3.2. Taxonomy
Aurantioporia B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847338
Differs from other genera by its resupinate, rhizomorphic basidiocarps with an orange pore surface, a dimitic hyphal system with arboriform skeletal hyphae, tissues becoming violet in KOH, ellipsoid, truncate, and slightly dextrinoid basidiospores.
Type species: Aurantioporia bambusicola (Choeyklin, T. Hatt. & E.B.G. Jones) B.K. Cui & Xing Ji
Etymology: Aurantioporia (Lat.) refers to the orange pore surface of the genus.
Basidiocarps are annual to perennial and resupinate with rhizomorphs. The pore surface is yellow to orange when fresh, grayish orange, and orange-brown to dark orange when dry; pores are round to angular; dissepiments thin, entire. The subiculum is extremely thin and cream to pale orange. The tubes are concolorous with pore surface. The hyphal system is dimitic; generative hyphae with clamp connections; skeletal hyphae arboriform, IKI−, CB+; tissues become violet to dark in KOH. Basidiospores are ellipsoid, truncate, hyaline, thick-walled, smooth, slightly dextrinoid, and CB+.
Notes: In our study,
P. aurantiaca (A. David & Rajchenb.) Decock & Ryvarden and
P. bambusicola Choeyklin, T. Hatt. & E.B.G. Jones formed a single well-supported clade (82% BS, 1.00 BPP,
Figure 1; 89% BS, 1.00 BPP,
Figure 2), distant from the
Perenniporia s. s. clade. Morphologically, the two species differ from
Perenniporia s. s. species by the combination of a rhizomorphic basidiocarp with an orange pore surface, a dimitic hyphal system with arboriform skeletal hyphae, and tissues becoming violet in KOH. Therefore,
Aurantioporia gen. nov. is proposed to include
Perenniporia aurantiaca and
P. bambusicola.
Aurantioporia aurantiaca (A. David & Rajchenb.) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847362
Basionym: Pyrofomes aurantiacus A. David & Rajchenb., Mycotaxon 22(2): 312 (1985).
≡ Perenniporia aurantiaca (A. David & Rajchenb.) Decock & Ryvarden, Mycol. Res. 103(9): 1140 (1999).
For a detailed description of
Perenniporia aurantiaca, see David and Rajchenberg [
50] and Decock and Ryvarden [
4].
Notes:
Aurantioporia aurantiaca was originally described in
Pyrofomes by David and Rajchenberg [
50] and later was transferred to
Perenniporia by Decock and Ryvarden [
4]. The sequence of
Aurantioporia aurantiaca from French Guyana (type locality) fell into the
Aurantioporia clade in our phylogeny.
Aurantioporia bambusicola (Choeyklin, T. Hatt. & E.B.G. Jones) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847363
Basionym: Perenniporia bambusicola Choeyklin, T. Hatt. & E.B.G. Jones, Fungal Diversity 36: 122 (2009).
For a detailed description of
Perenniporia bambusicola, see Choeyklin et al. [
51] and Cui et al. [
3].
Notes:
Aurantioporia bambusicola was first described in
Perenniporia from Thailand [
51]. It is characterized by resupinate basidiocarps with an orange pore surface, a dimitic hyphal system with non-dextrinoid skeletal hyphae, tissues becoming violet to dark in KOH, oblong-ellipsoid, and truncate basidiospores. This species only grows on bamboo.
Aurantioporia aurantiaca also shares an orange pore surface, but
Aurantioporia aurantiaca grows on hardwood trees [
51].
Specimen examined: CHINA. Yunnan, Cangyuan County, Banlao, on bamboo, 11 July 2013, Cui 11050 (BJFC).
Key to species of Aurantioporia
1. Growing on bamboo; distributed in Southeast Asia ………………………………………………………………………………………………… A. bambusicola
1. Growing on other hardwoods; distributed in neotropical areas………………………………………………………………………………………A. aurantiaca
Citrinoporia B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847346
Differs from other genera by its slightly cushion shape, yellow pore surface, a dimitic hyphal system with dextrinoid, and cyanophilous shortly arboriform vegetative hyphae and ellipsoid, truncate, thick-walled, dextrinoid, and cyanophilous basidiospores.
Type species:
Etymology: Citrinoporia (Lat.) refers to the yellowish pore surface of the genus.
Basidiocarps are annual to perennial and resupinate. The pore surface is white to yellow; pores are round. Subiculum is cream to buff, corky. Tubes are buff to pale brown and corky to hard corky. The hyphal system is dimitic: generative hyphae with clamp connections; skeletal hyphae arboriform, IKI−, CB+; tissues becoming pale brown to black in KOH. Cystidia is absent, cystidioles are present. Basidiospores are ellipsoid, truncate, hyaline, thick-walled, smooth, dextrinoid, and CB+.
Notes: In our ITS + nLSU and five-gene phylogenetic analyses,
P. citrinoalba B.K. Cui, C.L. Zhao & Y.C. Dai and
P. corticola (Corner) Decock clustered together and formed a clade distinct from the
Perenniporia s. s clade with full support (100% BS, 1.00 BPP,
Figure 1; 100% BS, 1.00 BPP,
Figure 2). Morphologically, this clade differs from the
Perenniporia s. s. by its yellow pore surface and dimitic hyphal system. Thus, the new genus is set up and these two new combinations are proposed. Based on five-gene phylogenetic analysis,
Citrinoporia is sister to
Aurantioporia as they have the same overall morphology, yellow pore surface, dimitic hyphal system, and ellipsoid, truncate basidiospores, but the former differs in absence of rhizomorphs.
Citrinoporia citrinoalba (B.K. Cui, C.L. Zhao & Y.C. Dai) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847364
Basionym: Perenniporia citrinoalba B.K. Cui, C.L. Zhao & Y.C. Dai, Fungal Diversity 97: 270 (2019)
For a detailed description of
Perenniporia citrinoalba, see Cui et al. [
3].
Notes:
Citrinoporia citrinoalba was newly described in
Perenniporia from tropical China [
2]. It is characterized by annual and resupinate basidiocarps with white to yellow pore surfaces, a dimitic hyphal system with dextrinoid and cyanophilous skeletal hyphae, tissues becoming pale brown to black in KOH, and broadly ellipsoid and truncate basidiospores.
Specimens examined: CHINA. Hainan, Qiongzhong County, Limushan Forest Park, on fallen trunk of Castanopsis, 15 June 2014, Dai 13643 (holotype, BJFC), on fallen angiosperm trunk, 18 November 2015, Cui 13615 (BJFC).
Citrinoporia corticola (Corner) B.K. Cui & Xing Ji, comb. Nov.
MycoBank: MB 847365
Basionym: Parmastomyces corticola Corner, Beih. Nova Hedwigia 96: 96 (1989).
≡ Perenniporia corticola (Corner) Decock, Mycologia 93(4): 776 (2001).
= Perenniporia dipterocarpicola T. Hatt. & S.S. Lee, Mycologia 91(3): 525 (1999)
Notes:
Citrinoporia corticola was originally described in
Parmastomyces Kotl. & Pouzar from Malaysia by Corner [
52] as having a monomitic hyphal system with simple-septate generative hyphae. Decock [
53] studied the type specimens of
Parmastomyces corticola and confirmed that this species has a dimitic hyphal system with clamped generative hyphae and transferred the species to
Perenniporia.
Citrinoporia corticola and
Citrinoporia citrinoalba share yellow pore surfaces, dimitic hyphal structures, and truncate basidiospores, but the basidiospores of
C. citrinoalba (5.5–6 × 4.7–5.2 µm) [
3] are larger than those of
Citrinoporia corticola (4.4–5 × 3.4–4 μm) [
53].
Specimens examined: MALAYSIA. Selangor, Kota Damansara, Community Forest Reserve, on angiosperm stump, 17 April 2018 Dai 18633, 18641 (BJFC).
Key to species of Citrinoporia
1. Basidiospores 4.4–5 μm; growing mainly on trees of Dipterocarpaceae……………………………………………………………………………… C. corticola
1. Basidiospores 5.5–6 µm; growing on trees of Fagaceae ……………………………………………………………………………………………… C. citrinoalba
Cystidioporia B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847348
Differs from other genera by its resupinate basidiocarps, slightly dextrinoid and cyanophilous skeletal hyphae, presence of thick-walled cystidia, and thick-walled, oblong-ellipsoid, truncate, slightly dextrinoid, and cyanophilous basidiospores.
Type species: Cystidioporia piceicola (Y.C. Dai) B.K. Cui & Xing Ji
Etymology: Cystidioporia (Lat.) refers to resembling Perenniporia but with cystidia.
Basidiocarps are annual to biennial, resupinate, soft corky when fresh, and hard corky when dry. Pore surface is cream to buff when fresh and pale yellowish upon drying. Pores are round and large; dissepiments are thin, entire. Subiculum is yellowish ochraceous and corky. Tubes are yellowish ochraceous or straw yellow and corky. Hyphal system is dimitic to trimitic; generative hyphae with clamp connections; skeletal hyphae is slightly dextrinoid, CB+; tissues unchanged in KOH. Cystidia present, thick-walled, strongly CB+. Basidiospores are ellipsoid, truncate, hyaline, thick-walled, smooth, slightly dextrinoid, and CB+.
Notes: In our present phylogenetic analyses (
Figure 1 and
Figure 2), two specimens of
Perenniporia piceicola Y.C. Dai formed a single clade distant from the
Perenniporia s. s. clade. Moreover, this species has thick-walled cystidia, large pores, and basidiospores (pores 2–3 per mm, basidiospores 11–14 × 5.4–7.5 µm) [
8] which are different from other species of
Perenniporia. Thus, the new genus is set up, and the following combination is proposed.
Cystidioporia piceicola (Y.C. Dai) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847366
Basionym: Perenniporia piceicola Y.C. Dai, Ann. Bot. Fenn. 39(3): 173 (2002).
For a detailed description of
Perenniporia piceicola, see Dai et al. [
8].
Notes:
Cystidioporia piceicola was originally described in
Perenniporia by Dai et al. [
8]; it is characterized by resupinate basidiocarps, thick-walled cystidia, large pores and basidiospores, and usually grows on
Picea and
Abies.
Specimens examined: CHINA. Yunnan, Lijiang, Yunshanping, on fallen trunk of Picea likiangensis, 18 June 1999, Dai 3089 (isotype, BJFC), on fallen trunk of Abies, 16 September 2018, Cui 17062 (BJFC), and on fallen trunk of Picea, 16 September 2018, Cui 17069 (BJFC).
Dendroporia B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847349
Differs from other genera by annual and resupinate basidiocarps with gray to pale brown pore surface, a dimitic hyphal system with weakly dextrinoid skeletal hyphae, tissues darkening in KOH, presence of dendrohyphidia and large rhomboid crystals, and hyaline to pale yellowish, thick-walled, ellipsoid, truncate, non-dextrinoid, and cyanophilous basidiospores.
Type species: Dendroporia cinereofusca (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji
Etymology: Dendroporia (Lat.) refers to the presence of dendrohyphidia.
Basidiocarps are annual, resupinate, adnate, and corky. Pore surface is gray to pale brown. Subiculum is thin and clay buff to pale brown. Tubes are concolorous with pore surface and corky. Hyphal system is dimitic; generative hyphae with clamp connections; skeletal hyphae weakly dextrinoid and CB+; tissues are brown to black in KOH. Dendrohyphidia present at dissepimental edges; cystidia are absent; cystidioles are present. Large rhomboid crystals are present. Basidiospores ellipsoid, truncate, hyaline to pale yellowish, thick-walled, smooth, IKI−, and CB+.
Notes: In our present phylogeny, two specimens of
Perenniporia cinereofusca B.K. Cui & C.L. Zhao formed a strongly supported clade distinct from the
Perenniporia s. s. clade (
Figure 1 and
Figure 2). Morphologically,
P. cinereofusca differs from species of
Perenniporia s. s. by its resupinate basidiocarps with a gray to pale brown pore surface, tissues darkening in KOH, hyaline to pale yellowish, and non-dextrinoid basidiospores. Thus, the new genus
Dendroporia is set up and the new combination
Dendroporia cinereofusca is proposed.
In the current phylogenetic studies,
Dendroporia is related to
Tropicoporia and
Sparsitubus, but with only weak support, and
Tropicoporia differs from
Dendroporia by its buff-yellow to grayish orange pore surface, non-dextrinoid skeletal hyphae, and dextrinoid basidiospores.
Sparsitubus differs from
Dendroporia in having effused reflexed to pileate basidiocarps and non-truncate, ornamented basidiospores [
54].
Dendroporia cinereofusca (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847367
Basionym: Perenniporia cinereofusca B.K. Cui & C.L. Zhao, Mycoscience 55: 419 (2014).
For a detailed description of
Perenniporia cinereofusca, see Zhao et al. [
20].
Notes:
Dendroporia cinereofusca was first described in
Perenniporia from tropical China [
20] and is characterized by its resupinate basidiocarps with gray to pale brown pore surfaces, a dimitic hyphal system with weakly dextrinoid skeletal hyphae, tissues becoming brown to black in KOH, the presence of dendrohyphidia, and hyaline to pale yellowish, truncate, and non-dextrinoid basidiospores.
Specimens examined: CHINA. Hainan, Ledong County, Jianfengling Nature Reserve, on fallen angiosperm trunk, 18 November 2007, Dai 9289 (holotype, BJFC); Lingshui County, Diaoluoshan Forest Park, on fallen angiosperm trunk, 20 November 2007, Cui 5280 (paratype, BJFC).
Luteoperenniporia B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847350
Differs from other genera by its resupinate basidiocarps with buff-yellow to cinnamon-buff pore surface, a dimitic hyphal system with weak to strong dextrinoid skeletal hyphae, the presence of cystidioles, and thick-walled, ellipsoid, and non-truncate, dextrinoid, and cyanophilous basidiospores.
Type species: Luteoperenniporia bannaensis (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji
Etymology: Luteoperenniporia (Lat.) refers to resembling Perenniporia but with a buff-yellow pore surface when dry.
Basidiocarps are annual to perennial and resupinate. Pore surface iscream, buff to pale cinnamon buff when fresh, and becoming buff, buff-yellow to cinnamon-buff upon drying; pores are round to angular; dissepiments thin, entire to lacerate. Subiculum is thin and buff to cinnamon-buff. Tubes are concolorous with pore surface and corky. Hyphal system is dimitic, generative hyphae with clamp connections; skeletal hyphae weakly to strongly dextrinoid, CB+; tissues are unchanged in KOH. Cystidia is absent; cystidioles are present. Basidiospores are ellipsoid, non-truncate, hyaline, thick-walled, smooth, dextrinoid, and CB+.
Notes: In the combined ITS + nLSU and five-gene phylogenetic analyses, the species of
Luteoperenniporia formed a single clade with high support (
Figure 1 and
Figure 2) distant from the
Perenniporia s. s. clade. Morphologically,
Luteoperenniporia differs from
Perenniporia s. s. by its buff, buff-yellow to cinnamon-buff pore surfaces, a dimitic hyphal system with weakly to strongly dextrinoid skeletal hyphae and non-truncate basidiospores. Therefore, three new combinations are proposed in
Luteoperenniporia, and the new species is described below.
Luteoperenniporia bannaensis (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847368
Basionym: Perenniporia bannaensis B.K. Cui & C.L. Zhao, Fungal Diversity 58: 52 (2013).
For a detailed description of
Perenniporia bannaensis, see Zhao et al. [
19].
Notes:
Luteoperenniporia bannaensis was recently described in
Perenniporia from China by Zhao et al. [
19]; it is closely related to
L. yinggelingensis in morphology and phylogeny; they share annual and resupinate basidiocarps, cream to buff pore surface and a dimitic hyphal system with dextrinoid and cyanophilous skeletal hyphae, and both species are distributed in the tropics. However,
L. yinggelingensis is distinguished from
L. bannaensis by its larger pores and basidiospores (pores 5–6 per mm, basidiospores 6.2–7.5 × 4.5–5.5 μm) [
3].
Specimens examined: CHINA. Yunnan, Xishuangbanna, Mengla County, Wangtianshu Nature Reserve, on fallen angiosperm trunk, 2 November 2009, Cui 8560 (holotype, BJFC), Cui 8562 (paratype, BJFC).
Luteoperenniporia mopanshanensis (C.L. Zhao) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847369
Basionym: Perenniporia mopanshanensis C.L. Zhao, Mycotaxon 134(1): 132 (2019).
For a detailed description of
Perenniporia mopanshanensis, see Zhao and Ma [
28].
Notes:
Luteoperenniporia mopanshanensis was recently described in
Perenniporia by Zhao and Ma [
28].
L. mopanshanensis and
L. bannaensis are both reported from Yunnan Province in southern China. They share resupinate basidiocarps, a dimitic hyphal system with strongly dextrinoid skeletal hyphae, non-truncate, and strongly dextrinoid and similar sized basidiospores. However,
L. bannaensis differs by having an annual growth habit and smaller pores (6–8 per mm) [
19].
Luteoperenniporia yinggelingensis (B.K. Cui & Y.C. Dai) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847370
Basionym: Perenniporia yinggelingensis B.K. Cui & Y.C. Dai, Fungal Diversity 97: 300 (2019).
For a detailed description of
Perenniporia yinggelingensis, see Cui et al. [
3].
Notes:
Luteoperenniporia yinggelingensis was newly described from a tropical area of China [
3]. It is characterized by annual and resupinate basidiocarps with slightly lacerate pores, a distinct sterile margin, and a dimitic hyphal system with weakly dextrinoid skeletal hyphae. Macroscopically,
L. yinggelingensis is close to
L. mopanshanensis, but the latter species has perennial basidiocarps, larger pores (3–5 per mm), indistinct sterile margins, and strongly dextrinoid skeletal hyphae [
28].
Specimens examined: CHINA. Hainan, Baisha County, Yinggeling Nature Reserve, on fallen angiosperm trunk, 17 November 2015, Cui 13625 (holotype, BJFC); on fallen angiosperm branch, 17 June 2016, Cui 13856 (BJFC).
Luteoperenniporia australiensis B.K. Cui & Xing Ji, sp. nov.;
Figure 3 and
Figure 4 MycoBank: MB 847371
Differs from other species of Luteoperenniporia by its annual to perennial growth habit, resupinate basidiocarps with buff to cinnamon-buff pore surfaces, a dimitic hyphal system with dextrinoid skeletal hyphae, and ellipsoid, non-truncate, dextrinoid basidiospores (6.2–7.5 × 4–5.2 μm).
Holotype: AUSTRALIA. Tasmania, Keogh’s Creek Walk, on fallen trunk of Eucalyptus, 15 May 2018, Cui 16743 (BJFC).
Etymology: australiensis (Lat.) refers to the country where the new species was found.
Fruitbody: Basidiocarps are annual to perennial, resupinate, without odor or taste when fresh, corky to hard corky when dry, and up to 14 cm long, 8.5 cm wide, and 7 mm thick at center. Pore surface is buff-yellow, pinkish buff to pale cinnamon-buff when fresh, and buff, pale yellowish-brown to cinnamon-buff upon drying; pores are round to angular, 4–6 per mm, dissepiments thin, entire to lacerate. Sterile margin is distinct to indistinct, buff, and up to 2 mm wide. Subiculum is thin, buff, and up to 1 mm thick. Tubes are concolorous with pore surface, corky to hard corky when dry, and up to 6 mm long.
Hyphal structure: Hyphal system is dimitic; generative hyphae bearing clamp connections; skeletal hyphae dextrinoid, and CB+; tissues are unchanged in KOH.
Subiculum: Generative hyphae is infrequent, hyaline, thin-walled, occasionally branched, and 1.5–2.5 μm in diameter; skeletal hyphae is dominant, hyaline, thick-walled with a wide to narrow lumen, rarely branched, interwoven, and 2–5 μm in diameter.
Tubes: Generative hyphae is infrequent, hyaline, thin-walled, occasionally branched, and 1–2.4 μm in diameter; skeletal hyphae is dominant, hyaline, thick-walled with a wide to narrow lumen, occasionally branched, interwoven, and 1–3 μm. Cystidia are absent; fusoid cystidioles are present, hyaline, thin-walled, and 15.5–22 × 5.8–8 μm. Basidia clavate, with four sterigmata and a basal clamp connection, 15–24.5 × 6.5–9.7 μm; basidioles are dominant and in shape similar to basidia, but slightly smaller.
Spores: Basidiospores are ellipsoid, non-truncate, hyaline, thick-walled, smooth, dextrinoid, CB+, (6–) 6.2–7.4 (–7.7) × (3.9–) 4–5.2 (–5.4) μm, L = 6.77 μm, W = 4.58 μm, Q = 1.44–1.54 (n = 90/3).
Notes:
Luteoperenniporia australiensis is characterized by its annual to perennial and resupinate basidiocarps, buff to cinnamon-buff pore surface, entire to lacerate pores, a dimitic hyphal system with dextrinoid skeletal hyphae, presence of cystidioles, and ellipsoid, non-truncate, dextrinoid, and cyanophilous basidiospores.
Luteoperenniporia bannaensis is similar to
L. australiensis by sharing a buff-yellow to pinkish buff pore surface, a dimitic hyphal system with dextrinoid, and cyanophilous skeletal hyphae. However,
L. bannaensis differs from
L. australiensis by its smaller pores (6–8 per mm) and smaller basidiospores (5.2–6 × 4–4.6 μm) [
19].
Luteoperenniporia yinggelingensis may be confused with
L. australiensis by having resupinate basidiocarps, and similar pore size and basidiospores. However,
L. yinggelingensis is distinguished from
L. australiensis mainly by its annual growth habit and cream to buff pore surface [
3].
Perenniporia subaurantiaca (Rodway & Cleland) P.K. Buchanan & Ryvarden is also described from Tasmania, it is similar to
Luteoperenniporia australiensis in its resupinate basidiocarps, dimitic hyphal system with dextrinoid and cyanophilous skeletal hyphae, and presence of cystidioles, but
P. subaurantiaca has greyish cream to greyish orange pore surfaces and larger basidiospores (7.2–9.5 × 4.2–5.5 μm) [
55]
Additional specimens (paratypes) examined: AUSTRALIA. Victoria, Yarra Ranges National Park, at the base of living Eucalyptus, 9 May 2018, Cui 16524 (BJFC), on fallen trunk of Eucalyptus, 9 May 2018, Cui 16525 (BJFC), 10 May 2018, Cui 16535 (BJFC), on living tree of Eucalyptus, 10 May 2018, Cui 16533, Cui 16534 (BJFC); Tasmania, Keogh’s Creek Walk, on fallen trunk of Eucalyptus, 15 May 2018, Cui 16742 (BJFC).
Key to species of Luteoperenniporia
1. Pores 6–8 per mm……………………………………………………………………………………………………………………………………………L. bannaensis
1. Pores 3–6 per mm…………………………………………………………………………………………………………………………………………………………2
2. Skeletal hyphae unbranched…………………………………………………………………………………………………………………………L. mopanshanensis
2. Skeletal hyphae branched ……………………………………………………………………………………………………………………………………………… 3
3. Basidiocarps annual, distributed in tropical areas…………………………………………………………………………………………………L. yinggelingensis
3. Basidiocarps annual to perennial, distributed in temperate to subtropical areas…………………………………………………………………L. australiensis
Macroporia B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847352
Differs from other genera by its annual and resupinate basidiocarps, a dimitic hyphal system with dextrinoid skeletal hyphae, the presence of thin-walled cystidioles, and hyaline, thick-walled, ellipsoid, truncate, dextrinoid, and cyanophilous basidiospores.
Type species: Macroporia macropora (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji
Etymology: Macroporia (Lat.) refers to the species with relatively large pores in Perenniporia.
Basidiocarps are annual, resupinate, adnate. Pore surface is white, cream to buff when fresh, and becoming buff, pinkish buff to yellowish buff upon drying; pores are angular; dissepiments are thin, entire to lacerate. Subiculum is thin, cream. Tubes are concolorous with pore surface, corky to hard corky. Hyphal system is dimitic, generative hyphae with clamp connections; skeletal hyphae branched, dextrinoid, CB+; tissues are unchanged in KOH. Cystidia is absent; cystidioles are usually present. Basidiospores are ellipsoid, truncate, hyaline, thick-walled, smooth, dextrinoid, and CB+.
Notes: In our ITS + nLSU and combined five-gene phylogenetic analyses,
Perenniporia lacerata B.K. Cui & C.L. Zhao,
P. macropora B.K. Cui & C.L. Zhao,
P. subrhizomorpha Xue W. Wang, L.W. Zhou & X.M. Tian and
P. tibetica B.K. Cui & C.L. Zhao grouped together and formed a well-supported clade (
Figure 1 and
Figure 2), which was distant from the
Perenniporia s. s. clade. Morphologically, species in the clade usually have larger pores than those of
Perenniporia s. s.; therefore, the new genus
Macroporia is proposed to accommodate the four species.
Macroporia lacerata (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847372
Basionym: Perenniporia lacerata B.K. Cui & C.L. Zhao, Mycoscience 54: 232 (2013).
For a detailed description of
Perenniporia lacerata, see Zhao and Cui [
18].
Notes:
Macroporia lacerata was originally described in
Perenniporia from China by Zhao and Cui [
18]. It is characterized by papery and thin basidiocarps, lacerate pores, a dimitic hyphal system with weakly dextrinoid skeletal hyphae, and ellipsoid, truncate, dextrinoid, and cyanophilous basidiospores.
Specimens examined: CHINA. Henan, Xiuwu County, Yuntaishan Park, on fallen angiosperm trunk, 3 September 2009, Cui 7220 (holotype, BJFC); Neixiang County, Baotianman Nature Reserve, on rotten angiosperm wood, 22 September 2009, Dai 11268 (paratype, BJFC).
Macroporia macropora (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847373
Basionym: Perenniporia macropora B.K. Cui & C.L. Zhao, Mycologia 105: 947 (2013).
For a detailed description of
Perenniporia macropora, see Zhao and Cui [
17].
Notes:
Macroporia macropora was first described in
Perenniporia from southern China by Zhao and Cui [
17]. It is distinguished by large pores (2–3 per mm), a dimitic hyphal system with dextrinoid and branched skeletal hyphae, the presence of dendrohyphidia, and ellipsoid and truncate basidiospores.
M. macropora is very close to
M. lacerata in the current phylogeny, but
M. lacerata differs from
M. macropora by its smaller (3–5 per mm) and lacerate pores, the absence of dendrohyphidia, and smaller basidiospores (6.1–7 × 5–5.7 μm) [
18].
Specimens examined: CHINA. Guangxi, Ningming County, Nonggang Nature Reserve, on fallen angiosperm branch, 8 July 2007, Zhou 407 (holotype, IFP), 7 July 2007, Zhou 297 (paratype, IFP).
Macroporia subrhizomorpha (Xue W. Wang, L.W. Zhou & X.M. Tian) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847374
Basionym: Perenniporia subrhizomorpha Xue W. Wang, L.W. Zhou & X.M. Tian, Phytotaxa 528(2): 129 (2021).
For a detailed description of
Perenniporia subrhizomorpha, see Tian et al. [
56].
Notes:
Macroporia subrhizomorpha was recently described in
Perenniporia as
P. subrhizomorpha by Tian et al. [
56].
Perenniporia rhizomorpha may be confused with
P. subrhizomorpha by sharing cream rhizomorphs and similar pore sizes, but the former species differs by its non-truncate basidiospores [
10].
Perenniporia tibetica also has rhizomorphs but differs from
P. subrhizomorpha by larger pores (2–3 per mm) [
15].
Macroporia tibetica (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847375
Basionym: Perenniporia tibetica B.K. Cui & C.L. Zhao, Mycoscience 53: 366 (2012).
For a detailed description of
Perenniporia tibetica, see Cui and Zhao [
15].
Notes:
Macroporia tibetica is characterized by resupinate basidiocarps with white to cream rhizomorphs, a dimitic hyphal system with slightly dextrinoid skeletal hyphae, and ellipsoid, truncate or not, dextrinoid, and cyanophilous basidiospores.
M. macropora and
M. tibetica share resupinate basidiocarps and similar pores, but the former differs in having dendrohyphidia and lacking rhizomorphs [
17].
Specimens examined: CHINA. Xizang, Linzhi County, Tongmai, on fallen angiosperm trunk, 16 September 2010, Cui 9457 (holotype, BJFC), Cui 9459 (paratype, BJFC).
Key to species of Macroporia
1. Pores lacerate ………………………………………………………………………………………………………………………………………………… M. lacerata
1. Pores entire ……………………………………………………………………………………………………………………………………………………………… 2
2. Basidiocarps without rhizomorphs……………………………………………………………………………………………………………………… M. macropora
2. Basidiocarps with rhizomorphs…………………………………………………………………………………………………………………………………………3
3. Pores 2–3 per mm………………………………………………………………………………………………………………………………………………M. tibetica
3. Pores 4–6 per mm …………………………………………………………………………………………………………………………………… M. subrhizomorpha
Macrosporia B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847353
Differs from other genera by its annual and resupinate basidiocarps, cinnamon-buff pore surface, a trimitic hyphal system with weakly dextrinoid and cyanophilous skeletal hyphae, and hyaline, thick-walled, ellipsoid, truncate, strongly dextrinoid, and cyanophilous basidiospores.
Type species: Macrosporia nanlingensis (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji
Etymology: Macrosporia (Lat.) refers to the large basidiospores.
Basidiocarps are annual, resupinate, adnate, corky when fresh, and becoming hard corky upon drying. Pore surface is cream-buff to yellowish buff when fresh, becoming cinnamon-buff upon drying; pores are round; dissepiments are thick, entire. Subiculum is cream to buff. Tubes are concolorous with the pore surface and hard corky. Hyphal system is trimitic; generative hyphae with clamp connections; skeletal and binding hyphae are weakly dextrinoid and CB+. Cystidia is absent; cystidioles are present. Basidiospores areellipsoid, truncate, hyaline, thick-walled, smooth, strongly dextrinoid, and CB+.
Notes: In our study, Perenniporia nanlingensis B.K. Cui & C.L. Zhao formed a single clade that was distant from the Perenniporia s. s. clade. Morphologically, it differs from Perenniporia s. s. by its annual and resupinate basidiocarps with cinnamon-buff pore surfaces and larger basidiospores. Therefore, Macrosporia gen. nov. is proposed to accommodate P. nanlingensis.
Macrosporia nanlingensis (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847376
Basionym: Perenniporia nanlingensis B.K. Cui & C.L. Zhao, Mycol. Prog. 11: 556 (2012).
For a detailed description of
Perenniporia nanlingensis, see Zhao and Cui [
16].
Notes:
Macrosporia nanlingensis was first described in
Perenniporia from southern China [
16]; it is characterized by annual and resupinate basidiocarps, a cinnamon-buff pore surface when dry, a trimitic hyphal system with slightly dextrinoid skeletal hyphae, and ellipsoid, truncate, strongly dextrinoid, and cyanophilous basidiospores that are usually longer than 9 μm.
Specimens examined: CHINA. Guangdong Province, Ruyang County, Nanling Nature Reserve, on dead angiosperm tree, 16 September 2009, Cui 7589 (holotype, BJFC), Cui 7620 (paratype, BJFC).
Minoporus B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847354
Differs from other genera by its annual and pileate basidiocarps, cream to pale buff pileal surface when fresh, a dimitic hyphal system with weakly amyloid and cyanophilous skeletal hyphae, and hyaline, thick-walled, ellipsoid, truncate, dextrinoid, and cyanophilous basidiospores.
Type species: Minoporus minor (Y.C. Dai & H.X. Xiong) B.K. Cui & Xing Ji
Etymology: Minoporus (Lat.) refers to the small pilei.
Basidiocarps are annual, pileate, solitary, and soft corky when fresh, becoming hard corky upon drying. Pilei are semicircular to spathulate. Pileal surface is cream to pale buff when fresh, becoming cinnamon-buff when dry. Pore surface is cream when fresh, becoming cinnamon-buff when dry; pores are round. Context is white to cream, corky. Tubes are concolorous with pore surface and hard corky. Hyphal system dimitic; generative hyphae with clamp connections; skeletal hyphae is weakly amyloid and CB+. Cystidia and cystidioles are absent. Basidiospores are ellipsoid, truncate, hyaline, thick-walled, smooth, dextrinoid, and CB+.
Notes: In the ITS + nLSU analysis, two specimens of
Perenniporia minor Y.C. Dai & H.X. Xiong formed a highly supported single clade (
Figure 1), which is distinct from the
Perenniporia s. s. clade and closely related to the
Perenniporiella clade. Further phylogeny (
Figure 2) inferred from the combined five-gene dataset indicated that the clade of
P. minor was distant from the
Perenniporia s. s. clade and closely related to the
Neoporia clade. However,
Neoporia has resupinate basidiocarps, dextrinoid skeletal hyphae, and non-truncate basidiospores;
Perenniporiella has dextrinoid skeletal hyphae and non-truncate basidiospores [
30].
Perenniporia minor differs from species of
Perenniporia s. s. by its annual and pileate basidiocarps with a cream to pale buff pileal surface, a dimitic hyphal system with weakly amyloid skeletal hyphae. Therefore,
Minoporus gen. nov. is proposed to accommodate
P. minor.
Minoporus minor (Y.C. Dai & H.X. Xiong) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847377
Basionym: Perenniporia minor Y.C. Dai & H.X. Xiong, Mycotaxon 105: 60 (2008).
For a detailed description of
Perenniporia minor, see Xiong et al. [
11].
Notes: This species was described from northeastern China by Xiong et al. [
11] and is characterized by annual and pileate basidiocarps, cream to pale buff when fresh and cinnamon buff when dry pileal surface, a dimitic hyphal system with weakly amyloid skeletal hyphae, and ellipsoid, truncate, dextrinoid, and cyanophilous basidiospores.
Specimens examined: CHINA. Jilin, Antu County, Changbaishan Nature Reserve, Huangsongpu, on fallen branch of Acer, 14 September 2007, Dai 9198 (holotype, IFP); Liaoning, Huanren County, Laotudingzi Nature Reserve, on fallen branch of Quercus, 31 July 2008, Cui 5738 (BJFC).
Neoporia B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847355
Differs from other genera by its annual and resupinate basidiocarps with a buff-yellow pore surface, a dimitic hyphal system with dextrinoid and cyanophilous skeletal hyphae, and ellipsoid, non-truncate, dextrinoid, and cyanophilous basidiospores.
Type species: Neoporia rhizomorpha (B.K. Cui, Y.C. Dai & Decock) B.K. Cui & Xing Ji
Etymology: Neoporia (Lat.) refers to the genus resembling Perenniporia.
Basidiocarps are annual, resupinate and corky when dry. Pore surface is cream to buff when fresh, buff-yellow to grayish orange upon drying; pores are round to angular. Subiculum is cream to buff. Tubes are concolorous with the pore surface and corky. Hyphal system is dimitic; generative hyphae with clamp connections; skeletal hyphae dextrinoid, CB+; tissues unchanged in KOH. Basidiospores are ellipsoid, non-truncate, hyaline, thick-walled, smooth, dextrinoid, and CB+.
Notes: In our phylogenetic analyses, three species previously included in
Perenniporia,
P. bostonensis C.L. Zhao,
P. koreana Y. Jang & J.J. Kim, and
P. rhizomorpha B.K. Cui, Y.C. Dai & Decock grouped together and formed a highly supported clade (100% BS, 1.00 BPP,
Figure 1; 100% BS, 1.00 BPP,
Figure 2), which was distant from the
Perenniporia s. s. clade. Morphologically, this clade differs from
Perenniporia s. s. by its dimitic hyphal system and non-truncate basidiospores. Therefore,
Neoporia gen. nov. is proposed to accommodate
P. bostonensis,
P. koreana and
P. rhizomorpha.
In the five-gene phylogenetic analysis,
Neoporia is related to
Luteoperenniporia,
Minoporus,
Poriella,
Vanderbyliella, and
Yuchengia.
Luteoperenniporia,
Neoporia,
Poriella,
Vanderbyliella, and
Yuchengia all have ellipsoid and non-truncate basidiospores. However,
Luteoperenniporia differs from
Neoporia in having a perennial growth habit;
Poriella differs from
Neoporia by having a perennial growth habit, cinnamon to ochraceous pore surfaces, and unbranched skeletal hyphae;
Vanderbyliella is different from
Neoporia by its pileate basidiocarps;
Yuchengia differs from
Neoporia in having amyloid skeletal hyphae and non-dextrinoid basidiospores [
35]. In addition,
Minoporus differs from
Neoporia by its pileate basidiocarps, amyloid skeletal hyphae, and truncate basidiospores.
Neoporia bostonensis (C.L. Zhao) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847378
Basionym: Perenniporia bostonensis C.L. Zhao, Phytotaxa 351(1): 67 (2018).
For a detailed description of
Perenniporia bostonensis, see Shen et al. [
27].
Notes:
Neoporia bostonensis was recently described in
Perenniporia from North America by Shen et al. [
27]. It is characterized by resupinate basidiocarps with cream to buff pore surfaces, a dimitic hyphal system with strongly dextrinoid and unbranched skeletal hyphae, and ovoid to broad ellipsoid, non-truncate, and dextrinoid basidiospores.
Neoporia koreana (Y. Jang & J.J. Kim) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847379
Basionym: Perenniporia koreana Y. Jang & J.J. Kim, Mycotaxon 130(1): 174 (2015).
For a detailed description of
Perenniporia bannaensis, see Jang et al. [
22].
Notes:
Neoporia koreana was originally described from Republic of Korea as
Perenniporia koreana by Jang et al. [
22]; it has annual and resupinate basidiocarps with grayish orange pore surfaces, dextrinoid skeletal hyphae, and ellipsoid and non-truncate basidiospores. The sequences of
Neoporia koreana from type specimens fell into
Neoporia in our phylogeny. Thus,
P. koreana is transferred to
Neoporia.
N. koreana is similar to
N. bostonensis in resupinate basidiocarps, similar sized pores, dextrinoid skeletal hyphae, and non-truncate basidiospores, but the former has larger basidiospores (6–7 × 3.9–5.2 μm) [
22] and the latter has smaller basidiospores (3.5–4.5 × 3–4 μm) [
27].
Neoporia rhizomorpha (B.K. Cui, Y.C. Dai & Decock) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847380
Basionym: Perenniporia rhizomorpha B.K. Cui, Y.C. Dai & Decock, Mycotaxon 99: 176 (2007).
For a detailed description of
Perenniporia rhizomorpha, see Cui et al. [
10].
Notes:
Neoporia rhizomorpha was first described in
Perenniporia based on morphological characters from China [
10]. It is unique in
Neoporia due to its resupinate basidiocarps with rhizomorphs.
Specimens examined: CHINA. Anhui, Huangshan, Yellow Mountain, on fallen angiosperm trunk, 13 October 2004, Dai 6165 (holotype, BJFC); Fujian, Wuyishan County, Wuyishan Nature Reserve, on fallen angiosperm branch, 19 October 2005, Cui 7248 (paratype, BJFC).
Key to species of Neoporia
1. Basidiocarps with rhizomorphs…………………………………………………………………………………………………………………………N. rhizomorpha
1. Basidiocarps without rhizomorphs…………………………………………………………………………………………………………………………………… 2
2. Basidiospores 3.5–4.5 × 3–4 μm ………………………………………………………………………………………………………………………… N. bostonensis
2. Basidiospores 6–7 × 3.9–5.2 μm………………………………………………………………………………………………………………………………N. koreana
Niveoporia B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847356
Differs from other genera by perennial basidiocarps with white pore surface when fresh, distinct rusty red to reddish brown sterile margin, a dimitic hyphal system with dextrinoid and cyanophilous skeletal hyphae, the presence of cystidioles, hyaline, and thick-walled, ellipsoid, and truncate basidiospores.
Type species: Niveoporia russeimarginata (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji
Etymology: Niveoporia (Lat.) refers to the white pore surface.
Basidiocarps are perennial, resupinate to pileate, corky to woody hard when dry. Pilei dimidiate to fan shaped. Pileal surface is clay-buff to reddish brown when fresh, grayish brown to umber brown when dry, glabrous, and concentrically sulcate. Pore surface is white when fresh and white to cream upon drying; pores are round. The sterile margin is sometimes distinct, rusty red to reddish brown. Context is buff to fawn. Tubes are cream to pale cinnamon. Hyphal system is dimitic; generative hyphae with clamp connections; skeletal hyphae dextrinoid, CB+; tissues unchanged in KOH. Cystidia is absent; cystidioles are present. Basidiospores are ellipsoid, truncate, hyaline, thick-walled, smooth, dextrinoid or not, and CB+.
Notes: In the phylogenetic analysis of this study,
P. decurrata F. Wu & X.H. Ji,
P. russeimarginata B.K. Cui & C.L. Zhao and an undescribed species grouped together and formed a well-supported clade (74% BS, 1.00 BPP,
Figure 1; 83% BS, 0.97 BPP,
Figure 2) which was separated from the
Perenniporia s. s. clade. Morphologically, species in the clade usually have pileate basidiocarps and white pore surfaces with reddish brown sterile margins, which are different from the species of
Perenniporia s. s. Thus,
Niveoporia gen. nov. is proposed to accommodate these species.
Niveoporia decurrata (Corner) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847381
Basionym: Perenniporia decurrata Corner, Beih. Nova Hedwigia 96: 105 (1989)
= Perenniporia chiangraiensis F. Wu & X.H. Ji, Mycosphere 8(8): 1103 (2017).
For a detailed description of
Perenniporia decurrata, see Corner [
52].
Notes:
Perenniporia decurrata was first described from Malaysia [
52].
Perenniporia chiangraiensis was recently described from Northern Thailand based on morphological characters and molecular data by Ji et al. [
25]. However, these authors overlooked
P. decurrata, which is a priority name for this species. The species is characterized by pileate basidiocarps with concentrically sulcate pileal surfaces, white pore surfaces, the presence of dendrohyphidia and cystidioles, and ellipsoid, truncate, thick-walled, and non-dextrinoid basidiospores.
Specimens examined: CHINA. Yunnan, Xishuangbanna, Menglun, on fallen angiosperm trunk, 12 September 2006, Yuan 2334 (IFP). THAILAND. Chiang Rai, Doi Mae Salong, on angiosperm tree root, 22 July 2016, Dai 16637 (BJFC).
Niveoporia russeimarginata (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847382
Basionym: Perenniporia russeimarginata B.K. Cui & C.L. Zhao, Mycologia 105(4): 947 (2013).
For a detailed description of
Perenniporia russeimarginata, see Zhao and Cui [
17].
Notes:
Niveoporia russeimarginata was first described in
Perenniporia from southern China [
17].
Perenniporia alboferruginea Decock, described from Cameroon in Africa, is similar to
N. russeimarginata in having resupinate basidiocarps with ferruginous red upper margins and a dimitic hyphal system. However,
P. alboferruginea differs by having an annual growth habit, larger pores (5–6 per mm), the absence of cystidioles, and non-dextrinoid basidiospores [
57].
Specimens examined: CHINA. Yunnan, Chuxiong, Zixishan Nature Reserve, on fallen angiosperm trunk, 1 August 2005, Yuan 1225 (holotype, IFP), Yuan 1262 (paratype, IFP).
Niveoporia subrusseimarginata B.K. Cui & Xing Ji, sp. nov.,
Figure 5 and
Figure 6 MycoBank: MB 847383
Differs from other Niveoporia species by its resupinate to pileate basidiocarps with rusty reddish brown sterile margins and pores measuring 5–6 per mm.
Holotype: CHINA. Yunnan, Binchuan County, Jizushan Park, on angiosperm stump, 14 September 2018, Cui 16991 (BJFC).
Etymology: Subrusseimarginata (Lat.) refers to the species resembling Niveoporia russeimarginata.
Fruitbody: Basidiocarps are perennial, resupinate, sometimes pileate, corky, without odor or taste when fresh, becoming woody hard upon drying. Pilei are irregular, projecting up to 2 cm, 5 cm wide, and 3 cm thick at the base; with resupinate up to 9 cm long, 5 cm wide, and 1.3 cm thick at center. Pileal surface is orange-brown to reddish brown when fresh, umber brown when dry, glabrous, and concentrically sulcate. The pore surface is white when fresh, cream upon drying; pores are round to angular, 5–6 per mm, dissepiments thick, entire. Sterile margin is distinct to indistinct, cinnamon to rusty reddish brown, and up to 2 mm wide. Subiculum is buff, thin, up to 0.5 mm thick. Tubes are buff, woody hard when dry, and up to 11.5 mm long.
Hyphal structure: Hyphal system is dimitic; generative hyphae bearing clamp connections; skeletal hyphae is weakly dextrinoid and CB+; tissues are unchanged in KOH.
Subiculum: Generative hyphae are infrequent, hyaline, thin-walled, unbranched, and 1–2.2 μm in diameter; skeletal hyphae are dominant, hyaline, thick-walled, occasionally branched, interwoven, and 1–2.5 μm in diameter.
Tubes: Generative hyphae are infrequent, hyaline, thin-walled, occasionally branched, and 1–2 μm in diameter; skeletal hyphae are dominant, hyaline, thick-walled, moderately branched, and interwoven, 1–2.3 μm. Cystidia are absent; fusoid cystidioles are present, hyaline, thin-walled, and 12.5–14 × 4.8–6 μm. Basidia are clavate, with four sterigmata and a basal clamp connection, 12.5–17.5 × 6.3–9 μm; basidioles are dominant and in shape similar to basidia, but slightly smaller.
Spores: Basidiospores are broadly ellipsoid, truncate or not, hyaline, thick-walled, smooth, weakly dextrinoid, CB+, (4–)4.2–4.8 (–5) × (3–)3.2–3.8(–4) μm, L = 4.48 μm, W = 3.51 μm, Q = 1.27–1.3 (n = 90/3).
Notes: Niveoporia subrusseimarginata is characterized by resupinate to pileate basidiocarps with white to cream pore surface, cinnamon to rust sterile margin, dimitic hyphal system with weakly dextrinoid skeletal hyphae, the presence of cystidioles, and broadly ellipsoid, truncate or not basidiospores.
Additional specimens (paratypes) examined: CHINA. Yunnan, Binchuan County, Jizushan Park, on the stump of Quercus, 14 September 2018, Cui 16973 (BJFC), on fallen angiosperm branch, 14 September 2018, Cui 16980 (BJFC), on angiosperm stump, 14 September 2018, Cui 16988, 16990 (BJFC).
Key to species of Niveoporia
1. Sterile margin indistinct …………………………………………………………………………………………………………………………………… N. decurrata
1. Sterile margin distinct, reddish brown…………………………………………………………………………………………………………………………………2
2. Pores 5–6 per mm………………………………………………………………………………………………………………………………… N. subrusseimarginata
2. Pores 6–8 per mm …………………………………………………………………………………………………………………………………… N. russeimarginata
Perenniporia Murrill, Mycologia 34(5): 595 (1942).
MycoBank: MB 18204
Type species: Perenniporia medulla-panis (Jacq.) Donk, Persoonia 5(1): 76 (1967).
Basidiocarps are annual to perennial and resupinate. Pore surface is white to cream when fresh, cream to buff when dry; pores are round and small. Subiculum is thin, cream, and corky. Tubes are concolorous with pore surface and corky. Hyphal system is dimitic to trimitic; generative hyphae with clamp connections; skeletal hyphae are non-dextrinoid to dextrinoid or amyloid, cyanophilous; tissues are unchanged in KOH. Cystidia are absent, cystidioles are present. Basidiospores are ellipsoid, truncate, hyaline, thick-walled, smooth, dextrinoid, and CB+.
Notes: In our phylogenetic analyses, the species of Perenniporia s. l. clustered into several clades, Perenniporia medulla-panis is grouped with P. substraminea B.K. Cui & C.L. Zhao and P. hainaniana B.K. Cui & C.L. Zhao. These three species share similar morphological characteristics and form the core clade of Perenniporia. The above concept of Perenniporia s. s. is determined from P. medulla-panis, P. hainaniana, and P. substraminea.
Specimens examined: Perenniporia hainaniana. China. Hainan, Changjiang County, Bawangling Nature Reserve, on angiosperm stump, 8 May 2009, Cui 6364 (holotype, BJFC). Perenniporia medulla-panis. China. Guangxi, Jinxiu County, Dayaoshan Nature Reserve, on living angiosperm tree, 15 July 2017, Cui 14515 (BJFC). Perenniporia substraminea. China. Zhejiang, Taishun County, Wuyanling Nature Reserve, on angiosperm stump, 22 August 2011, Cui 10177 (holotype, BJFC).
Key to species of Perenniporia s. s.
1. Pores > 8 per mm…………………………………………………………………………………………………………………………………………P. substraminea
1. Pores < 7 per mm…………………………………………………………………………………………………………………………………………………………2
2. Dendrohyphidia present at dissepimental edges………………………………………………………………………………………………………P. hainaniana
2. Dendrohyphidia absent at dissepimental edges…………………………………………………………………………………………………… P. medulla-panis
Poriella C.L. Zhao, Agronomy 11(7, no. 1308): 5 (2021).
MycoBank: MB 840061
Type species: Poriella subacida (Peck) C.L. Zhao, Agronomy 11(7, no. 1308): 6 (2021).
Basidiocarps are annual to perennial, resupinate to effused-reflexed, and corky when dry. Pore surface is dingy yellowish, cinnamon to ochraceous; pores are round to angular. Context is thin, cream, buff to pale ochraceous. Tubes are concolorous with pore surface, corky. Hyphal system is dimitic to trimitic; generative hyphae with clamp connections; skeletal hyphae is unbranched, strongly dextrinoid, and cyanophilous. Basidiospores are ellipsoid to subglobose, non-truncate, hyaline, thick-walled, smooth, non-dextrinoid to dextrinoid, and CB+.
Notes:
Poriella was newly set up by Chen et al. [
37] according to analyses of ITS, nLSU, mtSSU, and TEF1 datasets. The type species
Poriella subacida, originally described as
Polyporus subacidus Peck [
58], was usually treated in
Perenniporia in current studies [
2,
3,
59]. In our phylogeny, three species of
Perenniporia,
P. africana Ipulet & Ryvarden,
P. ellipsospora Ryvarden & Gilb., and
P. valliculorum Spirin et Zmitr. grouped together with
Poriella subacida and formed a highly supported clade (100% BS, 1.00 BPP,
Figure 1; 98% BS, 1.00 BPP,
Figure 2); morphologically, these three species have resupinate basidiocarps, a dimitic hyphal system with unbranched and strongly dextrinoid skeletal hyphae, and hyaline, thick-walled, and non-truncate basidiospores, which are quite consistent with the concept of
Poriella. Therefore, these three species are transferred to
Poriella based on molecular data and morphological characters.
Poriella africana (Ipulet & Ryvarden) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847384
Basionym: Perenniporia africana Ipulet & Ryvarden, Syn. Fung. 20: 94 (2005).
For a detailed description of
Perenniporia africana, see Ipulet and Ryvarden [
60] and Cui et al. [
3].
Notes:
Poriella africana was originally described from Uganda as
Perenniporia africana [
60]. It has perennial and resupinate basidiocarps, a dimitic hyphal system with unbranched and strongly dextrinoid skeletal hyphae, and subglobose to broadly ellipsoid and non-truncate basidiospores.
Specimens examined: CHINA. Anhui, She County, Qingliangfeng Nature Reserve, on fallen angiosperm trunk, 14 December 2009, Cui 8674, 8676 (BJFC).
Poriella ellipsospora (Ryvarden & Gilb.) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847385
Basionym: Perenniporia ellipsospora Ryvarden & Gilb., Mycotaxon 19: 140 (1984).
For a detailed description of
Perenniporia ellipsospora, see Cui et al. [
3].
Notes:
Poriella ellipsospora and
P. subacida both have resupinate basidiocarps, strongly dextrinoid skeletal hyphae, and ellipsoid and non-truncate basidiospores, but the former has larger pores (3–4 per mm) and dextrinoid basidiospores, and the latter has smaller pores (4–6 per mm) and non-dextrinoid basidiospores [
37].
Specimens examined: CHINA. Yunnan, Lanping County, Changyanshan Nature Reserve, on fallen angiosperm trunk, 18 September 2011, Cui 10276, 10284 (BJFC).
Poriella valliculorum (Spirin & Zmitr.) B.K. Cui & Xing Ji, comb. nov.
Mycobank: MB 847386
Basionym: Perenniporia valliculorum Spirin & Zmitr., Folia Cryptogamica Petropolitana (Sankt-Peterburg) 6: 51 (2005).
For a detailed description of
Perenniporia valliculorum, see Spirin et al. [
9].
Notes:
Poriella valliculorum was originally described from Russia as
Perenniporia valliculorum by Spirin et al. [
9]. It has a pale citric yellow to pale tan pore surface, strongly dextrinoid skeletal hyphae, and non-truncate basidiospores, and these characters fit
Poriella well. In addition, the sequence of
P. valliculorum from the type specimen fell into
Poriella in our phylogeny (
Figure 1 and
Figure 2). Therefore,
Perenniporia valliculorum is transferred to
Poriella.
Key to species of Poriella
1. Basidiospores non-dextrinoid ……………………………………………………………………………………………………………………………… P. subacida
1. Basidiospores dextrinoid…………………………………………………………………………………………………………………………………………………2
2. Pores < 5 per mm ………………………………………………………………………………………………………………………………………… P. ellipsospora
2. Pores > 5 per mm………………………………………………………………………………………………………………………………………………………… 3
3. Basidiocarps perennial…………………………………………………………………………………………………………………………………………P. africana
3. Basidiocarps annual to perennial ……………………………………………………………………………………………………………………… P. valliculorum
Rhizoperenniporia B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847357
Differs from other genera by its resupinate basidiocarps with rhizomorphs, a dimitic hyphal system with weakly dextrinoid skeletal hyphae, and hyaline, thick-walled, ellipsoid, truncate, dextrinoid, and cyanophilous basidiospores.
Type species: Rhizoperenniporia japonica (Yasuda) B.K. Cui & Xing Ji
Etymology: Rhizoperenniporia (Lat.) refers to resembling Perenniporia but with rhizomorphs.
Basidiocarps are annual to perennial, resupinate, and corky when dry. Rhizomorphs are present. Pore surface is grayish white to pale buff when dry; pores are round; dissepiments thick, entire. Subiculum is thin, cream. Tubes are concolorous with pore surface and corky. Hyphal system is dimitic; generative hyphae with clamp connections; skeletal hyphae are dextrinoid and CB+; tissues are unchanged in KOH. Cystidia are absent; cystidioles are present. Basidiospores are ellipsoid, truncate, hyaline, thick-walled, smooth, dextrinoid, and CB+.
Notes: In our phylogenetic analyses (
Figure 1 and
Figure 2), two specimens of
Perenniporia japonica (Yasuda) T. Hatt. & Ryvarden formed a single clade distant from the
Perenniporia s. s. clade. In addition,
P. japonica has basidiocarps with rhizomorphs, which are different from other species of
Perenniporia s. s. Therefore, the new genus
Rhizoperenniporia is proposed to include
P. japonica.
Rhizoperenniporia japonica (Yasuda) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847387
Basionym: Trametes japonica Yasuda, Bot. Mag., Tokyo 32: 356 (1918).
= Perenniporia japonica (Yasuda) T. Hatt. & Ryvarden, Mycotaxon 50: 36 (1994).
For a detailed description of
Perenniporia japonica, see Núñez and Ryvarden [
2] and Cui et al. [
3].
Notes: In
Perenniporia s. l.,
P. aurantiaca,
P. bambusicola,
P. rhizomorpha,
P. subrhizomorpha, and
P. tibetica also have resupinate basidiocarps with rhizomorphs. However,
P. aurantiaca differs from
P. japonica by an orange pore surface, arboriform vegetative hyphae, and tissues becoming violet in KOH [
4].
Perenniporia bambusicola is distinguished by an orange pore surface turning dark violet to black in KOH, arboriform vegetative hyphae, and growing only on bamboo [
51].
Perenniporia rhizomorpha differs by its non-truncate basidiospores [
10].
Perenniporia subrhizomorpha differs by the absence of cystidioles and larger basidiospores (5.7–6.5 × 4.3–5.5 μm) [
56].
Perenniporia tibetica differs by having larger pores and basidiospores (pores 2–3 per mm, basidiospores 6.7–8.7 × 5.3–6.8 μm) [
15].
Specimens examined: CHINA. Shanxi, Yangcheng County, Manghe Nature Reserve, on rotten wood of Vitex, 25 August 2016, Dai 17035 (BJFC); Huguan County, Baquan Gorge, on fallen trunk of Lonicera, 27 August 2016, Dai 17068, 17080 (BJFC).
Tropicoporia B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847358
Differs from other genera by its dimitic to trimitic hyphal system with usually non-dextrinoid and inamyloid skeletal hyphae, broadly ellipsoid to subglobose, truncate, dextrinoid, and cyanophilous basidiospores.
Type species: Tropicoporia aridula (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji
Etymology: Tropicoporia (Lat.) refers to the distribution of the genus in tropical areas.
Basidiocarps are annual to perennial, mostly resupinate, and pseudopileate to rarely pileate. Pore surface is cream, buff-yellow to grayish orange; pores are round to angular. Hyphal system is dimitic to trimitic; generative hyphae with clamp connections; skeletal hyphae are non-dextrinoid to slightly dextrinoid and CB+. Basidiospores are broadly ellipsoid to subglobose, truncate, hyaline, thick-walled, smooth, dextrinoid, and CB+.
Notes: The phylogenies inferred from the two combined datasets ITS + nLSU and ITS + nLSU + mtSSU + TEF1 + TBB1 showed that
Perenniporia aridula B.K. Cui & C.L. Zhao,
P. vanhulleae Decock & Ryvarden,
P. centrali-africana Decock & Mossebo, and
P. brasiliensis C.R.S. de Lira et al. grouped together and formed a clade distinct from the
Perenniporia s. s. clade, although the branch support was low (
Figure 1 and
Figure 2). Morphologically, the four species usually have non-dextrinoid skeletal hyphae and broadly ellipsoid to subglobose basidiospores, which are different from the species of
Perenniporia s. s.. Thus,
Tropicoporia gen. nov. is proposed to accommodate the four species.
In our study,
Tropicoporia was closely related to
Sparsitubus and then grouped with
Dendroporia with low support (
Figure 1 and
Figure 2). Morphologically,
Sparsitubus differs from
Tropicoporia by its effused-reflexed to pileate basidiocarps, and asperulate and non-truncate basidiospores [
54].
Dendroporia is distinguished from
Tropicoporia by its gray to pale brown pore surface, dextrinoid skeletal hyphae, and non-dextrinoid basidiospores.
Tropicoporia aridula (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847388
Basionym: Perenniporia aridula B.K. Cui & C.L. Zhao, Fungal Diversity 58: 48 (2013)
For a detailed description of
Perenniporia aridula, see Zhao et al. [
19].
Notes:
Tropicoporia aridula was first described in
Perenniporia by Zhao et al. [
19]. It is thus far known only from Southwest China.
Tropicoporia vanhulleae is closely related to
T. aridula in morphology and phylogeny, but the former has smaller basidiospores (5.5–6.0 × 4.5–5.5 µm) [
21].
Specimens examined: CHINA. Yunnan, Yuanjiang County, on fallen angiosperm trunk, 9 June 2011, Dai 12396 (holotype, BJFC), on fallen bamboo, 9 June 2011, Dai 12398 (paratype, BJFC).
Tropicoporia brasiliensis (Lira, A.M.S. Soares, Ryvarden & Gibertoni) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847389
Basionym: Perenniporia brasiliensis Lira, A.M.S. Soares, Ryvarden & Gibertoni, Persoonia 38: 355 (2017).
For a detailed description of
Perenniporia brasiliensis, see Lira et al in Crous et al. [
23].
Notes:
Tropicoporia brasiliensis was originally described from Brazil as
Perenniporia brasiliensis by Lira et al (in Crous et al. [
23]); it has subglobose to globose and small basidiospores (3–4 × 2–4 μm) [
23].
Tropicoporia centrali-africana (Decock & Mossebo) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847390
Basionym: Perenniporia centrali-africana Decock & Mossebo, Systematics and Geography of Plants 71(2): 608 (2002).
For a detailed description of
Perenniporia centrali-africana, see Decock and Mossebo [
61].
Notes:
Tropicoporia centrali-africana was originally described from Cameroon as
Perenniporia centrali-africana [
61]. Lira et al (in Crous et al. [
23]) also reported this species from Brazil. It differs from the other three species by its pileate basidiocarps.
Tropicoporia vanhulleae (Decock & Ryvarden) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847391
Basionym: Perenniporia vanhulleae Decock & Ryvarden, Index Fungorum 234: 1 (2015)
For a detailed description of
Perenniporia vanhulleae, see Decock and Ryvarden [
21].
Notes:
Tropicoporia vanhulleae was originally described from Africa as
Perenniporia vanhulleae by Decock and Ryvarden [
21]. The sequence of
T. vanhulleae from the type specimens fell into
Tropicoporia in our phylogeny. Therefore,
Perenniporia vanhulleae is transferred to
Tropicoporia.
Key to species of Tropicoporia
1. Basidiocarps resupinate to pileate………………………………………………………………………………………………………………… T. centrali-africana
1. Basidiocarps resupinate…………………………………………………………………………………………………………………………………………………2
2. Basidiospores < 5 µm………………………………………………………………………………………………………………………………………T. brasiliensis
2. Basidiospores > 5 µm……………………………………………………………………………………………………………………………………………………3
3. Basidiospores 6–7 μm………………………………………………………………………………………………………………………………………… T. aridula
3. Basidiospores 5.5–6.0 µm………………………………………………………………………………………………………………………………… T. vanhulleae
Truncatoporia B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847359
Differs from other genera by its resupinate to pileate basidiocarps, a dimitic to trimitic hyphal system with dextrinoid and cyanophilous skeletal hyphae, and thick-walled, ellipsoid, truncate, and cyanophilous basidiospores.
Type species: Truncatoporia truncatospora (Lloyd) B.K. Cui & Xing Ji
Etymology: Truncatoporia (Lat.) refers to the truncate basidiospores of the genus.
Basidiocarps are annual to perennial, resupinate to pileate, and corky. Pileal surface is brown to ochraceous. Pore surface is buff to pale yellowish buff upon drying; pores are round to angular; dissepiments thin, entire. Context cream buff to pale brown. Tubes are concolorous with pore surface. Hyphal system is dimitic to trimitic; generative hyphae with clamp connections; skeletal hyphae are dextrinoid and CB+; tissues are unchanged in KOH. Basidiospores are ellipsoid, truncate, hyaline, thick-walled, smooth, dextrinoid or not, and CB+.
Notes: In our phylogenetic analyses, two species previously included in
Perenniporia,
P. pyricola Y.C. Dai & B.K. Cui and
P. truncatospora (Lloyd) Ryvarden, grouped together and formed a well-supported clade (78% BS, 1.00 BPP,
Figure 1; 98% BS, 1.00 BPP,
Figure 2) distinct from the
Perenniporia s. s. clade. Therefore,
Truncatoporia gen. nov. is proposed to accommodate
P. pyricola and
P. truncatospora.
Truncatoporia pyricola (Y.C. Dai & B.K. Cui) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847392
Basionym: Perenniporia pyricola Y.C. Dai & B.K. Cui, Mycosystema 29(6): 815 (2010).
For a detailed description of
Perenniporia pyricola, see Dai [
62].
Notes:
Truncatoporia pyricola was first described in
Perenniporia from Northeast China [
62]. It has a distribution in northern China mainly on
Pyrus and
Prunus. The species has perennial and resupinate basidiocarps with cream to pale cinnamon, a dimitic hyphal system with dextrinoid and cyanophilous skeletal hyphae, thick-walled and truncate, dextrinoid, and cyanophilous basidiospores.
Specimens examined: CHINA. Liaoning, Anshan, Qianshan Park, on living tree of Pyrus, 2 August 2008, Dai 10265 (holotype, BJFC); Tianjin, Ji County, Panshan Mountain, Living tree of Crataegus, 6 August 2015, Dai 15496 (BJFC), Dai 15498 (BJFC).
Truncatoporia truncatospora (Lloyd) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847393
Basionym: Trametes truncatospora Lloyd, Mycol. Writ. 6: 853 (1919).
= Perenniporia truncatospora (Lloyd) Ryvarden, Acta Mycol. Sin. 5: 228 (1986).
For a detailed description of
Perenniporia truncatospora, see Cui et al. [
3].
Notes:
Truncatoporia truncatospora and
T. pyricola both have dextrinoid skeletal hyphae and truncate basidiospores, but the former has pileate basidiocarps, smaller pores (6–8 per mm), and non-dextrinoid basidiospores, and the latter has resupinate basidiocarps, larger pores (3–5 per mm) [
62], and dextrinoid basidiospores.
Specimen examined: CHINA. Sichuan, Ganluo County, Shengli, Gaoqiao Village, on living tree of Quercus, 14 September 2019, Cui 17770 (BJFC).
Key to species of Truncatoporia
1. Basidiocarps resupinate, 3–5 per mm………………………………………………………………………………………………………………………T. pyricola
1. Basidiocarps pileate, 6–8 per mm…………………………………………………………………………………………………………………… T. truncatospora
Vanderbyliella B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847360
Differs from other genera by its pileate basidiocarps with an orange brown pileal surface, a dimitic hyphal system with strongly dextrinoid skeletal hyphae, and hyaline, thick-walled, ellipsoid, non-truncate, and cyanophilous basidiospores.
Type species: Vanderbyliella tianmuensis (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji
Etymology: Vanderbyliella (Lat.) refers to the morphological similarity to Vanderbylia.
Basidiocarps are annual to perennial, pileate, and hard corky to woody hard when dry. Pileal surface is clay-buff, orange-brown to yellowish brown, glabrous, and concentrically sulcate. Pore surface is buff to pale brown when dry; pores are round to angular; dissepiments thin, entire. Context cream to pale brown, corky to hard corky. Tubes are buff to pale brown, hard corky to woody hard. Hyphal system is dimitic; generative hyphae with clamp connections; skeletal hyphae are strongly dextrinoid and CB+; tissues are unchanged in KOH. Cystidia are absent; cystidioles are present. Basidiospores are ellipsoid, non-truncate, hyaline, thick-walled, smooth, dextrinoid or not, and CB+.
Notes: In our phylogenetic studies,
Perenniporia tianmuensis B.K. Cui & C.L. Zhao and an unknown species, grouped together and formed a strongly supported clade (99% BS, 1.00 BPP,
Figure 1; 100% BS, 1.00 BPP,
Figure 2), which was distant from the
Perenniporia s. s. clade. Morphologically, these two species have pileate basidiocarps and non-truncate basidiospores, which are different from species of
Perenniporia s. s., so
Vanderbyliella gen. nov. is proposed.
Vanderbyliella tianmuensis (B.K. Cui & C.L. Zhao) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847394
Basionym: Perenniporia tianmuensis B.K. Cui & C.L. Zhao, Mycoscience 54: 236 (2013).
For a detailed description of
Perenniporia tianmuensis, see Zhao and Cui [
18].
Notes:
Vanderbyliella tianmuensis was first described in
Perenniporia from China by Zhao and Cui [
18]; it is characterized by annual and pileate basidiocarps, a dimitic hyphal system with strongly dextrinoid skeletal hyphae, and thick-walled, ellipsoid, non-truncate, dextrinoid, and cyanophilous basidiospores.
Specimens examined: CHINA. Zhejiang, Lin’an County, Tianmushan Nature Reserve, on the basis of dead angiosperm trees, 10 October 2005, Cui 2648 (holotype, BJFC).
Xanthoperenniporia B.K. Cui & Xing Ji, gen. nov.
MycoBank: MB 847361
Differs from other genera by its resupinate basidiocarps with yellow pore surface, weakly dextrinoid, and cyanophilous skeletal hyphae, hyaline, thick-walled, ellipsoid, truncate, and cyanophilous basidiospores.
Type species: Xanthoperenniporia tenuis (Schwein.) B.K. Cui & Xing Ji
Etymology: Xanthoperenniporia (Lat.) refers to resembling Tropicoporia but with a yellow pore surface.
Basidiocarps are annual to perennial, resupinate to reflexed-effused, and corky when dry. Pore surface is cream to yellow when fresh, buff to yellow; pores are round to angular. Subiculum is thin, cream, buff to pale yellowish brown. Tubes are concolorous with pore surface. Hyphal system is dimitic to trimitic, generative hyphae with clamp connections; skeletal hyphae are dextrinoid or weakly dextrinoid and CB+; tissues are unchanged in KOH. Cystidia are absent, cystidioles are usually present. Basidiospores are ellipsoid, truncate, hyaline, thick-walled, smooth, dextrinoid or not, and CB+.
Notes: In the combined five-gene phylogeny,
Perenniporia maackiae (Bondartsev & Ljub.) Parmasto,
P. punctata Hai J. Li & Jing Si,
P. subcorticola Chao G. Wang & F. Wu and
P. tenuis (Schwein.) Ryvarden clustered together and formed a single clade with good support (74% BS, 0.96 BPP,
Figure 1; 89% BS, 1.00 BPP,
Figure 2), which was distant from the
Perenniporia s. s. clade. Morphologically, these species differ from
Perenniporia s. s. in having a yellow pore surface. Therefore, the new genus
Xanthoperenniporia is set up, and four new combinations are proposed.
Xanthoperenniporia maackiae (Bondartsev & Ljub.) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847395
Basionym: Fomitopsis maackiae Bondartsev & Ljub., Botanicheskie Materialy 15: 103 (1962).
= Perenniporia maackiae (Bondartsev & Ljub.) Parmasto, Ann. Bot. fenn. 32(4): 223 (1995).
For a detailed description of
Perenniporia maackiae, see Cui et al. [
3].
Notes:
Xanthoperenniporia maackiae grows mainly on
Maackia.
Xanthoperenniporia subcorticola is similar to
X. maackiae by sharing resupinate basidiocarps with yellow pore surfaces, similar sized pores, and the presence of cystidioles, but
X. subcorticola has smaller basidiospores (4.2–5 × 3.5–4.2 µm) [
29], and
X. maackiae has larger basidiospores (5.4–6.3 × 3.8–5.0 µm).
Specimens examined: CHINA. Heilongjiang, Yichun, Dailing, Liangshui Nature Reserve, on fallen branch of Maackia, 26 August 2014, Cui 11531 (BJFC). Jilin, Antu County, Changbaishan Nature Reserve, on dead tree of Maackia, 11 September 2014, Dai 14780 (BJFC).
Xanthoperenniporia punctata (Hai J. Li & Jing Si) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847396
Basionym: Perenniporia punctata Hai J. Li & Jing Si, Phytotaxa 360(1): 56 (2018).
For a detailed description of
Perenniporia punctata, see Li et al. [
26].
Notes:
Xanthoperenniporia punctata was recently described in
Perenniporia from China by Li et al. [
26] and is characterized by annual and resupinate basidiocarps with a buff-yellow pore surface, a dimitic hyphal system with slightly dextrinoid skeletal hyphae, broadly ellipsoid to subglobose, and truncate and non-dextrinoid basidiospores.
Specimens examined: CHINA. Hubei, Yichang, Wufeng County, Chaibuxi National Forestry Park, on angiosperm stump, 15 August 2017, Dai 17923 (holotype, BJFC), on rotten wood of Quercus, 14 August 2017, Dai 17916 (paratype, BJFC).
Xanthoperenniporia subcorticola (Chao G. Wang & F. Wu) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847397
Basionym: Perenniporia subcorticola Chao G. Wang & F. Wu, MycoKeys 69: 62 (2020).
For a detailed description of
Perenniporia subcorticola, see Wang et al. [
29].
Notes:
Xanthoperenniporia subcorticola was recently described in
Perenniporia as
P. subcorticola Chao G. Wang & F. Wu by Wang et al. [
29]. It is similar to
Perenniporia corticola by having a yellow pore surface, dimitic hyphal system, and truncate and dextrinoid basidiospores of almost the same size, but
P. corticola differs from
P. subcorticola by having arboriform skeletal hyphae and dendrohyphidia [
53].
Specimens examined: CHINA. Fujian, Wuyishan County, Wuyishan Nature Reserve, on rotten wood of Pinus, 21 October 2005, Dai 7330 (holotype, BJFC).
Xanthoperenniporia tenuis (Schwein.) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847398
Basionym: Polyporus tenuis Schwein., Trans. Am. phil. Soc., New Series 4: 159 (1832).
= Perenniporia tenuis (Schwein.) Ryvarden, Norw. J Bot. 20: 9 (1973).
For a detailed description of
Perenniporia tenuis, see Cui et al. [
3].
Notes:
Xanthoperenniporia punctata is similar to
X. tenuis in having resupinate basidiocarps, pale yellow to buff-yellow pore surface, and similar sized basidiospores, but the former differs in its smaller pores (6–9 per mm), and the absence of cystidioles and non-dextrinoid basidiospores [
26], the latter has larger pores (4–6 per mm), the presence of cystidioles, and dextrinoid basidiospores.
Specimens examined: CHINA. Hubei, Yichang, Wufeng County, Chaibuxi National Forestry Park, on rotten angiosperm stump, 15 August 2017, Dai 17935 (BJFC). Shanxi, Yangcheng County, Manghe Nature Reserve, on fallen trunk of Vitex, 25 August 2016, Dai 17026 (BJFC).
Key to species of Xanthoperenniporia
1. Basidiospores non-dextrinoid .……………………………………………………………………………………………………………………………… X. punctata
1. Basidiospores dextrinoid …………………………………………………………………………………………………………………………………………………2
2. Basidiocarps resupinate to reflexed-effused, growing on Maackia……………………………………………………………………………………… X. maackiae
2. Basidiocarps resupinate, growing on other trees………………………………………………………………………………………………………………………3
3. Basidiocarps annual, pores 4–6 per mm ……………………………………………………………………………………………………………………… X. tenuis
3. Basidiocarps perennial, pores 7–8 per mm………………………………………………………………………………………………………………X. subcorticola
Yuchengia B.K. Cui & K.T. Steffen, Nordic J. Bot. 31(3): 333 (2013).
MycoBank: MB 563490
Type species: Yuchengia narymica (Pilát) B.K. Cui, C.L. Zhao & K.T. Steffen, Nordic J. Bot. 31(3): 333 (2013).
Basidiocarps are annual to perennial, resupinate, corky when fresh, and hard corky when dry. Pore surface is cream, yellowish buff to tan; pores are round to angular, dissepiments thin, entire. Subiculum is cream to pale ochraceous. Tubes are concolorous with pore surface. Hyphal system is dimitic; generative hyphae with clamp connections; skeletal hyphae amyloid or not, acyanophilous or weakly cyanophilous, dissolving in KOH. Cystidia are absent; cystidioles are present. Basidiospores are ellipsoid, truncate or not, hyaline, thick-walled, smooth, IKI−, and CB+.
Notes: The type species
Yuchengia narymica was first described as
Trametes narymica Pilát [
63] and later was transferred to
Perenniporia by Pouzar [
64]. Zhao et al. [
35] proposed
Yuchengia to accommodate
Perenniporia narymica based on molecular data and morphological characteristics.
Yuchengia kilemariensis (Spirin & Shirokov) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847399
Basionym: Perenniporia kilemariensis Spirin & Shirokov, Folia Cryptogamica Petropolitana (Sankt-Peterburg) 6: 38 (2005).
For a detailed description of
Perenniporia kilemariensis, see Spirin et al. [
9].
Notes:
Yuchengia kilemariensis was originally described from Russia as
Perenniporia kilemariensis by Spirin et al. [
9]. It has inamyloid skeletal hyphae and truncate basidiospores, which is different from the original descriptions of
Yuchengia. However, it has resupinate basidiocarps, a dimitic hyphal system with skeletal hyphae dissolving in KOH, and ellipsoid and non-dextrinoid basidiospores. These characters fit
Yuchengia well. Moreover, in ITS + nLSU and five-gene phylogenetic analysis, the sequence of
Y. kilemariensis from the type specimen fell into
Yuchengia (
Figure 1 and
Figure 2). Therefore,
P. kilemariensis is transferred to
Yuchengia.
Key to species of Yuchengia
Microporellus subadustus (Z.S. Bi & G.Y. Zheng) B.K. Cui & Xing Ji, comb. nov.
MycoBank: MB 847400
Basionym: Wrightoporia subadusta Z.S. Bi & G.Y. Zheng, Bull. Bot. Res., Harbin 7(4): 76 (1987).
= Perenniporia subadusta (Z.S. Bi & G.Y. Zheng) Y.C. Dai, Ann. Bot. fenn. 39(3): 180 (2002).
= Murinicarpus subadustus (Z.S. Bi & G.Y. Zheng) B.K. Cui & Y.C. Dai, Fungal Di-versity 97: 255 (2019).
= Perenniporia cystidiata Y.C. Dai, W.N. Chou & Sheng H. Wu, Mycotaxon 83: 209 (2002).
Notes: After studying type materials of
Wrightoporia subadusta and
Perennipori cystidiata, Cui et al. [
3] found that the two represent the same species and separated this species from
Perenniporia and proposed
Murinicarpus.
Murinicarpus has the same characters with
Microporellus Murrill, stipitate basidiocarps, dextrinoid skeletal hyphae, non-truncate and non-dextrinoid basidiospores [
33], and it is treated as a synonym of
Microporellus in the current study.
Key to species of Perenniporia and related genera
1. Basidiocarps stipitate………………………………………………………………………………………………………………………………………Microporellus
1. Basidiocarps resupinate, effused-reflexed to pileate…………………………………………………………………………………………………………………2
2. Basidiospores pale yellowish …………………………………………………………………………………………………………………………… Abundisporus
2. Basidiospores hyaline…………………………………………………………………………………………………………………………………………………… 3
3. Skeletal hyphae amyloid…………………………………………………………………………………………………………………………………………………4
3. Skeletal hyphae inamyloid………………………………………………………………………………………………………………………………………………6
4. Basidiocarps pileate……………………………………………………………………………………………………………………………………………Minoporus
4. Basidiocarps resupinate to effused-reflexed ………………………………………………………………………………………………………………………… 5
5. Basidiospores amyloid………………………………………………………………………………………………………………………………………Amylosporia
5. Basidiospores inamyloid………………………………………………………………………………………………………………………………………Yuchengia
6. Basidiocarps with rhizomorphs .……………………………………………………………………………………………………………………………………… 7
6. Basidiocarps without rhizomorphs…………………………………………………………………………………………………………………………………… 8
7. Pore surface orange to orange-brown ………………………………………………………………………………………………………………… Aurantioporia
7. Pore surface grayish white to pale buff…………………………………………………………………………………………………………… Rhizoperenniporia
8. Cystidia present………………………………………………………………………………………………………………………………………………………… 9
8. Cystidia absent………………………………………………………………………………………………………………………………………………………… 10
9. Basidiocarps pileate……………………………………………………………………………………………………………………………………Hornodermoporus
9. Basidiocarps resupinate……………………………………………………………………………………………………………………………………Cystidioporia
10. Basidiospores non-truncate………………………………………………………………………………………………………………………………………… 11
10. Basidiospores truncate……………………………………………………………………………………………………………………………………………… 15
11. Basidiocarps pileate ………………………………………………………………………………………………………………………………………………… 12
11. Basidiocarps resupinate .…………………………………………………………………………………………………………………………………………… 13
12. Basidiospores obovoid …………………………………………………………………………………………………………………………………… Vanderbylia
12. Basidiospores ellipsoid ………………………………………………………………………………………………………………………………… Vanderbyliella
13. Basidiocarps annual……………………………………………………………………………………………………………………………………………Neoporia
13. Basidiocarps annual to perennial……………………………………………………………………………………………………………………………………14
14. Hyphal system dimitic………………………………………………………………………………………………………………………………Luteoperenniporia
14. Hyphal system dimitic to trimitic …………………………………………………………………………………………………………………………… Poriella
15. Basidiocarps osseous ………………………………………………………………………………………………………………………………… Perenniporiopsis
15. Basidiocarps corky to woody hard…………………………………………………………………………………………………………………………………16
16. Pore surface white when fresh, usually with reddish brown sterile margin…………………………………………………………………………Niveoporia
16. Pore surface cream, buff, yellowish to cinnamon, without reddish brown sterile margin………………………………………………………………… 17
17. Basidiocarps resupinate to pileate………………………………………………………………………………………………………………………………… 18
17. Basidiocarps resupinate …………………………………………………………………………………………………………………………………………… 19
18. Basidiospores > 9 µm in length ………………………………………………………………………………………………………………………… Truncospora
18. Basidiospores < 9 µm in length…………………………………………………………………………………………………………………………Truncatoporia
19. Tissues brown to black in KOH …………………………………………………………………………………………………………………………………… 20
19. Tissues unchanged in KOH …………………………………………………………………………………………………………………………………………21
20. Dendrohyphidia present at dissepiment edges, basidiospores non-dextrinoid……………………………………………………………………Dendroporia
20. Dendrohyphidia usually absent at dissepiment edges, basidiospores dextrinoid…………………………………………………………………Citrinoporia
21. Pore surface yellow ……………………………………………………………………………………………………………………………… Xanthoperenniporia
21. Pore surface usually cream to cinnamon………………………………………………………………………………………………………………………… 22
22. Basidiospores ≥ 9 µm in length ………………………………………………………………………………………………………………………… Macrosporia
22. Basidiospores ≤ 9 µm in length …………………………………………………………………………………………………………………………………… 23
23. Skeletal hyphae usually non-dextrinoid ……………………………………………………………………………………………………………… Tropicoporia
23. Skeletal hyphae usually dextrinoid ……………………………………………………………………………………………………………………………… 24
24. Basidiospores < 6 µm in length ………………………………………………………………………………………………………………………… Perenniporia
24. Basidiospores > 6 µm in length .………………………………………………………………………………………………………………………… Macroporia










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
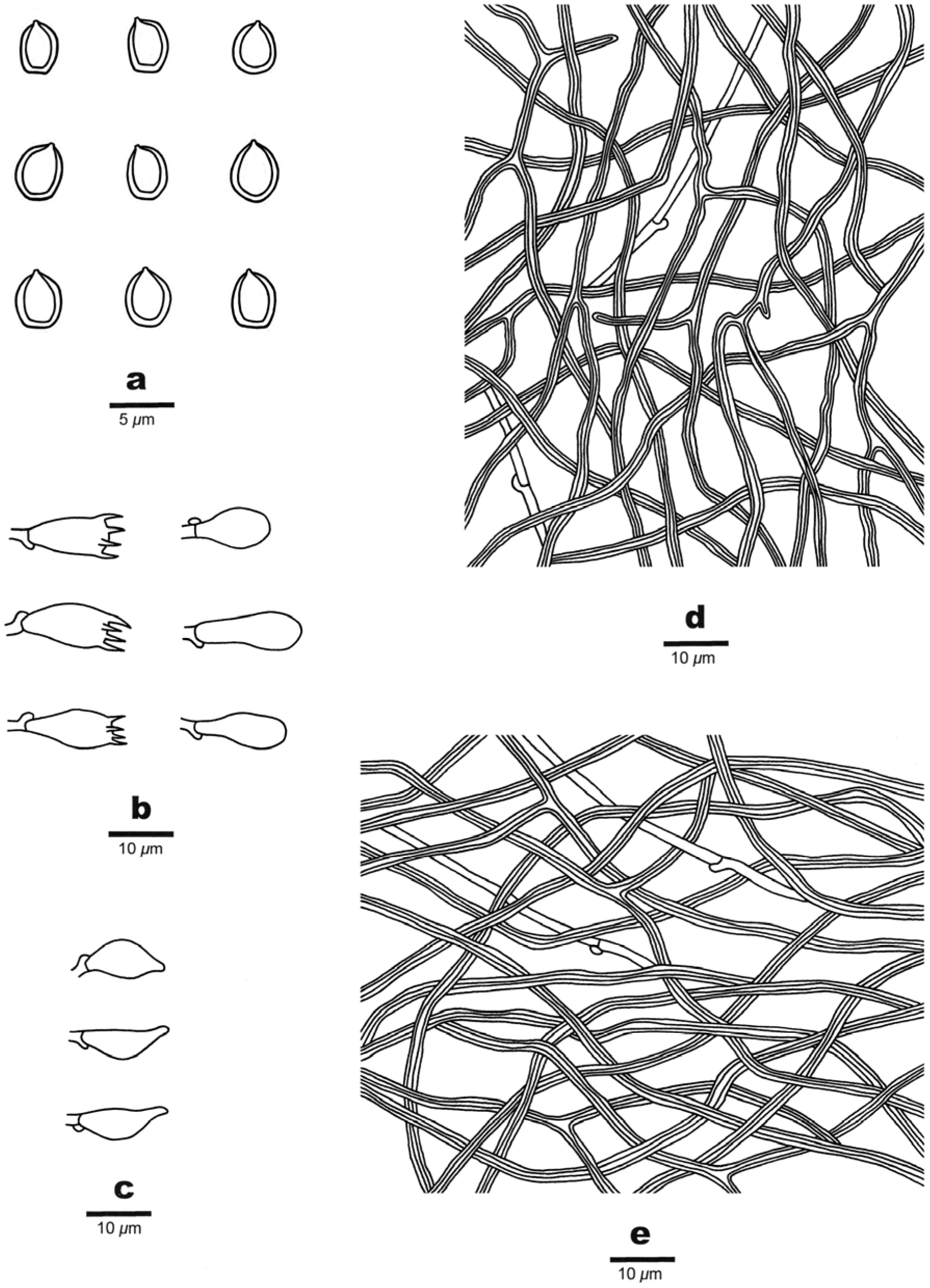{kind=link}