
The Importance of the Epi-Transcriptome in Translation Fidelity


Institute for Integrative Biology of the Cell (I2BC), CEA, CNRS, Université Paris-Saclay, 91198 Gif-sur-Yvette, France
*
Author to whom correspondence should be addressed.
Non-Coding RNA 2021, 7(3), 51; https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7030051
Submission received: 27 July 2021
/
Revised: 17 August 2021
/
Accepted: 22 August 2021
/
Published: 27 August 2021
(This article belongs to the Collection Research on RNA Modification)
{kind=link}
{kind=link}
{kind=link}
Abstract
:RNA modifications play an essential role in determining RNA fate. Recent studies have revealed the effects of such modifications on all steps of RNA metabolism. These modifications range from the addition of simple groups, such as methyl groups, to the addition of highly complex structures, such as sugars. Their consequences for translation fidelity are not always well documented. Unlike the well-known m6A modification, they are thought to have direct effects on either the folding of the molecule or the ability of tRNAs to bind their codons. Here we describe how modifications found in tRNAs anticodon-loop, rRNA, and mRNA can affect translation fidelity, and how approaches based on direct manipulations of the level of RNA modification could potentially be used to modulate translation for the treatment of human genetic diseases.
Keywords:
RNA modifications; ribosomes; tRNA; translation fidelity; m6A; PSI; Inosine; 2′-O-methylation1. Introduction
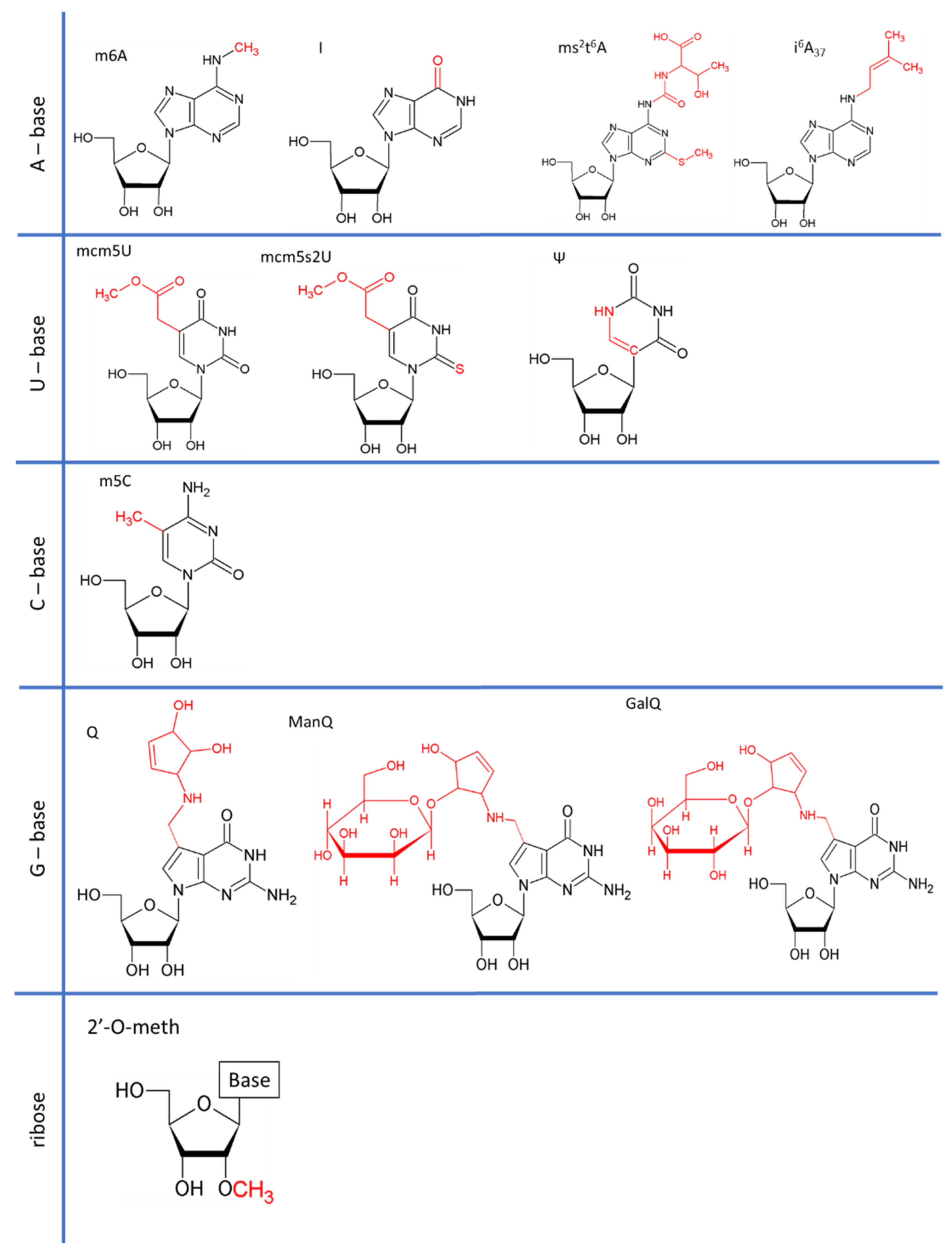
All types of RNA are subject to post-transcriptional modification. Since the discovery of RNA modifications in 1951, more than 150 RNA modifications have been found in coding and non-coding RNAs ranging from the addition of simple groups to the addition of highly complex structures (Figure 1) [1,2,3].
Their biological consequences are largely unknown, but the discovery that RNAs undergo dynamic, reversible chemical modifications marked the birth of the era of epi-transcriptomics. All aspects of RNA metabolism can be affected by RNA modifications [4], either directly, through changes to RNA folding or stability, or indirectly, through the action of “reader” proteins [5,6,7]. Several excellent reviews have already described in detail the role of RNA modifications in cancers and in cell-fate determination [8,9,10,11,12]. In this review, we mainly focus on the importance of RNA modifications in the two most abundant non-coding RNA families (transfer RNAs and ribosomal RNAs) and their consequences for translation fidelity. Translation proceeds via four steps: initiation, elongation, termination and recycling [13]. Generally, it begins with the fixation of the 43S ribosomal complex to the cap-binding protein eIF4F with the help of numerous translation initiation factors (eIFs) [14,15], followed by scanning of the pre-initiation complex (PIC) to the start codon [16]. The two ribosomal subunits are assembled when the PIC is present at the initiation codon by the universally conserved GTPase eIF5B [17,18]. Elongation begins with the delivery of a tRNA to the ribosomal A-site by elongation factors [19] and continues codon-by-codon until the ribosome reaches a stop codon. When a stop codon enters the ribosome, it is recognized by the termination complex eRF1/eRF3, causing the release of the peptide [20,21]. RLI1/ABCE1 splits the ribosome into its two subunits [22,23], which are then available for a new translation cycle [24].
Translation is not perfectly accurate, as it has a median error rate of 0.01% in humans [25,26]. There are 30 codons in the human code that depend on the incorporation of a near-cognate tRNA (pairing of two of the three bases). Codon-anticodon pairing is known to be flexible at the third position of the codon, but it is clear that RNA modifications alter translation accuracy [27]. The incorporation of a near-cognate tRNA can occur during elongation or termination, in which case it is known as readthrough [28].
In this review we will describe modifications affecting two abundant types of non-coding RNAs—tRNAs and rRNAs—and will consider how such modifications fine-tune translational accuracy. We will also discuss the importance of certain mRNA modifications affecting ribosome fidelity. There is a striking difference in the mode of action of modifications between coding and non-coding RNAs, in that modifications to rRNAs and tRNAs act directly on the folding and activity of the molecule, whereas most of the modifications to mRNAs act via reader proteins. This simplified presentation needs to be modulated because some tRNA modifications are required for the proper action of aminoacyl-tRNA synthetases (aaRS) [29].
2. Control of Translation Fidelity by Modifications to Cytosolic tRNAs
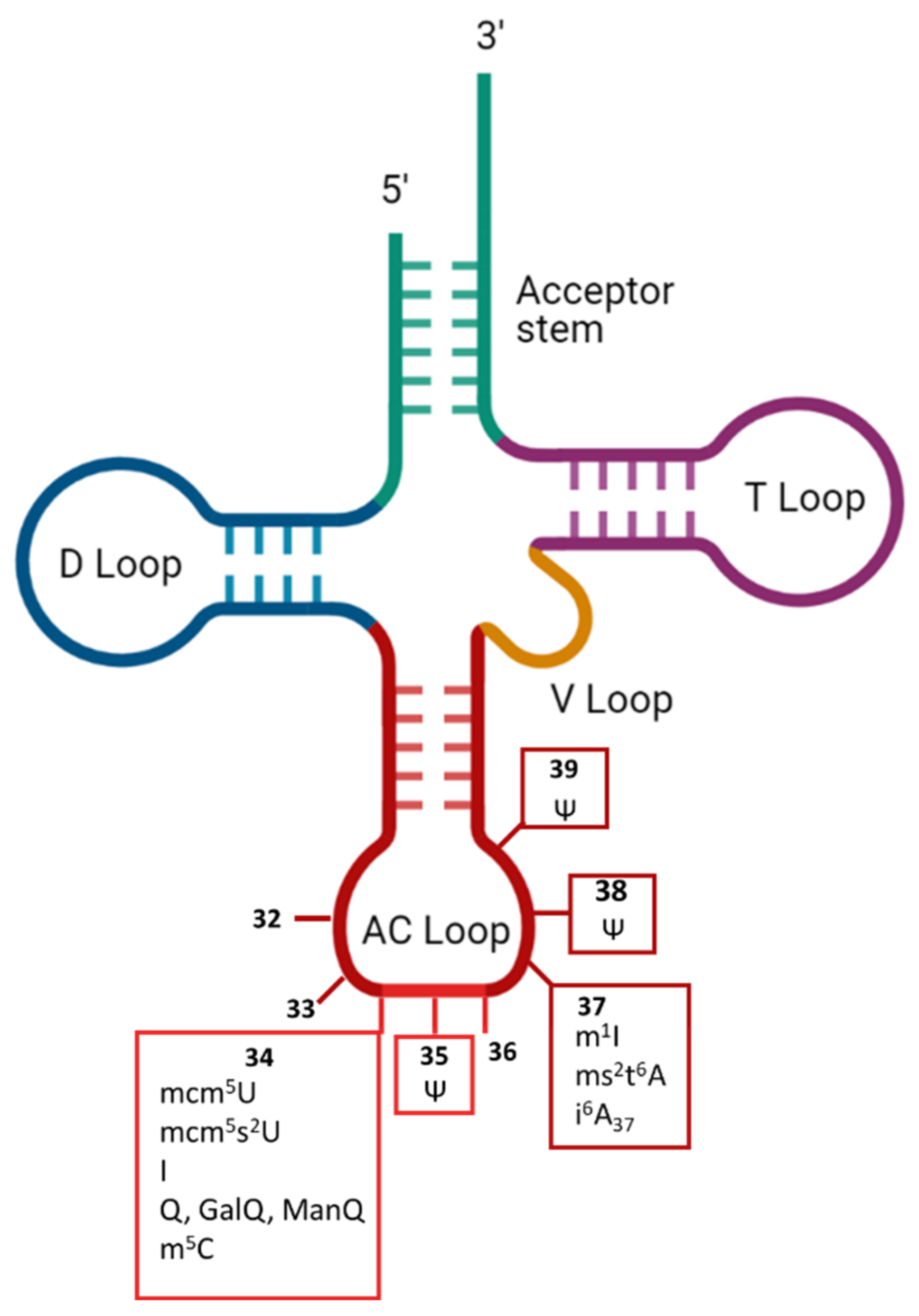
The type of RNA most frequently modified in cells is tRNA, in which about 17% of nucleotides are modified [30]. Modifications have been found in all five domains of tRNA (i.e., acceptor stem, D-loop, T-loop, V-loop and anticodon loop) (Figure 2).
Exceptions exist, but most of the modifications to the D- and T-loops affect the stability or folding of the molecule, whereas those in the anticodon region can affect either the recognition by aaRS or the fidelity of genetic code translation. The anticodon region occupies positions 34-35-36 (Figure 2) that directly base pair to the mRNA codon but are also recognized (together with the acceptor stem) by some aaRS [29]. Some modifications found in the anticodon loop specifically alter the reading frame maintenance. This is the case at position 37, which has been reported to affect the maintenance of the reading frame with the wybutosine yW modification [31]. Interestingly, in humans, no modification has yet been found at position 36, which pairs with the first position in the codon, whereas position 34 of tRNAs, which pairs with the third nucleotide of each codon, is one of the positions at which the various chemical modifications are most numerous [3,32,33]. The reasons for these differences remain unclear, but may reflect the importance of strict base-pairing at certain positions to prevent incorrect amino-acid incorporation, whereas flexibility may be more acceptable at other positions, at which it may not necessarily cause a change of amino acid, thanks to the redundancy of the genetic code. For a long time tRNA modifications have been considered irreversible. However, in 2016, the work of Fange et al. demonstrated that ALKBH1 can remove the methyl group from m1A58 in tRNAs [34], opening the possibility that tRNA modifications would be more dynamic than anticipated. Below, we review modifications of the anticodon loop known to affect the translation of the genetic code by affecting the efficacity of cognate or near-cognate tRNA incorporation.
2.1. mcm5U34 Modifications
The modifications observed at the U34 position of tRNA are 5-methoxycarbonylmethyluridine (mcm5U), 5-methoxycarbonylmethyl-2-thiouridine (mcm5s2U) and their derivatives. Catalysis begins with the addition of 5-methoxycarbonylmethyl (cm5) to uridine by the elongator complex [35,36]. The cm5U is then further modified by the addition of a methyl group by a heterodimeric complex, Trm9-Trm112 (ALKBH8 in mammals) [37]. Finally, the oxygen atom attached to the C2 atom of the uracil ring may undergo thiolation in a subset of tRNAs, catalyzed by the Ncs2-Ncs6 complex and resulting in a final mcm5s2U modification [38]. Ultimately, the U34 position is modified in most eukaryotic tRNAs.
The roles of mcm5U and mcm5s2U in codon–anticodon recognition have been studied in depth in yeast [39,40,41]. The rate of amino-acid misincorporation has been assessed by dual-luciferase reporter assays in a S. cerevisiae TRM9 mutant (absence of mcm5U and mcm5s2U) [40]. The Trm9 deletion decreases the fidelity of translation specifically for the Arg, Gln, Glu and Lys tRNAs. U34 modifications, thus promoting discrimination between some cognate and near-cognate codons. A second study in S. cerevisiae used mass spectrometry to specify amino-acid misincorporation during readthrough, in the presence or absence of U34 modifications. A similar phenotype was observed for tRNAArg under TRM9 gene deletion. However, the results obtained clearly differed from those for tRNAGln, for which U34 modifications of Gln enhance misincorporation. The impact of ALKBH8 protein deficiency has been investigated in mammals [37], through the generation of Alkbh8−/− mice. This mouse line has no mcm5U, mcm5s2U, or mcm5Um modifications to tRNAs, these modifications being replaced by the corresponding acid/amide forms: cm5U and/or ncm5U/ncm5s2U. Hypomodification of the selenocysteine tRNA (tRNASec) impairs its ability to decode the UGA stop codon in vitro. U34 modifications therefore play a role in codon–anticodon recognition in mammals, as in the yeast model. Despite the availability of this mammalian model, studies of the impact of U34 modifications on other tRNAs have yet to be published.
The physiological importance of U34 modifications has been demonstrated for mcm5s2U. This is especially well illustrated by the finding that loss of U34 modifications can lead to ribosome pausing, promotes proteotoxic stress and protein aggregation [42]. Indeed, the presence of the sulfur atom provides an extended chemical group, which stabilizes A-U or G-U pairing [43]. In conclusion, the weak interaction between A-U is strengthened by U34 modification, favoring translation fidelity, whereas stabilization of the unconventional G-U base-pairing favors the incorporation of near-cognate tRNAs.
2.2. I34/37
Inosine (I) results from deamination of the C6 of adenine [44]. Its editing is catalyzed by adenosine deaminases, which act directly on tRNA (ADATs) [45]. Inosine is present at tRNA positions 34 (8 tRNAs) and 37 (tRNAAla) in eukaryotes [45,46,47]. I34 is catalyzed by the heterodimeric enzyme ADAT (hetADAT), consisting of ADAT2 and ADAT3. I37 is deaminated by ADAT1 and further modified by methylation (m1I37) catalyzed by the tRNA methyltransferase TRM3 [48]. A deficiency of I34 has been reported to affect human health, with patients presenting bi-allelic ADAT3 mutations displaying intellectual disability [49].
I34 has been shown to enable tRNAs to pair with U, A and C nucleotides at the wobble position [27]. The chemical origin of the I34 base-pairing effect is the replacement of the hydrogen-donating amino group in the C6 position by a hydrogen-accepting oxygen [44]. Inosine contributes to the extension of the genetic code [50].
Presence of inosine in humans has been linked to a deviation of codon usage between prokaryotes and eukaryotes [51]. Indeed, bioinformatic analysis has shown that TAPSLIVR amino-acid stretches are more efficiently expressed with I34-tRNAs. This has resulted in a greater abundance (eight-fold) and length of such proteins in eukaryotic proteomes than in prokaryotic proteomes, which have only one I34-tRNA.
2.3. Q34 and Its Derivatives
Queuosine (Q), or 7-deazaguanosine, is a modified analog of guanosine incorporated at G34 of GUN anticodon tRNAs [52]. In mammals, Q34 is hypermodified at the Tyr and Asn anticodons, by the addition of a sugar (a galactose (GalQ) and a mannose (ManQ), respectively) to the C4 hydroxy group of the cyclo-pentenediol (Figure 1) [53,54,55]. Eukaryotes cannot synthesize Q de novo, and must therefore obtain it (or its derivative, queuine) as a micronutrient from the gut microbiota or through dietary intake [56,57,58]. Human cells must, therefore, take up queuine, to replace guanosine by queuosine. Interestingly, queuosine levels change during development [59,60]. It remains unclear whether these changes reflect differences in the need for translation fidelity during development or whether they simply reflect the availability of queuine in the diet and/or the possibility of its synthesis by the microbiota. Queuosine has also been associated with cell differentiation, the symptoms of poly-phenyl-ketonurea disease, cancer progression and microbiote diversity. Mannosyltransferase responsible for the generation of ManQ has been isolated from rat liver, but no galactosyltransferase has been identified yet for GalQ [61]. It remains unclear whether these sugar-modified tRNAs are involved in translation (i.e., are they still able to enter the ribosome?) or whether they act as regulatory RNAs, as reported for tRNA-derived fragments [62].
Despite its key position in the anticodon, the role of Q in translation fidelity has been little studied. The impact of Q on codon–anticodon pairing has been physically measured; the C-G pairing is slightly more stable than Q-G, and the Q-U pairing is more than twice as stable as C-U [63]. However, it remains unclear how Q is distinguished from G in front of C or U [64]. Q has been shown to modulate translation accuracy in E. coli [65]. However, a study in eukaryotes investigated the role of Q in tRNAHis decoding in Xenopus laevis oocytes [66]. GUG or QUG—tRNAHis from D. melanogaster was injected into the oocytes, and their ability to decode CAC or CAU codons was assessed. The results underline that Q34-tRNAHis decoded CAU more efficiently than the CAC codon, contrary to what was found for G34-tRNAHis. Additional studies are required to clarify the role of Q and to characterize the role of its hypermodified derivatives, GalQ and ManQ, in translation fidelity.
2.4. m5C34
5-methylcytosine (m5C) is an additional methyl group on C5 present on the cytosine at the position 34 of tRNALeu(CAA). In humans, the methyltransferase hTrm4 is responsible for this methylation position [67], whereas its yeast homolog Trm4 also modifies positions 48 and 49 [68]. In S.cereviae, tRNALeu(CAA) is surnamed “tRNASUP53” for its abilities to suppress the UAG stop codon. Interestingly, the presence of m5C34 on tRNASUP53 depends on the integrity of a 32 bases intron in the pre-tRNA.
The importance of m5C34 in tRNALeu(CAA) mis-incorporation has been assessed in the yeast model [68]. The suppressor activity of tRNASUP53 has been tested on medium minus tryptophan, using a trpl-J(Am) marker. The results showed that in the absence of m5C34 the cells do not grow on the medium, highlighting a decrease in the tRNASUP53 suppressor activity. Thus, m5C34 in tRNALeu(CAA) is considered to act as an enhancer of tRNA misincorporation modification. Unfortunately, for now, no replica of these results exists in a human or another eucaryotic model.
The lack of interest in this modification could be due to the low level of leucine incorporation at the UAG stop codon which has not been reported in any recent study; moreover, hTrm4 has not been linked to any disease in humans.
2.5. ms2t6A37
2-methylthio-N6-threonylcarbamoyladenosine (ms2t6A) biosynthesis is well known: the N6-threonyl carbamoyl adenosine (t6A) is methyl-thiolated to generate the ms2t6A at position 37 of the tRNALys(UUU) which is the only tRNA concerned by this modification in humans. The methyl-thiolation is done by Cdkal1 [69]. There is some structural evidence that ms2t6A at position 37 inhibits a noncanonical U33-A37 interaction and is likely to be needed to compensate for the relatively weak U-turn remodeling properties of mcm5s2U34 [70].
For now, studies of the role of ms2t6A in translation fidelity have been performed only in a bacteria model [71]. In B. subtilis, absence of mcm5s2U34 (ΔyqeV) revealed that ms2t6A37 in tRNALys(UUU) prevents the misreading of its cognate codons AAA and AAG, especially when the rate of translation is high. Unfortunately, ten years after this study, these encouraging data have not yet been reproduced in a eukaryotic model. However, in the same study, a knock-out of Cdkal1 in mouse has been performed and resulted in reduction of glucose-stimulated proinsulin synthesis. Thus, it has been hypothesized that, in the absence of Cdkal1, tRNALys(UUU) misreading increases, limiting the generation of mature insulin and C-peptide which relies on Lys 1 and 2 of proinsulin. This phenotype could explain the molecular pathogenesis of type 2 diabetes in patients carrying Cdkal1 risk alleles, new evidence for the importance of tRNA modifications in human health.
2.6. i6A37
N6-isopentenyladenosine (i6A37) results in the addition of an isopentenyl group onto N6 of A37 by an isopentenyl-transferase (IPTase). The nature of concerned tRNAs varies from one organism to another. In humans, i6A37 has been found to be synthetized by TRIT1 in cytosolic tRNASer (UGA, AGA, and CGA) and tRNA[Ser]Sec [72]. The last is relatively hypomodified (~40%). As a member of near-wobble modification, I6A37 is proposed to enhance A:U stacking by stabilizing the Watson-Crick base pair but the chemical reasons are not yet clear [73].
The role of i6A37 on translation fidelity was assessed in a S. cerevisiae model (Blanchet 2017). Mod5p enzyme synthetizes the i6A37 in the Tyr and Cys tRNAs. The ability of Tyr and Cys tRNAs to readthrough respectively UAA/UAG and UGA stop codons has been assessed in a ΔMod5p strain by mass spectrometry. In this strain, both tRNAs are less efficient at being mis-incorporated, revealing that i6A37 modification acts as an enhancer of translation plasticity in S. cerevisiae. In another study in S. pombe, the role of i6A37 has been assessed using a β-galactosidase codon-swap reporter [74]. This confirms that i6A37 increases the incorporation of tRNATyr at a near-cognate codon. Moreover, it shows that the modification enhances the incorporation of tRNACys at a cognate codon. Altogether, these data suggest that i6A37 promotes decoding activity generally.
To date, no study has been made in humans about the role of i6A37 in translation fidelity. However, it is clear that TRIT1 mutation is associated with severe diseases. Indeed, it has been defined as a tumor suppressor [75]. The mutation of this protein is also associated with encephalopathy and myoclonic epilepsy pathology [76].
2.7. Ψ35/38/39
Pseudouridine (Ψ) is an isomer of uridine in which uracil binds the ribose via a C1′-C5 rather than a C1′-N1 bond [77]. The pseudouridine synthases responsible for the catalysis of tRNA Ψs belong to the PUS RNA-independent family. Ψs are mainly found in the anticodon loop of tRNAs, at positions 35, 38 and 39. Catalysis is performed by two enzymes: PUS7 and PUS3. The effects of Ψ35, Ψ38 and Ψ39 on translation fidelity probably stems from their ability to improve the stacking of double-stranded RNA over that achieved with the uridine isomer [78]. More precisely, the presence of Ψ stabilizes the C3′-endo conformation, creating an additional N1 H-bond donor [79]. Thanks to these chemical characteristics, Ψs involved in (Ψ35) or next to (Ψ38-Ψ39) the anticodon enhances the stability and structure of pairing.
Studies in yeast (S. cerevisiae) have shown that the deletion of PUS3 triggers an increase in misincorporation events relative to the WT strain [39,80]. Even Ψ38 and Ψ39, at the end of the anticodon loop increase miscoding frequency. We have also studied the contribution of Ψ35 to the ability of tRNATyr to act as a near-cognate tRNA for stop-codon readthrough in a ΔPUS7 strain [39]. Mass spectrometry has shown that tyrosine incorporation is less efficient in the absence of the PUS7 gene. This finding highlights the importance of Ψ35 for the ability of tRNATyr to read through UAA and UAG codons. Together, these studies demonstrate that Ψ modifications to the anticodon loop of tRNAs have a major effect on translation fidelity. The tRNA pseudo-uridine synthases PUS3 and PUS7 have both been implicated in human health problems; they cause different diseases, and notably intellectual disability [81,82].
It is interesting to mention that some modifications away from the anticodon loop seems to play a role in translation. For example, absence of ac4C12 and dU20 of Leucine et Serine tRNAs causes a reduction in A-site occupancy at the corresponding codons. Widespread changes in the A-site occupancy have been also observed in the absence of m22G26 [83]. Since these modifications are not in the anticodon loop, they may influence either the charging of these tRNAs or their ability to bind stably the ribosomal A-site. Several studies suggest that interactions inside the anticodon loop are crucial for a correct modification of the tRNAs. These interconnections between modifications create a complex network. For example, in eukaryotes, the yW formation at m1G37 of tRNAPhe is greatly stimulated by the presence of Cm32 and Gm34 [84,85,86]; the presence of i6A37 or t6A37 in tRNASer stimulates the formation of m3C32 [87]; m5C38 is stimulated by prior Q34 formation [88]; and I34 editing in tRNAThr(AGU), is stimulated by prior C to U editing at position 32 [89]. Future structural studies of tRNA-modification enzymes complexes will probably explain such dependencies.
It is also possible that tRNA modifications act through other processes such as RNA sequestration, or the generation of tRNA-derived fragments (tRFs/tiRFs) [90] obtained by the endonuclease cleavage of tRNAs (mainly in the D-Loop, TΨC-Loop and anticodon-Loop), either in normal or stressed conditions. Such tRFs/tiRFs are involved in various physiological and pathological processes by modulating RNA stability or translation [62]. There is a clear link between tRNA modifications and tRFs/tiRFs generation. For example, Dnmt2, which methylates the C38 of tRNAAsp [91], protects tRNA from the degradation [92]. It has been also shown that ALKBH3 activity results in removal of m1A and m3C modifications, leading to a sensitivity to angiogenin cleavage [93].
3. Role of rRNA Modifications in Translation Fidelity
Ribosomal RNA is the most abundant non-coding RNA in the cytoplasm. It is the main constituent of the ribosome. In total, 200 modification sites have been mapped on the human ribosome, in which about 2% of the nucleotides are modified [30,94,95,96]. These modifications can modulate all stages in the life of the rRNA, from ribosome biogenesis to translation accuracy [97]. The most frequent modifications observed are pseudo-uridines and 2′-O-methylations, although base methylation and acetylation have been reported [30]. We focus here on the description of the two main modifications of rRNAs known to affect translation fidelity.
3.1. 2′-O-methylation (Nm)
2′-O-methylation (Nm) is a modification in which a sugar is added to the 2′C hydroxyl group of the nucleotide. The chemical impact of Nm on RNA has been investigated by several studies. It has been reported that Nm biases the sugar pucker equilibrium in favor of the C3′-endo conformation of pyrimidines [98]. Intra-residue steric repulsion occurs between the Nm, the 3′-phosphate, and the 2-carbonyl groups in the C2′-endo conformation, favoring the C3′ form. The Nm modification may, therefore, either stabilize or modulate RNA structures.
In human cells, Nm is mediated by the ribonucleoprotein complex consisting of the methylase fibrillarin (FBL) and the guide RNA (C/D box snoRNA) specific to the methylation site [99]. FBL is an essential protein, but it can be partially inactivated, leading to a decrease of up to 50% in the number of methylation sites in human cells [100,101]. More than 100 2′-O-methylation sites have been mapped on rRNAs, independently of nucleoside identity [102,103].
The role of Nm in miscoding has been explored in human cancer cells [104]. FBL overexpression, leading to hypermethylation of the ribosome, has been shown to trigger an increase in amino-acid incorporation at cognate or near-cognate codons. It is difficult to identify the 2′-O-methylation sites responsible for this phenotype, because site-specific inactivation experiments have not been performed yet on human cells. As FBL methylates all the sites, the only solution would be to inactivate each snoRNA specifically, one-by-one. A study of this type has been performed in yeast, in which knockouts of the various guide C/D box snoRNAs have been performed [105]. The impact of the loss of each snoRNA was evaluated by measuring stop codon readthrough efficiency. Nm-C1639 was identified as the most important of the Nm sites tested. The abolition of Nm at this P-site triggers a slight increase in UAG readthrough. This work revealed a role for Nm-C1639 in the maintenance of ribosome fidelity during termination. There is now a need to reproduce such systematic analyses of Nm sites in humans.
The role of rRNA’s Nm extends beyond miscoding events. The downregulation of FBL has been shown to alter IRES-dependent initiation and frameshifting. A single deletion of Am398 or Gm3745 in the 28S rRNA or of Am163 in the 18S rRNA is embryo-lethal in zebrafish [106]. Moreover, FBL overexpression has been reported during the differentiation of human stem cells, and in several cancer studies, suggesting a central role in these processes [100,104,107,108].
3.2. Pseudouridine
With the exception of position 50 in the 5S rRNA that is catalyzed by the enzyme PUS7, the formation of Ψs in rRNA is catalyzed by a ribonucleoprotein complex composed of the pseudo-uridine synthase DKC1 associated with H/ACA box snoRNAs [109,110]. In human rRNAs, Ψs are mapped with a Ψ/U ratio of 5–7%, with a total of about 100 sites [109,111,112,113,114]. DKC1 is as an essential protein, and mutations of its gene have been linked to X-linked dyskeratosis congenita disease. Patients may display alterations to skin color, nail dystrophy, bone marrow failure, and an increase in the risk of developing cancer and pulmonary fibrosis, although it is not clear whether these effects are related to the absence of Ψ from rRNA [110].
The role of Ψs in miscoding has been investigated in human cells [115]. SNORA24 (ACA24), a H/ACA box snoRNA guiding the Ψ609 and Ψ863 on the 18S rRNA, has been downregulated in HCC cells [116]. An analysis of ribosomal pre-translocation complex dynamics by sm-FRET indicated changes in tRNA conformation in the A-site in ribosomes lacking Ψ609 and Ψ863 relative to wild-type ribosomes, depending on the tRNA entering the ribosome. It has also been shown that lower levels of SNORA24 expression increase amino-acid misincorporation by 10%–20% and readthrough by 15% at UGA, but not at UAG codons.
The way in which Ψs in rRNAs decrease the accuracy of translation seems to depend on their abundance in the peptidyl transferase and decoding centers of the ribosome [77]. Ψs are known to generate an additional N1 H-bond donor and to stabilize the C3′-endo conformation [79]. This enables Ψs to increase RNA–RNA stability in the fidelity centers of the ribosome [117]. A decrease in the number of Ψ sites is, thus, accompanied by ribosome destabilization, resulting in a decrease in ribosome fidelity.
4. mRNA Modifications Influence the Reading of the Genetic Code
Many studies over the last decade have revealed the importance of mRNA modifications. These modifications are highly dynamic, with eraser proteins able to eradicate the modifications from the mRNA. The dynamic aspect of the modifications allows integration in a very efficient manner of the RNA metabolism and translation to the physiological state of the cell, considering the appearance of possible stresses.
4.1. Inosine
The formation of inosine on mRNAs is catalyzed by the adenosine deaminases ADAR1 and 2 [118]. The inosines of mRNAs, like those of tRNAs, play a major role in expansion of the genetic code, with 5072 identified editing sites in human coding sequences [119].
One of the best known examples of the importance of A-to-I editing in mRNA is the modification of the glutamate receptor subunit B (GluRB) precursor messenger RNA: CAG (Gln) → CIG (Arg) in exon 11. This site is modified by ADAR2 and is essential to ensure the impermeability of the glutamate receptor to Ca2+ ions [120]. A defect of this inosine site has, notably, been shown to contribute to neuronal death in amyotrophic lateral sclerosis [121]. The dysregulation of ADAR1 and 2 has also recently been observed in human hepatocellular carcinoma [122]. Patients with an upregulation of ADAR1 and a downregulation of ADAR2 have higher incidences of tumor recurrence and liver cirrhosis, and shorter disease-free survival times. These dysregulations are linked to changes in the number of inosine sites, with, in particular, hyper-editing of the FLNB mRNA and hypo-editing of the COPA mRNA [122]. Finally, ADAR1 seems to act as an oncogene, whereas ADAR2 acts as a tumor suppressor, in hepatocellular carcinoma.
Inosine in mRNAs is known to modulate alternative splicing and stability, but it clearly also plays an essential role as an enhancer of near-cognate tRNA incorporation, ensuring the activity of some proteins [118]. On the other hand, we did not find any significative difference in ribosome profiling between edited and non-edited mRNA in term of translation efficiency in A. Thaliana mitochondria [123]. The conservation of these essential CDS sites, rather than the cognate codon with a G, remains to be evaluated.
4.2. Pseudouridine
Unlike the Ψs found in rRNA, the reaction generating those found in mRNA is catalyzed by pseudo-uridine synthases, which are RNA-independent proteins [113,124,125,126], although the existence of some box H/ACA snoRNAs complementary to mRNAs raises the possibility that RNA-dependent pseudo-urylation of mRNAs also occurs [127,128]. mRNA Ψs are known to be modulated under cellular stress and during development, but no Ψ reader or eraser has yet been described [129]. Within the translated and untranslated regions of mRNAs, pseudo-uridine is present with a Ψ/U ratio of 0.2–0.6%, and 1889 sites have been identified by N3-CMC–enriched pseudo-uridine sequencing [113]. More than 60% of pseudo-uridine residues are located within the coding sequence, suggesting a link with translation [130,131].
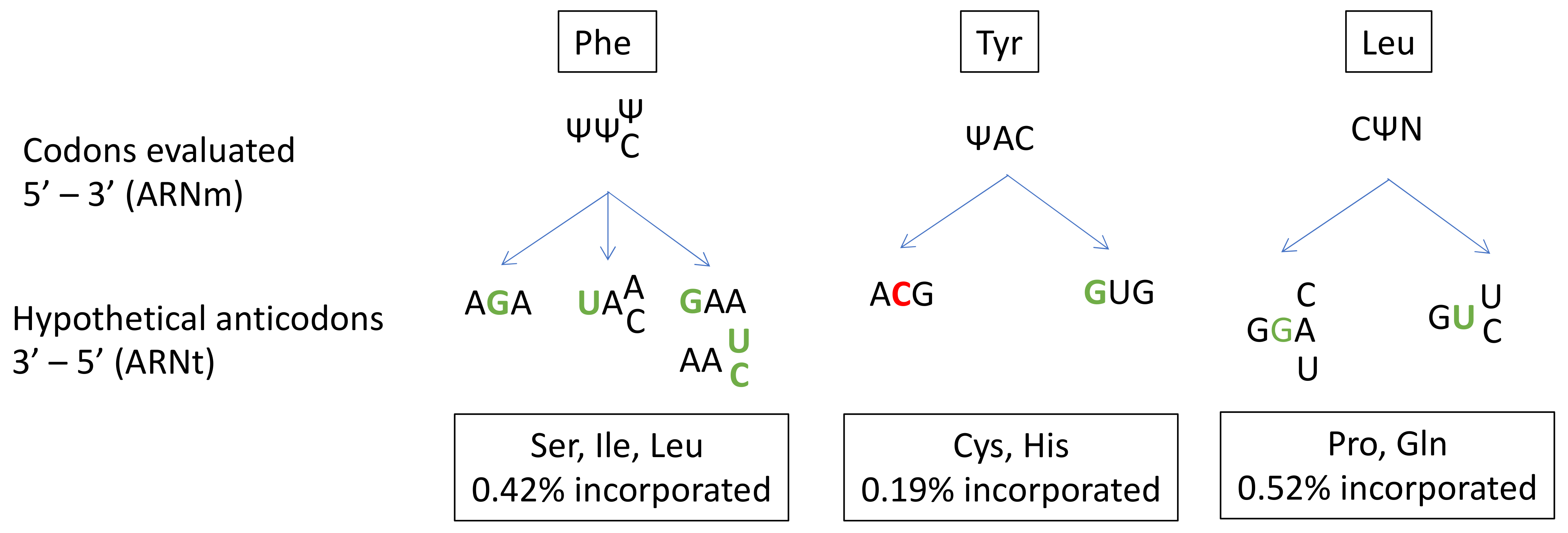
In prokaryotes, several studies have described the ability of Ψ to alter base-pairing and induce misincorporation [132,133,134]. However, far fewer studies have been performed on human cells [131]. Amino-acid misincorporation in front of a “U-codon” has been shown to occur at a rate of 1%. The presence of Ψ in mRNA induces the substitution of Ser, Ile or Leu for Phe at UUU/C codons; Cys or His substitution for Tyr at UAU/C codons; and Pro or Gln substitution for Leu at CUA/U/C/G codons (Figure 3).
Given the high frequency of Ψ in mRNA and its role in near-cognate tRNA recognition, Ψ modifications probably make a major contribution to translation fidelity. A closer look at codon/anticodon base-pairing in the case of the misincorporation of Cys at a Tyr codon reveals a central mismatch between an A and a C. This unfavorable interaction is probably compensated for by the strong ability of Ψ to stabilize the codon/anticodon structure by stacking interactions. Indeed, Ψ is known to enhance RNA structure stability. Despite its ability to form a supplementary N1-hydrogen bond, Ψ has the same Watson-Crick base-pairing properties as U [79].
4.3. m6A
The N6-methyladenosine (m6A) modification involves the addition of a methyl group to the N atom linked to the C6 of adenosine. Chemical predictions of the impact of m6A on RNA–RNA base-pairing suggest a disruption of this interaction due to the methyl group [135]. Indeed, this group must adopt an anti-conformation in the context of A–U pairing. This conformation is less energetically favorable than the syn conformation, leading to destabilization of the RNA–RNA accommodation.
In humans, more than 12000 m6A sites are estimated to be present on 7000 mRNAs [136,137]. About 35% of m6A sites are located within the coding region [138]. m6A is a dynamic modification that has been reported to interact with several enzymes called readers [139,140]. A heterodimeric methylase complex (METTL3-METTL14) is responsible for adding the methyl group. Once modified, the site can be recognized by reader proteins, most of which belong to the YTH-domain protein family (YTHDC and YTHDF), or eraser proteins, which are demethylases (such as FTO and ALKBH5).
The impact of m6A at the first or second position of the codon has been measured by quench flow techniques [141]. This modification delays tRNA incorporation, by slowing tRNA accommodation at site A of the ribosome. However, it has been reported that m6A at the middle position of the codon has a lesser effect on pairing for near-cognate than for cognate tRNAs [142]. This difference in kinetics suggests that tRNA misincorporation rates are likely to be higher in the presence of m6A at the middle position. However, contrary to these findings for prokaryotic systems, mass spectrometry assays in eukaryotes (wheat germ and HEK293T) identified no miscoding effect of the m6A modification [143,144]. The method used for eukaryote systems may be insufficiently sensitive to detect misincorporation in the context of cognate/near-cognate competition. Indeed, the same study found no miscoding effect of Ψ modification, contradicting the findings of another team published in the same year [131]. In the face of these conflicting data, further studies are required to clarify the impact of m6A on miscoding events.
m6A is one of the most commonly studied RNA modifications because of its broad influence on RNA maturation and degradation, RNA-protein interactions and translation efficiency, implicating this modification in a number of different biological processes [138,145,146,147,148,149]. Focusing on human health, altered m6A levels have been implicated in the regulation of the expression of genes relating to cancer pathogenesis and development [150].
4.4. m5C
5-methylcytosine (m5C) is a cytosine with an additional methyl group on C5. Like m6A, m5C is a dynamic modification, with writer, reader and eraser proteins. NSUN2 and the Aly/REF export factor are the principal m5C mRNA writer and reader proteins, respectively [151]. m5C has been mapped on several transcriptomes in humans [152,153,154,155]. Although NSUN2 and NSUN6 are well-known tRNA-modification enzymes, they also appear to modify mRNA. The number of m5C sites in mRNA has been estimated at about a thousand by bisulfite RNA sequencing [156]. Interestingly, viral RNAs are particularly rich in m5C modifications, suggesting that it could play a role in the discrimination of endogenous and exogenous RNAs.
The question of the impact of m5C on translation has been addressed by ribosome profiling in Hues9 human embryonic stem cells with a knockout of NSUN6 gene [157]. No global translational defect was observed, but the absence of NSUN6 was found to trigger stop codon enrichment at the P-site of the ribosome, possibly after readthrough, and an increase in ribosomes bound to the 3′UTR of mRNAs modified by NSUN6. These data suggest that m5C sites in the 3′UTR of mRNA enhance translation termination efficiency by decreasing the readthrough rate. It remains unclear how m5C in the 3′UTR affects termination. Another study in HEK293T cells assessed the impact of m5C at the three codon positions by mass spectrometry [144]. None of the three positions was found to modulate the misincorporation of amino acids.
m5C is linked to human health. Indeed, NSUN2 mutations are associated with growth retardation, neurodevelopmental defects, and have been identified as a possible treatment target for tumors [153,158,159,160,161]. Moreover, the m5C reader and eraser proteins cited above are known to display altered expression levels in various types of cancer [151].
5. Manipulation of RNA Modifications to Treat Human Diseases
The field of RNA modifications is undoubtedly a very promising area in human therapy. Synthetic modified mRNAs can be used in diverse therapeutic contexts, including cardiac regeneration, asthma, cystic fibrosis or lung diseases [162,163,164,165]. The best-known application is probably the current COVID-19 vaccines of Pfizer/BioNtech and Moderna. In these mRNA-based vaccines, all the uridine residues are replaced by N1-methyl-pseudouridines to prevent the recognition of the vaccine mRNA by host RNA sensors and to stimulate translation initiation by attenuating eIF2α phosphorylation [166,167]. For those interested in eiF2α stress response and translational regulations, please see the following review [168]. It is also possible to target mRNAs directly, through the use of artificial snoRNAs to replace a U residue with a Ψ at a specific position [132]. In this example, changing the U to a Ψ at the first position of a premature termination codon leads to the incorporation of several amino acids rather than a stopping of translation. This could restore production of the full-length protein, thereby correcting the genetic defect.
From another standpoint, RNA modifications affect diverse biological processes, and the correct incorporation of many of these modifications, at the correct sites, is required for normal development. Alterations to these modifications have been implicated in several diseases, including cancers and resistance to therapy of melanoma cells [169]. The role of m6A in cancer is very well documented, and m5C has also emerged as a major player in cancer development [170,171,172]. Given the crucial roles of writer, reader and eraser proteins in cell homeostasis, these proteins have naturally emerged as potential treatment targets [173]. Ribosome modifications are also of potential interest in this context, and DKC1 and FBL may serve as potential anticancer targets, as shown by the changes in their levels of expression in many cancers [104,174].
As discussed above, it is possible to target mRNA with an H/ACA snoRNA for the incorporation of a Ψ at a specific position. This approach could be used in genetic diseases caused by the presence of a premature termination codon (PTC). Proof-of-concept has been obtained through the demonstration that replacing the U of the stop codon with Ψ converts the stop codon into a sense codon [132]. Indeed, serine and threonine were found at ΨAA and ΨAG codons, whereas tyrosine and phenylalanine were found at ΨGA codons. In principle, it should be possible to change the modification status of tRNAs to modulate translation fidelity. This would be particularly useful in diseases linked to the appearance of a premature stop codon, which are treated with readthrough-inducing molecules. These molecules, such as aminoglycosides, target the ribosome, enabling it to read through the stop codon, but it should be possible to improve the incorporation of specific tRNAs by altering their modifications [175]. However, in this case, a delicate balance must be found between promoting high levels of readthrough without compromising normal tRNA usage. The recent publication describing the stimulation of UGA readthrough by inhibiting the Cm34 modification on tRNATrp with 2,6-diaminopurine (DAP) paves the way for the development of such therapeutic approaches [176]. We are still at the dawning of the epi-transcriptomic era, particularly as concerns human treatments, but this field promises to yield extraordinary advances.
6. Conclusions
With so many unanswered questions both in terms of molecular mechanisms and physiological consequences, the field of RNA modifications gains more and more interest. One of the current limitations is the difficulty to identify and quantify RNA modifications, especially in highly structured molecules such as tRNAs or rRNAs. Mass spectrometry approaches are extremely accurate, but require highly purified molecules and are hardly quantitative on a large scale. Deep-sequencing (NGS) approaches require either chemical modification of the RNA or immunoprecipitation with a specific antibody, with the associated problems of specificity [177]. Direct RNA sequencing (nanopore) holds a lot of promise with the possibility of directly detecting modified positions. However, this still requires the development of bioinformatics tools to allow a reliable and quantitative analysis. There is no guarantee that all tRNAs will be accessible, although preliminary report exists suggesting that some will be [178].
Author Contributions
Writing the manuscript C.V. and O.N.; Funding acquisition, O.N. All authors have read and agreed to the published version of the manuscript.
Funding
ANR ANR Rescue_ribosome program (17-CE12-0024-01) and ActiMeth (ANR-19-CE12-0004). This work was also supported by the epiRNA program from I2BC.
Institutional Review Board Statement
Not relevant.
Informed Consent Statement
Not relevant.
Data Availability Statement
Not relevant.
Acknowledgments
The English of this manuscript was corrected by Alex Edelman & Associates. We acknowledge support from the ANR ANR Rescue_ribosome program (17-CE12-0024-01) and ActiMeth (ANR-19-CE12-0004). CV is supported by a grant from FRM. This work has also been supported by the I2BC epiRNA program.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cohn, W.E.; Volkin, E. Nucleoside-5′-phosphates from ribonucleic acid. Nature 1951, 167, 483–484. [Google Scholar] [CrossRef]
- Cantara, W.A.; Crain, P.F.; Rozenski, J.; McCloskey, J.A.; Harris, K.A.; Zhang, X.; Vendeix, F.A.P.; Fabris, D.; Agris, P.F. The RNA modification database, RNAMDB: 2011 update. Nucleic Acids Res. 2011, 39, D195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machnicka, M.A.; Milanowska, K.; Oglou, O.O.; Purta, E.; Kurkowska, M.; Olchowik, A.; Januszewski, W.; Kalinowski, S.; Dunin-Horkawicz, S.; Rother, K.M.; et al. MODOMICS: A database of RNA modification pathways - 2013 update. Nucleic Acids Res. 2013, 41, D262. [Google Scholar] [CrossRef]
- Song, J.; Yi, C. Chemical Modifications to RNA: A New Layer of Gene Expression Regulation. ACS Chem. Biol. 2017, 12, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Nazar, R.N. Modification of rRNA as a quality control mechanism in ribosome biogenesis. FEBS Lett. 2002, 523, 182–186. [Google Scholar] [CrossRef] [Green Version]
- Agris, P.F. Decoding the genome: A modified view. Nucleic Acids Res. 2004, 32, 223–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjan, N.; Leidel, S.A. The epitranscriptome in translation regulation: mRNA and tRNA modifications as the two sides of the same coin? FEBS Lett. 2019, 593, 1483–1493. [Google Scholar] [CrossRef] [Green Version]
- Kadumuri, R.V.; Janga, S.C. Epitranscriptomic Code and Its Alterations in Human Disease. Trends Mol. Med. 2018, 24, 886–903. [Google Scholar] [CrossRef]
- Jonkhout, N.; Tran, J.; Smith, M.A.; Schonrock, N.; Mattick, J.S.; Novoa, E.M. The RNA modification landscape in human disease. RNA 2017, 23, 1754–1769. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.J.; Shi, H.; He, C. Epitranscriptomic influences on development and disease. Genome Biol. 2017, 18, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Haran, V.; Lenka, N. Deciphering the Epitranscriptomic Signatures in Cell Fate Determination and Development. Stem Cell Rev. Reports 2019, 15, 474–496. [Google Scholar] [CrossRef]
- Barbieri, I.; Kouzarides, T. Role of RNA modifications in cancer. Nat. Rev. Cancer 2020, 20, 303–322. [Google Scholar] [CrossRef]
- Kapp, L.D.; Lorsch, J.R. The molecular mechanics of eukaryotic translation. Annu. Rev. Biochem. 2004, 73, 657–704. [Google Scholar] [CrossRef]
- Pelletier, J.; Sonenberg, N. The Organizing Principles of Eukaryotic Ribosome Recruitment. Annu. Rev. Biochem. 2019, 88, 307–335. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.; Hellen, C.; Pestova, T. The mechanism of eukaryotic translation initiation and principles of its regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 113–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinnebusch, A. Structural Insights into the Mechanism of Scanning and Start Codon Recognition in Eukaryotic Translation Initiation. Trends Biochem. Sci. 2017, 42, 589–611. [Google Scholar] [CrossRef]
- Hashem, Y.; Frank, J. The Jigsaw Puzzle of mRNA Translation Initiation in Eukaryotes: A Decade of Structures Unraveling the Mechanics of the Process. Annu. Rev. Biophys. 2018, 47, 125–151. [Google Scholar] [CrossRef]
- Pestova, T.; Lomakin, I.; Lee, J.; Choi, S.; Dever, T.; Hellen, C. The joining of ribosomal subunits in eukaryotes requires eIF5B. Nature 2000, 403, 332–335. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, J.; Shin, B.; Kim, J.; Dever, T.; Puglisi, J.; Fernández, I. Structural basis for the transition from translation initiation to elongation by an 80S-eIF5B complex. Nat. Commun. 2020, 11, 1–18. [Google Scholar] [CrossRef]
- Blanchet, S.; Rowe, M.; Von der Haar, T.; Fabret, C.; Demais, S.; Howard, M.; Namy, O. New insights into stop codon recognition by eRF1. Nucleic Acids Res. 2015, 43, 3298–3308. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.; Shao, S.; Murray, J.; Hegde, R.; Ramakrishnan, V. Structural basis for stop codon recognition in eukaryotes. Nature 2015, 524, 493–496. [Google Scholar] [CrossRef]
- Young, D.; Guydosh, N.; Zhang, F.; Hinnebusch, A.; Green, R. Rli1/ABCE1 Recycles Terminating Ribosomes and Controls Translation Reinitiation in 3′UTRs In Vivo. Cell 2015, 162, 872–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dever, T.; Green, R. The elongation, termination, and recycling phases of translation in eukaryotes. Cold Spring Harb. Perspect. Biol. 2012, 4, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Mancera-Martínez, E.; Brito Querido, J.; Valasek, L.; Simonetti, A.; Hashem, Y. ABCE1: A special factor that orchestrates translation at the crossroad between recycling and initiation. RNA Biol. 2017, 14, 1279–1285. [Google Scholar] [CrossRef]
- Loftfield, R.B. The frequency of errors in protein biosynthesis. Biochem. J. 1963, 89, 82–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurland, C.; Gallant, J. Errors of heterologous protein expression. Curr. Opin. Biotechnol. 1996, 7, 489–493. [Google Scholar] [CrossRef]
- Crick, F.H.C. Codon—anticodon pairing: The wobble hypothesis. J. Mol. Biol. 1966, 19, 548–555. [Google Scholar] [CrossRef]
- Namy, O.; Rousset, J.P.; Napthine, S.; Brierley, I. Reprogrammed Genetic Decoding in Cellular Gene Expression. Mol. Cell 2004, 13, 157–168. [Google Scholar] [CrossRef]
- Muramatsu, T.; Nishikawa, K.; Nemoto, F.; Kuchino, Y.; Nishimura, S.; Miyazawa, T.; Yokoyama, S. Codon and amino-acid specificities of a transfer RNA are both converted by a single post-transcriptional modification. Nature 1988, 336, 179–181. [Google Scholar] [CrossRef]
- Sloan, K.E.; Warda, A.S.; Sharma, S.; Entian, K.D.; Lafontaine, D.L.J.; Bohnsack, M.T. Tuning the ribosome: The influence of rRNA modification on eukaryotic ribosome biogenesis and function. RNA Biol. 2017, 14, 1138–1152. [Google Scholar] [CrossRef]
- Tuorto, F.; Lyko, F. Genome recoding by tRNA modifications. Open Biol. 2016, 6, 160287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Crécy-Lagard, V.; Boccaletto, P.; Mangleburg, C.G.; Sharma, P.; Lowe, T.M.; Leidel, S.A.; Bujnicki, J.M. Survey and summary: Matching tRNA modifications in humans to their known and predicted enzymes. Nucleic Acids Res. 2019, 47, 2143–2159. [Google Scholar] [CrossRef] [PubMed]
- Schaffrath, R.; Leidel, S.A. Wobble uridine modifications–a reason to live, a reason to die?! RNA Biol. 2017, 14, 1209–1222. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Clark, W.; Luo, G.; Wang, X.; Fu, Y.; Wei, J.; Wang, X.; Hao, Z.; Dai, Q.; Zheng, G.; et al. ALKBH1-Mediated tRNA Demethylation Regulates Translation. Cell 2016, 167, 816–828.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsborn, T.; Tükenmez, H.; Mahmud, A.K.M.F.; Xu, F.; Xu, H.; Byström, A.S. Elongator, a conserved complex required for wobble uridine modifications in Eukaryotes. RNA Biol. 2014, 11, 1519–1528. [Google Scholar] [CrossRef] [Green Version]
- Bourgeois, G.; Létoquart, J.; van Tran, N.; Graille, M. Trm112, a protein activator of methyltransferases modifying actors of the eukaryotic translational apparatus. Biomolecules 2017, 7, 7. [Google Scholar] [CrossRef]
- Songe-Møller, L.; van den Born, E.; Leihne, V.; Vågbø, C.B.; Kristoffersen, T.; Krokan, H.E.; Kirpekar, F.; Falnes, P.Ø.; Klungland, A. Mammalian ALKBH8 Possesses tRNA Methyltransferase Activity Required for the Biogenesis of Multiple Wobble Uridine Modifications Implicated in Translational Decoding. Mol. Cell. Biol. 2010, 30, 1814–1827. [Google Scholar] [CrossRef] [Green Version]
- Noma, A.; Sakaguchi, Y.; Suzuki, T. Mechanistic characterization of the sulfur-relay system for eukaryotic 2-thiouridine biogenesis at tRNA wobble positions. Nucleic Acids Res. 2009, 37, 1335–1352. [Google Scholar] [CrossRef] [Green Version]
- Blanchet, S.; Cornu, D.; Hatin, I.; Grosjean, H.; Bertin, P.; Namy, O. Deciphering the reading of the genetic code by near-cognate tRNA. Proc. Natl. Acad. Sci. USA 2018, 115, 3018–3023. [Google Scholar] [CrossRef] [Green Version]
- Patil, A.; Chan, C.T.Y.; Dyavaiah, M.; Rooney, J.P.; Dedon, P.C.; Begley, T.J. Translational infidelity-induced protein stress results from a deficiency in Trm9-catalyzed tRNA modifications. RNA Biol. 2012, 9, 990–1001. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.J.O.; Esberg, A.; Huang, B.; Björk, G.R.; Byström, A.S. Eukaryotic Wobble Uridine Modifications Promote a Functionally Redundant Decoding System. Mol. Cell. Biol. 2008, 28, 3301–3312. [Google Scholar] [CrossRef] [Green Version]
- Nedialkova, D.D.; Leidel, S.A. Optimization of Codon Translation Rates via tRNA Modifications Maintains Proteome Integrity. Cell 2015, 161, 1606–1618. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.M.; Gamper, H.; Yang, W. Post-transcriptional modifications to tRNA - A response to the genetic code degeneracy. RNA 2015, 21, 642–644. [Google Scholar] [CrossRef]
- Licht, K.; Hartl, M.; Amman, F.; Anrather, D.; Janisiw, M.P.; Jantsch, M.F. Inosine induces context-dependent recoding and translational stalling. Nucleic Acids Res. 2019, 47, 3–14. [Google Scholar] [CrossRef]
- Rafels-Ybern, À.; Torres, A.G.; Grau-Bove, X.; Ruiz-Trillo, I.; Ribas de Pouplana, L. Codon adaptation to tRNAs with Inosine modification at position 34 is widespread among Eukaryotes and present in two Bacterial phyla. RNA Biol. 2018, 15, 500–507. [Google Scholar] [CrossRef] [Green Version]
- Gerber, A.P.; Keller, W. An adenosine deaminase that generates inosine at the wobble position of tRNAs. Science 1999, 286, 1146–1149. [Google Scholar] [CrossRef]
- Grosjean, H.; Auxilien, S.; Constantinesco, F.; Simon, C.; Corda, Y.; Becker, H.F.; Foiret, D.; Morin, A.; Jin, Y.X.; Fournier, M.; et al. Enzymatic conversion of adenosine to inosine and to N1-methylinosine in transfer RNAs: A review. Biochimie 1996, 78, 488–501. [Google Scholar] [CrossRef]
- Torres, A.G.; Piñeyro, D.; Filonava, L.; Stracker, T.H.; Batlle, E.; Ribas de Pouplana, L. A-to-I editing on tRNAs: Biochemical, biological and evolutionary implications. FEBS Lett. 2014, 588, 4279–4286. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.; Proven, M.; Halvardson, J.; Hagelskamp, F.; Kuchinskaya, E.; Phelan, B.; Bell, R.; Kellner, S.M.; Feuk, L.; Thuresson, A.C.; et al. Identification and rescue of a tRNA wobble inosine deficiency causing intellectual disability disorder. RNA 2020, 26, 1654–1666. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.G.; Rodríguez-Escribà, M.; Marcet-Houben, M.; Santos Vieira, H.G.; Camacho, N.; Catena, H.; Murillo Recio, M.; Rafels-Ybern, À.; Reina, O.; Torres, F.M.; et al. Human tRNAs with inosine 34 are essential to efficiently translate eukarya-specific low-complexity proteins. Nucleic Acids Res. 2021, 49, 7011–7034. [Google Scholar] [CrossRef] [PubMed]
- Rafels-Ybern, À.; Torres, A.G.; Camacho, N.; Herencia-Ropero, A.; Frigolé, H.R.; Wulff, T.F.; Raboteg, M.; Bordons, A.; Grau-Bove, X.; Ruiz-Trillo, I.; et al. The expansion of inosine at the wobble position of tRNAs, and its role in the evolution of proteomes. Mol. Biol. Evol. 2019, 36, 650–662. [Google Scholar] [CrossRef]
- Harada, F.; Nishimura, S. Possible Anticodon Sequences of tRNAHis, tRNAAsn, and tRNAAsp from Escherichia coli B. Universal Presence of Nucleoside Q in the First Position of the Anticondons of These Transfer Ribonucleic Acids. Biochemistry 1972, 11, 301–308. [Google Scholar] [CrossRef]
- Okada, N.; Shindo-okada, N.; Nishimura, S. Isolation of mammalian tRNAAsp and tRNATyr by lectin-sepharose affinity column chromatography. Nucleic Acids Res. 1977, 4, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.; De Barros, J.P.P.; Keith, G.; Baranowski, W.; Desgrès, J. Determination of queuosine derivatives by reverse-phase liquid chromatography for the hypomodification study of Q-bearing tRNAs from various mammal liver cells. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2004, 801, 237–247. [Google Scholar] [CrossRef]
- Nishimura, S. Structure, Biosynthesis, and Function of Queuosine in Transfer RNA; Elsevier: Amsterdam, The Netherlands, 1983. [Google Scholar]
- Farkas, W. Effect of diet on the queuosine family of tRNAs of germ-free mice. J. Biol. Chem. 1980, 255, 6832–6835. [Google Scholar] [CrossRef]
- Reyniers, J.; Pleasants, J.; Wostmann, B.; Katze, J.; Farkas, W. Administration of exogenous queuine is essential for the biosynthesis of the queuosine-containing transfer RNAs in the mouse. J. Biol. Chem. 1981, 256, 11591–11594. [Google Scholar] [CrossRef]
- Katze, J.R.; Gündüz, U.; Smith, D.L.; Cheng, C.S.; McCloskey, J.A. Evidence that the Nucleic Acid Base Queuine is Incorporated Intact into tRNA by Animal Cells. Biochemistry 1984, 23, 1171–1176. [Google Scholar] [CrossRef]
- Zaborske, J.M.; Bauer DuMont, V.L.; Wallace, E.W.J.; Pan, T.; Aquadro, C.F.; Drummond, D.A. A Nutrient-Driven tRNA Modification Alters Translational Fidelity and Genome-wide Protein Coding across an Animal Genus. PLoS Biol. 2014, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, J.R.; Sezzi, E.; Moriyama, E.N.; Gleason, J.M.; Caccone, A. Analysis of a Shift in Codon Usage in Drosophila. In Proceedings of the Journal of Molecular Evolution. J. Mol. Evol. 2003, 57, S214–S225. [Google Scholar] [CrossRef]
- Okada, N.; Nishimura, S. Enzymatic synthesis of Q nucleoside containing mannose in the anticodon of tRNA: Isolation of a novel mannosyltransferase from a cell-free extract of rat liver. Nucleic Acids Res. 1977, 4, 2931–2937. [Google Scholar] [CrossRef] [Green Version]
- Magee, R.; Rigoutsos, I. On the expanding roles of tRNA fragments in modulating cell behavior. Nucleic Acids Res. 2020, 48, 9433–9448. [Google Scholar] [CrossRef] [PubMed]
- Grosjean, H.J.; De Henau, S.; Crotherst, D.M. On the Physical Basis for Ambiguity in Genetic Coding Interactions. Proc. Natl. Acad. Sci. USA 1978, 75, 610–614. [Google Scholar] [CrossRef] [Green Version]
- Morris, R.C.; Brown, K.G.; Elliott, M.S. The effect of queuosine on trna structure and function. J. Biomol. Struct. Dyn. 1999, 16, 757–774. [Google Scholar] [CrossRef] [PubMed]
- Manickam, N.; Joshi, K.; Bhatt, M.J.; Farabaugh, P.J. Effects of tRNA modification on translational accuracy depend on intrinsic codon-anticodon strength. Nucleic Acids Res. 2015, 44, 1871–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, F.; Suter, B.; Grosjean, H.; Keith, G.; Kubli, E. Queuosine modification of the wobble base in tRNAHis influences “in vivo” decoding properties. EMBO J. 1985, 4, 823–827. [Google Scholar] [CrossRef] [PubMed]
- Brzezicha, B.; Schmidt, M.; Makałowska, I.; Jarmołowski, A.; Pieńkowska, J.; Szweykowska-Kulińska, Z. Identification of human tRNA: m5C methyltransferase catalysing intron-dependent m5C formation in the first position of the anticodon of the pre-tRNA(CAA)Leu. Nucleic Acids Res. 2006, 34, 6034–6043. [Google Scholar] [CrossRef] [PubMed]
- Strobel, M.C.; Abelson, J. Effect of intron mutations on processing and function of Saccharomyces cerevisiae SUP53 tRNA in vitro and in vivo. Mol. Cell. Biol. 1986, 6, 2663–2673. [Google Scholar] [CrossRef] [Green Version]
- Arragain, S.; Handelman, S.K.; Forouhar, F.; Wei, F.Y.; Tomizawa, K.; Hunt, J.F.; Douki, T.; Fontecave, M.; Mulliez, E.; Atta, M. Identification of eukaryotic and prokaryotic methylthiotransferase for biosynthesis of 2-methylthio-N6-threonylcarbamoyladenosine in tRNA. J. Biol. Chem. 2010, 285, 28425–28433. [Google Scholar] [CrossRef] [Green Version]
- Durant, P.C.; Baji, A.C.; Sundaram, M.; Kumar, R.K.; Davis, D.R. Structural Effects of Hypermodified Nucleosides in the Escherichia coli and Human tRNALys Anticodon Loop: The Effect of Nucleosides s2U, mcm5U, mcm5s2U, mnm5s2U, t6A, and ms2t6A. Biochemistry 2005, 44, 8078–8089. [Google Scholar] [CrossRef]
- Wei, F.Y.; Suzuki, T.; Watanabe, S.; Kimura, S.; Kaitsuka, T.; Fujimura, A.; Matsui, H.; Atta, M.; Michiue, H.; Fontecave, M.; et al. Deficit of tRNALys modification by Cdkal1 causes the development of type 2 diabetes in mice. J. Clin. Investig. 2011, 121, 3598–3608. [Google Scholar] [CrossRef] [Green Version]
- Lamichhane, T.N.; Mattijssen, S.; Maraia, R.J. Human Cells Have a Limited Set of tRNA Anticodon Loop Substrates of the tRNA Isopentenyltransferase TRIT1 Tumor Suppressor. Mol. Cell. Biol. 2013, 33, 4900–4908. [Google Scholar] [CrossRef] [Green Version]
- U, S.; S, B.; N, F.-V. The modified base isopentenyladenosine and its derivatives in tRNA. RNA Biol. 2017, 14, 1197–1208. [Google Scholar] [CrossRef] [Green Version]
- Lamichhane, T.N.; Blewett, N.H.; Crawford, A.K.; Cherkasova, V.A.; Iben, J.R.; Begley, T.J.; Farabaugh, P.J.; Maraia, R.J. Lack of tRNA Modification Isopentenyl-A37 Alters mRNA Decoding and Causes Metabolic Deficiencies in Fission Yeast. Mol. Cell. Biol. 2013, 33, 2918–2929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinola, M.; Falvella, F.S.; Galvan, A.; Pignatiello, C.; Leoni, V.P.; Pastorino, U.; Paroni, R.; Chen, S.; Skaug, V.; Haugen, A.; et al. Ethnic differences in frequencies of gene polymorphisms in the MYCL1 region and modulation of lung cancer patients’ survival. Lung Cancer 2007, 55, 271–277. [Google Scholar] [CrossRef]
- Yarham, J.W.; Lamichhane, T.N.; Pyle, A.; Mattijssen, S.; Baruffini, E.; Bruni, F.; Donnini, C.; Vassilev, A.; He, L.; Blakely, E.L.; et al. Defective i6A37 Modification of Mitochondrial and Cytosolic tRNAs Results from Pathogenic Mutations in TRIT1 and Its Substrate tRNA. PLoS Genet. 2014, 10, e1004424. [Google Scholar] [CrossRef] [Green Version]
- De Zoysa, M.D.; Yu, Y.T. Posttranscriptional RNA Pseudouridylation. In Enzymes; Academic Press: Cambridge, MA, USA, 2017; Volume 41, pp. 151–167. [Google Scholar]
- Davis, D.R.; Veltri, C.A.; Nielsen, L. An rna model system for investigation of pseudouridine stabilization of the codon-anticodon interaction in trnaLys, tRNAHis and tRNATyr. J. Biomol. Struct. Dyn. 1998, 15, 1121–1132. [Google Scholar] [CrossRef]
- Deb, I.; Popenda, Ł.; Sarzyńska, J.; Małgowska, M.; Lahiri, A.; Gdaniec, Z.; Kierzek, R. Computational and NMR studies of RNA duplexes with an internal pseudouridine-adenosine base pair. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecointe, F.; Namy, O.; Hatin, I.; Simos, G.; Rousset, J.P.; Grosjean, H. Lack of Pseudouridine 38/39 in the anticodon arm of yeast cytoplasmic tRNA decreases in vivo recoding efficiency. J. Biol. Chem. 2002, 277, 30445–30453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Brouwer, A.P.M.; Abou Jamra, R.; Körtel, N.; Soyris, C.; Polla, D.L.; Safra, M.; Zisso, A.; Powell, C.A.; Rebelo-Guiomar, P.; Dinges, N.; et al. Variants in PUS7 Cause Intellectual Disability with Speech Delay, Microcephaly, Short Stature, and Aggressive Behavior. Am. J. Hum. Genet. 2018, 103, 1045–1052. [Google Scholar] [CrossRef] [Green Version]
- Shaheen, R.; Han, L.; Faqeih, E.; Ewida, N.; Alobeid, E.; Phizicky, E.M.; Alkuraya, F.S. A homozygous truncating mutation in PUS3 expands the role of tRNA modification in normal cognition. Hum. Genet. 2016, 135, 707–713. [Google Scholar] [CrossRef] [Green Version]
- Chou, H.J.; Donnard, E.; Gustafsson, H.T.; Garber, M.; Rando, O.J. Transcriptome-wide Analysis of Roles for tRNA Modifications in Translational Regulation. Mol. Cell 2017, 68, 978–992.e4. [Google Scholar] [CrossRef] [Green Version]
- Guy, M.P.; Phizicky, E.M. Conservation of an intricate circuit for crucial modifications of the tRNAPhe anticodon loop in eukaryotes. RNA 2015, 21, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Guy, M.P.; Podyma, B.M.; Preston, M.A.; Shaheen, H.H.; Krivos, K.L.; Limbach, P.A.; Hopper, A.K.; Phizicky, E.M. Yeast Trm7 interacts with distinct proteins for critical modifications of the tRNAPhe anticodon loop. RNA 2012, 18, 1921–1933. [Google Scholar] [CrossRef] [Green Version]
- Guy, M.P.; Shaw, M.; Weiner, C.L.; Hobson, L.; Stark, Z.; Rose, K.; Kalscheuer, V.M.; Gecz, J.; Phizicky, E.M. Defects in tRNA Anticodon Loop 2′-O-Methylation Are Implicated in Nonsyndromic X-Linked Intellectual Disability due to Mutations in FTSJ1. Hum. Mutat. 2015, 36, 1176–1187. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Marcus, E.; D’Silva, S.; Phizicky, E.M.S. cerevisiae Trm140 has two recognition modes for 3-methylcytidine modification of the anticodon loop of tRNA substrates. RNA 2017, 23, 406–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, M.; Hartmann, M.; Schuster, I.; Bender, S.; Thüring, K.L.; Helm, M.; Katze, J.R.; Nellen, W.; Lyko, F.; Ehrenhofer-Murray, A.E. Dynamic modulation of Dnmt2-dependent tRNA methylation by the micronutrient queuine. Nucleic Acids Res. 2015, 43, 10952–10962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio, M.A.T.; Ragone, F.L.; Gaston, K.W.; Ibba, M.; Alfonzo, J.D. C to U editing stimulates A to I editing in the anticodon loop of a cytoplasmic threonyl tRNA in Trypanosoma brucei. J. Biol. Chem. 2006, 281, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.; Han, T.; Li, G. Novel insights into the roles of tRNA-derived small RNAs. RNA Biol. 2021, 1–11. [Google Scholar] [CrossRef]
- Goll, M.G.; Kirpekar, F.; Maggert, K.A.; Yoder, J.A.; Hsieh, C.L.; Zhang, X.; Golic, K.G.; Jacobsen, S.E.; Bestor, T.H. Methylation of tRNAAsp by the DNA methyltransferase homolog Dnmt2. Science 2006, 311, 395–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, M.; Pollex, T.; Hanna, K.; Tuorto, F.; Meusburger, M.; Helm, M.; Lyko, F. RNA methylation by Dnmt2 protects transfer RNAs against stress-induced cleavage. Genes Dev. 2010, 24, 1590–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Qi, M.; Shen, B.; Luo, G.; Wu, Y.; Li, J.; Lu, Z.; Zheng, Z.; Dai, Q.; Wang, H. Transfer RNA demethylase ALKBH3 promotes cancer progression via induction of tRNA-derived small RNAs. Nucleic Acids Res. 2019, 47, 2533–2545. [Google Scholar] [CrossRef] [Green Version]
- Shubina, M.Y.; Musinova, Y.R.; Sheval, E.V. Proliferation, cancer, and aging-novel functions of the nucleolar methyltransferase fibrillarin? Cell Biol. Int. 2018, 42, 1463–1466. [Google Scholar] [CrossRef]
- Natchiar, S.K.; Myasnikov, A.G.; Kratzat, H.; Hazemann, I.; Klaholz, B.P. Visualization of chemical modifications in the human 80S ribosome structure. Nature 2017, 551, 472–477. [Google Scholar] [CrossRef]
- Taoka, M.; Nobe, Y.; Yamaki, Y.; Sato, K.; Ishikawa, H.; Izumikawa, K.; Yamauchi, Y.; Hirota, K.; Nakayama, H.; Takahashi, N.; et al. Landscape of the complete RNA chemical modifications in the human 80S ribosome. Nucleic Acids Res. 2018, 46, 9289–9298. [Google Scholar] [CrossRef]
- Baudin-Baillieu, A.; Namy, O. Saccharomyces cerevisiae, a powerful model for studying rRNA modifications and their effects on translation fidelity. Int. J. Mol. Sci. 2021, 22, 7419. [Google Scholar] [CrossRef]
- Assi, H.A.; Shi, H.; Liu, B.; Clay, M.; Erharter, K.; Kreutz, C.; Holley, C.; Al-Hashimi, H. 2′-O-methylation alters the RNA secondary structural ensemble. bioRxiv 2020, 2020, 121996. [Google Scholar] [CrossRef]
- Watkins, N.J.; Bohnsack, M.T. The box C/D and H/ACA snoRNPs: Key players in the modification, processing and the dynamic folding of ribosomal RNA. Wiley Interdiscip. Rev. RNA 2012, 3, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Erales, J.; Marchand, V.; Panthu, B.; Gillot, S.; Belin, S.; Ghayad, S.E.; Garcia, M.; Laforêts, F.; Marcel, V.; Baudin-Baillieu, A.; et al. Evidence for rRNA 2′-O-methylation plasticity: Control of intrinsic translational capabilities of human ribosomes. Proc. Natl. Acad. Sci. USA 2017, 114, 12934–12939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Marchand, V.; Motorin, Y.; Lafontaine, D.L.J. Identification of sites of 2′-O-methylation vulnerability in human ribosomal RNAs by systematic mapping. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Krogh, N.; Jansson, M.D.; Häfner, S.J.; Tehler, D.; Birkedal, U.; Christensen-Dalsgaard, M.; Lund, A.H.; Nielsen, H. Profiling of 2′-O-Me in human rRNA reveals a subset of fractionally modified positions and provides evidence for ribosome heterogeneity. Nucleic Acids Res. 2016, 44, 7884–7895. [Google Scholar] [CrossRef] [Green Version]
- Incarnato, D.; Anselmi, F.; Morandi, E.; Neri, F.; Maldotti, M.; Rapelli, S.; Parlato, C.; Basile, G.; Oliviero, S. High-throughput single-base resolution mapping of RNA 2-O-methylated residues. Nucleic Acids Res. 2017, 45, 1433–1441. [Google Scholar] [CrossRef] [Green Version]
- Marcel, V.; Ghayad, S.E.; Belin, S.; Therizols, G.; Morel, A.P.; Solano-Gonzàlez, E.; Vendrell, J.A.; Hacot, S.; Mertani, H.C.; Albaret, M.A.; et al. P53 Acts as a Safeguard of Translational Control by Regulating Fibrillarin and rRNA Methylation in Cancer. Cancer Cell 2013, 24, 318–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baudin-baillieu, A.; Fabret, C.; Liang, X.H.; Piekna-Przybylska, D.; Fournier, M.J.; Rousset, J.P. Nucleotide modifications in three functionally important regions of the Saccharomyces cerevisiae ribosome affect translation accuracy. Nucleic Acids Res. 2009, 37, 7665–7677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higa-Nakamine, S.; Suzuki, T.; Uechi, T.; Chakraborty, A.; Nakajima, Y.; Nakamura, M.; Hirano, N.; Suzuki, T.; Kenmochi, N. Loss of ribosomal RNA modification causes developmental defects in zebrafish. Nucleic Acids Res. 2012, 40, 391–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe-Susaki, K.; Takada, H.; Enomoto, K.; Miwata, K.; Ishimine, H.; Intoh, A.; Ohtaka, M.; Nakanishi, M.; Sugino, H.; Asashima, M.; et al. Biosynthesis of Ribosomal RNA in Nucleoli Regulates Pluripotency and Differentiation Ability of Pluripotent Stem Cells. Stem Cells 2014, 32, 3099–3111. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Xu, T.; Ganapathy, S.; Shadfan, M.; Long, M.; Huang, T.H.M.; Thompson, I.; Yuan, Z.M. Elevated snoRNA biogenesis is essential in breast cancer. Oncogene 2014, 33, 1348–1358. [Google Scholar] [CrossRef] [PubMed]
- Schattner, P.; Barberan-Soler, S.; Lowe, T.M. A computational screen for mammalian pseudouridylation guide H/ACA RNAs. RNA 2006, 12, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Jack, K.; Bellodi, C.; Landry, D.M.; Niederer, R.O.; Meskauskas, A.; Musalgaonkar, S.; Kopmar, N.; Krasnykh, O.; Dean, A.M.; Thompson, S.R.; et al. RRNA Pseudouridylation Defects Affect Ribosomal Ligand Binding and Translational Fidelity from Yeast to Human Cells. Mol. Cell 2011, 44, 660–666. [Google Scholar] [CrossRef] [Green Version]
- Hughes, D.G.; Maden, E.H. The pseudouridine contents of the ribosomal ribonucleic acids of three vertebrate species. Numerical correspondence between pseudouridine residues and 2′-O-methyl groups is not always conserved. Biochem. J. 1978, 171, 781–786. [Google Scholar] [CrossRef] [Green Version]
- Maden, B.E.H. The Numerous Modified Nucleotides in Eukaryotic Ribosomal RNA. Prog. Nucleic Acid Res. Mol. Biol. 1990, 39, 241–303. [Google Scholar] [CrossRef]
- Li, X.; Zhu, P.; Ma, S.; Song, J.; Bai, J.; Sun, F.; Yi, C. Chemical pulldown reveals dynamic pseudouridylation of the mammalian transcriptome. Nat. Chem. Biol. 2015, 11, 592–597. [Google Scholar] [CrossRef]
- Bakin, A.V.; Ofengand, J. Mapping of pseudouridine residues in RNA to nucleotide resolution. Methods Mol. Biol. 1998, 77, 297–309. [Google Scholar]
- McMahon, M.; Contreras, A.; Holm, M.; Uechi, T.; Forester, C.M.; Pang, X.; Jackson, C.; Calvert, M.E.; Chen, B.; Quigley, D.A.; et al. A single H/ACA small nucleolar RNA mediates tumor suppression downstream of oncogenic RAS. Elife 2019, 8, e48847. [Google Scholar] [CrossRef] [PubMed]
- Kiss, A.M.; Jády, B.E.; Bertrand, E.; Kiss, T. Human Box H/ACA Pseudouridylation Guide RNA Machinery. Mol. Cell. Biol. 2004, 24, 5797–5807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.H.; Liu, Q.; Fournier, M.J. rRNA Modifications in an Intersubunit Bridge of the Ribosome Strongly Affect Both Ribosome Biogenesis and Activity. Mol. Cell 2007, 28, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Nishikura, K. Functions and regulation of RNA editing by ADAR deaminases. Annu. Rev. Biochem. 2010, 79, 321–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, M.; Yano, T.; Kawabata, H.; Ueda, H.; Suzuki, T. Inosine cyanoethylation identifies A-to-I RNA editing sites in the human transcriptome. Nat. Chem. Biol. 2010, 6, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Sommer, B.; Köhler, M.; Sprengel, R.; Seeburg, P.H. RNA editing in brain controls a determinant of ion flow in glutamate-gated channels. Cell 1991, 67, 11–19. [Google Scholar] [CrossRef]
- Kawahara, Y.; Ito, K.; Sun, H.; Aizawa, H.; Kanazawa, I.; Kwak, S. RNA editing and death of motor neurons: There is a glutamate-receptor defect in patients with amyotrophic lateral sclerosis. Nature 2004, 427, 801. [Google Scholar] [CrossRef]
- Chan, T.H.M.; Lin, C.H.; Qi, L.; Fei, J.; Li, Y.; Yong, K.J.; Liu, M.; Song, Y.; Chow, R.K.K.; Ng, V.H.E.; et al. A disrupted RNA editing balance mediated by ADARs (Adenosine DeAminases that act on RNA) in human hepatocellular carcinoma. Gut 2014, 63, 832–843. [Google Scholar] [CrossRef]
- Planchard, N.; Bertin, P.; Quadrado, M.; Dargel-Graffin, C.; Hatin, I.; Namy, O.; Mireau, H. The translational landscape of Arabidopsis mitochondria. Nucleic Acids Res. 2018, 46, 6218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlile, T.M.; Rojas-Duran, M.F.; Zinshteyn, B.; Shin, H.; Bartoli, K.M.; Gilbert, W.V. Pseudouridine profiling reveals regulated mRNA pseudouridylation in yeast and human cells. Nature 2014, 515, 143–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlile, T.M.; Martinez, N.M.; Schaening, C.; Su, A.; Bell, T.A.; Zinshteyn, B.; Gilbert, W.V. mRNA structure determines modification by pseudouridine synthase 1. Nat. Chem. Biol. 2019, 15, 966–974. [Google Scholar] [CrossRef]
- Safra, M.; Nir, R.; Farouq, D.; Slutzkin, I.V.; Schwartz, S. TRUB1 is the predominant pseudouridine synthase acting on mammalian mRNA via a predictable and conserved code. Genome Res. 2017, 27, 393–406. [Google Scholar] [CrossRef] [Green Version]
- Hüttenhofer, A.; Brosius, J.; Bachellerie, J.P. RNomics: Identification and function of small, non-messenger RNAs. Curr. Opin. Chem. Biol. 2002, 6, 835–843. [Google Scholar] [CrossRef]
- Cavaillé, J.; Buiting, K.; Kiefmann, M.; Lalande, M.; Brannan, C.I.; Horsthemke, B.; Bachellerie, J.P.; Brosius, J.; Hüttenhofer, A. Identification of brain-specific and imprinted small nucleolar RNA genes exhibiting an unusual genomic organization. Proc. Natl. Acad. Sci. USA 2000, 97, 14311–14316. [Google Scholar] [CrossRef] [Green Version]
- Vandivier, L.E.; Gregory, B.D. Reading the Epitranscriptome: New Techniques and Perspectives. In Enzymes; Academic Press: Cambridge, MA, USA, 2017; Volume 41, pp. 269–298. [Google Scholar]
- Gilbert, W.V.; Bell, T.A.; Schaening, C. Messenger RNA modifications: Form, distribution, and function. Science 2016, 352, 1408–1412. [Google Scholar] [CrossRef] [Green Version]
- Eyler, D.E.; Franco, M.K.; Batool, Z.; Wu, M.Z.; Dubuke, M.L.; Dobosz-Bartoszek, M.; Jones, J.D.; Polikanov, Y.S.; Roy, B.; Koutmou, K.S. Pseudouridinylation of mRNA coding sequences alters translation. Proc. Natl. Acad. Sci. USA 2019, 116, 23068–23074. [Google Scholar] [CrossRef]
- Karijolich, J.; Yu, Y.T. Converting nonsense codons into sense codons by targeted pseudouridylation. Nature 2011, 474, 395–399. [Google Scholar] [CrossRef] [Green Version]
- Fernández, I.S.; Ng, C.L.; Kelley, A.C.; Wu, G.; Yu, Y.T.; Ramakrishnan, V. Unusual base pairing during the decoding of a stop codon by the ribosome. Nature 2013, 500, 107–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svidritskiy, E.; Madireddy, R.; Korostelev, A.A. Structural Basis for Translation Termination on a Pseudouridylated Stop Codon. J. Mol. Biol. 2016, 428, 2228–2236. [Google Scholar] [CrossRef] [Green Version]
- Jordan Ontiveros, R.; Stoute, J.; Liu, K.F. The chemical diversity of RNA modifications. Biochem. J. 2019, 476, 1227–1245. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive analysis of mRNA methylation reveals enrichment in 3′ UTRs and near stop codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Dong, L.; Liu, X.M.; Guo, J.; Ma, H.; Shen, B.; Qian, S.B. m6A in mRNA coding regions promotes translation via the RNA helicase-containing YTHDC2. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Meyer, K.D.; Jaffrey, S.R. Rethinking m6A readers, writers, and erasers. Annu. Rev. Cell Dev. Biol. 2017, 33, 319–342. [Google Scholar] [CrossRef] [Green Version]
- Balacco, D.L.; Soller, M. The m6A Writer: Rise of a Machine for Growing Tasks. Biochemistry 2019, 58, 363–378. [Google Scholar] [CrossRef]
- Choi, J.; Ieong, K.W.; Demirci, H.; Chen, J.; Petrov, A.; Prabhakar, A.; O’Leary, S.E.; Dominissini, D.; Rechavi, G.; Soltis, S.M.; et al. N6-methyladenosine in mRNA disrupts tRNA selection and translation-elongation dynamics. Nat. Struct. Mol. Biol. 2016, 23, 110–115. [Google Scholar] [CrossRef] [Green Version]
- Ieong, K.-W.; Indrisiunaite, G.; Prabhakar, A.; Puglisi, J.D.; Ehrenberg, M. N 6-Methyladenosines in mRNAs reduce the accuracy of codon reading by transfer RNAs and peptide release factors. Nucleic Acids Res. 2021, 49, 2684–2699. [Google Scholar] [CrossRef]
- You, C.; Dai, X.; Wang, Y. Position-dependent effects of regioisomeric methylated adenine and guanine ribonucleosides on translation. Nucleic Acids Res. 2017, 45, 9059–9067. [Google Scholar] [CrossRef] [Green Version]
- Hoernes, T.P.; Heimdörfer, D.; Köstner, D.; Faserl, K.; Nußbaumer, F.; Plangger, R.; Kreutz, C.; Lindner, H.; Erlacher, M.D. Eukaryotic translation elongation is modulated by single natural nucleotide derivatives in the coding sequences of mRNAs. Genes 2019, 10, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N 6-methyladenosine-dependent regulation of messenger RNA stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, B.S.; Roundtree, I.A.; Lu, Z.; Han, D.; Ma, H.; Weng, X.; Chen, K.; Shi, H.; He, C. N6-methyladenosine modulates messenger RNA translation efficiency. Cell 2015, 161, 1388–1399. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N6 -methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature 2015, 518, 560–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Chen, K.; Song, B.; Ma, J.; Wu, X.; Xu, Q.; Wei, Z.; Su, J.; Liu, G.; Rong, R.; et al. M6A-Atlas: A comprehensive knowledgebase for unraveling the N6-methyladenosine (m6A) epitranscriptome. Nucleic Acids Res. 2021, 49, D134–D143. [Google Scholar] [CrossRef]
- Meyer, K.D.; Patil, D.P.; Zhou, J.; Zinoviev, A.; Skabkin, M.A.; Elemento, O.; Pestova, T.V.; Qian, S.B.; Jaffrey, S.R. 5′ UTR m6A Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Li, H.; Wu, A.; Peng, Y.; Shu, G.; Yin, G. Functions of N6-methyladenosine and its role in cancer. Mol. Cancer 2019, 18, 1–15. [Google Scholar] [CrossRef] [Green Version]
- García-Vílchez, R.; Sevilla, A.; Blanco, S. Post-transcriptional regulation by cytosine-5 methylation of RNA. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 240–252. [Google Scholar] [CrossRef] [Green Version]
- Squires, J.E.; Patel, H.R.; Nousch, M.; Sibbritt, T.; Humphreys, D.T.; Parker, B.J.; Suter, C.M.; Preiss, T. Widespread occurrence of 5-methylcytosine in human coding and non-coding RNA. Nucleic Acids Res. 2012, 40, 5023–5033. [Google Scholar] [CrossRef]
- Blanco, S.; Dietmann, S.; Flores, J.V.; Hussain, S.; Kutter, C.; Humphreys, P.; Lukk, M.; Lombard, P.; Treps, L.; Popis, M.; et al. Aberrant methylation of t RNA s links cellular stress to neuro-developmental disorders. EMBO J. 2014, 33, 2020–2039. [Google Scholar] [CrossRef]
- Hussain, S.; Sajini, A.A.; Blanco, S.; Dietmann, S.; Lombard, P.; Sugimoto, Y.; Paramor, M.; Gleeson, J.G.; Odom, D.T.; Ule, J.; et al. NSun2-mediated cytosine-5 methylation of vault noncoding RNA determines its processing into regulatory small RNAs. Cell Rep. 2013, 4, 255–261. [Google Scholar] [CrossRef]
- Huang, T.; Chen, W.; Liu, J.; Gu, N.; Zhang, R. Genome-wide identification of mRNA 5-methylcytosine in mammals. Nat. Struct. Mol. Biol. 2019, 26, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Schumann, U.; Zhang, H.N.; Sibbritt, T.; Pan, A.; Horvath, A.; Gross, S.; Clark, S.J.; Yang, L.; Preiss, T. Multiple links between 5-methylcytosine content of mRNA and translation. BMC Biol. 2020, 18, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selmi, T.; Hussain, S.; Dietmann, S.; Heiß, M.; Borland, K.; Flad, S.; Carter, J.M.; Dennison, R.; Huang, Y.L.; Kellner, S.; et al. Sequence- and structure-specific cytosine-5 mRNA methylation by NSUN6. Nucleic Acids Res. 2021, 49, 1006–1022. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Rafiq, M.A.; Noor, A.; Hussain, S.; Flores, J.V.; Rupp, V.; Vincent, A.K.; Malli, R.; Ali, G.; Khan, F.S.; et al. Mutation in NSUN2, which encodes an RNA methyltransferase, causes autosomal-recessive intellectual disability. Am. J. Hum. Genet. 2012, 90, 856–863. [Google Scholar] [CrossRef] [Green Version]
- Abbasi-Moheb, L.; Mertel, S.; Gonsior, M.; Nouri-Vahid, L.; Kahrizi, K.; Cirak, S.; Wieczorek, D.; Motazacker, M.M.; Esmaeeli-Nieh, S.; Cremer, K.; et al. Mutations in NSUN2 cause autosomal- Recessive intellectual disability. Am. J. Hum. Genet. 2012, 90, 847–855. [Google Scholar] [CrossRef] [Green Version]
- Martinez, F.J.; Lee, J.H.; Lee, J.E.; Blanco, S.; Nickerson, E.; Gabrie, S.; Frye, M.; Al-Gazali, L.; Gleeson, J.G. Whole exome sequencing identifies a splicing mutation in NSUN2 as a cause of a Dubowitz-like syndrome. J. Med. Genet. 2012, 49, 380–385. [Google Scholar] [CrossRef] [Green Version]
- Blanco, S.; Bandiera, R.; Popis, M.; Hussain, S.; Lombard, P.; Aleksic, J.; Sajini, A.; Tanna, H.; Cortés-Garrido, R.; Gkatza, N.; et al. Stem cell function and stress response are controlled by protein synthesis. Nature 2016, 534, 335–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, K.; Zangi, L. Modified mRNA as a Therapeutic Tool for the Heart. Cardiovasc. Drugs Ther. 2020, 34, 871–880. [Google Scholar] [CrossRef]
- Mays, L.E.; Ammon-Treiber, S.; Mothes, B.; Alkhaled, M.; Rottenberger, J.; Müller-Hermelink, E.S.; Grimm, M.; Mezger, M.; Beer-Hammer, S.; Von Stebut, E.; et al. Modified Foxp3 mRNA protects against asthma through an IL-10-dependent mechanism. J. Clin. Investig. 2013, 123, 1216–1228. [Google Scholar] [CrossRef]
- Haque, A.K.M.A.; Dewerth, A.; Antony, J.S.; Riethmüller, J.; Schweizer, G.R.; Weinmann, P.; Latifi, N.; Yasar, H.; Pedemonte, N.; Sondo, E.; et al. Chemically modified hCFTR mRNAs recuperate lung function in a mouse model of cystic fibrosis. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sahu, I.; Haque, A.K.M.A.; Weidensee, B.; Weinmann, P.; Kormann, M.S.D. Recent Developments in mRNA-Based Protein Supplementation Therapy to Target Lung Diseases. Mol. Ther. 2019, 27, 803–823. [Google Scholar] [CrossRef] [Green Version]
- Nance, K.D.; Meier, J.L. Modifications in an Emergency: The Role of N1-Methylpseudouridine in COVID-19 Vaccines. ACS Cent. Sci. 2021, 7, 748–756. [Google Scholar] [CrossRef] [PubMed]
- Svitkin, Y.V.; Cheng, Y.M.; Chakraborty, T.; Presnyak, V.; John, M.; Sonenberg, N. N1-methyl-pseudouridine in mRNA enhances translation through eIF2α-dependent and independent mechanisms by increasing ribosome density. Nucleic Acids Res. 2017, 45, 6023–6036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wek, R.C. Role of eIF2α kinases in translational control and adaptation to cellular stress. Cold Spring Harb. Perspect. Biol. 2018, 10, a032870. [Google Scholar] [CrossRef]
- Rapino, F.; Delaunay, S.; Rambow, F.; Zhou, Z.; Tharun, L.; De Tullio, P.; Sin, O.; Shostak, K.; Schmitz, S.; Piepers, J.; et al. Codon-specific translation reprogramming promotes resistance to targeted therapy. Nature 2018, 558, 605–609. [Google Scholar] [CrossRef]
- Dong, S.; Wu, Y.; Liu, Y.; Weng, H.; Huang, H. N6-methyladenosine Steers RNA Metabolism and Regulation in Cancer. Cancer Commun. 2021, cac2.12161. [Google Scholar] [CrossRef]
- Huang, J.; Chen, Z.; Chen, X.; Chen, J.; Cheng, Z.; Wang, Z. The role of RNA N6-methyladenosine methyltransferase in cancers. Mol. Ther. Nucleic Acids 2021, 23, 887–896. [Google Scholar] [CrossRef]
- Chen, X.; Li, A.; Sun, B.F.; Yang, Y.; Han, Y.N.; Yuan, X.; Chen, R.X.; Wei, W.S.; Liu, Y.; Gao, C.C.; et al. 5-methylcytosine promotes pathogenesis of bladder cancer through stabilizing mRNAs. Nat. Cell Biol. 2019, 21, 978–990. [Google Scholar] [CrossRef]
- Nombela, P.; Miguel-López, B.; Blanco, S. The role of m6A, m5C and Ψ RNA modifications in cancer: Novel therapeutic opportunities. Mol. Cancer 2021, 20, 1–30. [Google Scholar] [CrossRef]
- Ruggero, D.; Grisendi, S.; Piazza, F.; Rego, E.; Mari, F.; Rao, P.H.; Cordon-Cardo, C.; Pandolfi, P.P. Dyskeratosis congenita and cancer in mice deficient in ribosomal RNA modification. Science 2003, 299, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Bidou, L.; Bugaud, O.; Belakhov, V.; Baasov, T.; Namy, O. Characterization of new-generation aminoglycoside promoting premature termination codon readthrough in cancer cells. RNA Biol. 2017, 14, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Trzaska, C.; Amand, S.; Bailly, C.; Leroy, C.; Marchand, V.; Duvernois-Berthet, E.; Saliou, J.M.; Benhabiles, H.; Werkmeister, E.; Chassat, T.; et al. 2,6-Diaminopurine as a highly potent corrector of UGA nonsense mutations. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntyre, A.B.R.; Gokhale, N.S.; Cerchietti, L.; Jaffrey, S.R.; Horner, S.M.; Mason, C.E. Limits in the detection of m6A changes using MeRIP/m6A-seq. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, N.; Poodari, V.; Jain, M.; Olsen, H.; Akeson, M.; Abu-Shumays, R. Direct Nanopore Sequencing of Individual Full Length tRNA Strands. bioRxiv 2021. [Google Scholar] [CrossRef]
Figure 1.
Structure of different RNA modifications discussed in this review. In black: basic structure of the base or ribose concerned. Red: chemical modification. m6A: N6-methyladenosine. I: inosine. ms2t6A: 2-methylthio-N6-threonylcarbamoyladenosine. I6A: N6-isopentenyladenosine. mcm5U: 5-methoxycarbonylmethyluridine. mcm5s2U: 5-methoxycarbonylmethyl-2-thiouridine. Ψ: pseudo-uridine. m5C: 5-methylcytosine. Q: queuosine. ManQ: mannosyl-queuosine. GalQ: galactosyl-queuosine. Nm: 2′-O-methylation.
Figure 1.
Structure of different RNA modifications discussed in this review. In black: basic structure of the base or ribose concerned. Red: chemical modification. m6A: N6-methyladenosine. I: inosine. ms2t6A: 2-methylthio-N6-threonylcarbamoyladenosine. I6A: N6-isopentenyladenosine. mcm5U: 5-methoxycarbonylmethyluridine. mcm5s2U: 5-methoxycarbonylmethyl-2-thiouridine. Ψ: pseudo-uridine. m5C: 5-methylcytosine. Q: queuosine. ManQ: mannosyl-queuosine. GalQ: galactosyl-queuosine. Nm: 2′-O-methylation.

Figure 2.
Overview of human cytosolic tRNAS anticodon loop modifications known to shape translation fidelity. AC loop: anticodon loop. Ψ: pseudo-uridine. I: inosine. m1I: N1-methylinosine. mcm5U: 5-methoxycarbonylmethyluridine. mcm5s2U: 5-methoxycarbonylmethyl-2-thiouridine. Q: queuosine. GalQ: galactosyl-queuosine. ManQ: mannosyl-queuosine. ms2t6A: 2-methylthio-N6-threonylcarbamoyladenosine. m5C: 5-methylcytosine. i6A: N6-isopentenyladenosine. 32–39: anticodon loop positions.
Figure 2.
Overview of human cytosolic tRNAS anticodon loop modifications known to shape translation fidelity. AC loop: anticodon loop. Ψ: pseudo-uridine. I: inosine. m1I: N1-methylinosine. mcm5U: 5-methoxycarbonylmethyluridine. mcm5s2U: 5-methoxycarbonylmethyl-2-thiouridine. Q: queuosine. GalQ: galactosyl-queuosine. ManQ: mannosyl-queuosine. ms2t6A: 2-methylthio-N6-threonylcarbamoyladenosine. m5C: 5-methylcytosine. i6A: N6-isopentenyladenosine. 32–39: anticodon loop positions.

Figure 3.
Prediction of anticodon substitution in front of codons with pseudo-uridine (Ψ), based on the amino acid mis-incorporated in the study of Eyler et.al. (2019). Nucleotides in green indicate a mismatch in front of Ψ. Nucleotides in red indicate a mismatch next to Ψ. N: A, U, C or G base. Possibilities of codon-anticodon pairings with more than one mismatch are not represented.
Figure 3.
Prediction of anticodon substitution in front of codons with pseudo-uridine (Ψ), based on the amino acid mis-incorporated in the study of Eyler et.al. (2019). Nucleotides in green indicate a mismatch in front of Ψ. Nucleotides in red indicate a mismatch next to Ψ. N: A, U, C or G base. Possibilities of codon-anticodon pairings with more than one mismatch are not represented.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Valadon, C.; Namy, O. The Importance of the Epi-Transcriptome in Translation Fidelity. Non-Coding RNA 2021, 7, 51. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7030051
AMA Style
Valadon C, Namy O. The Importance of the Epi-Transcriptome in Translation Fidelity. Non-Coding RNA. 2021; 7(3):51. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7030051
Chicago/Turabian StyleValadon, Charlène, and Olivier Namy. 2021. "The Importance of the Epi-Transcriptome in Translation Fidelity" Non-Coding RNA 7, no. 3: 51. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7030051
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.