
Synthetic, Photosynthetic, and Chemical Strategies to Enhance Carbon Dioxide Fixation

1
Department of Natural Sciences, Bowie State University, 14000 Jericho Park Rd., Bowie, MD 20715, USA
2
Independent Researcher, El Paso, TX 79968, USA
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to the work.
‡
Formerly at Department of Chemistry & Biochemistry, University of Texas El Paso, 500 W. University Ave., El Paso, TX 79968, USA.
C 2022, 8(1), 18; https://0-doi-org.brum.beds.ac.uk/10.3390/c8010018
Submission received: 28 December 2021
/
Revised: 9 February 2022
/
Accepted: 18 February 2022
/
Published: 15 March 2022
Abstract
:The present human population is more than three times what it was in 1950. With that, there is an increasing demand for the consumption of fossil fuels for various anthropogenic activities. This consumption is the major source of carbon dioxide emission causing greenhouse effects leading to global warming. The dependency on fossil fuels around the globe is such that it would be hard to move away from it any time soon. Hence, we must work on strategies to improve carbon dioxide fixation as we are making advancements in clean energy technology. This review explores the natural carbon dioxide fixation pathways in plants and various microorganisms and discusses their limitations and alternative strategies. It explains what necessitates the exploration of synthetic pathways and discusses strategies and matrices to consider while evaluating various pathways. This review also discusses the recent breakthroughs in the field of nanosciences that could accelerate chemical methods of carbon dioxide fixation.
Keywords:
rubisco; cyanobacteria; C4 pathway; CAM; acetogens; diatoms; nanoparticles; syngas; dinoflagellates; synthetic pathways1. Introduction
Carbon is the most important element of organic matter, forming the major part of its dry weight. Elemental carbon exists in the atmosphere as carbon dioxide (CO2) and is dissolved in water, rocks, and soils as carbonic acid (H2CO3), bicarbonates, and carbonates [1]. In fossil fuels, it exists as carbon rings or chains and in biomolecules such as nucleic acids, carbohydrates, proteins, and lipids. Atmospheric CO2 is the major cause for global warming through the greenhouse effect; hence, carbon dioxide emissions must be strictly regulated to minimize the greenhouse effect [2,3,4]. The Intergovernmental Panel on Climate Change (IPCC) report from 2018 forewarns us that catastrophe in the form of floods, poverty, drought, etc., will be unleashed by a mere change in global temperature of a degree. In the Paris agreement in February 2021, 186 countries that are responsible for 90% of global emissions pledged carbon emission reduction through 2025 or 2030. India (the third-largest carbon emitter) has committed to generating 50% of its energy needs by renewable means by 2030 and reaching net-zero carbon emissions by 2070. China (the largest carbon emitter) has not made any such commitments, since they are the manufacturing hub and are wary of disrupting the supply chain worldwide. Nevertheless, irrespective of the verbal commitment, China leads the world in electricity production through renewable energy resources and also in the investment in renewable energy innovation and capacity building according to the UN Environmental Program Report.
Though the top three major carbon-emitting countries, i.e., China, the United States, and India, are aggressively working on cutting down their emissions by reducing their dependency on fossil fuels, it will still take a few more decades to slow down before we see any observable effects. This fact necessitates that we explore scientific methods that could reduce emitted carbon via synthetic, photosynthetic, and chemical methods. This review provides a brief overview of the theories related to photosynthetic pathways, explores their potential for enhancing fixation capacity, provides an overview of approaches to identify novel synthetic carbon fixation pathways, and gives a brief overview of non-photosynthetic approaches and speculates on their potential.
2. Engineering Photosynthetic Plants
One of the main contributors to biological carbon fixation is plants, during the process of photosynthesis. Plants are divided into three main categories based on their photosynthetic pathways, i.e., C3, C4, and crassulacean acid metabolism (CAM) [5]. These pathways are differentiated by their respective methods of carbon fixation and their ability to combat photorespiration. Photorespiration is the process by which the enzyme RuBisCo reacts with oxygen when there is a lack of CO2, wasting necessary energy and creating more CO2 [6,7]. C3 plants account for approximately 85% of the plant population. Characterized by its production of a three-carbon compound, 3-Phosphoglycerate, C3 photosynthesis does not have a mechanism capable of reducing the occurrence of photorespiration [5].
The C3 cycle, also known as the Calvin–Benson–Bassham cycle, has been determined to be the least efficient of the group. This is because in other pathways such as the reductive tricarboxylic acid cycles, the consumption of ATP is reserved only for inauspicious reactions that ultimately expedite carbon fixation such as RuBP-based carboxylation and carboxyl reduction. Additionally, to better conserve cellular resources, most of these unfavorable reactions are coupled with exergonic reactions, which, due to their spontaneous nature, diminish the need for the utilization of ATP [7]. Conversely, the Calvin cycle has no such functionality and frivolously uses ATP for these processes that have been optimized in other pathways.
C4 and CAM plants differ from C3 in their internal mechanisms to combat the occurrence of photorespiration. C4 plants are the least prevalent, accounting for approximately 5% of the plant population. In C4 plants, photosynthesis occurs in two different types of cells, mesophyll cells and bundle sheath cells. During C4 photosynthesis, CO2 reacts with the enzyme phosphoenolpyruvate carboxylase (PEPC), creating oxaloacetate. Oxaloacetate is then reduced into malate and transported to the bundle sheath cell, where decarboxylation occurs, allowing the CO2 molecule to enter the Calvin cycle [5]. By carbon fixation and the Calvin cycle occurring in two separate cells, RuBisCo is surrounded by a higher concentration of CO2, greatly reducing the chance of photorespiration. Approximately 10% of the plant population are CAM plants. Instead of photosynthesis occurring in two separate cells, like C4 plants, CAM plants complete photosynthesis in two separate parts of the day. During the night, CAM plants open their stomata to accumulate CO2, create oxaloacetate, and malate through the same mechanism as C4 plants, but during the day, their stomata are closed, and the CO2 proceeds to enter the Calvin cycle [5].
2.1. Engineering C4 Cycle into C3 Plants
With C3 photosynthesis being the least effective method of carbon fixation, genetically engineering effective photorespiration evasion mechanisms is a primary goal. The introduction of the C4 photosynthetic processes into C3 plants would help to minimize the occurrence of photorespiration. C4 plants evolved from ancestral C3 plants due to environmental conditions, such as decreased CO2 availability. C3 plants already have the necessary enzymes required to complete the processes of C4 photosynthesis; the main hindrance is the location where the photosynthesis takes place [8,9]. For C3 plants to exhibit C4 photosynthesis, the plants would need to incorporate bundle sheath cells into their processes, since C3 photosynthesis only takes place in mesophyll cells. Modification necessary to initiate this process includes increasing vein density, increasing the size and chlorophyll richness of bundle sheath cells, changing the organization of bundle sheath cells and mesophyll cells, engineering different forms of chloroplast for the respective cells, and compartmentalizing necessary enzymes into the respective cells [9]. Attempts to engineer single-cell and two-cell C4 photosynthesis mechanisms have occurred, to no avail. Since C3 plants already have the enzymes necessary to carry out C4 photosynthetic mechanisms, once the C4 mechanisms were put in place, overexpression and misexpression become an issue. It is hypothesized that the causes of failure revolve around the inability to control diffusion between the chloroplast and cytosol, affecting the necessarily high concentration of CO2 [9]. In order to effectively induce C4 photosynthetic mechanisms in C3 plants, extensive gene modifying technologies will be necessary, but these technologies are still in development. The technology would need the ability to insert genes necessary for the C4 cycle and possibly to remove genes that may suppress the cycle. The use of CRISPR/Cas9 is likely to be useful in knocking out unnecessary C3 functions (Schuler et al., 2016). Recent developments in next-generation sequencing (NGS) have led to breakthroughs in C4 research. Through NGS, five C4 plant species have been sequenced. Following this development, previously unknown components of C4 cycle biochemistry were discovered, and molecular insight regarding Kranz leaf anatomy became available [9].
2.2. Engineering CAM into C3 Plants
Another method to increase the effectiveness of carbon fixation in C3 plants would be to genetically engineer the photosynthetic mechanisms of CAM into the plants. Similar to C4 plants, CAM organisms also evolved from C3 plants, but their mechanisms for evolution are widely unknown. While CAM photosynthetic mechanisms and C4 plants share their regulatory elements with C3 plants, CAM seems to be a more likely candidate for genetically engineering C3 plants, based on its single-cell structure [10]. CAM plants were likely to evolve to increase the plants’ ability to survive in arid conditions, such as those of desert cacti. For CAM to be engineered into C3 plants, CAM-specific genes would need to be transferred into the plant, some pre-existing genes would require modifications, and traits specific to leaf structure might need to be modified. However, it is unknown how many genes would need to be modified for this process to be successful [11]. A few known modifications that C3 plants would need are a mechanism for nocturnal opening and diurnal closing of stomata, a mechanism for the decarboxylation of malate, and an anatomical modification to increase leaf succulence [10,11]. CAM engineering is still out of reach for modern plant biotechnology, since it only has the capability of modifying a few genes at a time. Synthetic biological approaches to CAM engineering prove to offer more potential. For synthetic approaches to CAM engineering, it is necessary to compile a library containing the different parts of CAM genes; design a circuit that includes carboxylation, decarboxylation, stomatal control, and anatomical structures; assemble multi-gene constructs; transfer the constructs into the chromosomes of C3 plants; and evaluate the newly engineered plants’ functionality. A few of the necessary parts for library construction include the promoter, terminator, protein-coding sequence, insulator sequence, recombination site, and linker sequences. After the library assembly, the previously mentioned steps would repeat in a cycle until the performances of the engineered plants are optimized [11].
3. Carbon Dioxide Fixation by Microbes
Even before eukaryotic plants evolved, single-celled organisms such as cyanobacteria had come up with mechanisms to fix atmospheric carbon dioxide into useful products [12]. Even though the mechanisms are primitive evolutionarily in comparison to eukaryotic plants, certain species of microorganisms have more efficient carbon-capture mechanisms and unique sets of proteins working together to make carbon sequestering possible [13,14,15].
Microorganisms that fix carbon in extreme conditions such as acetogens and methanogens have come up with exceptionally efficient methods of fixing carbon [13]. A high variety of enzymes exist in these ancient microbes as compared to plant systems and could provide new insight into carbon sequestering. The metabolic pathways used by acetogens and methanogens are evolutionarily among the oldest and have had a great impact on later lifeforms [13,16].
While the carbon-capturing enzymes in plants are incompetent and problematic, the ones in methanogens and acetogens are highly efficient, and downstream processes use different strategies to minimize the cost of carbon fixation in them. The two-step carbon-fixing pathway followed by these extremophiles is the reverse of all other carbon-fixing pathways [15,17]. Reduction of carbon followed by carboxylation is followed by methanogens and acetogens. This pathway, namely, the reductive acetyl-CoA pathway, is least energy-demanding and thus favored by early carbon-sequestering microorganisms.
3.1. Archaea Bacteria
This is a group of unicellular, primitive extremophiles living in harsh environments that have suboptimal temperatures and no oxygen. Among the three subdivisions, namely, halophiles, thermoacidophiles, and methanogens, the latter is of concern to our study. Methanogens are used to produce methane, a biofuel. A study showed Methanobacteriaceae, Methanospirillaceae, and Methanosarcinacea have high potential in producing methane by converting carbon dioxide emitted from industries [16].
3.2. Acetogens
Bacteria such as Moorella thermoacetica, Clostridium ljungdahlii, and Clostridium ragsdalei are anaerobic acetogens that have been known for syngas (CO2, CO, and H2) fermentation [14]. Acetogens, such as methanogens, use the Wood–Ljungdahl pathway/reductive acetyl-CoA pathway to fix carbon dioxide into acetic acid or ethanol. Electrons required for the fermentation of the syngas are derived from hydrogen gas via hydrogenase enzyme or from carbon monoxide through carbon monoxide dehydrogenase. Pathways other than reductive acetyl-CoA are needed for ATP production, because net ATP production in acetogens and methanogens via this pathway is zero [14,18].
The pathways followed by the acetogens and methanogens include the use of various energy-conserving enzymes that can use low-potential enzymes, which makes survival in difficult environments possible [13,15,18,19,20]. Though all these mechanisms are fascinating, our interest lies mostly in the extraordinary carbon dioxide capture mechanism. The enzyme responsible for the reduction and carboxylation of carbon dioxide is formyl-methanofuran dehydrogenase complex, a molybdenum/tungsten-containing protein complex. While every other formyl-methanofuran dehydrogenase enzyme complex releases carbon dioxide, in methanogens and acetogens, it fixes carbon [21]. What makes the carbon capture extraordinary are the unique/specialized enzymes that capture and fix carbon dioxide with low-energy electrons.
The main enzyme that is relatively conserved in both acetogens and methanogens is carbon monoxide-dehydrogenase/acetyl-CoA synthase, which is involved in the pathway involving the reduction of carbon dioxide. It uses Fe- and Ni-containing clusters to make this process possible. The C-cluster containing Fe-[NiFe3S4] uses low-potential electrons to reduce carbon dioxide to CO, which is then channeled to acetyl-CoA synthase to be fixed on Ni-[Fe4S4] containing clusters. The conversions of these reactions require metal catalysts and hydrophobic channels. A system of 46-[Fe4S4] clusters combined with catalytic enzymes and hydrophobic/hydrophilic channels working together in the presence of an electron donor make this no-energy carbon fixation possible in methanogens [21]. Acetogens have a wider variety of enzymes as compared to methanogens [22].
3.3. Methanogens
Methanogens have an astoundingly complex biochemical makeup to convert carbon dioxide into methane. More than 200 genes are involved in coding for the enzymes, which are unheard of in plants. These genes are specific to archaea and cannot be found in other organisms. Electrochemical sodium potential is used to derive energy for certain steps of the reaction [23].
Another group of microbes involved in carbon fixation that use the same reductive acetyl-CoA pathway is Clostridium spp. Similar to certain acetogens, these species of bacteria also require hydrogen gas to function. These are found in anaerobic conditions and are highly sensitive to atmospheric oxygen [24].
3.4. Algae
Red algae are of importance to our discussion because of a few convergently evolved enzymes. RubisCO tends to form toxic metabolites due to its oxygenase activity, which then have to be repaired. RubisCO is inhibited by inhibitors such as xylulose 1,5-bisphosphate (XuBP) or Ribulose-1,5-bisphosphate. RubisCO in plants needs to be remodeled once they are inactivated by inhibitory action on xylulose 1,5-bisphosphate (XuBP)/Ribulose-1,5-bisphosphate (RuBP) binding. Reactivation is achieved by motor enzymes called rubisco activase, which, with the help of ATP hydrolysis, mediate reactivation of RubisCO [25].
Rubisco activases have a major role to play in the activity of the RubisCO enzyme. These belong to a superfamily of proteins called ATPases associated with various cellular activities (AAA+) [25]. Rubisco activases act as molecular chaperons “reactivate” RubisCO after an inactivation event. Red algae RubisCO has better kinetics and is a target for synthetic pathways. However, these need rubisco activases to achieve the desired rate of carbon dioxide fixation. Green algal rubisco activases and plant activases differ from red algal activase enzymes and are thought to be inherited from proteobacterial horizontal gene transfer of gene cbbX [26,27].
Algal species which have properties such as highest carbon capture and incorporation, maximum carbon dioxide stress tolerance, maximum nutrients assimilation, pH, and thermal fluctuation tolerance are looked into for industrial purposes [28]. Although for carbon dioxide assimilation such rigid norms for selection are not necessary, all the qualities are highly desirable. Botryococcus braunii, Scenedesmus obliquus, Nannochloropsis oculate, and Chlorella vulgaris are a few species that are of industrial relevance in biofuel production. These algal species meet the required criteria of high carbon dioxide assimilation and stress tolerance [29].
Marine autotropic species have an advantage over the terrestrial alternatives because of the presence of large bodies of water, in which the area for growth is not limited. The land is a limited resource, and competition for requirements such as food and human residency would always be a priority [30]. These issues would only increase with time, but with marine algal species, this problem would not arise. Other advantages are the high assimilation rate and byproducts that could be made from algae. These advantages give algae an edge among other options.
3.5. Proteobacteria
Bacteria belonging to the phylum of proteobacteria are also known to fix carbon by using the reverse tri carboxylic acid (TCA) cycle (also known as reductive TCA cycle) directly into the cytoplasm. Certain members of this phylum are of great relevance to carbon-sequestering efforts due to their ability to convert it into commercially valuable products such as bioplastics and medicine. In a study using the extracts of T. denitrificans and Candidatus Arcobacter sulfidicus to test for carbon-fixing pathways, no RubisCO was detected in either of the two bacteria, but the activity of enzymes such as ATP citrate lyase and 2-oxoglutarate: ferredoxin oxidoreductase, which are enzymes involved in the reductive TCA cycle (Figure 1), were found in it [31]. Other groups of proteobacteria may contain the RubisCO enzyme, such as Acidithiobacillus ferrooxidans, which is a well-studied acidophilic bacteria [32]. Detailed studies on the activities of rubisco activases involved in proteobacteria also pave the way for synthetic pathways [33].
3.6. Engineering Microbes for Carbon Dioxide Fixation
Naturally occurring carbon-dioxide-fixing autotrophs have limitations that need to be dealt with before they can be used for macroscale carbon sequestering. Autotrophs such as acetogens and methanogens have efficient carbon capture and carbon assimilation enzymes but require certain conditions such as suboptimal temperature, pH, and anaerobic conditions for growth. Certain proteobacteria need specific requirements, such as low pH, for growth. Fungi assimilate carbon quickly but would require dead decaying matter for growth and take much-needed terrestrial space. Algae seem to be the best option yet, but with the pressing demands for carbon sequestration, more efficient microbes may be needed.
Scientists are exploring possibilities for combining synthetic pathways from different organisms to propose a new pathway that is superior to natural pathways [30]. Genetically engineering microorganisms to produce unique and efficient pathways for carbon sequestering is an important field of research. Researchers have already used genetically engineered E. coli for carbon sequestration [34]. Among other factors, the motivation to choose E. coli lies in their accelerated doubling time, being cost-effective and easier to genetically engineer as compared to multicellular organisms. Various groups are exploring the possibility of artificially adding genes to code for carbon-fixing enzymes in them. Table 1 compares the advantages and disadvantages of various carbon fixation pathways in bacteria and algae.
In one study, two sequential enzymes along with carbonic anhydrase from cyanobacteria for carbon fixation were incorporated into E. coli. The carbon dioxide fixation rate was 19.6 mg/Lh, which is comparable to certain species of cyanobacteria and algae [35]. Synthetic pathways have also been designed, some of which have been demonstrated in vivo. The crotonyl-CoA/ethylmalonyl-CoA/hydroxybutyryl-CoA (CETCH) pathway is an example of a synthetic carbon dioxide fixation pathway. It uses a kinetically faster carboxylase and 17 enzymes from nine different organisms to achieve this synthetic pathway. It implements enoyl-CoA carboxylase/Reductase (a carboxylase found in alpha-proteobacteria and Streptomycetes), which is insensitive to oxygen and has two to four times as much catalytic efficiency as compared to RubisCO [36].
Cell-free conversion of carbon dioxide to methanol was carried out by using three dehydrogenases from Clostridium ljungdahlii, Burkholderia multivorans, and Saccharomyces cerevisiae. Formate dehydrogenase, formaldehyde dehydrogenase, and alcohol dehydrogenase were respectively isolated, cloned, purified, and manipulated for this study [37]. Another study used 13 enzymes from five different organisms to construct a synthetic pathway that fixed carbon dioxide into malate [38].
4. Phytoplankton
Oceans are the largest reservoirs of carbon on the planet. It is strongly believed that the ocean’s capacity to store carbon is not infinite and the pH will change by 0.3 to 0.4 units by the turn of this century [44]. In oceans, 90% of the inorganic form of carbon is in bicarbonate, with the release of a proton that contributes to lowering the ocean pH causing acidification [45].
Marine phytoplankton account for more than half of the carbon dioxide fixation of Earth. The export of carbon is highest at the photic zone of the ocean, which is dominated by phytoplankton. The term phytoplankton comes from the Greek words phyto (plants) and plankton (wanderers or drifters). Plankton can also be zooplanktons that feed on phytoplankton and release fecal pellets that are made of dissolved carbon particles. Phytoplankton constitute organisms across multiple kingdoms such as Kingdom Monera, e.g., Cyanobacteria; Kingdom Protista, e.g., fire algae, diatoms, dinoflagellates, etc.; and Kingdom Plantae, e.g., Brown algae (Sargassum); while Kingdom Animalia consists of zooplankton [46].
The growth of the phytoplankton mainly depends upon three factors, i.e., nutrients, sunlight, and carbon dioxide. Phytoplankton-like plants have chlorophyll that fixes carbon dioxide to glucose using the Rubisco enzyme. Phytoplankton-like plants are sensitive to nutrient availability, and hence, depending on the species, their growth is limited in the ocean based on nutrient availability. Other factors that affect phytoplankton growth are water depth and temperature, pH and salinity, and the diversity of predators that feed on them. When all factors are optimal, there phytoplankton boom over the surface of the ocean to an extent that can be captured by satellite images, as they cover hundreds of miles over the ocean, jeopardizing other oceanic flora and fauna [47,48].
4.1. Cyanobacteria
Cyanobacteria are a class of prokaryotic microorganisms that are blue-green photo-autotropic Gram-negative bacteria or algae. These are the predecessors of green plants and possess carboxysomes, which are carbon dioxide concentration mechanisms. The enzymes used for carbon capture are the same as in plants, namely, RubisCO, and carbonic anhydrase. Carbon-concentrating mechanisms exist in cyanobacteria, similar to algae that increase the carboxylase activity of RubisCO enzyme by increasing carbon dioxide concentration near the fixing enzyme.
Among all the naturally found carbon-sequestering organisms, algae seem to be most efficient in converting carbon dioxide to biomass and fuel. Microalgae include microorganisms such as diatoms, euglenoids, and green, blue, golden, red, brown, and yellow algae, which fix carbon dioxide using RubisCO and are mostly single-celled. With a biomass accumulation rate that is 100-fold faster than terrestrial plants, microalgae are a very promising candidate for carbon-sequestering units [49,50]. Algae are high in lipids and could have direct use in the production of biofuels [51]. Biochar derived from algae has been studied and found to have excellent properties to be used as a fertilizer [52]. Microalgae have carbon-concentrating mechanisms, which increase the concentration of carbon dioxide near the RubisCO enzyme by separating it into a different membrane compartment, into which carbon dioxide and bicarbonates are transported. Uluva and Laminaria species of microalgae are prime choices for bioenergy production [34].
Every 1 kg of algae can fix 1.83 kg of carbon dioxide in ideal conditions [29,53]. Atmospheric carbon dioxide concentrations range from 0.03–0.06% (v/v), and algae can grow efficiently at 2% or higher. Certain species such as Scenedesmus sp. have a carbon dioxide tolerance even at 10–20% (v/v) concentration of carbon dioxide. An increase in biomass is the most significant pointer to indicate carbon dioxide sequestering [33].
4.2. Dinoflagellates
Dinoflagellate are a major part of oceanic phytoplankton that use Rubisco form II as a primary enzyme for carbon fixation [54]. The form II has both lower specificity and affinity for carbon dioxide/oxygen. This suggests that they have an advanced mechanism to increase carbon dioxide concentration, since they cannot fix carbon dioxide at lower ambient carbon dioxide concentration [55]. Of the various isomers of carbonic anhydrase (α, β, γ, and δ), δ-carbonic anhydrase has been implicated in lowering carbon dioxide concentration [54]. The enzyme carbonic anhydrase (CA) catalyzes rapid conversion of carbon dioxide and bicarbonate ion [56]. Many dinoflagellates have carbon-concentrating mechanisms (CCMs) to transport carbon into the cell [54]. One of the biggest bio-indicators of global warming is the bleaching of coral reefs. Coral reefs that belong to Kingdom Animalia are a complex system of multiple species belonging to phylum coelenterates. The base of the ecosystem is built upon dinoflagellates that live as intracellular photosynthetic symbionts. Loss of these dinoflagellates is believed to be the primary cause of coral bleaching that eroded 85% of the Great Barrier Reef in Australia in 2016 [57,58]. How do coral reefs become bleached? A slight change in water temperature (2 °F) disturbs the photosynthetic electron transfer and in the process damages the PsbA (D1), a photosystem II reaction center protein. This disruption of photosynthesis triggers the bleaching of coral reefs [59,60,61,62,63].
While genetically engineering cyanobacteria has some success, engineering genes to improve the carbon fixation in diatoms and dinoflagellates has been a challenge due to the lack of critical gene transformation strategies. Initial studies of transformation using silicon carbide whiskers were suggested, but the experiments were difficult to reproduce [64,65]. Experiments with glass beads were performed for transient expressions, but even these were hard to reproduce [57,66].
Recent studies showed that the stable transformation of dinoflagellates is reproducibly possible when they bombarded microparticle-containing plasmid-like minicircles that carried chloroplast gene psbA. They successfully transformed two minicircles providing resistance to chloramphenicol and astrazine and in the process assisting the selection of successful transformants. Inability to stably transform has less to do with the inherent genetics of protists than it has to do with the DNA’s inability to cross the membrane barrier [57].
4.3. Diatoms
Diatoms exhibit a wide range of metabolic diversity due to their evolution that includes endosymbiosis among diverse lineages. This is why they can be used to make a wide range of compounds. Previous studies have shown that manipulating carbon dioxide levels of Navicula pelliculosa, P. tricornutum, T. pseudonana, and Asterionella formosa [67] can increase the accumulation of both lipids and carbohydrates. Diatoms T. weissflogii and P. tricornutum have exhibited excellent absorption systems for carbon dioxide and bicarbonate at quantities typical of ocean surface waters. It has also been observed that their absorption rates can adjust to a wide range of inorganic carbon supplies [68]. Mixotrophic cultivation regimes, in addition to photoautotrophy, can aid in the production of higher biomass concentrations and productivities [69,70,71].
Marine photosynthetic organisms such as diatom and other microalgae uptake carbon dioxide using a mechanism called carbon-concentrating mechanism or CCM [69,70]. In diatoms, the carbon dioxide travels inside the cells similar to plants and concentrates near the site where Rubisco is located for fixation. This mechanism is important because it helps regulate the carbon flux and raise it to higher concentrations inside the cell compared to the outside for optimal Rubisco activity [69,70,71,72,73]. Very few mechanisms are known for the majority of diatoms. For the ones that are known, they use the Rubisco Calvin–Benson–Bassham (CBB) cycle in their chloroplasts, carbonic anhydrases, Glyceraldehyde-3-phosphate dehydrogenase (GAPDH), Phosphoglycerate kinase, Fructose 1,6 bis phosphatase, Sedoheptulose-1,7-biphosphatase, and various transporters [65,66,70,71].
Presently, there is a dearth of literature with regards to carbon fixation in diatoms. Studies have shown that the assimilation of carbon dioxide is not linearly proportional to the availability. P. tricornutum showed a higher fixation rate with supplemented carbon dioxide concentration but had reduced biomass productions. The carbon fixation rate did not change when subjected to diverse pH ranges. There is diversity among diatoms for carbon substrate specificity for fixation purposes. While P. tricornutum assimilates in the form of bicarbonate, T. pseudonana takes up carbon dioxide predominantly [67,68,69,70,71,72,73].
4.4. What Are the Challenges in Genetically Modifing Diatoms?
Though diatoms are being used as bioreactors for environmentally sustainable manufacturing of valuable metabolites, there is little research to explore their potential for carbon sequestration and fixation [70,72,73]. There is a huge opportunity available for research on this aspect; hence, it is surprising that this has been less explored. We predict one of the reasons is the environmental impact of genetically modified microorganisms. Diatoms, like other microalgae, provide huge potential as promising candidates for carbon sequestration and fixation. Genetically modified diatoms have the potential for explosive growth like other microalgae and are hard to predict. They can have a huge environmental impact, jeopardizing the marine ecological balance [74]. The genetically modified diatoms also need to be tested for the new metabolites that they may start producing post-modification that may prove toxic in large quantities to the grazers that are at the top of the food chain. Hence, marine ecosystem modeling need to be used to test and simulate the challenges associated before introducing a new species in the wild. New experimental marine models need to be developed that can assist in predicting and simulating the underlying challenges in a marine ecosystem.
5. Photosynthesis and Rubisco: An Evolutionary Perspective
When oxygenic photosynthesis evolved around 2.4 billion years ago, it is believed that all the oxygen-evolving organisms used the ribulose bisphosphate carboxylase-oxygenase (Rubisco)- photosynthetic carbon reduction cycle (PCRC) despite there being five other major CO2 assimilation pathways [75,76]. Rubisco-PCRC was dominant, with the highest specific activity in presence of carbon dioxide at high concentrations without the presence of oxygen. The other five pathways are (a) reverse tricarboxylic acid cycle (TCAC), (b) Hydroxypropionate, (c) Hydroxypropionate-4-hydroxybutyrate, (d) dicarboxylate-4-hydroxybutyrate, and (e) Wood–Ljungdahl pathway. What is unique about the Rubisco-PCRC pathway compared to the other five pathways is that oxygen is a competitive inhibitor for the Rubisco-PCRC pathway, while it is not for all the other five pathways. Rubisco enzyme is present both in prokaryotes and eukaryotes [77,78,79,80]. Four major forms of Rubisco have been identified (I, II, II/III, and IV) that are known to catalyze carboxylation and oxygenation of RuBP (Ribulose 1,5 Bisphosphate) [81,82]. Form II of Rubisco is associated with anoxygenic photosynthesis by proteobacteria before the evolution of oxygenic photosynthesis. The rise in oxygen concentration in the atmosphere with the proliferation of cyanobacteria over one billion years ago caused a decrease in CO2, ushering in the Great Oxidation Event (GOE) [75]. This change in atmospheric oxygen led to the evolution of oxygenic photosynthesis. The first time when Rubisco was exposed to oxygen led to the oxygenation of RuBP that in turn produced 2 phosphoglycolates, ushering in the evolution of the photorespiration pathway [75]. This photosynthesis with the evolution of oxygen eased the evolution of aerobic respiration, which is a bio-energetically more efficient process in comparison to anaerobic fermentation [75]. It is believed that the evolution of form I of Rubisco is closely linked to the increase in oxygen in the atmosphere [75]. The lowering of carbon-dioxide-to-oxygen ratios in the aerobic environment led to the evolution of more carbon-dioxide-specific enzymes starting to take place.
5.1. Can Engineering Photosynthesis Provide Solution?
Plants play a major role in fixing the global carbon footprint. Modern agriculture techniques with the use of fertilizer and irrigation provide evidence that carbon fixation could be a rate-limiting step. Many C3 plants when exposed to twice the carbon dioxide concentration have been shown to produce a significantly higher amount of biomass [83]. This suggests that plant growth could be regulated by manipulating biochemical pathways related to carbon fixation [84]. Studies have shown that Arabidopsis grew faster with more soluble sugars and enhanced shoot and root biomass when the natural photorespiration pathways were replaced with bacterial photorespiration pathways [85]. In another study with tobacco plants overexpressing an enzyme (sedoheptulose-1,7-bisphosphatase) that operates in the reductive pentose phosphate cycle, (rPP); showed enhanced biomass as well as a 30% increase in photosynthesis [86,87].
5.2. Why Is It Important to Look beyond Rubisco?
Diverse autotrophic organisms use the reductive pentose phosphate cycle (rPP cycle) for carbon dioxide assimilation. This cycle is rate-limited by the low catalytic rate of Rubisco (ribulose-1,5-bisphosphate carboxylase/oxygenase) [88,89,90]. The turnover of Rubisco and the low catalysis rate are negatively correlated, which suggests that the Rubisco enzyme is perhaps naturally optimized after evolving for millions of years [90,91,92]. Hence, optimizing Rubisco using genetic engineering may not prove to be a profitable endeavor [92]. This also suggests that it is imperative to look beyond the rPP cycle and identify Rubisco-independent pathways that have a higher carbon fixation rate.
5.3. What Are the Other Natural Carbon-Fixing Metabolic Pathways?
To date, apart from the rPP cycle, there are five other natural carbon-fixing metabolic pathways that have been identified. They are (a) 3-hydroxypropionate cycle; (b) reductive tricarboxylic acid (rTCA) cycle; (c) oxygen sensitive reductive acetyl-CoA (rAcCoA) pathways) (Figure 2); (d) dicarboxylate/4-hydroxybutyrate cycle (Figure 3); and (e) 3-hydroxypropionate/4-hydroxybutyrate cycle (Figure 3) [90,92,93,94].
6. Exploring Other Alternatives: Introduction to Synthetic Pathways
To explore other alternatives, it is important to be aware of various factors that are to be considered while searching for alternative pathways. Studies have been conducted exploring approximately 5000 enzymes from the KEGG database to seek potential carbon fixation pathways [95]. The pathways were analyzed and evaluated based on four criteria, i.e., kinetics, thermodynamics, the efficiency of energetics, and topological compatibility. All the criteria were compared to the six naturally occurring carbon fixation pathways to validate the results [96]. Other studies have included a few other factors in addition to the ones mentioned above such as toxicity of metabolites produced and regulation of the cycle [97,98,99].
Combining the six criteria from the above studies would provide a comprehensive matrix to analyze and establish novel carbon fixation pathways.
6.1. How to Use the Criteria Matrix to Select Synthetic Carbon Pathways?
6.1.1. Kinetic Analysis
While considering the kinetics of theoretical pathways, the Kcat/Km of each of the enzymes should have a high value for Kcat and a low value for Km. This will ensure that there is high enzyme specificity in addition to a high rate of catalysis [100]. Now the question is, where do we start? The best way to get started is to look for rate-limiting steps and identify enzymes associated with them. An example could be finding carboxylation enzymes that have higher specific activity and have higher affinities for carbon dioxide and bicarbonate ions under ambient conditions. Enzymes such as phosphoenolpyruvate (PEP) carboxylase, pyruvate carboxylase, acetyl-CoA, and propionyl-CoA carboxylases are some of the examples.
6.1.2. Resource Consumption and Energetic Efficiency
Too much dependence on kinetics could be detrimental, as it does not provide information on resource consumption for each pathway. It is important to calculate the energetic cost simultaneously for each pathway under consideration. The energetic cost for each pathway can be classified into NADPH cost and ATP cost. While calculating NADPH cost, one needs to consider the number of moles of NADPH consumed to produce one mole of a product. Under NADPH, other redox carriers such as ferredoxins and FADH2 are also counted. Similarly, for the ATP cost, consumption of 1 mole of non-redox carriers such as nucleotide triphosphates (NTPs), coenzyme A thioesters, and phosphoesters for the production of 1 mole of product needs to be calculated.
6.1.3. Thermodynamic Analysis
While analyzing kinetics and resource consumption, it is imperative to concurrently conduct a thermodynamic analysis of the pathways. At ambient conditions and carbon dioxide concentrations, the Gibbs free energy should be negative, i.e., (∆Go << 0). Some of the natural pathways such as the rTCA cycle [7] and the rAcCoA pathway do not function under ambient carbon dioxide concentrations if the ranges of pH and ionic strengths are broad, as the (∆Go > 0). Interestingly, the rTCA cycle can function under a broad range of pH and ionic strengths if the carbon dioxide concentrations are 100 times higher than the ambient concentration. Similarly, the rAcCoA pathway can function at pH < 8 under high carbon dioxide concentrations. This is possible especially for C4 and CAM plants that use mechanisms to concentrate carbon dioxide. Hence, it could be safely concluded that both the rTCA cycle and the rAcCoA pathway work under anaerobic conditions. This is important, as it is a known fact that anaerobes use pathways that have low energetic yields. Hence, using such pathways would limit our success in enhancing carbon fixation yield. The only way these two pathways can be feasible is if the plants are grown under high-carbon-dioxide conditions.
6.1.4. Analysis of Distributed Thermodynamic Bottlenecks
Theoretically, analyzing the energetics of the whole pathway as stated above can predict whether the pathway is feasible or not. It can also help us predict when there are no constraints on the concentration of various metabolites. In reality, the concentrations of metabolites have an upper and lower limit. Hence, analysis of the energetics of the overall pathway does not guarantee feasibility. There is always a chance the predictions can fail despite being ∆Go < 0, as there are sub-pathways within the major pathway that are concentration-dependent. These sub-pathways are known as thermodynamic bottlenecks.
To analyze thermodynamic bottlenecks, the major pathway is analyzed by analyzing the Gibbs free energy change at an ambient temperature of the sub-cycles. ∆Go of each sub-cycle is experimentally analyzed at multiple pH ranges and multiple ionic concentrations without accounting for the ATP hydrolysis. If the ∆Go of all the sub-pathways for all the pH and ionic range is negative, then it can be safely concluded that the following major pathway is feasible and a promising candidate for future carbon fixation studies [101,102,103].
6.1.5. Metabolic Regulation and Compatibility
As the pathways are analyzed, it is important to simultaneously consider the integration and compatibility of the network with other pathways inside the cell. The key parameters to consider are a number of enzymes involved in the fixation cycle and conducting random sampling followed by flux analysis to determine compatibility with the other endogenous network in the cell. When attempting to design a synthetic pathway, it is lucrative to engineer it in such a way that there are as few of these manual checkpoints as possible to reduce the number of complications that would arise from the need to maintain them. However, there are exceptions. An instance where having regulation points would be useful is when one wants to limit the number of irreversible enzymes that produce intermediates to prevent flux imbalances [7].
6.1.6. Analysis of Metabolites
The metabolic intermediates should be as unreactive and non-toxic to the cell as possible. Not only this but having the intermediates be considerably hydrophobic should be avoided in order to prevent them from leaking out from within the cell [79]. Additionally, metabolites containing low concentrations of cofactors should be used sparingly, if at all [7]. This will reduce the number of challenges that will arise from having to regulate cofactors and prevent them from depleting.
6.1.7. Application of Synthetic Pathways
Malonyl-CoA-Oxaloacetate-Glyoxylate (MOG) is one of the synthetic pathways discovered using the method mentioned above. Promising candidates for carbon-fixing enzymes were identified, and the pathway was predicted to be thermodynamically feasible [102,103]. MOG showed similarity with carboxylation products, namely Malonyl-CoA and oxaloacetate. It was called the Malonyl-CoA-Oxaloacetate-Glyoxylate (MOG) pathway because of their common carboxylation and export products. There are limitations like some other pathways, such as requiring a certain temperature for function, and glyoxylate produces carbon dioxide via decarboxylation [7,79].
7. Chemical Alternatives for Fixing Carbon Dioxide
7.1. Carbon Nanotube (CNT) Synthesis
Synthesis of carbon nanotubes from carbon dioxide is one of the most promising non-photosynthetic methods for carbon capture, utilization, and storage [103]. The carbon material developed from gaseous carbon dioxide can have multiple utilities. Compared to amorphous carbon, carbon nanotubes are superior due to their superior electronic/ionic properties and mechanical, chemical, and thermal stability [104,105,106]. Hence, they are used to make supercapacitors, lithium-ion batteries, and fuel cells with high performance. This method, though promising, did not seem feasible, as it was hard to scale up due to extreme experimental conditions. For instance, it required supercritical carbon dioxide with magnesium and lithium as reducing agents [104,106]. This made it more reactive compared to regular carbon dioxide. To conduct such an experiment, one needs to maintain a high temperature around 1000 °C and pressure of 10 kbar, which made it highly improbable to scale up [107,108,109,110]. Recent advances in this production technique have not only made it more feasible but scalable as well. CNTS can now be synthesized from gaseous carbon dioxide by one-step chemical vapor deposition in the presence of sodium borohydride as a reducing agent and nickel catalyst at 500 °C and 1 atm pressure [105]. Another recent advancement was made using another greenhouse gas; i.e., reforming of methane was done by reacting it with carbon dioxide in the presence of a catalyst to produce syngas using a series combination of two reactors (carbon generator or CARGEN), which converts the gasses into multi-walled carbon nanotubes. Carbon nanotubes are in use in electronics, lithium-ion batteries, aerospace technology, and carbon fiber, to name a few applications [111].
7.2. Fixing Carbon Dioxide into Plastics
Recent research has shown that using a technique known as flame spray pyrolysis (FSP) generates zinc oxide nanoparticles that can convert carbon dioxide into syngas [112]. Syngas when mixed with hydrogen and carbon monoxide at various ratios in turn can generate various chemicals such as diesel, alcohol, and plastics [111,112]. With this method, carbon dioxide can be converted into precursors, which in turn can be used to generate various chemicals including plastics [111].
7.3. Metal-Organic Frameworks (MOFs)
MOFs hold a lot of promise because they are porous and have a large surface area [113]. Though they are being used as a heterogeneous catalyst, more improvements are required to lower the cost of production and energy requirement. Recently, a MOF was created that has acrylamide groups that are accessible and has a copper group exposed, thereby increasing the efficiency of carbon dioxide adsorption [114]. In these MOFs, they extended the tricarboxylate ligand backbone using click chemistry that helped create larger pores. In another study, Zn-MOF-184 was made that had strong Lewis acidity due to zinc cations, and it showed the highest catalytic activity upon cycloaddition of carbon dioxide under mild conditions, i.e., 80 °C for 6 h under mild solvent conditions and balloon pressure [115]. This is an evolving field and holds a lot of promise for further improvement and research for sequestering carbon dioxide. Presently, there is enough information to develop intuitive models to optimize acid-base properties to improve and expedite the search for MOFs with remarkably improved catalytic properties [116].
7.4. Nanotechnology Based Carbon Dioxide Fixation
7.4.1. Metal-Based Nanocatalyst
Similar to MOFs, metal-based magnetic nanocatalysts hold a lot of promise. In one of the recent studies, a copper-based magnetic nanocatalyst was found to be a simple yet efficient way of carbon dioxide fixation. This nanocatalyst adds epoxide and carbon dioxide to form cyclic carbonates through cycloaddition [117].
Nickel-based nanoparticles hold a lot of promise, as they can catalyze the hydration of carbon dioxide. This is important, as during carbon dioxide absorption, the hydration reaction is the rate-limiting step. These nanoparticles showed optimal activity around 20–30 °C with pH less than 8. When these nanoparticles were mixed with potassium carbonate solution, the adsorption capacity was enhanced by 77%. At a commercial level, heating at 150 °C regenerated the potassium-carbonate-saturated nanofluid. Interestingly, the nanoparticles did not show any surface oxidation post-regeneration [118].
7.4.2. Nanoparticles for Capturing Post-Combustion Carbon Dioxide
Fossil fuel derivatives such as coal, natural gas, and crude oil are utilized to generate approximately 67% of the world power supply. Hence, one of the largest sources of carbon dioxide emitters is constituted by the power plants that consume fossil fuels such as coal and natural gas to generate electricity [119]. Water-based nanofluids have been tested for the elimination of carbon dioxide. It was found that aluminum oxide and calcium carbonate were the best-performing nanoparticles. The assessment for efficiency was made based on the surface area, temperature, and pressure to adsorb carbon dioxide.
8. Conclusions
Global warming due to the accumulation of carbon dioxide is a reality whose effect could already be observed looking at the receding ice at both the poles and coral bleaching [120]. This warming has resulted in the rise in the sea level, erratic weather changes, and droughts, to name a few effects [120]. This review evaluates various research avenues and highlights the challenges for each avenue. Interestingly, present analyzing of carbon usage places too much emphasis on carbon footprint. The developed nations have exported their carbon footprint to the developing nations such as India, China and other Southeast Asian countries. Hence, they are responsible for providing goods to the world and at the same time are also held responsible for increasing carbon footprint (https://www.euronews.com/green/2021/02/06/why-we-re-all-to-blame-for-china-and-india-s-filthy-co2-emissions (accessed on 17 February 2022))). There is no technological breakthrough yet to minimize pollution from major polluting industries such as steel, cement, various metal-ore plants etc. Therefore, the only way to reduce carbon footprint is to stop production. If these countries cut down production, the whole supply chain around the world will be disrupted, which is already in a mess thanks to COVID. Due to this reason, advances in chemical carbon fixation are imperative to minimize the carbon emission from high-polluting industries. As stated previously, since the cutting down to zero emissions by India and China will not happen in the next couple of decades, synthetic and photosynthetic methods of carbon fixation need to be explored further.
Many (including heads of states) have doubted the scientific community as to whether the global warming due to greenhouse gasses is really a matter of concern. The same people argue that nature is the best engineer and would reset the imbalance on its own, and hence one should not be really be worried. The naysayers need to understand that nature has reset balance before by mass extinction and the earth has already undergone five mass extinctions (https://www.cnn.com/2022/01/30/americas/earth-extinction-andrew-knoll-crafoord-prize-scn/index.html/ (accessed on 17 February 2022)). First, the Ordovician-Silurian extinction happened 440 million years ago; second, the Devonian extinction happened 365 million years ago; third, the Permian-triassic extinction happened 250 million years ago; fourth, the Triassic-Jurassic extinction took place 210 million years ago; and lastly, the fifth and the most recent extinction, the Cretaceous-Tertiary extinction, happened 65 million years ago [121,122,123,124,125,126,127,128]. According to Knoll and colleagues, the Permian extinction can be attributed to a rapid rise in carbon dioxide due to volcanic activities. Present scientific evidence also suggests that we are rapidly progressing towards a sixth mass extinction [121,122,123,124,125,126,127,128]. Hence, it is imperative that we regulate the rise in carbon dioxide through anthropogenic activities. In this review, we discussed the various methods that have been tried to fix carbon dioxide. We also discussed the limitations and challenges for some of the methods.
Our research group is working on developing an efficient fuel cell system that can generate a reasonable amount of power with a minimal carbon footprint. We are also exploring developing efficient nanomaterials to deliver nutrients to plants with minimal carbon footprint and also easy assimilation. The alternative synthetic pathways for carbon dioxide fixation are also being explored. Reducing global warming by reduction of carbon dioxide is a massive problem and cannot be a one-step process with a single solution. It has to be a cumulative effect of multiple efforts across fields. Hence, fixing carbon dioxide using bimolecular and chemical methods has been discussed, as they would play a key role in achieving net-zero emissions in the next few decades.
Author Contributions
Conceptualization, S.R.; Investigation, S.R., J.A., N.J., M.L. and N.C. Writing—original draft preparation, S.R., J.A., N.J., M.L. and N.C.; Writing—review and editing, S.R., J.A., N.J., M.L. and N.C.; Supervision, S.R.; Project administration, S.R.; Funding acquisition, S.R. All authors have read and agreed to the published version of the manuscript.
Funding
Bowie State University supported N.J., J.T.A., M.L., and the research in S.R.’s lab.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Special thanks to A. Bissahoyo, G. Amoussou, J. Olanbiwonnu, G. Acquaah, G. Ude, and C. Esimai for their constant support to the students and our research, without which this manuscript would not have been possible.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study, in the writing of the manuscript, or in the decision to publish the results.
References
- Irfan, M.; Bai, Y.; Zhou, L.; Kazmi, M.; Yuan, S.; Maurice Mbadinga, S.; Yang, S.Z.; Liu, J.F.; Sand, W.; Gu, J.D.; et al. Direct microbial transformation of carbon dioxide to value-added chemicals: A comprehensive analysis and application potentials. Bioresour. Technol. 2019, 288, 121401. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Opgenorth, P.H.; Wernick, D.G.; Rogers, S.; Wu, T.Y.; Higashide, W.; Malati, P.; Huo, Y.X.; Cho, K.M.; Liao, J.C. Integrated electromicrobial conversion of CO2 to higher alcohols. Science 2012, 335, 1596. [Google Scholar] [CrossRef] [PubMed]
- Pacala, S.; Socolow, R. Stabilization wedges: Solving the climate problem for the next 50 years with current technologies. Science 2004, 305, 968–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiti, R.K.; González Rodriguez, H.; Ivanova, N.S. Autoecology and Ecophysiology of Woody Shrubs and Trees: Concepts and Applications; Wiley Blackwell: Chichester, UK, 2016. [Google Scholar]
- Kumar, V.; Sharma, A.; Soni, J.; Pawar, N. Physiological response of C3, C4 and CAM plants in changeable climate. India Pharma Innov. Int. J. 2017, 6, 70–79. [Google Scholar]
- Driever, S.M.; Kromdijk, J. Will C3 crops enhanced with the C4 CO2-concentrating mechanism live up to their full potential (yield)? J. Exp. Bot. 2013, 64, 3925–3935. [Google Scholar] [CrossRef] [Green Version]
- Bar-Even, A. Daring metabolic designs for enhanced plant carbon fixation. Plant Sci. 2018, 273, 71–83. [Google Scholar] [CrossRef]
- Aubry, S.; Brown, N.J.; Hibberd, J.M. The role of proteins in C3 plants prior to their recruitment into the C4 pathway. J. Exp. Bot. 2011, 62, 3049–3059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuler, M.L.; Mantegazza, O.; Weber, A.P.M. Engineering C4 photosynthesis into C3 chassis in the synthetic biology age. Plant J. 2016, 87, 51–65. [Google Scholar] [CrossRef]
- Borland, A.M.; Hartwell, J.; Weston, D.J.; Schlauch, K.A.; Tschaplinski, T.J.; Tuskan, G.A.; Yang, X.; Cushman, J.C. Engineering crassulacean acid metabolism to improve water-use efficiency. Trends Plant Sci. 2014, 19, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Cushman, J.C.; Borland, A.M.; Edwards, E.J.; Wullschleger, S.D.; Tuskan, G.A.; Owen, N.A.; Griffiths, H.; Smith, J.A.C.; De Paoli, H.C.; et al. A roadmap for research on crassulacean acid metabolism (CAM) to enhance sustainable food and bioenergy production in a hotter, drier world. New Phytol. 2015, 207, 491–504. [Google Scholar] [CrossRef]
- Fujita, E.; Muckerman, J.T.; Himeda, Y. Interconversion of CO2 and formic acid by bio-inspired Ir complexes with pendent bases. BBA Bioenerg. 2013, 1827, 1031–1038. [Google Scholar] [CrossRef] [Green Version]
- Lemaire, O.N.; Jespersen, M.; Wagner, T.; Scott, K. CO2-Fixation Strategies in Energy Extremophiles: What Can We Learn From Acetogens? Front. Microbiol. 2020, 11, 486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, P.; Rismani-yazdi, H.; Stephanopoulos, G. Anaerobic CO2 Fixation by the Acetogenic Bacterium Moorella thermoacetica. AIChe J. 2013, 59, 3176–3183. [Google Scholar] [CrossRef]
- Berg, I.A.; Kockelkorn, D.; Vera, W.H.R.; Say, R.F. Autotrophic carbon fixation in archaea. Nat. Rev. Genet. 2010, 8, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Haiza, N.; Yasin, M.; Maeda, T.; Hu, A.; Yu, C.; Wood, K.T. CO2 sequestration by methanogens in activated sludge for methane production. Appl. Energy 2015, 142, 426–434. [Google Scholar] [CrossRef]
- Martin, W.F.; Thauer, R.K. Energy in Ancient Metabolism. Cell 2017, 168, 953–955. [Google Scholar] [CrossRef] [Green Version]
- Schuchmann, K.; Müller, V. Autotrophy at the thermodynamic limit of life: A model for energy conservation in acetogenic bacteria. Nat. Rev. Genet. 2014, 12, 809–821. [Google Scholar] [CrossRef]
- Moal, G.; Lagoutte, B. Biochimica et Biophysica Acta Photo-induced electron transfer from photosystem I to NADP +: Characterization and tentative simulation of the in vivo environment. BBA Bioenerg. 2012, 1817, 1635–1645. [Google Scholar] [CrossRef] [Green Version]
- Peters, J.W.; Beratan, D.N.; Bothner, B.; Dyer, R.B.; Harwood, C.S.; Heiden, Z.M.; Hille, R.; Jones, A.K.; King, P.W.; Lu, Y.; et al. A new era for electron bifurcation. Curr. Opin. Chem. Biol. 2018, 47, 32–38. [Google Scholar] [CrossRef]
- Wagner, T.; Ermler, U.; Shima, S. The methanogenic CO2 reducing-and-fixing enzyme is bifunctional and contains 46 [4Fe-4S] clusters. Struct. Biol. 2015, 592, 1406–1416. [Google Scholar]
- Wang, S.; Huang, H.; Kahnt, J.; Mueller, A.P.; Köpke, M.; Thauer, K. NADP-Specific Electron-Bifurcating [FeFe]-Hydrogenase in a Functional Complex with Formate Dehydrogenase in Clostridium autoethanogenum Grown on CO. Bacteriology 2013, 195, 4373–4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyu, Z.; Shao, N.; Akinyemi, T.; Whitman, W.B. Methanogenesis. Curr. Biol. 2018, 28, 727–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, G. Variations of the Acetyl-eoA Pathway in Diversely Related Microorganisms That Are Not Acetogens. In Acetogenesis; Chapman & Hall Microbiology Series; Springer: Boston, MA, USA, 1994. [Google Scholar]
- Mueller-cajar, O.; Stotz, M.; Wendler, P.; Hartl, F.U.; Bracher, A.; Hayer-hartl, M. Structure and function of the AAA 1 protein CbbX, a red-type Rubisco activase. Nature 2011, 479, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Maier, U.; Fraunholz, M.; Zauner, S.; Penny, S.; Douglas, S. A Nucleomorph-Encoded CbbX and the Phylogeny of RuBisCo Regulators. Mol. Biol. Evol. 1995, 17, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Delwiche, C.F.; Palmer, J.D. Rampant Horizontal Transfer and Duplication Eubacteria and Plastids of Rubisco Genes in eubacteria and plastids. Mol. Biol. Evol. 1996, 13, 873–882. [Google Scholar] [CrossRef] [Green Version]
- Singh, U.B.; Ahluwalia, A.S. Microalgae: A promising tool for carbon sequestration. Mitig. Adapt. Strat. Glob. Chang. 2012, 18, 73–95. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Show, P.L.; Chang, J.; Ling, T.C.; Juan, J.C. Biosequestration of atmospheric CO2 and flue gas-containing CO2 by microalgae. Bioresour. Technol. 2014, 184, 190–201. [Google Scholar] [CrossRef]
- Pimentel, D.; Whitecraft, M.; Scott, Z.R.; Zhao, L.; Satkiewicz, P.; Scott, T.J.; Phillips, J.; Szimak, D.; Singh, G.; Gonzalez, D.O.; et al. Will Limited Land, Water, and Energy Control Human Population Numbers in the Future? Hum. Ecol. 2010, 38, 599–611. [Google Scholar] [CrossRef]
- Hu, M.; Wirsen, C.O.; Fuchs, G.; Taylor, C.D.; Sievert, S.M. Evidence for Autotrophic CO2 Fixation via the Reductive Tricarboxylic Acid Cycle by Members of the ε Subdivision of Proteobacteria. Am. Soc. Microbiol. 2005, 187, 3020–3027. [Google Scholar] [CrossRef]
- Zhang, S.; Yan, L.; Xing, W.; Chen, P.; Zhang, Y.; Wang, W. Acidithiobacillus ferrooxidans and its potential application. Extremophiles 2018, 22, 563–579. [Google Scholar] [CrossRef]
- Loganathan, N.; Tsai, Y.C.; Mueller-cajar, O. Characterization of the heterooligomeric red-type rubisco activase from red algae. Proc. Natl. Acad. Sci. USA 2016, 113, 14019–14024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerotto, C.; Norici, A.; Giordano, M. Toward Enhanced Fixation of CO2 in Aquatic Biomass: Focus on Microalgae. Front. Energy Res. 2020, 8, 213. [Google Scholar] [CrossRef]
- Gong, F.; Liu, G.; Zhai, X.; Zhou, J.; Cai, Z.; Li, Y. Quantitative analysis of an engineered CO2-fixing Escherichia coli reveals great potential of heterotrophic CO2 fixation. Biotechnol. Biofuels 2015, 8, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwander, T.; Burgener, S.; Erb, T.J. A synthetic pathway for the fixation of carbon dioxide in vitro. Science 2016, 354, 900–904. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Singh, R.; Li, J.; Sung, B.H.; Cho, B.; Kim, D.R.; Kim, S.C.; Kalia, C.; Zhang, Y.P.; Zhao, H.; et al. Insights into cell-free conversion of CO2 to chemicals by a multienzyme cascade reaction. ACS Catal. 2018, 8, 11085–11093. [Google Scholar] [CrossRef]
- Shi, T.; Liu, S.; Zhang, Y.P.J. CO2 fixation for malate synthesis energized by starch via in vitro metabolic engineering. Metab. Eng. 2019, 55, 152–160. [Google Scholar] [CrossRef]
- Basen, M.; Geiger, I.; Henke, L. A Genetic System for the Thermophilic Acetogenic Bacterium. Appl. Environ. Microbiol. 2018, 84, e02210-17. [Google Scholar] [CrossRef] [Green Version]
- Mcgrath, J.M.; Long, S.P. Can the Cyanobacterial Carbon-Concentrating Mechanism Increase Photosynthesis in Crop Species? A Theoretical Analysis. Plant Physiol. 2014, 164, 2247–2261. [Google Scholar] [CrossRef] [Green Version]
- Rae, B.D.; Long, B.M.; Förster, B.; Nguyen, N.D.; Velanis, C.N.; Atkinson, N.; Hee, W.Y.; Mukherjee, B.; Price, G.D.; Mccormick, A.J. Progress and challenges of engineering a biophysical carbon dioxide-concentrating mechanism into higher plants. J. Exp. Bot. 2017, 68, 3717–3737. [Google Scholar] [CrossRef]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A Common Factor in Human Diseases. BioMed Res. Int. 2017, 2017, 9351507. [Google Scholar] [CrossRef] [Green Version]
- Munive, A.; Moulin, L. Nodulation of legumes by members of the β -subclass of Proteobacteria. Nature 2001, 411, 948–950. [Google Scholar]
- Oelkers, H.E.; Cole, R.D. Carbon Dioxide Sequestration A Solution to a Global Problem. Elements 2008, 4, 305–310. [Google Scholar] [CrossRef]
- Poschenrieder, C.; Fernández, J.A.; Rubio, L.; Pérez, L.; Terés, J.; Barceló, J. Transport and Use of Bicarbonate in Plants: Current Knowledge and Challenges Ahead. Int. J. Mol. Sci. 2018, 19, 1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruk, C.; Mazzeo, N.; Lacerot, G.; Reynolds, C.S. Classification schemes for phytoplankton: A local validation of a functional approach to the analysis of species temporal replacement. J. Plankton Res. 2002, 24, 901–912. [Google Scholar] [CrossRef]
- Brierley, A.S. Plankton. Curr. Biol. 2017, 27, R478–R483. [Google Scholar] [CrossRef] [PubMed]
- Sethi, D.; Butler, T.O.; Shuhaili, F.; Vaidyanathan, S. Diatoms for Carbon Sequestration and Bio-Based Manufacturing. Biology 2020, 9, 217. [Google Scholar] [CrossRef]
- Benedetti, M.; Vecchi, V.; Barera, S.; Dall’Osto, L. Biomass from microalgae: The potential of domestication towards sustainable biofactories. Microb. Cell Factories 2018, 17, 173. [Google Scholar] [CrossRef] [Green Version]
- Giordano, M.; Beardall, J.; Raven, J.A. CO2 concentrating mechanisms in algae: Mechanisms, environmental modulation, and evolution. Annu. Rev. Plant. Biol. 2005, 56, 99–131. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Factories 2018, 17, 36. [Google Scholar] [CrossRef]
- Mona, S.; Malyan, S.K.; Saini, N.; Deepak, B.; Pugazhendhi, A.; Kumar, S.S. Chemosphere Towards sustainable agriculture with carbon sequestration, and greenhouse gas mitigation using algal biochar. Chemosphere 2021, 275, 129856. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, W.; Wang, J.; Chen, Y.; Shen, S.; Liu, T. Bioresource Technology Utilization of simulated flue gas for cultivation of Scenedesmus dimorphus. Bioresour. Technol. 2013, 128, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.; Salois, P.; Markovic, P.; Hastings, J.W. A nuclear-encoded form II RuBisCO in dinoflagellates. Science 1995, 268, 1622–1624. [Google Scholar] [CrossRef] [PubMed]
- Whitney, S.M.; Andrews, T.J. The CO2 specificity of single-subunit ribulose-bisphosphate carboxylase from the dinoflagellate. Funct. Plant Biol. 1998, 25, 131–138. [Google Scholar] [CrossRef]
- Lapointe, M.; Mackenzie, T.D.; Morse, D. An external delta-carbonic anhydrase in a free-living marine dinoflagellate may circumvent diffusion-limited carbon acquisition. Plant Physiol. 2008, 147, 1427–1436. [Google Scholar] [CrossRef] [Green Version]
- Nimmo, I.C.; Barbrook, A.C.; Lassadi, I.; Chen, J.E.; Geisler, K.; Smith, A.G.; Aranda, M.; Purton, S.; Waller, R.F.; Nisbet, R.E.R.; et al. Genetic transformation of the dinoflagellate chloroplast. eLife 2019, 8, e45292. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef]
- Spalding, M.D.; Brown, B.E. Warm-water coral reefs and climate change. Science 2015, 350, 769–771. [Google Scholar] [CrossRef]
- Slavov, C.; Schrameyer, V.; Reus, M.; Ralph, P.J.; Hill, R.; Büchel, C.; Larkum, A.W.; Holzwarth, A.R. “Super-quenching” state protects Symbiodinium from thermal stress—Implications for coral bleaching. Biochim. Biophys. Acta 2016, 1857, 840–847. [Google Scholar] [CrossRef]
- Rehman, A.U.; Szabó, M.; Deák, Z.; Sass, L.; Larkum, A.; Ralph, P.; Vass, I. Symbiodinium sp. cells produce light-induced intra- and extracellular singlet oxygen, which mediates photodamage of the photosynthetic apparatus and has the potential to interact with the animal host in coral symbiosis. New Phytol. 2016, 212, 472–484. [Google Scholar] [CrossRef] [Green Version]
- Warner, M.E.; Fitt, W.K.; Schmidt, G.W. Damage to photosystem II in symbiotic dinoflagellates: A determinant of coral bleaching. Proc. Natl. Acad. Sci. USA 1999, 96, 8007–8012. [Google Scholar] [CrossRef] [Green Version]
- Howe, C.J.; Nisbet, R.E.; Barbrook, A.C. The remarkable chloroplast genome of dinoflagellates. J. Exp. Bot. 2008, 59, 1035–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Te, M.R.; Lohuis; Miller, D.J. Genetic transformation of dinoflagellates (Amphidinium and Symbiodinium): Expression of GUS in microalgae using heterologous promoter constructs. Plant J. 1998, 13, 427–435. [Google Scholar] [CrossRef]
- Walker, T.L.; Collet, C.; Purton, S. Algal transgenics in the genomic era1. J. Phycol. 2005, 41, 1077–1093. [Google Scholar] [CrossRef]
- Ortiz-Matamoros, M.F.; Villanueva, M.A.; Islas-Flores, T. Transient transformation of cultured photosynthetic dinoflagellates (Symbiodinium spp.) with plant-targeted vectors. Cienc. Mar. 2015, 41, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Jensen, E.L.; Yangüez, K.; Carrière, F.; Gontero, B. Storage Compound Accumulation in Diatoms as Response to Elevated CO2 Concentration. Biology 2019, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Burkhardt, S.; Amoroso, G.; Riebesell, U.; Sültemeyer, D. CO2 and HCO3− uptake in marine diatoms acclimated to different CO2 concentrations. Limnol. Oceanogr. 2001, 46, 1378–1391. [Google Scholar] [CrossRef] [Green Version]
- Clement, R.; Jensen, E.; Prioretti, L.; Maberly, S.C.; Gontero, B. Diversity of CO2-concentrating mechanisms and responses to CO2 concentration in marine and freshwater diatoms. J. Exp. Bot. 2017, 68, 3925–3935. [Google Scholar] [CrossRef]
- Granum, E.; Raven, J.A.; Leegood, R.C. How do marine diatoms fix 10 billion tonnes of inorganic carbon per year? Can. J. Bot. 2005, 83, 898–908. [Google Scholar] [CrossRef]
- Hopkinson, B.M.; Dupont, C.L.; Matsuda, Y. The physiology and genetics of CO2 concentrating mechanisms in model diatoms. Curr. Opin. Plant Biol. 2016, 31, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhou, Z.; Li, Y.; Wang, Y.; Xu, M.; Zhou, B.; Lu, K.; Wang, Y. The Bloom-Forming Dinoflagellate Karenia mikimotoi Adopts Different Growth Modes When Exposed to Short or Long Period of Seawater Acidification. Toxins 2021, 13, 629. [Google Scholar] [CrossRef]
- Pierella Karlusich, J.J.; Bowler, C.; Biswas, H. Carbon Dioxide Concentration Mechanisms in Natural Populations of Marine Diatoms: Insights from Tara Oceans. Front. Plant Sci. 2021, 12, 657821. [Google Scholar] [CrossRef] [PubMed]
- Flynn, K.J.; Mitra, A.; Greenwell, H.C.; Sui, J. Monster potential meets potential monster: Pros and cons of deploying genetically modified microalgae for biofuels production. Interface Focus 2013, 3, 20120037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothschild, L.J. The evolution of photosynthesis…again? Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 2787–2801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farquhar, G.D. Models describing the kinetics of ribulose biphosphate carboxylase-oxygenase. Arch. Biochem. Biophys. 1979, 193, 456–468. [Google Scholar] [CrossRef]
- Farquhar, J.; Zerkle, A.L.; Bekker, A. Geological constraints on the origin of oxygenic photosynthesis. Photosynth. Res. 2011, 107, 11–36. [Google Scholar] [CrossRef]
- Berg, I.A. Ecological aspects of the distribution of different autotrophic CO2 fixation pathways. Appl. Environ. Microbiol. 2011, 77, 1925–1936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar-Even, A.; Noor, E.; Milo, R. A survey of carbon fixation pathways through a quantitative lens. J. Exp. Bot. 2012, 63, 2325–2342. [Google Scholar] [CrossRef] [Green Version]
- Iñiguez, C.; Capó-Bauçà, S.; Niinemets, Ü.; Stoll, H.; Aguiló-Nicolau, P.; Galmés, J. Evolutionary trends in RuBisCO kinetics and their co-evolution with CO2 concentrating mechanisms. Plant J. 2020, 101, 897–918. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Ramya RC, S.; Mueller-Cajar, O. Surveying the expanding prokaryotic Rubisco multiverse. FEMS Microbiol. Lett. 2017, 364, fnx156. [Google Scholar] [CrossRef] [Green Version]
- Tabita, F.R.; Satagopan, S.; Hanson, T.E.; Kreel, N.E.; Scott, S.S. Distinct form I, II, III, and IV Rubisco proteins from the three kingdoms of life provide clues about Rubisco evolution and structure/function relationships. J. Exp. Bot. 2008, 59, 1515–1524. [Google Scholar] [CrossRef]
- Lee, J.W.; Mets, L.; Greenbau, E. Improvement of photosynthetic CO2 fixation at high light intensity through reduction of chlorophyll antenna size. Appl. Biochem. Biotechnol. 2002, 98–100, 37–48. [Google Scholar] [CrossRef]
- Lefebvre, S.; Lawson, T.; Fryer, M.; Zakhleniuk, O.V.; Lloyd, J.C.; Raines, C.A. Increased sedoheptulose-1, 7-bisphosphatase activity in transgenic tobacco plants stimulates photosynthesis and growth from an early stage in development. Plant Physiol. 2005, 138, 451–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sage, R.F. Variation in the k cat of Rubisco in C3 and C4 plants and some implications for photosynthetic performance at high and low temperature. J. Exp. Bot. 2002, 53, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Kebeish, R.; Niessen, M.; Thiruveedhi, K.; Bari, R.; Hirsch, H.-J.; Rosenkranz, R.; Stäbler, N.; Schönfeld, B.; Kreuzaler, F.; Peterhänsel, C. Chloroplastic photorespiratory bypass increases photosynthesis and biomass production in Arabidopsis thaliana. Nat. Biotechnol. 2007, 25, 593–599. [Google Scholar] [CrossRef]
- Raines, C.A. Transgenic approaches to manipulate the environmental responses of the C3 carbon fixation cycle. Plant Cell Environ. 2006, 29, 331–339. [Google Scholar] [CrossRef]
- Tcherkez, G.G.; Farquhar, G.D.; Andrews, T.J. Despite slow catalysis and confused substrate specificity, all ribulose bisphosphate carboxylases may be nearly perfectly optimized. Proc. Natl. Acad. Sci. USA 2006, 103, 7246–7251. [Google Scholar] [CrossRef] [Green Version]
- Kapralov, M.V.; Filatov, D.A. Widespread positive selection in the photosynthetic Rubisco enzyme. BMC Evol. Biol. 2007, 7, 73. [Google Scholar] [CrossRef] [Green Version]
- Ragsdale, S.W.; Pierce, E. Acetogenesis and the Wood-Ljungdahl pathway of CO2 fixation. Biochim. Biophys. Acta 2008, 1784, 1873–1898. [Google Scholar] [CrossRef] [Green Version]
- Savir, Y.; Noor, E.; Milo, R.; Tlusty, T. Cross-species analysis traces adaptation of Rubisco toward optimality in a low-dimensional landscape. Proc. Natl. Acad. Sci. USA 2010, 107, 3475–3480. [Google Scholar] [CrossRef] [Green Version]
- Evans, M.; Buchanan, B.B.; Arnon, D.I. A new ferredoxin-dependent carbon reduction cycle in a photosynthetic bacterium. Proc. Natl. Acad. Sci. USA 1966, 55, 928. [Google Scholar] [CrossRef] [Green Version]
- Huber, H.; Gallenberger, M.; Jahn, U.; Eylert, E.; Berg, I.A.; Kockelkorn, D.; Eisenreich, W.; Fuchs, G. A dicarboxylate/4-hydroxybutyrate autotrophic carbon assimilation cycle in the hyperthermophilic Archaeum Ignicoccus hospitalis. Proc. Natl. Acad. Sci. USA 2008, 105, 7851–7856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herter, S.; Fuchs, G.; Bacher, A.; Eisenreich, W. A bicyclic autotrophic CO2 fixation pathway in Chloroflexus aurantiacus. J. Biol. Chem. 2002, 277, 20277–20283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar-Even, A.; Noor, E.; Lewis, N.E.; Milo, R. Design and analysis of synthetic carbon fixation pathways. Proc. Natl. Acad. Sci. USA 2010, 107, 8889–8894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Papin, J.A.; Stelling, J.; Price, N.D.; Klamt, S.; Schuster, S.; Palsson, B.O. Comparison of network-based pathway analysis methods. Trends Biotechnol. 2004, 22, 400–405. [Google Scholar] [CrossRef]
- Schilling, C.H.; Letscher, D.; Palsson, B.O. Theory for the systemic definition of metabolic pathways and their use in interpreting metabolic function from a pathway-oriented perspective. J. Biol. 2000, 203, 229–248. [Google Scholar] [CrossRef] [Green Version]
- Schuster, S.; Dandekar, T.; Fell, D.A. Detection of elementary flux modes in biochemical networks: A promising tool for pathway analysis and metabolic engineering. Trends Biotechnol. 1999, 17, 53–60. [Google Scholar] [CrossRef]
- Lorsch, J.R. Practical Steady-State Enzyme Kinetics. In Methods in Enzymology; Elsevier: Amsterdam, The Netherland, 2014; pp. 3–15. [Google Scholar] [CrossRef]
- Manichaikul, A.; Ghamsari, L.; Hom, E.F.; Lin, C.; Murray, R.R.; Chang, R.L.; Balaji, S.; Hao, T.; Shen, Y.; Chavali, A.K.; et al. Metabolic network analysis integrated with transcript verification for sequenced genomes. Nat. Methods 2009, 6, 589–592. [Google Scholar] [CrossRef] [Green Version]
- Mittenthal, J.E.; Clarke, B.; Waddell, T.G.; Fawcett, G. A new method for assembling metabolic networks, with application to the Krebs citric acid cycle. J. Biol. 2001, 208, 361–382. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, X.; Sun, X.; Ma, Y. Shape-controlled synthesis of nanocarbons through direct conversion of carbon dioxide. Sci. Rep. 2013, 3, 3534. [Google Scholar] [CrossRef]
- Licht, S.; Douglas, A.; Ren, J.; Carter, R.; Lefler, M.; Pint, C.L. Carbon Nanotubes Produced from Ambient Carbon Dioxide for Environmentally Sustainable Lithium-Ion and Sodium-Ion Battery Anodes. ACS Cent. Sci. 2016, 2, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.M.; Lim, W.-G.; Kang, D.; Park, J.H.; Lee, H.; Lee, J.; Lee, J.W. Transformation of carbon dioxide into carbon nanotubes for enhanced ion transport and energy storage. Nanoscale 2020, 12, 7822–7833. [Google Scholar] [CrossRef] [PubMed]
- Lou, Z.; He, M.; Zhao, D.; Li, Z.; Shang, T. Synthesis of carbon nanorods by reduction of carbon bisulfide. J. Alloy. Compd. 2010, 507, 38–41. [Google Scholar] [CrossRef]
- Wu, H.; Li, Z.; Ji, D.; Liu, Y.; Li, L.; Yuan, D.; Zhang, Z.; Ren, J.; Lefler, M.; Wang, B.; et al. One-pot synthesis of nanostructured carbon materials from carbon dioxide via electrolysis in molten carbonate salts. Carbon 2016, 106, 208–217. [Google Scholar] [CrossRef] [Green Version]
- Lehman, J.H.; Terrones, M.; Mansfield, E.; Hurst, K.E.; Meunier, V. Evaluating the characteristics of multiwall carbon nanotubes. Carbon 2011, 49, 2581–2602. [Google Scholar] [CrossRef]
- DiLeo, R.A.; Landi, B.J.; Raffaelle, R.P. Purity assessment of multiwalled carbon nanotubes by Raman spectroscopy. J. Appl. Phys. 2007, 101, 064307. [Google Scholar] [CrossRef] [Green Version]
- Motiei, M.; Hacohen, Y.R.; Calderon-Moreno, J.; Gedanken, A. Preparing carbon nanotubes and nested fullerenes from supercritical CO2 by a chemical reaction. J. Am. Chem. Soc. 2001, 123, 8624–8625. [Google Scholar] [CrossRef]
- Challiwala, M.S.; Choudhury, H.A.; Wang, D.; El-Halwagi, M.M.; Weitz, E.; Elbashir, N.O. A novel CO2 utilization technology for the synergistic co-production of multi-walled carbon nanotubes and syngas. Sci. Rep. 2021, 11, 1417. [Google Scholar] [CrossRef]
- Daiyan, R.; Lovell, E.C.; Huang, B.; Zubair, M.; Leverett, J.; Zhang, Q.; Lim, S.; Horlyck, J.; Tang, J.; Lu, X.; et al. Uncovering Atomic-Scale Stability and Reactivity in Engineered Zinc Oxide Electrocatalysts for Controllable Syngas Production. Adv. Energy Mater. 2020, 10, 2001381. [Google Scholar] [CrossRef]
- Beyzavi, M.H.; Stephenson, C.J.; Liu, Y.; Karagiaridi, O.; Hupp, J.T.; Farha, O.K. Metal–Organic Framework-Based Catalysts: Chemical Fixation of CO2 with Epoxides Leading to Cyclic Organic Carbonates. Front. Energy Res. 2015, 2, 63. [Google Scholar] [CrossRef] [Green Version]
- Li, P.-Z.; Wang, X.-J.; Liu, J.; Phang, H.S.; Li, Y.; Zhao, Y. Highly Effective Carbon Fixation via Catalytic Conversion of CO2 by an Acylamide-Containing Metal–Organic Framework. Chem. Mater. 2017, 29, 9256–9261. [Google Scholar] [CrossRef]
- Tran, Y.B.N.; Nguyen, P.T.K.; Luong, Q.T.; Nguyen, K.D. Series of M-MOF-184 (M = Mg, Co, Ni, Zn, Cu, Fe) Metal–Organic Frameworks for Catalysis Cycloaddition of CO2. Inorg. Chem. 2020, 59, 16747–16759. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Cui, C.; Shi, C.; Wu, Z.S.; Yang, J.; Cheng, R.; Guang, T.; Wang, H.; Lu, H.; Wang, X. High-Energy-Density Hydrogen-Ion-Rocking-Chair Hybrid Supercapacitors Based on Ti3C2Tx MXene and Carbon Nanotubes Mediated by Redox Active Molecule. ACS Nano 2019, 13, 6899–6905. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.K.; Gaur, R.; Yadav, M.; Goswami, A.; Zbořil, R.; Gawande, M.B. An efficient copper-based magnetic nanocatalyst for the fixation of carbon dioxide at atmospheric pressure. Sci. Rep. 2019, 8, 1901. [Google Scholar] [CrossRef] [PubMed]
- Bhaduri, G.A.; Alamiry, M.A.H.; Šiller, L. Nickel Nanoparticles for Enhancing Carbon Capture. J. Nanomater. 2015, 2015, 581785. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Mangalapuri, R.; Ahmadi, M.H.; Vo, D.-V.N.; Solanki, R.; Kumar, P. The role of nanotechnology on post-combustion CO2 absorption in process industries. Int. J. Low-Carbon Technol. 2020, 15, 361–367. [Google Scholar] [CrossRef]
- Normile, D. Round and Round: A Guide to the Carbon Cycle. Science 2009, 325, 1642–1643. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceballos, G.; Ehrlich, P.R.; Raven, P.H. Vertebrates on the brink as indicators of biological annihilation and the sixth mass extinction. Proc. Natl. Acad. Sci. USA 2020, 117, 13596–13602. [Google Scholar] [CrossRef] [PubMed]
- Knoll, A.H.; Bambach, R.K.; Payne, J.L.; Pruss, S.; Fischer, W.W. Paleophysiology and end-Permian mass extinction. Earth Planet. Sci. Lett. 2007, 256, 295–313. [Google Scholar] [CrossRef]
- Ahmed, A.; Ahmed, Q.; Odelade, K.A. Microbial Inoculants for Improving Carbon Sequestration in Agroecosystems to Mitigate Climate Change Microbial Inoculants for Improving Carbon Sequestration in Agroecosystems to Mitigate Climate Change. In Handbook of Climate Change Resilience; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Gayathri, R.; Mahboob, S.; Govindarajan, M.; Al-ghanim, K.A.; Ahmed, Z.; Al-mulhm, N.; Vodovnik, M.; Vijayalakshmi, S. A review on biological carbon sequestration: A sustainable solution for a cleaner air environment, less pollution and lower health risks. J. King Saud Univ.-Sci. 2020, 33, 101282. [Google Scholar] [CrossRef]
- Fernandez, C.W.; Koide, R.T. Soil Biology & Biochemistry Initial melanin and nitrogen concentrations control the decomposition of ectomycorrhizal fungal litter. Soil Biol. Biochem. 2014, 77, 150–157. [Google Scholar] [CrossRef]
- Pal, A.; Pandey, S. Role of Glomalin in Improving Soil Fertility: A Review. Int. J. Plant Soil Sci. 2014, 3, 1112–1129. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-baracaldo, P.; Bianchini, G.; Wilson, J.D.; Knoll, A.H. Cyanobacteria and biogeochemical cycles through Earth history. Trends Microbiol. 2021, 30, 143–157. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of reverse tricarboxylic acid cycle.

Figure 2.
Schematic representation of reductive acetyl-CoA pathway.

Figure 3.
Schematic representation of dicarboxylate/4-hydroxybutyrate and 3-hydroxypropionate/4-hydroxybutyrate cycle. Green arrows indicate the dicarboxylate/4-hydroxybutyrate metabolic pathway, and blue arrows represent 3-hydroxypropionate/4-hydroxybutyrate cycle pathway. Black arrows represent common metabolic pathway between the two pathways.
Figure 3.
Schematic representation of dicarboxylate/4-hydroxybutyrate and 3-hydroxypropionate/4-hydroxybutyrate cycle. Green arrows indicate the dicarboxylate/4-hydroxybutyrate metabolic pathway, and blue arrows represent 3-hydroxypropionate/4-hydroxybutyrate cycle pathway. Black arrows represent common metabolic pathway between the two pathways.


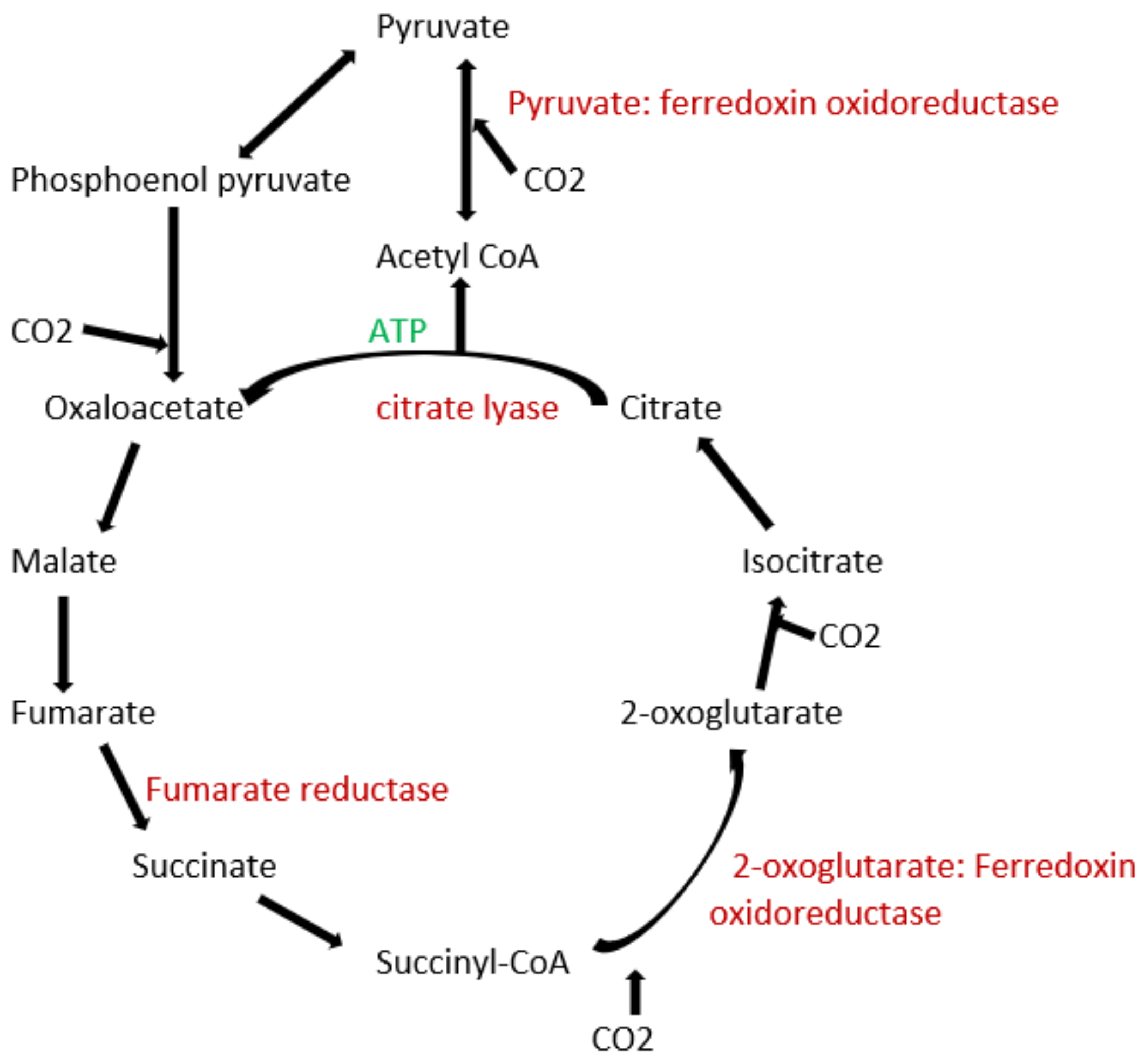{kind=link}
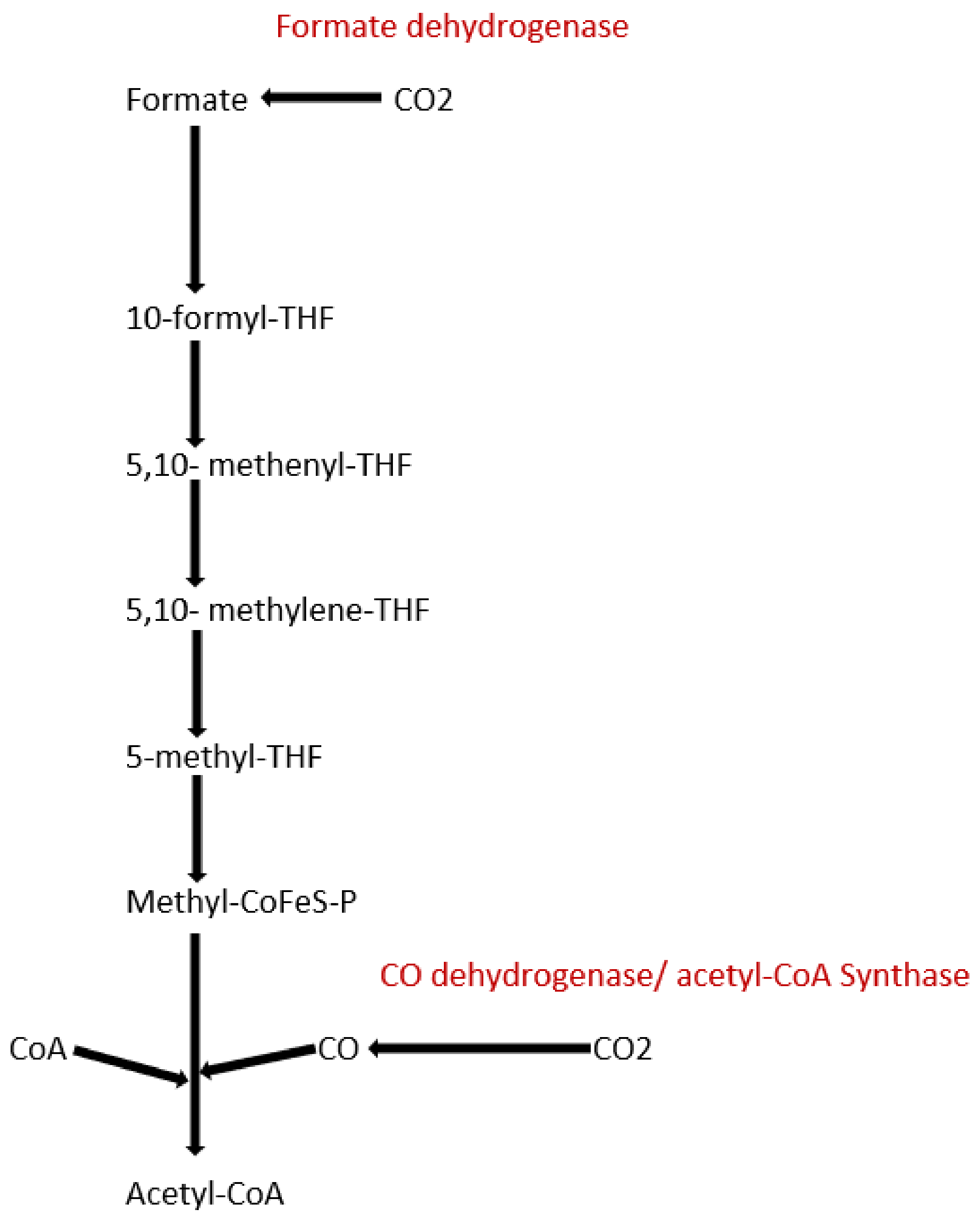{kind=link}
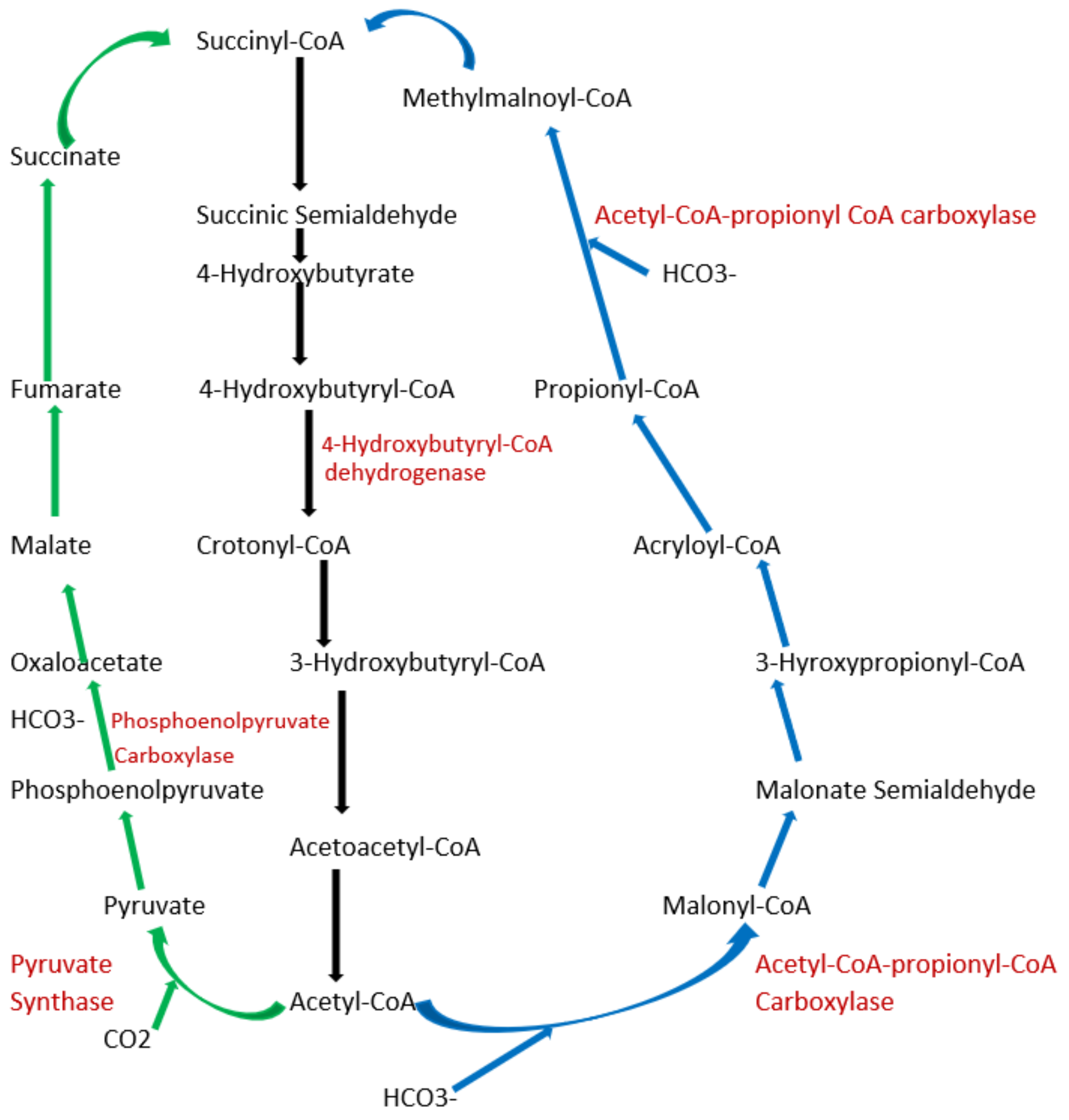{kind=link}
Table 1.
Comparative analysis of carbon fixation in bacterial species and algae.
| Microorganism | Description | Pathway | Advantage | Disadvantage | Reference |
|---|---|---|---|---|---|
| Methanogens | Prokaryotic, anaerobic, archaea | Reductive acetyl-CoA pathway | Efficient carbon capture, require low energy for carbon fixation, used for waste water treatment | Require specific conditions for growth, slow growth rate, oxygen-sensitive | [13,16] |
| Acetogens | Prokaryotic, anaerobic, bacteria | Reductive acetyl-CoA pathway | Efficient carbon capture, require low energy for fixation | Require specific conditions for growth, slow growth rate, oxygen-sensitive | [13,39] |
| Cyanobacteria | Prokaryotic, aerobic, bacteria | Calvin cycle | Carbon-concentrating mechanism present | Use inefficient RubisCO for carbon-fixation | [39,40] |
| Algae | Eukaryotic, single- or multi-cellular | Calvin cycle | Quick conversion of carbon for biomass and useful byproducts, may contain carbon-concentrating mechanisms, red algae contain better rubisco activases than other species. | Use inefficient RubisCO for carbon-fixation | [22,29,41] |
| Proteobacteria | Usually, Gram-negative bacteria, includes nitrogen-fixing bacteria, pathogenic | Reductive acetyl-CoA cycle, Calvin cycle | Different pathways across species, production of bioplastics and medicines as byproducts | Some require high pH, certain may use RubisCO | [35,42,43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ray, S.; Abraham, J.; Jordan, N.; Lindsay, M.; Chauhan, N. Synthetic, Photosynthetic, and Chemical Strategies to Enhance Carbon Dioxide Fixation. C 2022, 8, 18. https://0-doi-org.brum.beds.ac.uk/10.3390/c8010018
AMA Style
Ray S, Abraham J, Jordan N, Lindsay M, Chauhan N. Synthetic, Photosynthetic, and Chemical Strategies to Enhance Carbon Dioxide Fixation. C. 2022; 8(1):18. https://0-doi-org.brum.beds.ac.uk/10.3390/c8010018
Chicago/Turabian StyleRay, Supriyo, Jason Abraham, Nyiah Jordan, Mical Lindsay, and Neha Chauhan. 2022. "Synthetic, Photosynthetic, and Chemical Strategies to Enhance Carbon Dioxide Fixation" C 8, no. 1: 18. https://0-doi-org.brum.beds.ac.uk/10.3390/c8010018
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.