
Fungal Community Composition as Affected by Litter Chemistry and Weather during Four Years of Litter Decomposition in Rainshadow Coastal Douglas-Fir Forests

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Litter Bags and Field Sites
2.2. Sample Collection and Processing
2.3. Data Analysis
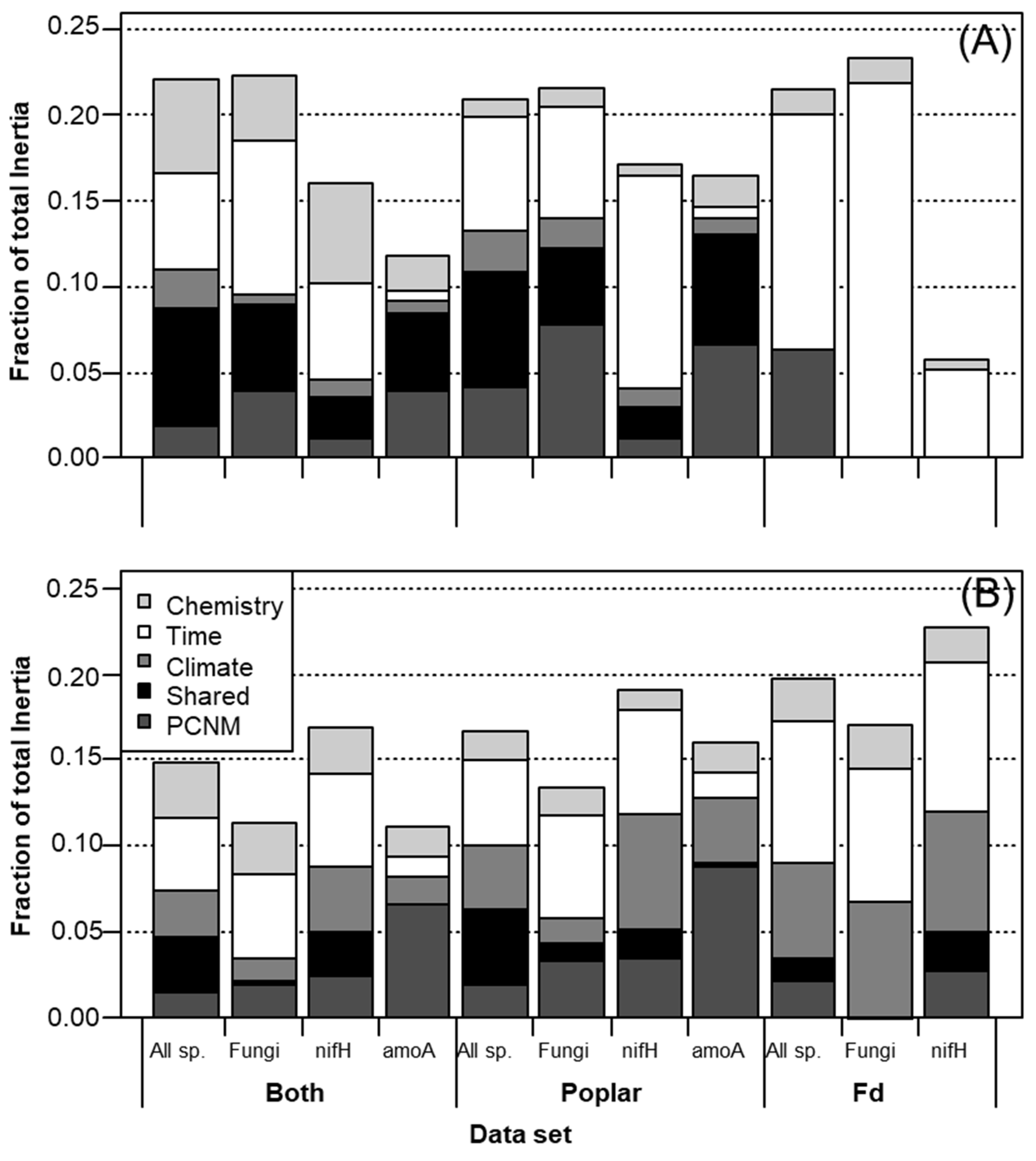
3. Results
3.1. DNA Quality
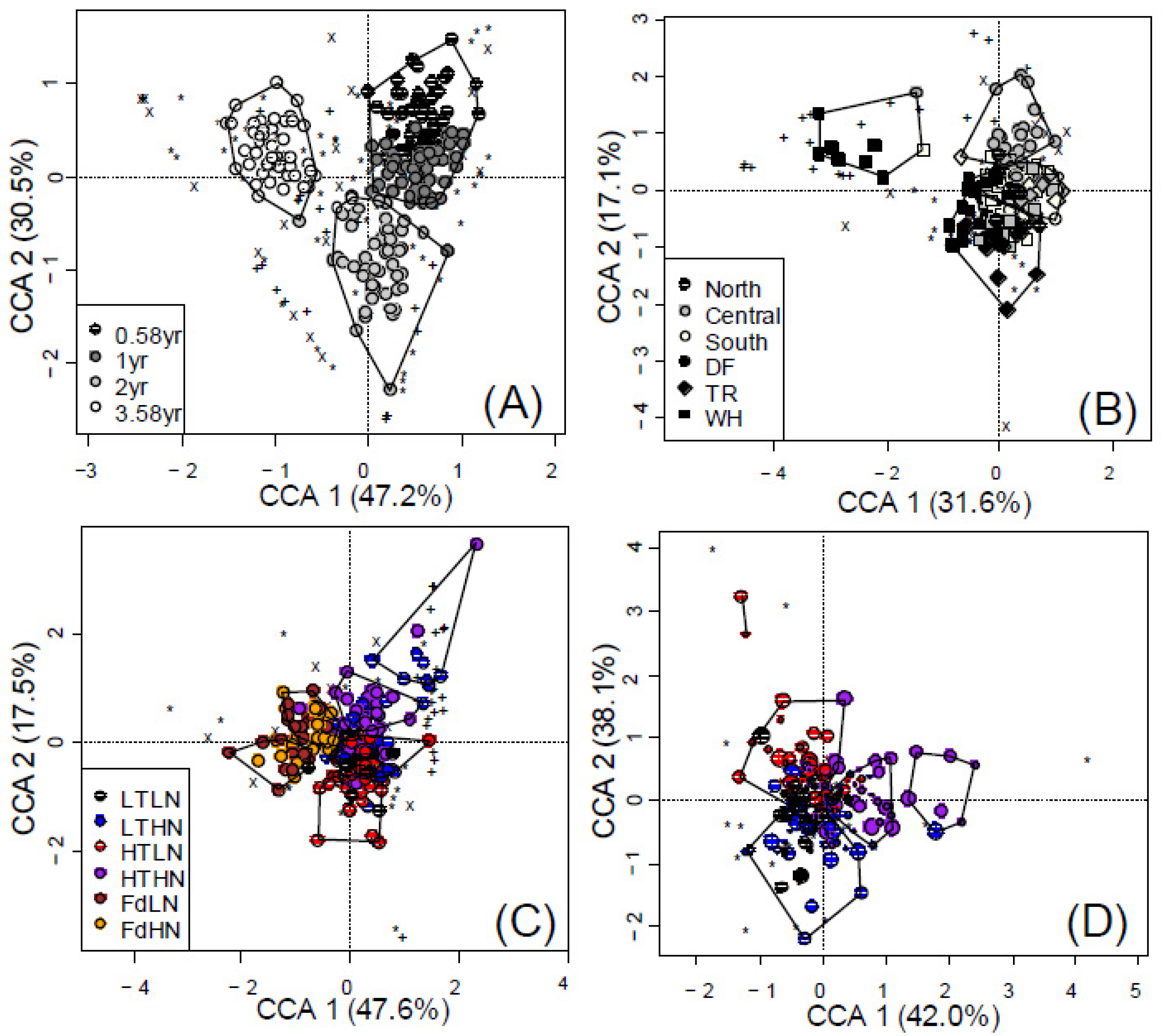
3.2. Temporal Effects
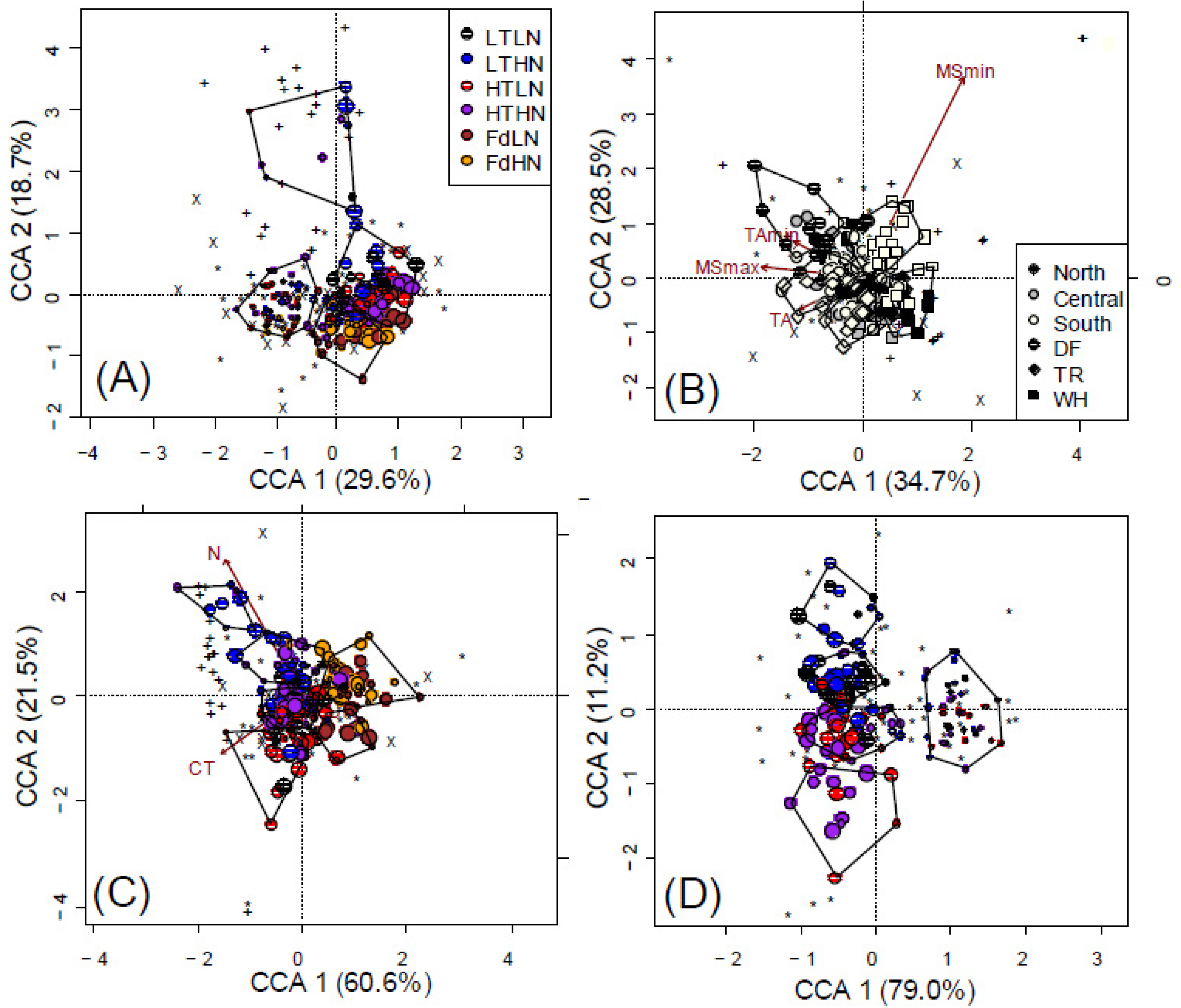
3.3. Litter Chemistry Effects
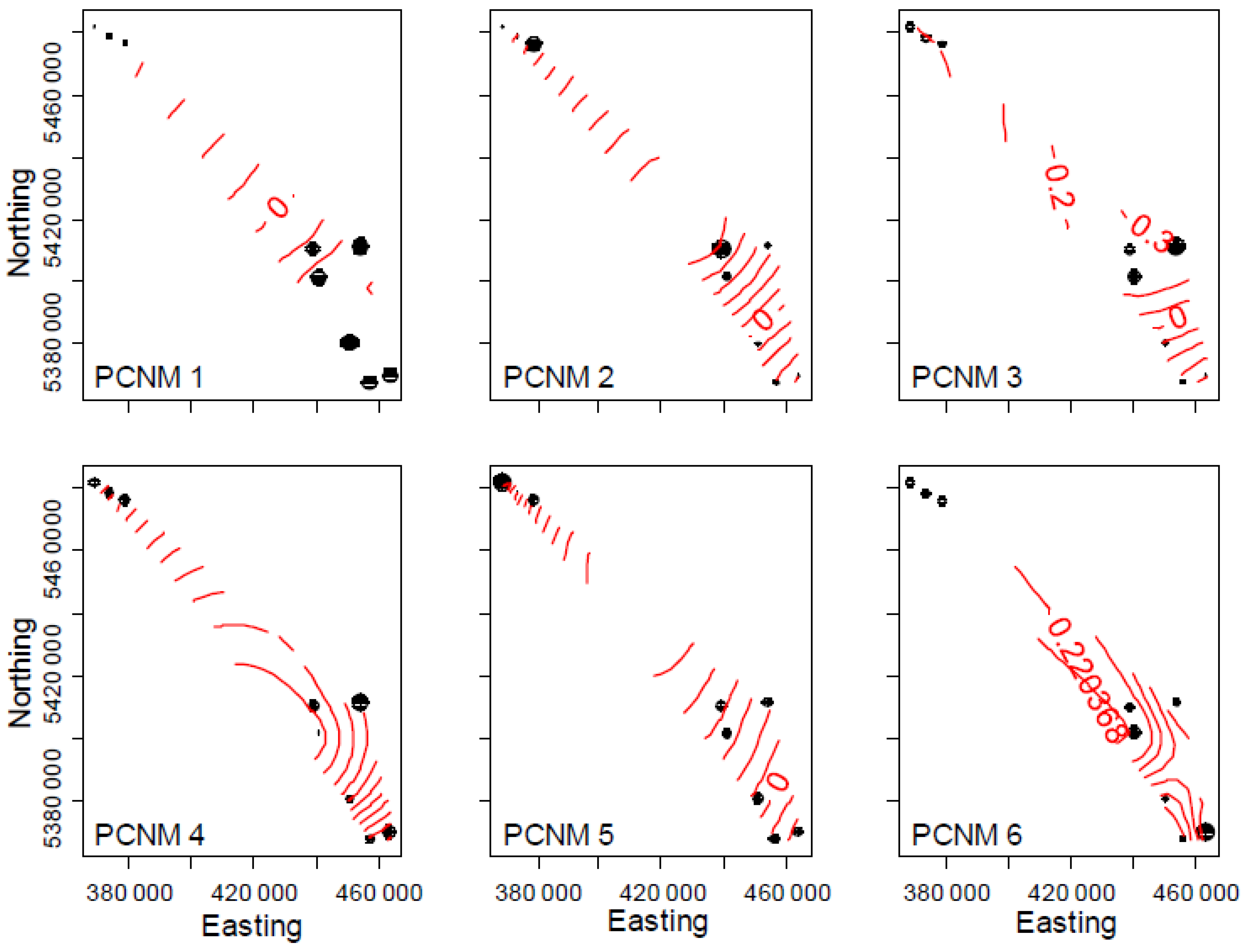
3.4. Spatial and Climatic Effects
4. Discussion
4.1. Ammonifying Communities Are Altered by Weather and Litter Nitrogen
4.2. Weather Can Account for Early Succession in Fungal Composition
4.3. Short-Term Effects on Fungal Communities Driven by Litter Chemistry
4.4. Diazotrophs Differ among Litter Types
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schlesinger, W.H.; Andrews, J.A. Soil respiration and the global carbon cycle. Biogeochemistry 2000, 48, 7–20. [Google Scholar] [CrossRef]
- Palosuo, T.; Liski, J.; Trofymow, J.A.; Titus, B.D. Litter decomposition affected by climate and litter quality—Testing the Yasso model with litterbag data from the Canadian intersite decomposition experiment. Ecol. Mod. 2005, 189, 183–198. [Google Scholar] [CrossRef]
- Moore, T.R.; Trofymow, J.A.; Prescott, C.E.; Fyles, J.; Titus, B.D. Patterns of carbon, nitrogen and phosphorus dynamics in decomposing foliar litter in Canadian forests. Ecosystems 2006, 9, 46–62. [Google Scholar] [CrossRef]
- Prescott, C.E. Litter decomposition: What controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 2010, 101, 133–149. [Google Scholar] [CrossRef]
- Solomon, S.; Hegerl, G.; Heimann, M.; Hewitson, B.; Hoskins, B.; Joos, F.; Jouzel, J.; Kattsov, V.; Lohmann, U.; Matsuno, T.; et al. Contribution of Working Group 1, Fourth Assessment Report of the IPCC; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Raich, J.W.; Potter, C.S. Global patterns of carbon-dioxide emissions from soils. Glob. Biogeochem. Cycles 1995, 9, 23–36. [Google Scholar] [CrossRef] [Green Version]
- Van Gestel, M.; Merckx, R.; Vlassak, K. Microbial biomass responses to soil drying and rewetting—The fate of fast-growing and slow-growing microorganisms in soils from different climates. Soil Biol. Biochem. 1993, 25, 109–123. [Google Scholar] [CrossRef]
- de Vries, F.T.; Shade, A. Controls on soil microbial community stability under climate change. Front. Microbiol. 2013, 4, 265. [Google Scholar] [CrossRef] [Green Version]
- Kurz, W.A.; Dymond, C.C.; White, T.M.; Stinson, G.; Shaw, C.H.; Rampley, G.J.; Smyth, C.; Simpson, B.N.; Neilson, E.T.; Trofymow, J.A.; et al. CBM-CFS3: A model of carbon-dynamics in forestry and land-use change implementing IPCC standards. Ecol. Mod. 2009, 220, 480–504. [Google Scholar] [CrossRef]
- Tarnocai, C.; Canadell, J.G.; Schuur, E.A.G.; Kuhry, P.; Mazhitova, G.; Zimov, S. Soil organic carbon pools in the northern circumpolar permafrost region. Glob. Biogeochem. Cycles 2009, 23, 1–11. [Google Scholar] [CrossRef]
- Mansfield, J.L.; Curtis, P.S.; Zak, D.R.; Pregitzer, K.S. Genotypic variation for condensed tannin production in trembling aspen (Populus tremuloides, Salicaceae) under elevated CO2 and in high- and low-fertility soil. Amer. J. Bot. 1999, 86, 1154–1159. [Google Scholar] [CrossRef] [Green Version]
- Moore, T.R.; Trofymow, J.A.; Taylor, B.; Prescott, C.; Camire, C.; Duschene, L.; Fyles, J.; Kozak, L.; Kranabetter, M.; Morrison, I.; et al. Rates of litter decomposition in Canadian forests. Glob. Chang. Biol. 1999, 5, 75–82. [Google Scholar] [CrossRef]
- Norby, R.J.; Cotrufo, M.F.; Ineson, P.; O’Neill, E.G.; Canadell, J.G. Elevated CO2, litter chemistry, and decomposition: A synthesis. Oecologia 2001, 127, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Parsons, W.F.J.; Bockheim, J.G.; Lindroth, R.L. Independent, interactive, and species-specific responses of leaf litter decomposition to elevated CO2 and O3 in a northern hardwood forest. Ecosystems 2008, 11, 505–519. [Google Scholar] [CrossRef]
- Driebe, E.M.; Whitham, T.G. Cottonwood hybridization affects tannin and nitrogen content of leaf litter and alters decomposition. Oecologia 2000, 123, 99–107. [Google Scholar] [CrossRef]
- Hattenschwiler, S.; Vitousek, P.M. The role of polyphenols in terrestrial ecosystem nutrient cycling. Trends Ecol. Evol. 2000, 15, 238–243. [Google Scholar] [CrossRef]
- Hattenschwiler, S.; Hagerman, A.E.; Vitousek, P.M. Polyphenols in litter from tropical montane forests across a wide range in soil fertility. Biogeochemistry 2003, 64, 129–148. [Google Scholar] [CrossRef]
- Smyth, C.E.; Trofymow, J.A.; Kurz, W.A.; CIDET Working Group. Decreasing Uncertainty in CBM-CFS3 Estimates of Forest Soil C Sources and Sinks through Use of Long-Term Data from the Canadian Intersite Decomposition Experiment; Inf. Rep. BC-X-422; Natural Resources Canada, Canadian Forest Service: Victoria, BC, Canada, 2010. [Google Scholar]
- Shay, P.E.; Constabel, C.P.; Trofymow, J.A. Evidence for the role and fate of water-insoluble condensed tannins in the short-term reduction of carbon loss during litter decay. Biogeochemistry 2018, 137, 127–141. [Google Scholar] [CrossRef] [Green Version]
- Kraus, T.E.C.; Dahlgren, R.A.; Zasoski, R.J. Tannins in nutrient dynamics of forest ecosystems—A review. Plant Soil 2003, 256, 41–66. [Google Scholar] [CrossRef]
- Constabel, C.P. Molecular controls of proanthocyanidin synthesis and structure: Prospects for genetic engineering in crop plants. J. Agric. Food Chem. 2018, 66, 9882–9888. [Google Scholar] [CrossRef]
- Barbehenn, R.V.; Constabel, C.P. Tannins in plant-herbivore interactions. Phytochemistry 2011, 72, 1551–1565. [Google Scholar] [CrossRef]
- Hagerman, A.E.; Riedl, K.M.; Jones, G.A.; Sovik, K.N.; Ritchard, N.T.; Hartzfeld, P.W.; Riechel, T.L. High molecular weight plant polyphenolics (tannins) as biological antioxidants. J. Agric. Food Chem. 1998, 46, 1887–1892. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A. Antimicrobial properties of tannins. Phytochemistry 1991, 30, 3875–3883. [Google Scholar] [CrossRef]
- Tarascou, I.; Souquet, J.M.; Mazauric, J.P.; Carrillo, S.; Coq, S.; Canon, F.; Fulcrand, H.; Cheynier, V. The hidden face of food phenolic composition. Arch. Biochem. Biophys. 2010, 501, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.H.; Lin, Y.M.; Ye, G.F.; Liu, X.W.; Lin, G.H. Changes in the N and P concentrations, N: P ratios, and tannin content in Casuarina equisetifolia branchlets during development and senescence. J. For. Res. 2008, 13, 302–311. [Google Scholar] [CrossRef] [Green Version]
- Joanisse, G.D.; Bradley, R.L.; Preston, C.M.; Munson, A.D. Soil enzyme inhibition by condensed litter tannins may drive ecosystem structure and processes: The case of Kalmia angustifolia. New Phytol. 2007, 175, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Strickland, M.S.; Rousk, J. Considering fungal:bacterial dominance in soils—Methods, controls, and ecosystem implications. Soil Biol. Biochem. 2010, 42, 1385–1395. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Carreiro, M.M.; Repert, D.A. Allocation of extracellular enzymatic activity in relation to litter composition, N deposition, and mass loss. Biogeochemistry 2002, 60, 1–24. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Polme, S.; Koljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1078. [Google Scholar] [CrossRef] [Green Version]
- Peay, K.G.; Schubert, M.G.; Nguyen, N.H.; Bruns, T.D. Measuring ectomycorrhizal fungal dispersal: Macroecological patterns driven by microscopic propagules. Mol. Ecol. 2012, 21, 4122–4136. [Google Scholar] [CrossRef]
- Peay, K.G.; Bruns, T.D. Spore dispersal of basidiomycete fungi at the landscape scale is driven by stochastic and deterministic processes and generates variability in plant-fungal interactions. New Phytol. 2014, 204, 180–191. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Howarth, R.W. Nitrogen limitation on land and in the sea—How can it occur. Biogeochemistry 1991, 13, 87–115. [Google Scholar] [CrossRef]
- Manzoni, S.; Trofymow, J.A.; Jackson, R.B.; Porporato, A. Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter. Ecol. Monogr. 2010, 80, 89–106. [Google Scholar] [CrossRef]
- Glick, B.R.; Patten, C.L.; Holguin, G.; Penrose, D.M. Biochemical and Genetic Mechanisms used by Plant Growth Promoting Bacteria; Imperial College Press: London, UK, 1991. [Google Scholar] [CrossRef]
- Barnard, R.; Leadley, P.W.; Hungate, B.A. Global change, nitrification, and denitrification: A review. Glob. Biogeochem. Cycles 2005, 19, 13. [Google Scholar] [CrossRef]
- Winder, R.S.; Lamarche, J.; Constabel, C.P.; Hamelin, R.C. The effects of high-tannin leaf litter from transgenic poplars on microbial communities in microcosm soils. Front. Microbiol. 2013, 4, 290. [Google Scholar] [CrossRef] [Green Version]
- Levy-Booth, D.J.; Winder, R.S. Quantification of nitrogen reductase and nitrate reductase gene abundance in soil of thinned and clear-cut Douglas-fir stands using real-time PCR. Appl. Environ. Microbiol. 2010, 76, 7116–7125. [Google Scholar] [CrossRef] [Green Version]
- Saetre, P.; Baath, E. Spatial variation and patterns of soil microbial community structure in a mixed spruce-birch stand. Soil Biol. Biochem. 2000, 32, 909–917. [Google Scholar] [CrossRef]
- Shay, P.E.; Winder, R.S.; Trofymow, J.A. Nutrient-cycling microbes in coastal Douglas-fir forests: Regional-scale correlation between communities, in situ climate, and other factors. Front. Microbiol. 2015, 6, 1097. [Google Scholar] [CrossRef] [Green Version]
- Voriskova, J.; Baldrian, P. Fungal community on decomposing leaf litter undergoes rapid successional changes. Int. Soc. Microbiol. Ecol. J. 2013, 7, 477–486. [Google Scholar]
- Ramette, A.; Tiedje, J.M. Multiscale responses of microbial life to spatial distance and environmental heterogeneity in a patchy ecosystem. Proc. Nat. Acad. Sci. USA 2007, 104, 2761–2766. [Google Scholar] [CrossRef] [Green Version]
- Fromin, N.; Hamelin, J.; Tarnawski, S.; Roesti, D.; Jourdain-Miserez, K.; Forestier, N.; Teyssier-Cuvelle, S.; Gillet, F.; Aragno, M.; Rossi, P. Statistical analysis of denaturing gel electrophoresis (DGE) fingerprinting patterns. Environ. Microbiol. 2002, 204, 634–643. [Google Scholar]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Nat. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monroy, F.; van der Putten, W.H.; Yergeau, E.; Mortimer, S.R.; Duyts, H.; Bezemer, T.M. Community patterns of soil bacteria and nematodes in relation to geographic distance. Soil Biol. Biochem. 2012, 45, 1–7. [Google Scholar] [CrossRef]
- Trofymow, J.A.; CIDET Working Group. The Canadian Intersite Decomposition Experiment (CIDET): Project and Site Establishment Report; Inf. Rep. BC-X-378; Natural Resources Canada, Canadian Forest Service: Victoria, BC, Canada, 1998; 126p. [Google Scholar]
- Lamit, L.J.; Bowker, M.A.; Holeski, L.M.; Naesborg, R.R.; Wooley, S.C.; Zinkgraf, M.; Lindroth, R.L.; Whithama, T.G.; Gehring, C.A. Genetically-based trait variation within a foundation tree species influences a dominant bark lichen. Fungal Ecol. 2011, 4, 103–109. [Google Scholar] [CrossRef]
- Rehill, B.J.; Whitham, T.G.; Martinsen, G.D.; Schweitzer, J.A.; Bailey, J.K.; Lindroth, R.L. Developmental trajectories in cottonwood phytochemistry. J. Chem. Ecol. 2006, 32, 2269–2285. [Google Scholar] [CrossRef] [Green Version]
- Shay, P.E.; Trofymow, J.A.; Constabel, C.P. An improved butanol-HCl assay for quantification of water-soluble, acetone:methanol-soluble, and insoluble proanthocyanidins (condensed tannins). Plant Methods 2017, 13, 63. [Google Scholar] [CrossRef] [Green Version]
- Pojar, J.; Meidinger, D. British Columbia: The Environmental Settings. In Ecosystems of British Columbia; Meidinger, D.V., Pojar, J., Eds.; Special Report Series 6; BC Ministry of Forests: Victoria, BC, Canada, 2011; pp. 40–66. [Google Scholar]
- Sheffield, V.C.; Cox, D.R.; Lerman, L.S.; Myers, R.M. Attachment of a 40-base pair G +C-rich sequence (GC-clamp) to genomic DNA fragments by the polymerase chain reaction results in improved detection of single-base changes. Proc. Nat. Acad. Sci. USA 1987, 86, 232–236. [Google Scholar] [CrossRef] [Green Version]
- Kowalchuk, G.A.; Stephen, J.R.; DeBoer, W.; Prosser, J.I.; Embley, T.M.; Woldendorp, J.W. Analysis of ammonia-oxidizing bacteria of the beta subdivision of the class Proteobacteria in coastal sand dunes by denaturing gradient gel electrophoresis and sequencing of PCR-amplified 16S ribosomal DNA fragments. Appl. Environ. Microbiol. 1997, 63, 1489–1497. [Google Scholar] [CrossRef] [Green Version]
- Vainio, E.J.; Hantula, J. Direct analysis of wood-inhabiting fungi using denaturing gradient gel electrophoresis of amplified ribosomal DNA. Mycol. Res. 2000, 104, 927–936. [Google Scholar] [CrossRef]
- Widmer, F.; Shaffer, B.T.; Porteous, L.A.; Seidler, R.J. Analysis of nifH gene pool complexity in soil and litter at a Douglas fir forest site in the Oregon Cascade Mountain Range. Appl. Environ. Microbiol. 1999, 65, 374–380. [Google Scholar] [CrossRef] [Green Version]
- Burgmann, H.; Meier, S.; Bunge, M.; Widmer, F.; Zeyer, J. Effects of model root exudates on structure and activity of a soil diazotroph community. Environ. Microbiol. 2005, 7, 1711–1724. [Google Scholar] [CrossRef]
- Nicolaisen, M.H.; Ramsing, N.B. Denaturing gradient gel electrophoresis (DGGE) approaches to study the diversity of ammonia-oxidizing bacteria. J. Microbiol. Methods 2002, 50, 189–203. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; Borcard, D.; Peres-Neto, P.R. Analyzing beta diversity: Partitioning the spatial variation of community composition data. Ecol. Monogr. 2005, 75, 435–450. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package; R Package Version 2.3-3.; 2016. Available online: http://CRAN.R-project.org/package=vegan (accessed on 15 December 2016).
- Borcard, D.; Legendre, P.; Avois-Jacquet, C.; Tuomisto, H. Dissecting the spatial structure of ecological data at multiple scales. Ecology 2004, 85, 1826–1832. [Google Scholar] [CrossRef] [Green Version]
- Cerdeira, J.O.; Silva, P.D.; Cadima, J.; Minhoto, M. Subselect: Selecting Variable Subsets; R Package Version 0.12-5; 2015. Available online: http://CRAN.R-project.org/package=subselect (accessed on 1 November 2015).
- Josse, J.; Husson, F. Handling missing values in exploratory multivariate data analysis methods. J. Soc. Fr. Stat. 2013, 153, 79–99. [Google Scholar]
- Gijsman, A.J. Soil-water content as a key factor determining the source of nitrogen (NH4+ or NO3−) absorbed by Douglas fir (Pseudotsuga menziesii) and the pattern of rhizosphere pH along its roots. Can. J. For. Res. 1991, 21, 616–625. [Google Scholar] [CrossRef]
- Turner, D.P.; Sollins, P.; Leuking, M.; Rudd, N. Availability and uptake of inorganic nitrogen in a mixed old-growth coniferous forest. Plant Soil 1993, 148, 163–174. [Google Scholar] [CrossRef]
- Hamann, A.; Wang, T.L. Potential effects of climate change on ecosystem and tree species distribution in British Columbia. Ecology 2006, 87, 2773–2786. [Google Scholar] [CrossRef] [Green Version]
- Schimel, J.P.; Balser, T.C.; Wallenstein, M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 2007, 88, 1386–1394. [Google Scholar] [CrossRef]
- Lin, W.-R.; Wang, P.-H.; Chen, W.-C.; Lai, C.-M.; Winder, R.S. Responses of soil fungal populations and communities to the thinning of Cryptomeria japonica forests. Microbes Environ. 2016, 31, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Joanisse, G.D.; Bradley, R.L.; Preston, C.M.; Bending, G.D. Sequestration of soil nitrogen as tannin-protein complexes may improve the competitive ability of sheep laurel (Kalmia angustifolia) relative to black spruce (Picea mariana). New Phytol. 2009, 181, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Averill, C.; Turner, B.L.; Finzi, A.C. Mycorrhiza-mediated competition between plants and decomposers drives soil carbon storage. Nature 2014, 505, 543. [Google Scholar] [CrossRef]
- Legay, N.; Baxendale, C.; Grigulis, K.; Krainer, U.; Kastl, E.; Schloter, M.; Bardgett, R.D.; Arnoldi, C.; Bahn, M.; Dumont, M.; et al. Contribution of above- and below-ground plant traits to the structure and function of grassland soil microbial communities. Ann. Bot. 2014, 114, 1011–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, E.B.; Knelman, J.E.; Schindlbacher, A.; Siciliano, S.; Breulmann, M.; Yannarell, A.; Beman, J.M.; Abell, G.; Philippot, L.; Prosser, J.; et al. Microbes as engines of ecosystem function: When does community structure enhance predictions of ecosystem processes? Front. Microbiol. 2016, 7, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Litter Type | C (mg g−1) | N (mg g−1) | C/N | Phenols (mg g−1) | CT (mg g−1) |
|---|---|---|---|---|---|---|
| Populus angustifolia | LT LN | 457.0 ± 1.1 | 8.1 ± 0.2 | 56.39 ±1.45 | 18.3 ± 3.4 | 66.4 ± 8.1 |
| LT HN | 461.4 ± 0.7 | 15.7 ± 0.3 | 29.42 ±0.55 | 19.2 ± 4.4 | 85.8 ± 6.4 | |
| HT LN | 481.8 ± 1.2 | 6.9 ± 0.2 | 70.36 ±1.72 | 60.8 ± 6.8 | 264.9 ± 7.9 | |
| HT HN | 477.5 ± 1.3 | 14.3 ± 0.1 | 33.32 ±0.40 | 54.3 ± 4.8 | 291.5 ± 5.3 | |
| Pseudotsuga menziesii | Fd LN | 512.6 ± 1.5 | 6.1 ± 0.1 | 83.52 ±1.37 | 19.0 ± 3.1 | 20.8 ± 0.4 |
| Fd HN | 511.0 ± 0.5 | 11.2 ± 0.2 | 45.46 ±0.76 | 18.4 ± 1.5 | 20.9 ±0.3 | |
| Species | Litter Type | NPE a (mg g−1) | WSE b (mg g−1) | AHF c (mg g−1) | AUR d (mg g−1) | Ash (mg g−1) |
| Populus angustifolia | LT LN | 137.6 ± 15.2 | 130.7 ± 0.1 | 304.2 ± 5.0 | 414.8 ± 6.6 | 12.7 ± 0.6 |
| LT HN | 152.4 ± 5.6 | 128.4 ± 7.5 | 307.8 ± 4.4 | 398.1 ± 2.7 | 13.3 ± 1.6 | |
| HT LN | 177.7 ± 15.2 | 154.0 ± 11.1 | 283.8 ± 10.9 | 373.1 ± 5.4 | 11.4 ± 0.8 | |
| HT HN | 230.2 ± 6.5 | 132.3 ± 12.4 | 274.3 ± 3.0 | 353.0 ± 5.5 | 10.1 ± 0.5 | |
| Pseudotsuga menziesii | Fd LN | 99.5 ± 3.5 | 119.5 ± 1.0 | 362.6 ± 0.6 | 394.0 ± 3.4 | 24.4 ± 0.6 |
| Fd HN | 121.6 ± 3.2 | 117.2 ± 1.5 | 355.1 ± 1.2 | 382.6 ± 0.5 | 23.4 ± 0.4 |
| Latitude | BEC Zone | Soil Moisture (%) a | Soil Temp. (°C) a | Air Temp. (°C) b | PET c |
|---|---|---|---|---|---|
| North | DF | 13.93 [3.85, 40.97] | 9.55 [3.73, 15.37] | 8.99 [−4.86, 23.90] | 88.2 |
| TR | 12.97 [2.77, 45.56] | 9.64 [2.84, 15.85] | 9.60 [−4.30, 27.18] | 92.5 | |
| WH | 9.05 [3.03, 20.62] | 8.97 [2.96, 14.71] | 8.97 [−5.90, 26.32] | 90.5 | |
| Central | DF | 9.83 [2.85, 23.07] | 9.87 [3.37, 16.92] | 10.09 [−3.52, 26.62] | 93.6 |
| TR | 9.2 [1.92, 19.52] | 9.68 [2.47, 17.09] | 9.44 [−6.67, 29.03] | 92.7 | |
| WH | 13.86 [3.50, 26.84] | 8.58 [2.14, 15.29] | 8.72 [−5.56, 31.26] | 90.3 | |
| South | DF | 11.93 [4.03, 24.72] | 9.94 [3.91, 16.18] | 9.59 [−5.18, 30.00] | 89.3 |
| TR | 13.1 [3.47, 25.85] | 8.96 [2.76, 14.48] | 8.44 [−6.97, 28.16] | 87.5 | |
| WH | 11.25 [6.00, 18.68] | 8.40 [2.31, 13.55] | 8.61 [−5.97, 28.50] | 87.8 |
| (A) Structure | Poplar and Fir | Poplar Only | Fir Only a | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| All taxa | Fungi | NifH | AmoA | All taxa | Fungi | NifH | AmoA | All taxa | Fungi | NifH | ||
| Constrained by treatments | time | <0.001 | <0.001 | <0.001 | ns | <0.001 | <0.001 | <0.001 | ns | <0.001 | <0.001 | ns |
| litter type | <0.001 | 0.003 | 0.005 | ns | ns | ns | ns | ns | ns | ns | ns | |
| site | <0.001 | <0.001 | 0.008 | 0.003 | <0.001 | <0.001 | 0.031 | <0.001 | 0.013 | 0.026 | ns | |
| model | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.001 | <0.001 | <0.001 | <0.001 | ns | |
| Constrained by DNA | concentration | ns | ns | ns | 0.02 | 0.027 | ns | ns | 0.008 | 0.043 | ns | 0.018 |
| 260/280 b | 0.003 | 0.033 | 0.005 | ns | ns | ns | 0.036 | ns | ns | ns | ns | |
| 260/230 b | ns | ns | ns | ns | ns | ns | ns | ns | 0.026 | 0.002 | ns | |
| model | 0.002 | 0.029 | 0.005 | 0.023 | 0.021 | ns | 0.035 | 0.009 | 0.016 | 0.002 | 0.018 | |
| Partially constrained by DNA | time | <0.001 | <0.001 | <0.001 | ns | <0.001 | NA | 0.001 | ns | <0.001 | <0.001 | ns |
| litter type | 0.003 | 0.034 | 0.048 | ns | ns | NA | ns | ns | ns | ns | ns | |
| site | <0.001 | <0.001 | 0.008 | <0.001 | <0.001 | NA | 0.022 | <0.001 | 0.068 | 0.047 | ns | |
| model | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | NA | 0.001 | <0.001 | <0.001 | <0.001 | ns | |
| (B) Composition | Poplar and Fir | Poplar Only | Fir Only a | |||||||||
| All taxa | Fungi | NifH | AmoA | All taxa | Fungi | NifH | AmoA | All taxa | Fungi | NifH | ||
| Constrained by treatments | time | 0.005 | <0.001 | <0.001 | ns | <0.001 | <0.001 | <0.001 | ns | <0.001 | <0.001 | <0.001 |
| litter type | 0.005 | <0.001 | <0.001 | ns | 0.037 | 0.035 | ns | ns | 0.04 | 0.032 | ns | |
| site | 0.005 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.002 | |
| model | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Constrained by DNA | concentration | 0.005 | <0.001 | 0.034 | 0.009 | 0.002 | 0.008 | ns | <0.001 | ns | 0.029 | ns |
| 260/280 b | 0.007 | 0.009 | <0.001 | ns | ns | ns | 0.004 | ns | ns | ns | <0.007 | |
| 260/230 b | ns | ns | ns | ns | 0.03 | ns | 0.007 | ns | ns | ns | ns | |
| model | <0.001 | <0.001 | <0.001 | 0.006 | 0.012 | 0.006 | <0.001 | <0.001 | ns | 0.043 | 0.008 | |
| Partially constrained by DNA | time | <0.001 | <0.001 | <0.001 | ns | <0.001 | <0.001 | <0.001 | ns | NA | <0.001 | <0.001 |
| litter type | <0.001 | <0.001 | <0.001 | ns | 0.045 | 0.041 | ns | ns | NA | 0.076 | ns | |
| site | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | NA | <0.001 | <0.001 | |
| model | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | NA | <0.001 | <0.001 | |
| (A) Structure | Poplar and Fir | Poplar Only | Fir Only a | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| All taxa | Fungi | NifH | AmoA | All taxa | Fungi | NifH | AmoA | All taxa | Fungi | NifH | ||
| Partially constrained model significance | time | <0.001 | <0.001 | <0.001 | 0.366 | <0.001 | <0.001 | <0.001 | 0.346 | <0.001 | <0.001 | 0.1 |
| litter chem. | <0.001 | 0.004 | <0.001 | 0.004 | 0.508 | 0.376 | 0.611 | 0.183 | 0.461 | 0.451 | 0.734 | |
| climate | 0.192 | 0.546 | 0.121 | 0.631 | 0.188 | 0.064 | 0.162 | 0.614 | ns | ns | ns | |
| PCNM | 0.12 | 0.011 | 0.076 | 0.008 | 0.012 | <0.001 | 0.157 | 0.002 | 0.012 | ns | ns | |
| Predictor significance | litter chem. | N (0.026) CT (0.008) AUR (<0.001) | CT (0.027) AUR (0.029) | Ash (<0.001) | N (0.007) | CT (0.564) N (0.297) | CT (0.173) N (0.85) | CT (0.766) N (0.341) | CT (0.711) N b (0.078) | N (0.484) | N (0.424) | N (0.717) |
| climate | MS (0.068) MSmin (0.075) MSmax (0.72) TA (0.735) TAmin (ns) | MSmin (0.239) TAmin (0.959) | TA (0.121) | MS (0.771) TA (0.378) | MS (0.131) MSmax (0.433) TA (0.3) | MSmin (0.071) | TA (0.195) | MS (0.796) TA (0.332) | ns | ns | ns | |
| PCNM | PCNM 1 (0.007) PCNM 3 (0.826) PCNM 4 (0.682) PCNM 5 (ns) | PCNM 1 (0.003) PCNM 3 (0.672) PCNM 4 (0.462) | PCNM 3 (0.078) | PCNM 1 (0.003) PCNM 2 (0.583) PCNM 5 (0.122) | PCNM 1 (<0.001) PCNM 3 (0.219) PCNM 5 (0.832) | PCNM 1 (0.001) PCNM 3 (0.429) | PCNM 3 (0.151) | PCNM 1 (0.002) PCNM 2 (0.512) PCNM 5 (0.044) | PCNM 4 (0.01) | ns | ns | |
| (B) Composition | Poplar and Fir | Poplar Only | Fir Only a | |||||||||
| All taxa | Fungi | NifH | AmoA | All taxa | Fungi | NifH | AmoA | All taxa | Fungi | NifH | ||
| Model significance | time | <0.001 | <0.001 | <0.001 | 0.033 | <0.001 | <0.001 | <0.001 | 0.031 | <0.001 | <0.001 | <0.001 |
| litter chem. | <0.001 | <0.001 | <0.001 | 0.002 | 0.023 | 0.051 | 0.718 | 0.228 | 0.08 | 0.074 | 0.268 | |
| climate | <0.001 | 0.03 | <0.001 | 0.008 | <0.001 | 0.153 | <0.001 | 0.002 | 0.003 | <0.001 | 0.003 | |
| PCNM | 0.004 | <0.001 | <0.001 | <0.001 | 0.002 | <0.001 | 0.002 | <0.001 | 0.296 | ns | 0.131 | |
| Predictor significance | litter chem. | N (<0.001) CT (<0.001) AUR (<0.001) | CT (0.003) AUR (<0.001) Ash (<0.001) | Ash (<0.001) | N (0.002) | CT d (0.162) N (0.024) | CT b,c (0.059) N b (0.141) | CT (0.577) N (0.692) | CT (0.533) N b (0.132) | N (0.076) | N (0.077) | N (0.271) |
| climate | MSmin (0.009) MSmax (0.005) TA (0.03) TAmin (0.683) TSrange d (ns) PET (ns) | MSmin (0.012) TSrange (0.334) | MSmin (0.034) TAmin (0.026) TSmin (<0.001) TSrange (0.016) PET (ns) | TAmin (0.013) | MSmin (0.016) MSmax (0.006) TA (0.014) TAmin (0.628) TSrange (ns) PET (ns) | MSmin (0.474) MSmax (0.078) | MSmin (0.005) TA (0.01) TAmin (0.058) TSrange (0.001) PET (0.006) | TAmin (0.001) TAmax (0.049) | TAmin (0.076) TSrange (0.005) | TAmax (0.003) TSrange (0.01) | MS (0.095) TAmin (0.002) | |
| PCNM | PCNM 1 (0.002) PCNM 3 (0.215) PCNM 5 (ns) PCNM 6 (ns) | PCNM 1 (<0.001) PCNM 6 (0.01) | PCNM 1 (0.023) PCNM 3 (0.002) PCNM 4 (0.093) PCNM 5 (ns) | PCNM 1 (<0.001) PCNM 5 (0.001) | PCNM 1 (0.004) PCNM 3 (0.074) PCNM 5 (ns) PCNM 6 (ns) | PCNM 1 (<0.001) PCNM 3 (0.251) PCNM 6 (0.007) | PCNM 1 (0.139) PCNM 3 (0.001) PCNM 4 (0.078) | PCNM 1 (0.002) PCNM 5 (<0.001) | PCNM 3 (0.304) | ns | PCNM 3 (0.131) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shay, P.-E.; Winder, R.S.; Constabel, C.P.; Trofymow, J.A. Fungal Community Composition as Affected by Litter Chemistry and Weather during Four Years of Litter Decomposition in Rainshadow Coastal Douglas-Fir Forests. J. Fungi 2022, 8, 735. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8070735
Shay P-E, Winder RS, Constabel CP, Trofymow JA. Fungal Community Composition as Affected by Litter Chemistry and Weather during Four Years of Litter Decomposition in Rainshadow Coastal Douglas-Fir Forests. Journal of Fungi. 2022; 8(7):735. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8070735
Chicago/Turabian StyleShay, Philip-Edouard, Richard S. Winder, C. Peter Constabel, and J. A. (Tony) Trofymow. 2022. "Fungal Community Composition as Affected by Litter Chemistry and Weather during Four Years of Litter Decomposition in Rainshadow Coastal Douglas-Fir Forests" Journal of Fungi 8, no. 7: 735. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8070735