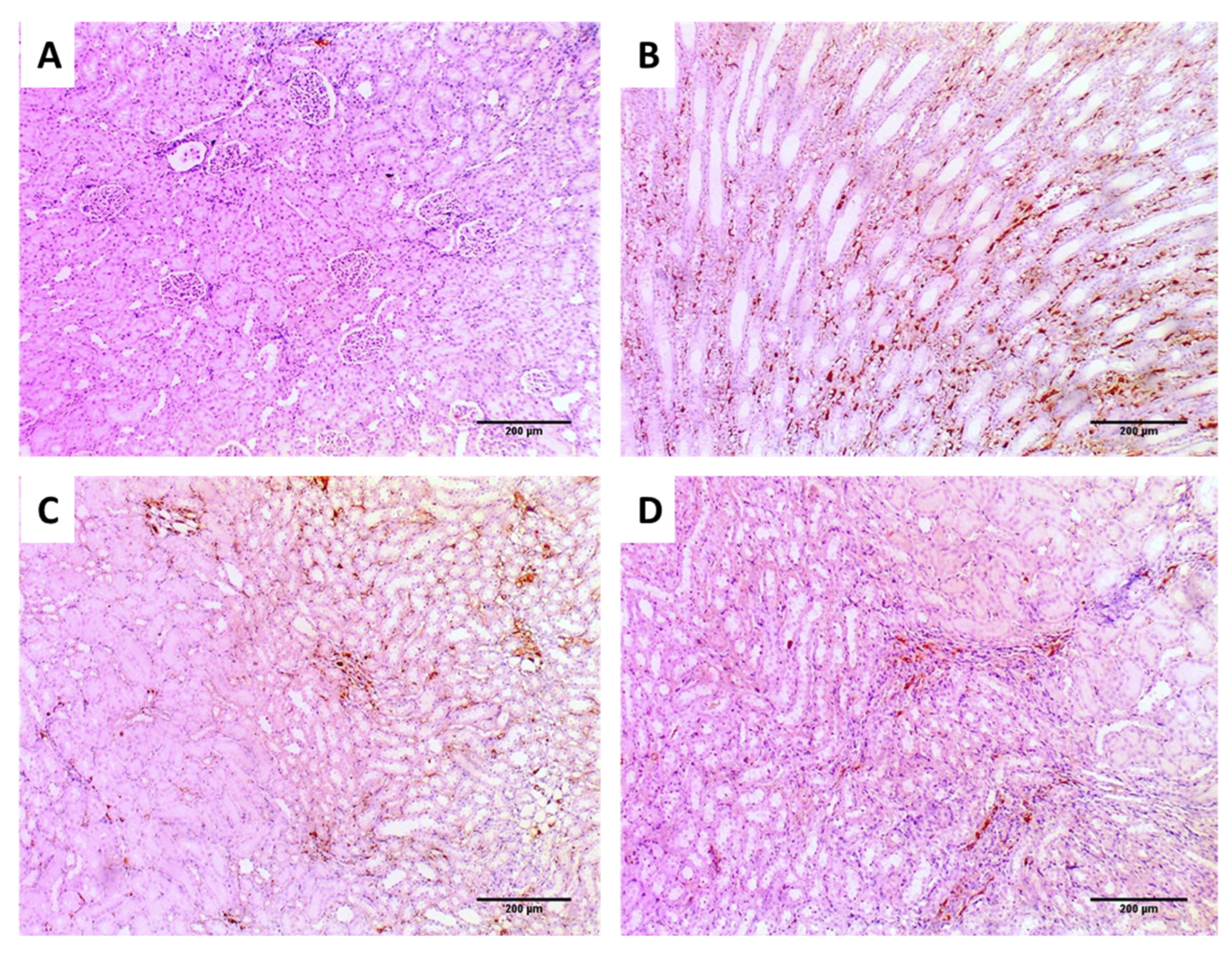
2.6.4. Immunohistochemical Studies
2.7. Statistical Analysis
3. Results
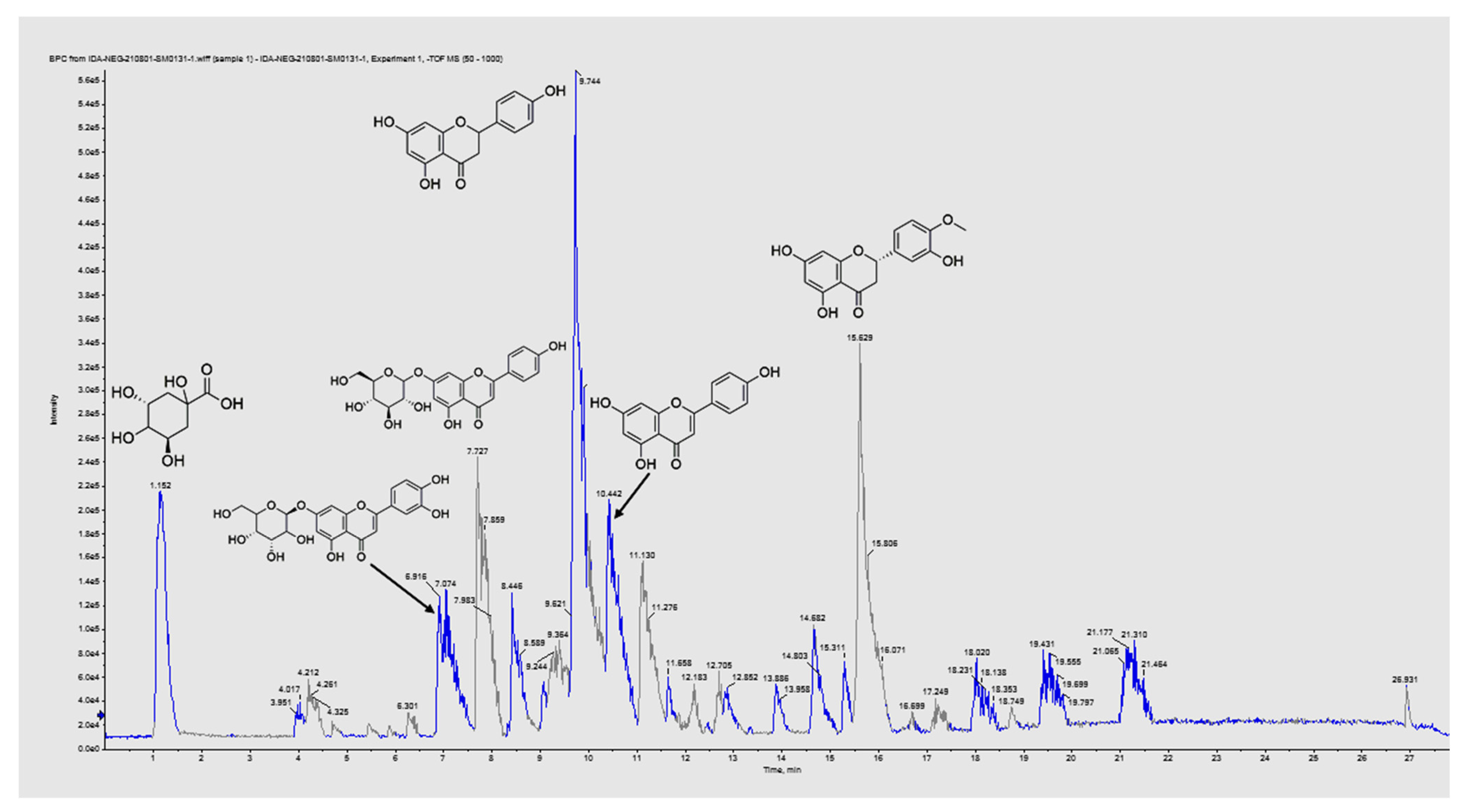
3.1. Phytochemical Profiling of EVME
3.1.1. Identification of Carboxylic, Phenolic, and Fatty Acids
3.1.2. Identification of Flavonoid Derivatives
3.1.3. Identification of Coumarins and Other Derivatives
3.2. In Vitro Antifungal Activity
3.2.1. Antifungal Susceptibility Testing and Determination of MIC
3.2.2. Time–Kill Study
3.2.3. Nucleotide Leakage
3.2.4. Sorbitol Protection Assay
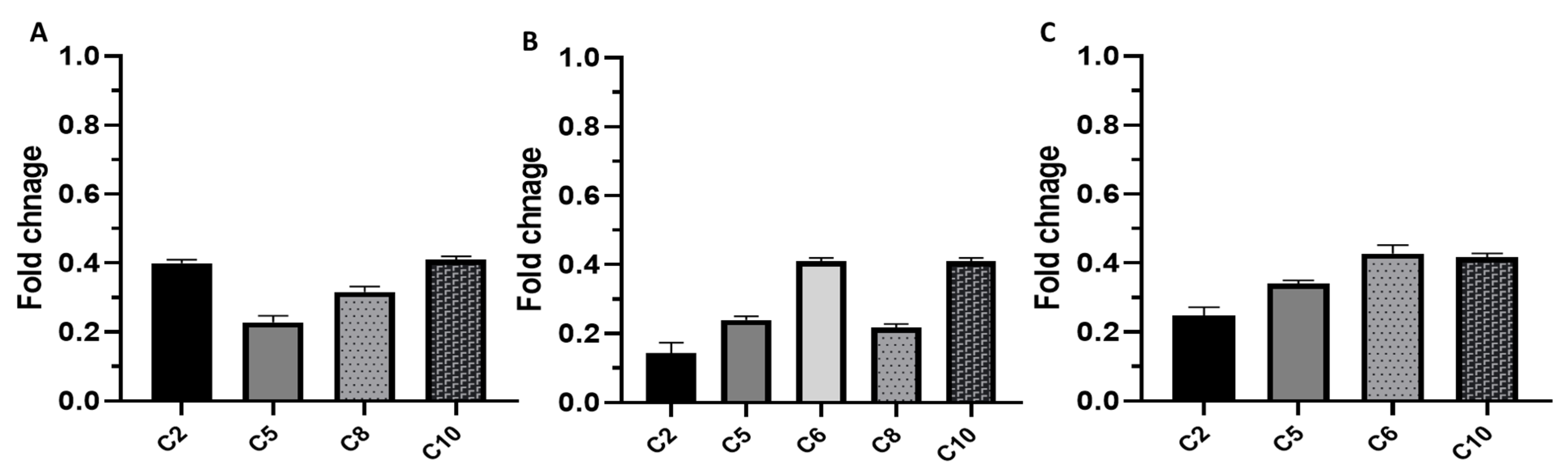
3.2.5. Phenotypic and Genotypic Detection of the Impact of EVME on Efflux Pumps
3.3. In Vivo Antifungal Activity
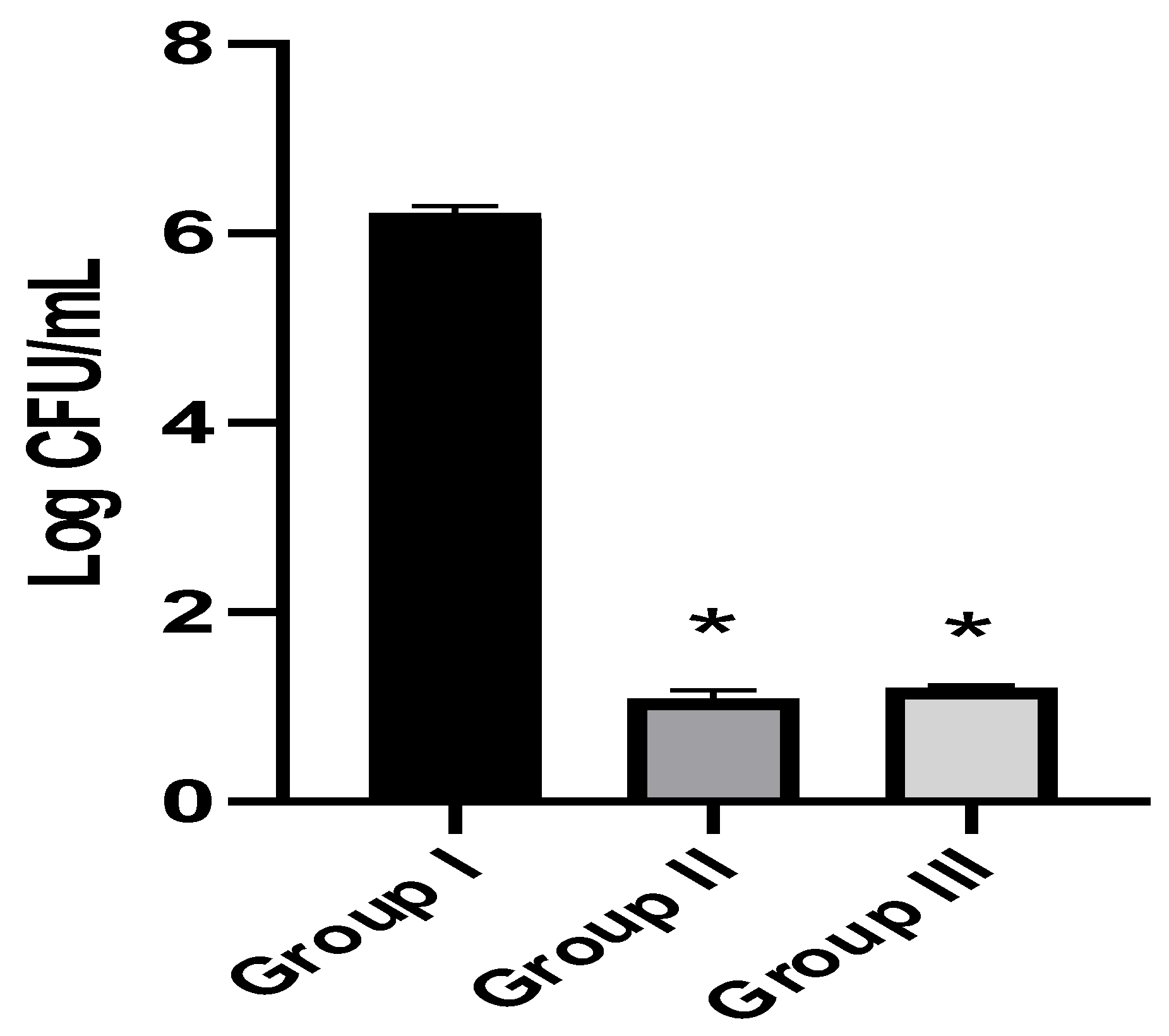
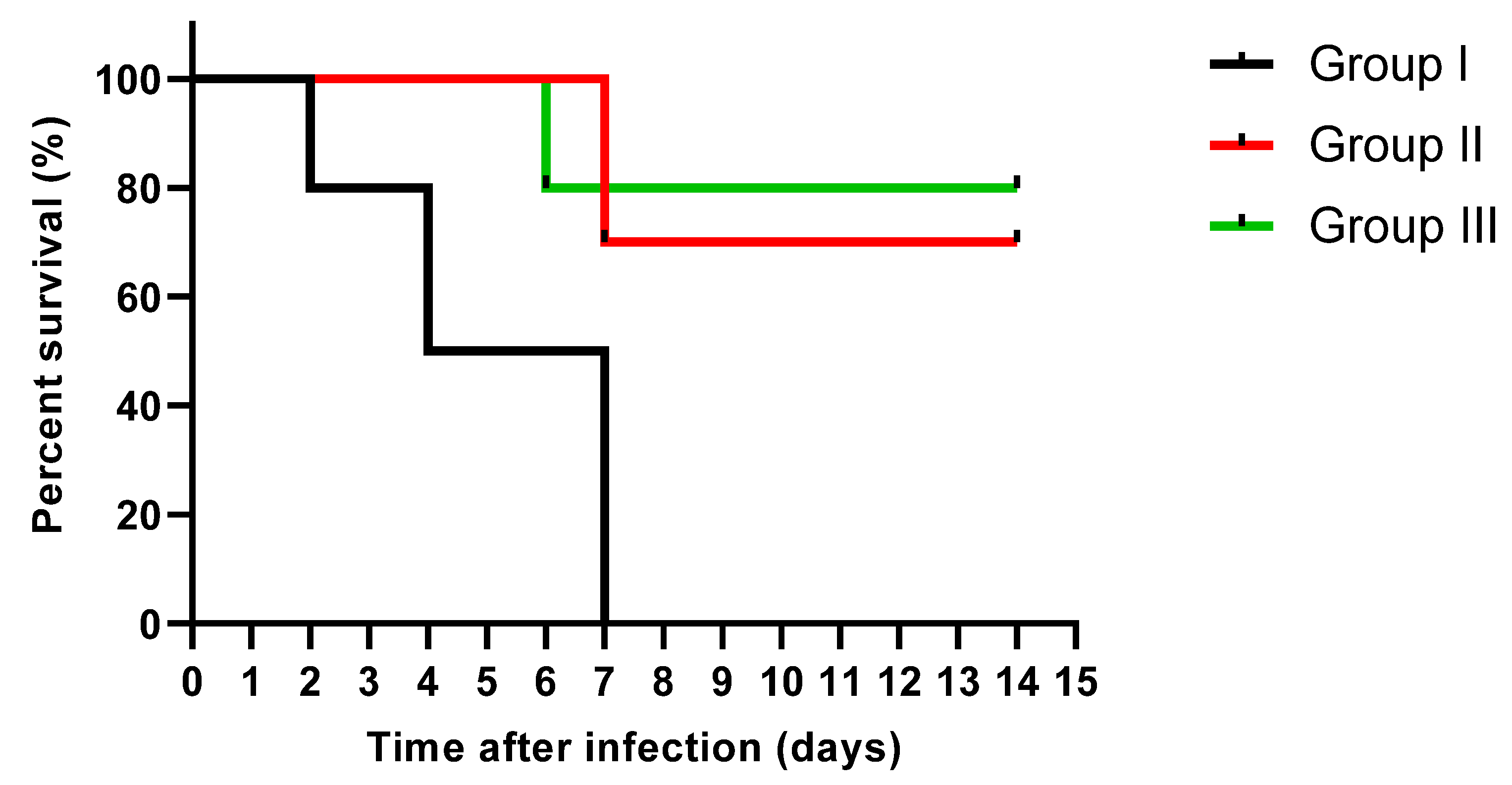
3.3.1. Fungal Burden in the Kidney Tissues and Survival Rate
3.3.2. Kidney Function Test
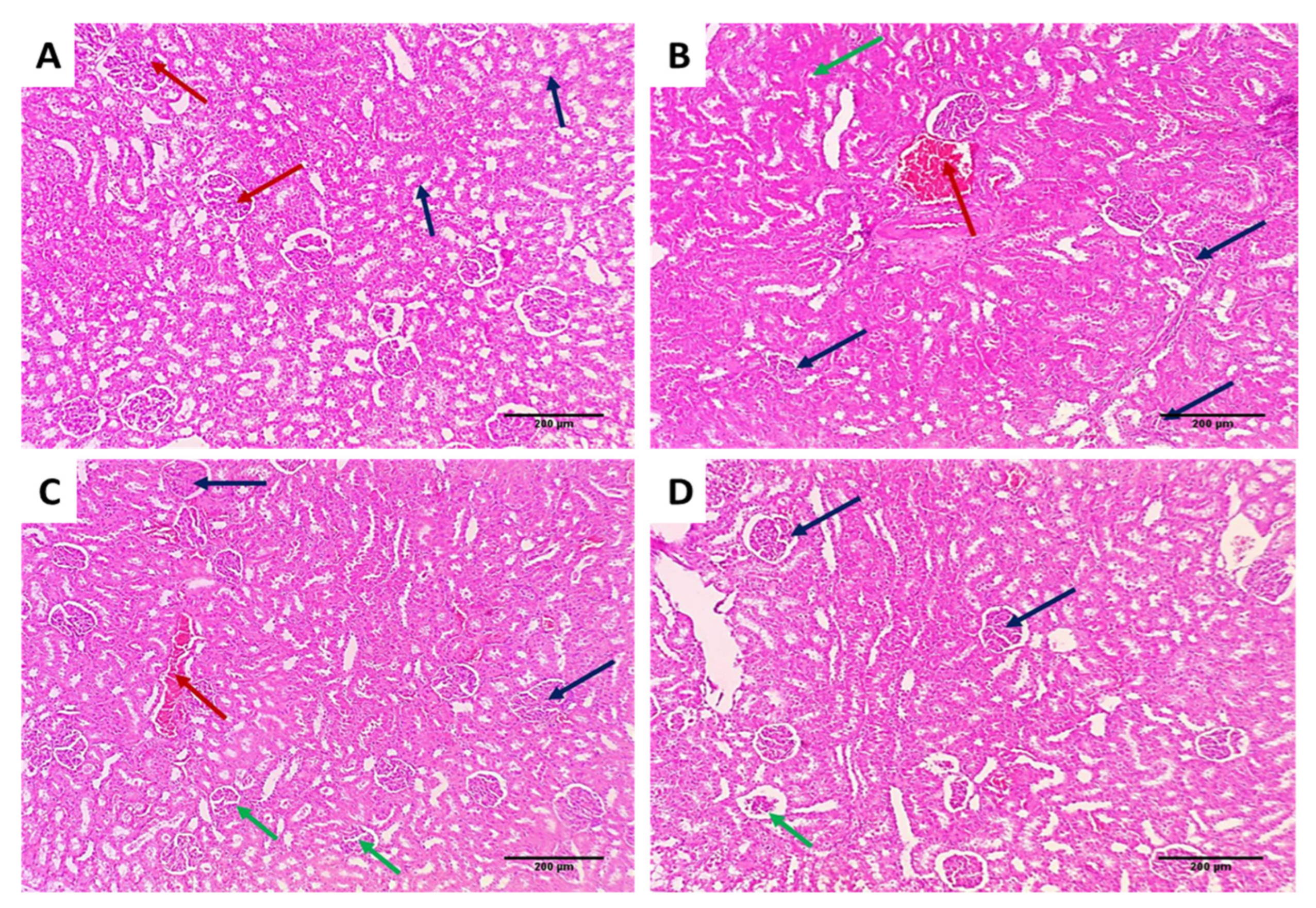
3.3.3. Histological Assessment
3.3.4. Immunohistochemical Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arastehfar, A.; Gabaldón, T.; Garcia-Rubio, R.; Jenks, J.D.; Hoenigl, M.; Salzer, H.J.; Ilkit, M.; Lass-Flörl, C.; Perlin, D.S. Drug-Resistant Fungi: An Emerging Challenge Threatening Our Limited Antifungal Armamentarium. Antibiotics 2020, 9, 877. [Google Scholar] [CrossRef] [PubMed]
- Talapko, J.; Juzbašić, M.; Matijević, T.; Pustijanac, E.; Bekić, S.; Kotris, I.; Škrlec, I. Candida albicans—The Virulence Factors and Clinical Manifestations of Infection. J. Fungi 2021, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.R.; Cardno, T.S.; Strouse, J.J.; Ivnitski-Steele, I.; Keniya, M.V.; Lackovic, K.; Monk, B.C.; Sklar, L.A.; Cannon, R.D. Targeting efflux pumps to overcome antifungal drug resistance. Future Med. Chem. 2016, 8, 1485–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Ganiny, A.M.; Kamel, H.A.; Yossef, N.E.; Mansour, B.; El-Baz, A.M. Repurposing pantoprazole and haloperidol as efflux pump inhibitors in azole resistant clinical Candida albicans and non-albicans isolates. Saudi Pharm. J. 2022, 30, 245–255. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Santos, G.C.; Vasconcelos, C.C.; Lopes, A.J.; de Sousa Cartágenes, M.D.S.; Filho, A.K.; do Nascimento, F.R.; de Andrade Monteiro, C. Candida Infections and Therapeutic Strategies: Mechanisms of Action for Traditional and Alternative Agents. Front Microbiol. 2018, 9, 1351. [Google Scholar] [CrossRef] [PubMed]
- El-Shahir, A.A.; El-Wakil, D.A.; Abdel Latef, A.A.H.; Youssef, N.H. Bioactive Compounds and Antifungal Activity of Leaves and Fruits Methanolic Extracts of Ziziphus spina-christi L. Plants 2022, 11, 746. [Google Scholar] [CrossRef]
- Alhassan, K.A.; Indabawa, A.S.; Shah, M.M. Phytochemical analysis, proximate composition and antibacterial activities of Ziziphus species (Z. jujube and Z. spina-christi). J. Appl. Adv. Res. 2019, 4, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and Other Phenolic Compounds from Medicinal Plants for Pharmaceutical and Medical Aspects: An Overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef]
- Mahlo, S.M.; Chauke, H.R.; McGaw, L.; Eloff, J. Antioxidant and antifungal activity of selected medicinal plant extracts against phytopathogenic fungi. Afr. J. Tradit. Complementary Altern. Med. 2016, 13, 216–222. [Google Scholar] [CrossRef] [Green Version]
- Treutlein, J.; Vorster, P.; Wink, M. Molecular relationships in Encephalartos (Zamiaceae, Cycadales) based on nucleotide sequences of nuclear ITS 1&2, rbcL, and genomic ISSR fingerprinting. Plant Biol. 2005, 7, 79–90. [Google Scholar]
- Negm, W.A.; El-Aasr, M.; Attia, G.; Alqahtani, M.J.; Yassien, R.I.; Abo Kamer, A.; Elekhnawy, E. Promising Antifungal Activity of Encephalartos laurentianus de Wild against Candida albicans Clinical Isolates: In Vitro and In Vivo Effects on Renal Cortex of Adult Albino Rats. J. Fungi 2022, 8, 426. [Google Scholar] [CrossRef]
- Temraz, A. Novel illudalane sesquiterpenes from Encephalartos villosus Lehm. antimicrobial activity. Nat. Prod. Res. 2016, 30, 2791–2797. [Google Scholar] [CrossRef] [PubMed]
- Negm, W.A.; El-Kadem, A.H.; Elekhnawy, E.; Attallah, N.G.; Al-Hamoud, G.A.; El-Masry, T.A.; Zayed, A. Wound-Healing Potential of Rhoifolin-Rich Fraction Isolated from Sanguisorba officinalis Roots Supported by Enhancing Re-Epithelization, Angiogenesis, Anti-Inflammatory, and Antimicrobial Effects. Pharmaceuticals 2022, 15, 178. [Google Scholar] [CrossRef] [PubMed]
- Alotaibi, B.; Mokhtar, F.A.; El-Masry, T.A.; Elekhnawy, E.; Mostafa, S.A.; Abdelkader, D.H.; Elharty, M.E.; Saleh, A.; Negm, W.A. Antimicrobial Activity of Brassica rapa L. Flowers Extract on Gastrointestinal Tract Infections and Antiulcer Potential Against Indomethacin-Induced Gastric Ulcer in Rats Supported by Metabolomics Profiling. J. Inflamm. Res. 2021, 14, 7411. [Google Scholar] [CrossRef]
- Attallah, N.G.M.; Negm, W.A.; Elekhnawy, E.; Elmongy, E.I.; Altwaijry, N.; El-Haroun, H.; El-Masry, T.A.; El-Sherbeni, S.A. Elucidation of Phytochemical Content of Cupressus macrocarpa Leaves: In Vitro and In Vivo Antibacterial Effect against Methicillin-Resistant Staphylococcus aureus Clinical Isolates. Antibiotics 2021, 10, 890. [Google Scholar] [CrossRef]
- Alotaibi, B.; Negm, W.A.; Elekhnawy, E.; El-Masry, T.A.; Elseady, W.S.; Saleh, A.; Alotaibi, K.N.; El-Sherbeni, S.A. Antibacterial, Immunomodulatory, and Lung Protective Effects of Boswelliadalzielii Oleoresin Ethanol Extract in Pulmonary Diseases: In Vitro and In Vivo Studies. Antibiotics 2021, 10, 1444. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antifungal Susceptibility Testing of Yeasts. CLSI Supplement M60 2017. Available online: https://clsi.org/media/1895/m60ed1_sample.pdf (accessed on 2 January 2022).
- Lemos, A.S.O.; Florêncio, J.R.; Pinto, N.C.C.; Campos, L.M.; Silva, T.P.; Grazul, R.M.; Pinto, P.F.; Tavares, G.D.; Scio, E.; Apolônio, A.C.M.; et al. Antifungal Activity of the Natural Coumarin Scopoletin Against Planktonic Cells and Biofilms from a Multidrug-Resistant Candida tropicalis Strain. Front. Microbiol. 2020, 11, 1525. [Google Scholar] [CrossRef]
- Negm, W.A.; Abo El-Seoud, K.A.; Kabbash, A.; Kassab, A.A.; El-Aasr, M. Hepatoprotective, cytotoxic, antimicrobial and antioxidant activities of Dioon spinulosum leaves Dyer Ex Eichler and its isolated secondary metabolites. Nat. Prod. Res. 2021, 35, 5166–5176. [Google Scholar] [CrossRef]
- Khudhair, D.H.; Al-Gareeb, A.I.; Al-Kuraishy, H.M.; El-Kadem, A.H.; Elekhnawy, E.; Negm, W.A.; Saber, S.; Cavalu, S.; Tirla, A.; Alotaibi, S.S. Combination of Vitamin C and Curcumin Safeguards against Methotrexate-Induced Acute Liver Injury in Mice by Synergistic Antioxidant Effects. Front. Med. 2022, 9, 866343. [Google Scholar] [CrossRef]
- Salgado, F.L.; Artigiani-Neto, R.; Lopes-Filho, G.d.J. Growth factors and COX2 in wound healing: An experimental study with Ehrlich tumors. ABCD. Arq. Bras. De Cir. Dig. (São Paulo) 2016, 29, 223–226. [Google Scholar] [CrossRef] [Green Version]
- Al Kadhi, O.; Melchini, A.; Mithen, R.; Saha, S. Development of a LC-MS/MS method for the simultaneous detection of tricarboxylic acid cycle intermediates in a range of biological matrices. J. Anal. Methods Chem. 2017, 2017, 5391832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avula, B.; Wang, Y.-H.; Smillie, T.J.; Khan, I.A. Determination of shikimic acid in fruits of Illicium species and various other plant samples by LC–UV and LC–ESI–MS. Chromatographia 2009, 69, 307–314. [Google Scholar] [CrossRef]
- Ruan, J.; Yan, J.; Zheng, D.; Sun, F.; Wang, J.; Han, L.; Zhang, Y.; Wang, T. Comprehensive chemical profiling in the ethanol extract of Pluchea indica aerial parts by liquid chromatography/mass spectrometry analysis of its silica gel column chromatography fractions. Molecules 2019, 24, 2784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosco, R.; Daeseleire, E.; Van Pamel, E.; Scariot, V.; Leus, L. Development of an ultrahigh-performance liquid chromatography–electrospray ionization–tandem mass spectrometry method for the simultaneous determination of salicylic acid, jasmonic acid, and abscisic acid in rose leaves. J. Agric. Food Chem. 2014, 62, 6278–6284. [Google Scholar] [CrossRef] [PubMed]
- Serrano, C.A.; Villena, G.K.; Rodríguez, E.F. Phytochemical profile and rosmarinic acid purification from two Peruvian Lepechinia Willd. species (Salviinae, Mentheae, Lamiaceae). Sci. Rep. 2021, 11, 7260. [Google Scholar] [CrossRef]
- Yuan, Y.; Hou, W.; Tang, M.; Luo, H.; Chen, L.-J.; Guan, Y.H.; Sutherland, I.A. Separation of flavonoids from the leaves of Oroxylum indicum by HSCCC. Chromatographia 2008, 68, 885–892. [Google Scholar] [CrossRef]
- El Sayed, A.M.; Basam, S.M.; Marzouk, H.S.; El-Hawary, S. LC–MS/MS and GC–MS profiling as well as the antimicrobial effect of leaves of selected Yucca species introduced to Egypt. Sci. Rep. 2020, 10, 17778. [Google Scholar] [CrossRef]
- Xiao, Y.; Hu, Z.; Yin, Z.; Zhou, Y.; Liu, T.; Zhou, X.; Chang, D. Profiling and distribution of metabolites of procyanidin B2 in mice by UPLC-DAD-ESI-IT-TOF-MSn technique. Front. Pharmacol. 2017, 8, 231. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.A.; dos Santos, L.R.; de Freitas, J.S.; de Sousa, R.P.; de Farias, R.R.S.; Júnior, G.M.V.; Rai, M.; Chaves, M.H. First report of flavonoids from leaves of Machaerium acutifolium by DI-ESI-MS/MS. Arab. J. Chem. 2022, 15, 103765. [Google Scholar] [CrossRef]
- Yuzuak, S.; Ballington, J.; Xie, D.-Y. HPLC-qTOF-MS/MS-based profiling of flavan-3-ols and dimeric proanthocyanidins in berries of two muscadine grape hybrids FLH 13-11 and FLH 17-66. Metabolites 2018, 8, 57. [Google Scholar] [CrossRef] [Green Version]
- Aouadi, K.; Hajlaoui, H.; Arraouadi, S.; Ghannay, S.; Snoussi, M.; Kadri, A. HPLC/MS Phytochemical Profiling with Antioxidant Activities of Echium humile Desf. Extracts: ADMET Prediction and Computational Study Targeting Human Peroxiredoxin 5 Receptor. Agronomy 2021, 11, 2165. [Google Scholar] [CrossRef]
- Šibul, F.; Orčić, D.; Berežni, S.; Anačkov, G.; Mimica-Dukić, N. HPLC–MS/MS profiling of wild-growing scentless chamomile. Acta Chromatographica 2020, 32, 86–94. [Google Scholar] [CrossRef]
- Carolus, H.; Van Dyck, K.; Van Dijck, P. Candida albicans and Staphylococcus species: A threatening twosome. Front. Microbiol. 2019, 10, 2162. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.A.; Rorato, V.C.; Almeida-Apolonio, A.A.; Rodrigues, A.B.; Barros, A.L.; Sangalli, A.; Arena, A.C.; Mota, J.S.; Grisolia, A.B.; Oliveira, K.M.D. In vitro antifungal activity of Myracrodruon urundeuva Allemão against human vaginal Candida species. An. Acad. Bras. Cienc. 2017, 89, 2423–2432. [Google Scholar] [CrossRef] [Green Version]
- Bonifácio, B.V.; dos Santos Ramos, M.A.; da Silva, P.B.; Negri, K.M.S.; de Oliveira Lopes, É.; de Souza, L.P.; Vilegas, W.; Pavan, F.R.; Chorilli, M.; Bauab, T.M. Nanostructured lipid system as a strategy to improve the anti-Candida albicans activity of Astronium sp. Int. J. Nanomed. 2015, 10, 5081. [Google Scholar] [CrossRef] [Green Version]
- Negm, W.A.; El-Aasr, M.; Kamer, A.A.; Elekhnawy, E. Investigation of the Antibacterial Activity and Efflux Pump Inhibitory Effect of Cycas thouarsii R. Br. Extract against Klebsiella pneumoniae Clinical Isolates. Pharmaceuticals 2021, 14, 756. [Google Scholar] [CrossRef]
- Elekhnawy, E.; Negm, W.A.; El-Aasr, M.; Kamer, A.A.; Alqarni, M.; Batiha, G.E.-S.; Obaidullah, A.J.; Fawzy, H.M. Histological assessment, anti-quorum sensing, and anti-biofilm activities of Dioon spinulosum extract: In vitro and in vivo approach. Sci. Rep. 2022, 12, 180. [Google Scholar] [CrossRef]
- Kaur, N.; Bains, A.; Kaushik, R.; Dhull, S.B.; Melinda, F.; Chawla, P. A Review on Antifungal Efficiency of Plant Extracts Entrenched Polysaccharide-Based Nanohydrogels. Nutrients 2021, 13, 2055. [Google Scholar] [CrossRef]
- Ayoub, I.M.; Korinek, M.; Hwang, T.-L.; Chen, B.-H.; Chang, F.-R.; El-Shazly, M.; Singab, A.N.B. Probing the antiallergic and anti-inflammatory activity of biflavonoids and dihydroflavonols from Dietes bicolor. J. Nat. Prod. 2018, 81, 243–253. [Google Scholar] [CrossRef]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2010, 69, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef] [PubMed]
- Grover, J.; Jachak, S.M. Coumarins as privileged scaffold for anti-inflammatory drug development. RSC Adv. 2015, 5, 38892–38905. [Google Scholar] [CrossRef]
- Qian, W.; Liu, M.; Fu, Y.; Zhang, J.; Liu, W.; Li, J.; Li, X.; Li, Y.; Wang, T. Antimicrobial mechanism of luteolin against Staphylococcus aureus and Listeria monocytogenes and its antibiofilm properties. Microb. Pathog. 2020, 142, 104056. [Google Scholar] [CrossRef] [PubMed]
- Attallah, N.G.; El-Sherbeni, S.A.; El-Kadem, A.H.; Elekhnawy, E.; El-Masry, T.A.; Elmongy, E.I.; Altwaijry, N.; Negm, W.A. Elucidation of the Metabolite Profile of Yucca gigantea and Assessment of its Cytotoxic, Antimicrobial, and Anti-Inflammatory Activities. Molecules 2022, 27, 1329. [Google Scholar] [CrossRef] [PubMed]
- Buchmann, D.; Schultze, N.; Borchardt, J.; Böttcher, I.; Schaufler, K.; Guenther, S. Synergistic antimicrobial activities of epigallocatechin gallate, myricetin, daidzein, gallic acid, epicatechin, 3-hydroxy-6-methoxyflavone and genistein combined with antibiotics against ESKAPE pathogens. J. Appl. Microbiol. 2022, 132, 949–963. [Google Scholar] [CrossRef]
- Negm, W.; Abo El-Seoud, K.; Kabbash, A.; El-Aasr, M. Investigation of the Biological Activity Some Gymnosperm Plants Belong to Cycadales Order. J. Adv. Med. Pharm. Res. 2020, 1, 9–13. [Google Scholar] [CrossRef]
- Elmongy, E.I.; Negm, W.A.; Elekhnawy, E.; El-Masry, T.A.; Attallah, N.G.; Altwaijry, N.; Batiha, G.E.-S.; El-Sherbeni, S.A. Antidiarrheal and Antibacterial Activities of Monterey Cypress Phytochemicals: In Vivo and In Vitro Approach. Molecules 2022, 27, 346. [Google Scholar] [CrossRef]
- Patel, J. Synthesis, characterization and antimicrobial screening of some furofused coumarines. Int. J. Adv. Res. Dev. 2018, 3, 1–8. [Google Scholar]
- Widelski, J.; Luca, S.V.; Skiba, A.; Chinou, I.; Marcourt, L.; Wolfender, J.-L.; Skalicka-Wozniak, K. Isolation and antimicrobial activity of coumarin derivatives from fruits of Peucedanum luxurians Tamamsch. Molecules 2018, 23, 1222. [Google Scholar] [CrossRef] [Green Version]
- Öz, Y.; Özdemir, H.G.; Gökbolat, E.; Kiraz, N.; Ilkit, M.; Seyedmousavi, S. Time-Kill Kinetics and In Vitro Antifungal Susceptibility of Non-fumigatus Aspergillus Species Isolated from Patients with Ocular Mycoses. Mycopathologia 2016, 181, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Attallah, N.G.; Negm, W.A.; Elekhnawy, E.; Altwaijry, N.; Elmongy, E.I.; El-Masry, T.A.; Alturki, E.A.; Yousef, D.A.; Shoukheba, M.Y. Antibacterial Activity of Boswellia sacra Flueck. Oleoresin Extract against Porphyromonas gingivalis Periodontal Pathogen. Antibiotics 2021, 10, 859. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Kong, J.; Ju, J.; Zhang, Y.; Guo, Y.; Cheng, Y.; Qian, H.; Xie, Y.; Yao, W. Membrane damage mechanism contributes to inhibition of trans-cinnamaldehyde on Penicillium italicum using Surface-Enhanced Raman Spectroscopy (SERS). Sci. Rep. 2019, 9, 490. [Google Scholar] [CrossRef] [PubMed]
- Fischer, D.; Gessner, G.; Fill, T.P.; Barnett, R.; Tron, K.; Dornblut, K.; Kloss, F.; Stallforth, P.; Hube, B.; Heinemann, S.H. Disruption of membrane integrity by the bacterium-derived antifungal jagaricin. Antimicrob. Agents Chemother. 2019, 63, e00707-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elekhnawy, E.A.; Sonbol, F.I.; Elbanna, T.E.; Abdelaziz, A.A. Evaluation of the impact of adaptation of Klebsiella pneumoniae clinical isolates to benzalkonium chloride on biofilm formation. Egypt. J. Med. Hum. Genet. 2021, 22, 51. [Google Scholar] [CrossRef]
- Elekhnawy, E.; Sonbol, F.; Abdelaziz, A.; Elbanna, T. An investigation of the impact of triclosan adaptation on Proteus mirabilis clinical isolates from an Egyptian university hospital. Braz. J. Microbiol. 2021, 52, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Kralik, P.; Ricchi, M. A Basic Guide to Real Time PCR in Microbial Diagnostics: Definitions, P., and Everything. Front. Microbiol. 2017, 8, 108. [Google Scholar] [CrossRef] [Green Version]
- Attallah, N.G.; Elekhnawy, E.; Negm, W.A.; Hussein, I.A.; Mokhtar, F.A.; Al-Fakhrany, O.M. In Vivo and In Vitro Antimicrobial Activity of Biogenic Silver Nanoparticles against Staphylococcus aureus Clinical Isolates. Pharmaceuticals 2022, 15, 194. [Google Scholar] [CrossRef]
- Wang, W.-Y.; Tan, M.-S.; Yu, J.-T.; Tan, L. Role of pro-inflammatory cytokines released from microglia in Alzheimer’s disease. Ann. Transl. Med. 2015, 3, 136. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Error | Metabolite Name | Rt Min | Molecular Formula | Identification | Fragments or MS2 Ions m/z | Precursor m/z [M–H]− | Theoretical Mass | Ref. | |
|---|---|---|---|---|---|---|---|---|---|---|
| Phenolic or Organic Acids Derivatives | ||||||||||
| 1 | −4.7 | D-Malic acid | 1.098 | C4H6O5 | Beta hydroxy acids and derivatives | 59.01, 71.01, 115.00 133.01 | 133.0150 | 134.0230 | [22] | |
| 2 | −2.1 | Shikimic acid | 1.111 | C7H10O5 | Shikimic acids and derivatives | 73.02, 137.04, 173.04 | 173.0460 | 174.0540 | [23] | |
| 3 | −0.1 | Maleic acid | 1.112 | C4H4O4 | Dicarboxylic acids and derivatives | 71.01, 115.01 | 115.0032 | 116.0112 | ||
| 4 | −0.2 | D-Quinic acid | 1.136 | C7H12O6 | Quinic acids and derivatives | 85.02, 145.04, 173.04, 191.05 | 191.0562 | 192.0642 | ||
| 5 | −0.9 | 3,4-Dihydroxy benzoic acid | 1.257 | C7H6O4 | Hydroxybenzoic acid derivatives | 109.03, 153.01 | 153.0187 | 154.0267 | [24] | |
| 6 | 3.1 | Caffeic acid | 1.344 | C9H8O4 | Hydroxycinnamic acids | 59.02, 89.02, 135.04, 179.05 | 179.0545 | 180.0625 | ||
| 7 | 4.4 | 3-(4-Hydroxy-3-methoxy-phenyl) prop-2-enoic acid | 2.076 | C10H10O4 | Hydroxycinnamic acids | 134.03, 193.05 | 193.0524 | 194.0604 | ||
| 8 | 0.8 | Salicylic acid | 4.442 | C7H6O3 | Salicylic acid | 91.02, 108.01, 136.01, 137.02 | 137.0238 | 138.0318 | [25] | |
| 9 | 0.8 | (+/−)-cis,trans-abscisic acid | 5.286 | C15H20O4 | Abscisic acids and derivatives | 151.07, 219.13, 263.11 | 263.1278 | 264.1358 | [25] | |
| 10 | −0.7 | 5-Methoxy salicylic acid | 6.857 | C8H8O4 | M-Methoxybenzoic acids and derivatives | 108.02, 152.01, 167.03 | 167.0352 | 168.0432 | ||
| 11 | 6.0 | Rosmarinic acid | 7.136 | C18H16O8 | Coumaric acids and derivatives | 181.04, 329.09, 359.11 | 359.1105 | 360.1185 | [26] | |
| 12 | −3.1 | Isocitrate | 27.721 | C6H8O7 | Tricarboxylic acids and derivatives | 150.96, 190.95 | 191.1050 | 192.1130 | ||
| Flavonoids and related metabolites | ||||||||||
| 13 | −3.7 | Baicalein-7-O-glucuronide | 1.293 | C21H18O11 | 445.07 | Flavonoid-7-O-glucuronides | 102.95, 191.05, 269.02 377.07, 445.14 | 445.1807 | 446.1887 | [27] |
| 14 | −2.1 | Isorhamnetin-3-O-rutinoside | 4.930 | C28H32O16 | 623.12 | Flavonoid-3-O-glycosides | 285.03, 470.87, 579.00, 623.12 | 623.1269 | 624.1349 | [28] |
| 15 | −3.1 | Procyanidin B2 | 5.006 | C30H26O12 | 577.13 | Biflavonoids and polyflavonoids | 407.07, 425.08, 577.13 | 577.1369 | 578.1449 | [28] |
| 16 | 5.0 | Kaempferol-3-O-alpha-L-rhamnoside | 5.223 | C21H20O10 | 431.18 | Flavonoid-3-O-glycosides | 151.04, 179.05, 362.88, 431.19 | 431.1890 | 432.1970 | |
| 17 | −3.6 | Daidzein-8-C-glucoside | 5.452 | C21H20O9 | 415.15 | Isoflavonoid C-glycosides | 210.92, 286.92, 415.16 | 415.1572 | 416.1652 | [29, 30] |
| 18 | −0.1 | Naringenin-7-O-glucoside | 6.170 | C21H22O10 | 433.20 | Flavonoid-7-O-glycosides | 271.06, 387.18, 433.10 | 433.2068 | 434.2148 | |
| 19 | −2.2 | (+−)-Taxifolin | 6.492 | C15H12O7 | 303.05 | Flavanonols | 125.02, 179.00, 1990.95, 285.04, 303.04 | 303.0520 | 304.0600 | |
| 20 | 2.5 | Hyperoside | 6.542 | C21H20O12 | 463.08 | Flavonoid-3-O-glycosides | 218.94, 286.94. 300.02, 326.92, 394.90, 463.08 | 463.0858 | 464.0938 | |
| 21 | 0.5 | Apigenin 8-C-glucoside | 6.542 | C21H20O10 | 431.09 | Flavonoid 8-C-glycosides | 269.04,311.05, 362.89, 431.20 | 431.0956 | 432.1036 | |
| 22 | −0.1 | Formononetin | 6.734 | C16H12O4 | 267.08 | 4’-O-methylisoflavones | 133.02, 193.04, 252.06, 267.08 | 267.0875 | 268.0955 | |
| 23 | 0.9 | Procyanidin B1 | 6.785 | C30H26O12 | 577.14 | Biflavonoids and polyflavonoids | 245.04, 425.08, 577.12 | 577.1337 | 578.1417 | [31] |
| 24 | 0.6 | Luteolin-7-O-glucoside | 6.913 | C21H20O11 | 447.09 | Flavonoid-7-O-glycosides | 285.03, 327.04, 447.09 | 447.0923 | 448.1003 | |
| 25 | 8.9 | Isookanin-7-glucoside | 6.926 | C21H22O11 | 449.09 | Flavonoid-7-O-glycosides | 151.00, 287.04, 449.09 | 449.0952 | 450.1032 | |
| 26 | 0.4 | Isorhamnetin-3-O-glucoside | 7.417 | C22H22O12 | 477.10 | Flavonoid-3-O-glycosides | 285.03, 299.01, 315.04, 477.09 | 477.1033 | 478.1113 | |
| 27 | −0.1 | Kaempferol-7-neohesperidoside | 7.539 | C27H30O15 | 593.14 | Flavonoid-7-O-glycosides | 269.04, 593.13, | 593.1483 | 594.1563 | |
| 28 | −0.9 | Apigenin-7-O-glucoside | 7.700 | C21H20O10 | 431.09 | Flavonoid-7-O-glycosides | 268.03, 311.06, 431.09 | 431.0988 | 432.1068 | |
| 29 | 10.9 | Kaempferol-3-Glucuronide | 7.893 | C21H18O12 | 461.10 | Flavonoid-3-O-glucuronides | 256.92, 299.09, 392.89, 461.09 | 461.1031 | 462.1111 | |
| 30 | −3.7 | 3’ 4’ 5 7-Tetrahydroxy flavanone | 8.678 | C13H12O6 | 287.05 | Flavanones | 135.04, 151.00, 218.94, 287.06 | 287.0984 | 288.1064 | |
| 31 | −0.2 | Kaempferol-3-O-alpha-L-arabinoside | 8.751 | C20H18O10 | 417.11 | Flavonoid-3-O-glucuronides | 179.05, 362.88, 431.19 | 417.1194 | 418.1274 | |
| 32 | −0.1 | Naringenin | 9.744 | C15H12O5 | 271.09 | Flavanones | 228.07, 255.05, 271.09 | 271.0985 | 272.1065 | [32] |
| 33 | −2.6 | 3’-Methoxy-4’,5,7-trihydroxy flavonol | 10.368 | C16H12O7 | 315.05 | Flavonols | 257.04, 300.02, 315.04 | 315.0528 | 316.0608 | |
| 34 | −3.0 | Apigenin | 10.428 | C15H10O5 | 269.04 | Flavones | 117.03, 151.00, 181.06, 225.05, 269.04 | 269.0467 | 270.0547 | [32] |
| 35 | 0.8 | Luteolin | 11.175 | C15H10O6 | 285.07 | Flavones | 151.00, 270.06, 263.11 | 285.0763 | 286.0843 | [32] |
| 36 | 0.7 | Rhoifolin | 11.421 | C27H30O14 | 577.13 | Flavonoid-7-O-glycosides | 291.19, 464.94, 577.13 | 577.1346 | 578.1426 | |
| 37 | −4.0 | 3 5 7-Trihydroxy-4’-methoxyflavone | 12.886 | C16H12O6 | 299.09 | Flavonols | 119.04, 179.03, 193.05, 299.09 | 299.0558 | 300.0638 | |
| 38 | −0.1 | 4’,5,7-Trihydroxy flavonol | 13.890 | C15H10O6 | 285.11 | Flavonols | 119.04, 149.99, 165.02, 285.07 | 285.1115 | 286.1195 | |
| 39 | −0.4 | Hesperetin | 14.598 | C16H14O6 | 301.10 | 4’-O-methylated flavonoids | 138.03, 151.03, 286.08, 301.10 | 301.0692 | 302.0772 | [33] |
| 40 | −0.2 | Luteolin-3’, 7-di-O-glucoside | 17.799 | C27H30O16 | 609.13 | Flavonoid-7-O-glycosides | 564.90, 594.11, 609.13 | 609.1440 | 610.1520 | |
| Fatty acids | ||||||||||
| 41 | −0.8 | 3-Hydroxy-3-Methyl glutaric acid | 1.159 | C6H10O5 | 161.04 | Hydroxy fatty acids | 57.03, 99.04, 161.04 | 161.0452 | 162.0532 | |
| 42 | −2.7 | 2-Isopropyl malic acid | 1.331 | C7H12O5 | 175.05 | Hydroxy fatty acids | 101.02, 113.05, 175.06 | 175.0599 | 176.0679 | |
| 43 | −0.5 | Citraconic acid | 1.394 | C5H6O4 | 129.01 | Methyl-branched fatty acids | 84.99, 129.01 | 129.0189 | 130.0269 | |
| 44 | 1.5 | Citramalate | 1.407 | C5H8O5 | 147.06 | Hydroxy fatty acids | 72.98, 87.00, 129.01, 147.06 | 147.0647 | 148.0727 | |
| 45 | −4.0 | Gamma-Linolenic acid | 18.900 | C18H30O2 | 277.14 | Linoleic acids and derivatives | 141.09, 233.15, 277.13 | 277.1465 | 278.1545 | |
| Others | ||||||||||
| 46 | −0.3 | E-3,4,5’-Trihydroxy-3’-glucopyranosyl stilbene | 1.123 | C20H22O9 | 405.01 | Stilbene glycosides | 190.96, 191.05, 243.02, 369.03, 405.09 | 405.1208 | 406.1288 | |
| 47 | −0.6 | Esculin | 1.172 | C15H16O9 | 339.12 | Coumarin glycosides | 149.01, 175.02, 202.90, 295.10, 339.11 | 339.1296 | 340.1376 | |
| 48 | 0.2 | Catechin | 5.452 | C15H14O6 | 289.07 | Catechins | 109.02, 125.02, 179.03, 203.03, 245.07, 289.06 | 289.0711 | 290.0791 | [31] |
| 49 | 3.0 | Hinokitiol | 6.815 | C10H12O2 | 163.07 | Tropolones | 119.08, 135.08, 163.07 | 163.0751 | 164.0831 | |
| 50 | −0.4 | Daphnetin | 7.295 | C9H6O4 | 177.01 | 7,8-Dihydroxycoumarins | 117.09, 133.03, 149.02, 163.01, 177.01 | 177.018 | 178.0260 | |
| 51 | 1.0 | E-4,5’-Dihydroxy-3-methoxy-3’-glucopyranosyl stilbene | 9.547 | C21H24O9 | 419.07 | Stilbene glycosides | 257.02, 343.01, 363.12, 419.06 | 419.0748 | 420.0828 | |
| Efflux Pump Activity | No. of Isolates | |
|---|---|---|
| Before Treatment | After Treatment | |
| Strong | 3 | 1 |
| Moderate | 3 | 1 |
| Weak | 4 | 6 |
| None | 2 | 4 |
| Measured Parameter | Group I | Group II | Group III |
|---|---|---|---|
| Blood urea level (mg/dL) | 69 ± 1.40 * | 47.6 ± 1.10 | 48 ± 0.90 |
| Serum creatinine level (mg/dL) | 0.95 ± 0.06 * | 0.7 ± 0.02 | 0.69 ± 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqahtani, M.J.; Elekhnawy, E.; Negm, W.A.; Mahgoub, S.; Hussein, I.A. Encephalartos villosus Lem. Displays a Strong In Vivo and In Vitro Antifungal Potential against Candida glabrata Clinical Isolates. J. Fungi 2022, 8, 521. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050521
Alqahtani MJ, Elekhnawy E, Negm WA, Mahgoub S, Hussein IA. Encephalartos villosus Lem. Displays a Strong In Vivo and In Vitro Antifungal Potential against Candida glabrata Clinical Isolates. Journal of Fungi. 2022; 8(5):521. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050521
Chicago/Turabian StyleAlqahtani, Moneerah J., Engy Elekhnawy, Walaa A. Negm, Sebaey Mahgoub, and Ismail A. Hussein. 2022. "Encephalartos villosus Lem. Displays a Strong In Vivo and In Vitro Antifungal Potential against Candida glabrata Clinical Isolates" Journal of Fungi 8, no. 5: 521. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050521