Ethanol Production from Cheese Whey and Expired Milk by the Brown Rot Fungus Neolentinus lepideus

Department of Chemistry and Biotechnology, Graduate School of Engineering, Tottori University, 4-101 Koyama, Tottori 680-8552, Japan
*
Author to whom correspondence should be addressed.
Fermentation 2019, 5(2), 49; https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation5020049
Submission received: 19 April 2019
/
Revised: 10 June 2019
/
Accepted: 13 June 2019
/
Published: 16 June 2019
(This article belongs to the Special Issue Microbial Production of Added-value Products from Renewable Resources 2.0)
Abstract
:The basidiomycete brown rot fungus Neolentinus lepideus is capable of assimilating and fermenting lactose to ethanol with a conversion yield comparable to those of lactose-fermenting yeasts. The ability of the fungus to ferment lactose is not influenced by the addition of glucose or calcium. Therefore, N. lepideus may be useful in ethanol production from materials composed mainly of lactose, such as cheese whey or expired cow’s milk. Whey is a by-product of cheese manufacturing, and approximately 50% of the total worldwide production of whey is normally disposed of without being utilized. We found that N. lepideus produced ethanol directly from cheese whey with a yield of 0.35 g of ethanol per gram of lactose consumed, and it also fermented expired milk containing lactose, protein, and fat with a similar yield. Our findings revealed that the naturally occurring basidiomycete fungus possesses a unique ability to produce ethanol from cheese whey and expired milk. Thus, N. lepideus may be useful in facilitating ethanol production from dairy wastes in a cost-effective and environmentally friendly manner.
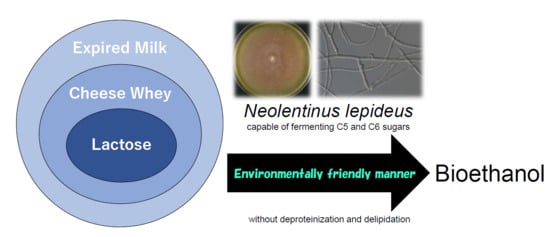
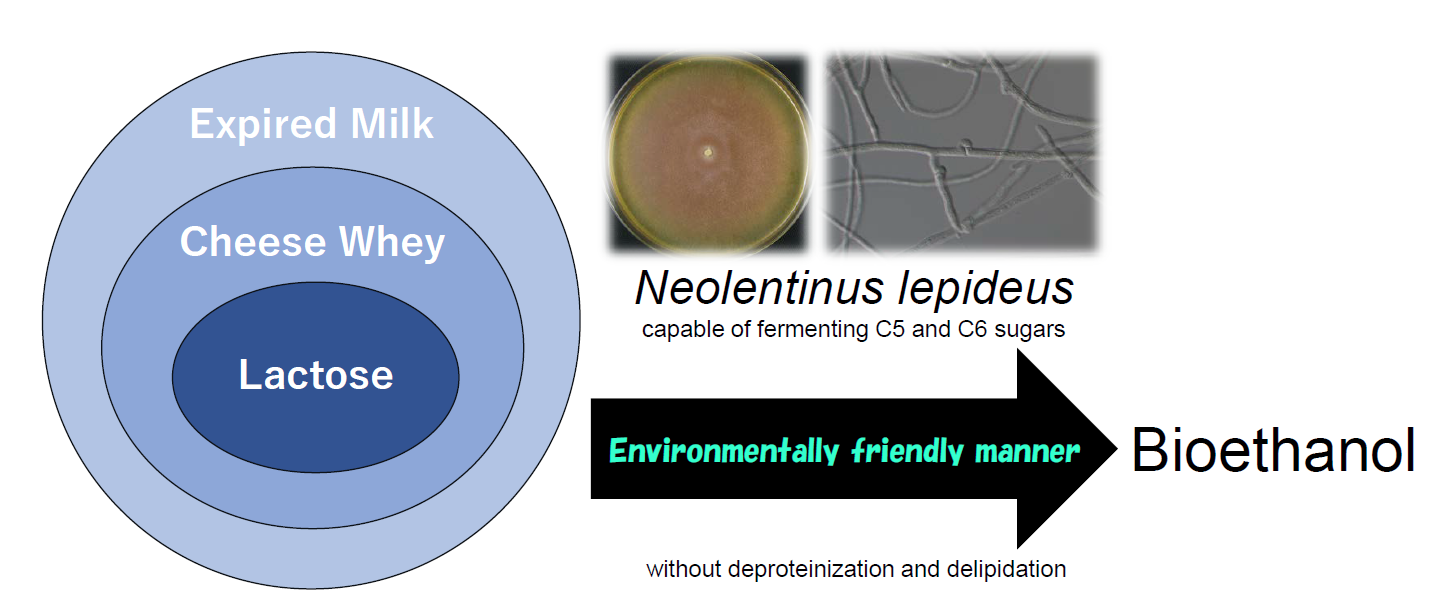
1. Introduction
The reduction of atmospheric carbon dioxide (CO2), which is a cause of global warming, involves the reduction of fossil fuel usage, being one of the main contributors for CO2, and is one of the most important global issues. From the viewpoint of reducing global greenhouse gas emissions, bioethanol has attracted attention in recent years as an alternative to fossil fuels. Plants, such as sugar cane and corn, which are raw materials for bioethanol, absorb CO2 from the atmosphere and grow, thus off-setting the CO2 released upon their burning. As it is a so-called carbon neutral fuel, the use of bioethanol is not counted in carbon dioxide emissions. Moreover, it can be used as a car fuel by mixing it with gasoline, and thus, it is now produced in many countries. Currently, there is a need to shift to ethanol production from lignocellulosic biomass, agricultural residues, and food waste as these renewable raw materials do not compete with food and do not affect the ecosystem [1,2].
Whey is the main by-product of cheese manufacturing. Generally, the production of 1 kg of cheese generates approximately 9 L of whey [3]. Whey is composed of lactose, proteins, minerals, vitamins, and fat. The worldwide production of whey was reported to be over 160 million tons per year with a 1–2% annual growth rate [4,5,6]. Whey, discarded as liquid waste, has a high biochemical oxygen demand and chemical oxygen demand due to the high volumes produced and its high content of organic matter, and it is a serious environmental problem. There are three types of whey that can be used as raw materials. First, plain cheese whey (a supernatant liquid generated in cheese production), whey permeate (protein in cheese whey is removed using an ultrafiltration membrane), and whey powder (cheese whey is dried and concentrated. Many studies have been performed on the conversion of whey to useful value-added chemicals, including ethanol and citric acid [7,8,9,10].
The main ingredient of whey, lactose, is a sugar derived from milk; it is a disaccharide consisting of galactose and glucose. Both β-galactosidase and lactose permease are indispensable for microorganisms to ferment lactose. Lactic acid bacteria, such as Lactobacillus, Lactococcus, and Leuconostoc, are the representative lactose-consuming microorganisms. They are able to convert lactose to lactic acid, but not to ethanol. However, a strain of the homo-lactic fermentative bacterium, Lactococcus lactis, has been genetically engineered to be able to efficiently produce ethanol from whey [11]. Similarly, Escherichia coli can assimilate lactose, but not ferment it, and some recombinant E. coli capable of fermenting whey have been constructed [12,13,14].
Some yeasts that have β-galactosidase and lactose permease are capable of directly metabolizing lactose. Among them, only a few yeasts, such as Kluyveromyces fragilis, Kluyveromyces marxianus, and Candida pseudotropicalis, can ferment lactose directly [15]. The representative yeast, Saccharomyces cerevisiae, is able to ferment glucose and galactose, but the wild strain cannot ferment lactose, because it lacks β-galactosidase activity [16], and pretreatment with acid or enzymatic hydrolysis is required for it to ferment lactose. Recombinant strains of S. cerevisiae with lactose-metabolizing genes have been constructed; these strains can consume and ferment lactose [17,18,19,20]. In addition, there have been various studies on ethanol production from various kinds of whey by Kluyveromyces spp., C. pseudotropicalis, and recombinant S. cerevisiae [6,21,22,23,24,25,26].
Most filamentous fungi have a low assimilative capacity for lactose. In fungi, there are two principal pathways for the catabolism of lactose. One pathway is observed in Hypocrea jecorina (Trichoderma reesei) and Aspergillus niger; they secrete β-galactosidase, which hydrolyzes lactose in the extracellular medium, and the resulting glucose and galactose monomers are transported into the cells by glucose and galactose permeases. The other pathway is observed in Aspergillus nidulans, Neurospora crassa, and Fusarium graminearum, in which lactose is transported into the cells by lactose permease, and is hydrolyzed intracellularly by β-galactosidase [27]. However, there have been no reports on fungi capable of fermenting lactose.
In our previous study, we isolated some basidiomycete fungi capable of producing ethanol from sugars, lignocellulosic biomass, and kitchen wastes [28,29,30,31,32,33]. The brown rot fungus Neolentinus lepideus could produce ethanol not only from xylose, but also from hexoses, including glucose, galactose, and lactose. In particular, among the ethanol-fermenting basidiomycete fungi, N. lepideus showed a superior ability to ferment lactose [31]. Therefore, we decided to examine the applicability of this fungus for ethanol production from cheese whey or expired cow’s milk. The aim of the present study was to characterize the ability of N. lepideus to ferment lactose, and to evaluate the potential of this fungus to efficiently produce bioethanol from dairy wastes.
2. Materials and Methods
2.1. Fungal Strain and Culture Conditions
The N. lepideus strain RS1911 used in this study has been reported before [31]. The fungus was initially grown on an MYG agar plate (10 g/L malt extract, 4 g/L yeast extract, 4 g/L glucose, and 15 g/L agar powder) at 28 °C for 7 days. Three 0.5 cm2 pieces of the mycelial mat were used to inoculate 50 mL of MYG liquid medium in 500 mL Erlenmeyer flasks and capped with silico plugs (Shin-Etsu Polymer Co., Ltd., Tokyo, Japan) for mycelial growth; the flasks were incubated for 7 days at 28 °C without shaking. The cultivated cells were inoculated into a 500-mL Erlenmeyer flask containing 50 mL of T medium (20 g/L lactose, 1 g/L yeast extract, 10 g/L KH2PO4, 2 g/L (NH4)2SO4, and 0.5 g/L MgSO4·7H2O) sealed with a silicon rubber plug (As One Co., Ltd., Osaka, Japan), and purged with nitrogen to confer an anaerobic condition for fermentation. The cultures were incubated at 28 °C without shaking. To investigate the fermentative characteristics of the fungus with dairy materials, whey or milk without any nutrient supplementation was used instead of the T medium.
2.2. Analytical Methods
At periodic intervals during fermentation, 1 mL samples were collected, centrifuged for 10 min at 15,000× g in an Eppendorf benchtop centrifuge, and then filtered through a 0.22 μm membrane filter (Millex-GP; Millipore Corp., Billerica, MA, USA). The ethanol and lactose concentrations in the culture filtrates were determined using a high-performance liquid chromatography system (Shimadzu Co., Ltd., Kyoto, Japan) equipped with a refractive index detector, RID-10A, and a Shodex KS-801 column (8.0 mm × 300 mm; Showa Denko Co., Ltd., Tokyo, Japan), and operated at 80 °C with a distilled water mobile phase at a flow rate of 0.6 mL/min. The theoretical yield of ethanol was defined as 0.538 g of ethanol per gram of glucose (4 mol of ethanol per mole of lactose).
3. Results and Discussion
3.1. Effect of the Lactose Concentration on Fermentation
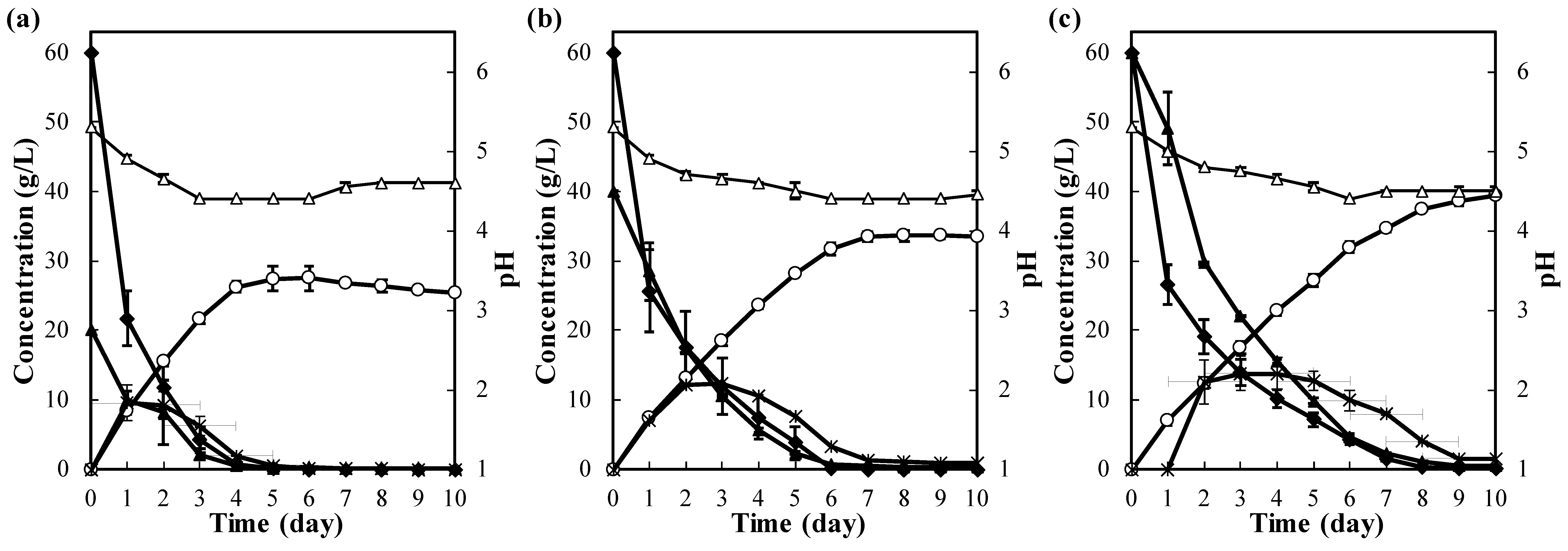
The effect of lactose concentration on fermentation by N. lepideus was investigated. As shown in Figure 1, 20, 60, and 100 g/L of lactose, as the sole carbon source, were consumed completely after 2, 4, and 6 days of incubation, respectively. No remarkable accumulation of galactose or glucose was detected in the culture filtrate at this point. Since no lactic acid was detected during cultivation, the pH of the culture filtrate did not change much. From 20, 60, and 100 g/L of lactose, the observed maximum ethanol concentrations were 7.6, 20.7, and 31.5 g/L, respectively, resulting in ethanol yields of 0.38, 0.35, and 0.32 g per gram of sugar utilized, respectively. The ethanol yield of this naturally occurring basidiomycete fungus was equivalent to those of K. fragilis (0.35 g/g), recombinant S. cerevisiae (0.33 g/g), and recombinant E. coli (0.38 g/g) [14,15,18]. This indicated that among naturally occurring microorganisms, strain RS1911 had a relatively high ability to ferment lactose. Although lactase (β-galactosidase) from ascomycete fungi, such as the genus Aspergillus or Penicillium, are well known [15,34,35,36,37], little information is available for basidiomycete fungi. Lactose does not exist in regular mushroom habitats in the first place, so it is presumed that lactase is not particularly necessary for growth. Fermentation of the lactose medium here suggested the presence of the same enzyme. Transcriptome analysis of the fermentation process showed increased transcription of the putative β-galactosidase gene (data not shown).
3.2. Effect of Glucose on Lactose Fermentation
Kluyveromyces yeasts are able to ferment lactose, but catabolite repression indicated that glucose is the preferred carbon source when available in the culture medium [38]. We investigated the effect of glucose on the fermentation by N. lepideus RS1911 by adding 20, 40, and 60 g/L of glucose to T medium containing 60 g/L lactose. We found that in the media with 20, 40, and 60 g/L of glucose, N. lepideus RS1911 consumed all of the lactose in 6, 6, and 9 days, respectively, yielding maximum ethanol concentrations of 27.5, 33.6, and 39.4 g/L, respectively. This corresponded to yields of 0.34, 0.33, and 0.33 g of ethanol per gram of consumed sugar, respectively (Figure 2). No significant catabolite repression was observed as N. lepideus simultaneously fermented both lactose and glucose in the media, although the consumption of lactose decreased when the concentration of glucose increased.
3.3. Effect of Calcium on Lactose Fermentation
In general, 100 mL of milk contains approximately 110 mg of calcium. The effect of calcium on the fermentative ability of N. lepideus was investigated. As shown in Figure 3, the addition of 500, 750, and 1000 mg of CaCl2 (final concentration) to T medium containing 60 g/L of lactose had no influence on lactose consumption or fermentation as compared to the levels in calcium-free medium (Figure 1b). Regardless of the concentration of added calcium, the maximum ethanol concentration was approximately 21 g/L, resulting in an ethanol yield of 0.35 g per gram of lactose. Thus, it turned out that basidiomycete fungi’s fermentation ability is not affected by the presence of calcium. In general, 0.1% or more of calcium is present in trees [39], so it is considered that, for the fungi growing there, fermentation may not be particularly affected under the same calcium concentration. Therefore, this indicates that strain RS1911 has an ability to ferment materials containing calcium, such as whey and milk.
3.4. Direct Ethanol Production from Cheese Whey
The cheese whey from cow’s milk used in this study comprised 5.4% lactose, 0.9% protein, and 0.3% fat. There have been no reports of cultivating basidiomycete fungi with cheese whey. The direct production of ethanol by N. lepideus RS1911 from whey, whey diluted 2-fold, and whey condensed 2-fold, was investigated. N. lepideus RS1911 consumed all of the lactose in whey, whey diluted 2-fold, and whey condensed 2-fold in 4, 2, and 8 days, respectively, yielding maximum ethanol concentrations of 16.9, 8.7, and 32.6 g/L, respectively, which corresponded to yields of 0.35, 0.38, and 0.32 g of ethanol per gram of lactose, respectively (Figure 4). A small amount of galactose and glucose liberated from lactose was found and were completely consumed in each whey sample. As a result, the ethanol yield per gram of lactose in each whey sample was similar to that in T medium containing lactose (Figure 1). This showed that regardless of the concentration of the whey, the fungus was capable of converting the lactose in the whey to ethanol with a similar yield. Although it has been shown that deproteinized whey is more likely to be suitable for fermentation than raw whey in performing the bioconversion of cheese whey to ethanol [6,7,8,9], it has been suggested that N. lepideus RS1911 has different properties from these. Previous studies have reported that the ethanol yields from whey without any additional nutrient supplements were 0.44, 0.37, and 0.32 g per gram of lactose with recombinant bacterium E. coli, recombinant yeast S. cerevisiae, and wild yeast K. marxianus, respectively [12,16,24]. No accumulation of lactic acid was detected during fermentation. Therefore, the decrease in pH is considered to be due to the generation of CO2 associated with ethanol production. The present experiment showed that the basidiomycete fungus N. lepideus can ferment whey similar to the lactose-fermenting yeasts. However, as shown in Table 1, ethanol productivity from whey by this fungus is lower than K. marxianus with similar ethanol yields. In order to improve the ethanol productivity using this strain, it will be necessary to further investigate optimizing fermentation conditions such as pH value, temperature, rate of aeration, inoculum size, and supplementation of nitrogen source.
3.5. Direct Ethanol Production from Whole Milk, High-Fat Milk, and Low-Fat Milk
Most investigations on ethanol production from cheese whey, such as whey permeate or whey powder, by recombinant S. cerevisiae or recombinant bacteria have reported relatively high ethanol yields. However, to our knowledge, there has been no report on direct ethanol production from milk containing protein and fat, which can affect fermentation. To further explore the fermentative properties of N. lepideus RS1911, we investigated the direct fermentation of expired whole milk, high-fat milk, and low-fat milk, all of which contain more fat and protein than whey. N. lepideus RS1911 consumed all the lactose in whole milk, low-fat milk, and high-fat milk after 6 days of incubation, yielding maximum ethanol concentrations of 16.2, 16.9, and 18.3 g/L, respectively, which corresponded to yields of 0.34, 0.34, and 0.34 g of ethanol per gram of lactose, respectively (Figure 5). Small amounts of liberated glucose and galactose were detected in the culture medium; this was attributed to the rapid uptake and conversion of the milk. The pH did not change because no significant amount of lactic acid was detected during fermentation. We estimated that N. lepideus RS1911 was able to directly convert milk to ethanol with a theoretical yield of approximately 63%. Whole cow milk comprises 4.8% lactose, 3.3% protein, and 3.8% fat. Low-fat milk comprises 5.0% lactose, 3.3% protein, and 2.1% fat. High-fat milk comprises 5.5% lactose, 3.9% protein, and 4.6% fat. The amounts of ethanol produced depended on the original lactose concentration, and not on the fat concentration. The ethanol yield per gram of lactose was equivalent to that in the T medium containing 6% lactose (Figure 1b) or in whey (Figure 4a). The fat and protein levels in the milk did not appear to affect the ethanol conversion by the fungus. Until now, studies on bioethanol production from dairy waste have used whey as their raw material [6,11,26], and there is no example of using milk as the main ingredient. There is an example of ethanol production using skim milk, but it was not the main raw material and instead it was used as a supplement on cane molasses fermentation [40]. Results here showed that N. lepideus was capable of converting lactose in milk containing fat and protein to ethanol with a relatively good yield. As the fermentation progressed, it was observed that the culture fluid gradually clarified. This is believed to be due to the decomposition of the milk protein by N. lepideus during the fermentation. As described above, when whey was used as a raw material, ethanol productivity was lower than other microorganisms, but the ability of this basidiomycete fungus to convert milk directly to ethanol without pretreatment is considered to be very interesting. To our knowledge, little evidence exists for direct ethanol production from milk by naturally occurring microorganisms. The present study is the first to report that a basidiomycete fungus, which has no relation to dairy products in nature, can ferment dairy wastes, such as whey and expired milk, without the need for any pre-treatment steps, such as deproteinization or defatting. Thus, our present findings indicate that N. lepideus RS1911 is a microbe that can potentially act as a biocatalyst in the fermentation of dairy wastes or unutilized resources to ethanol.
4. Conclusion
This is the first study to report the ability of the brown rot fungus N. lepideus to efficiently ferment whey and milk. The fungi are not only capable of assimilating carbon sources in woods, but also have the beneficial ability to ferment dairy products, despite them having no relation to the living environment of basidiomycete fungi. Thus, N. lepideus appears to be a promising microorganism for the production of bioethanol from various biomass materials. Further studies are required to optimize the bioethanol yield and productivity of this microorganism. The direct conversion of ethanol from these materials by the naturally occurring basidiomycete fungus would be advantageous for bioethanol production by consolidated bioprocessing because it would cost less, have a lower environmental footprint, and not require the use of hazardous chemicals.
Author Contributions
K.O. conceived, designed, analyzed the data, and wrote the manuscript. S.N., R.K., and S.K. carried out the experimental work and contributed with valuable discussions.
Funding
This study was supported in part by a Grant-in-Aid for Scientific Research (No. 24510099) from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
Acknowledgments
The authors are thankful to Tomohisa Shimoda, Daisen Nyugyo Agricultural Cooperative Association, for providing the cheese whey.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zabed, H.; Sahu, J.N.; Suely, A.; Boyce, A.N.; Faruq, G. Bioethanol production from renewable sources: Current perspectives and technological progress. Renew. Sustain. Energy Rev. 2017, 71, 475–501. [Google Scholar] [CrossRef]
- Huang, H.; Qureshi, N.; Chen, M.-H.; Liu, W.; Singh, V. Ethanol production from food waste at high solids content with vacuum recovery technology. J. Agric. Food Chem. 2015, 63, 2760–2766. [Google Scholar] [CrossRef] [PubMed]
- Kosikowski, F.V. Whey utilization and whey products. J. Dairy Sci. 1979, 62, 1149–1160. [Google Scholar] [CrossRef]
- Smithers, G.W. Whey and whey proteins—from ‘gutter-to-gold’. Int. Dairy J. 2008, 18, 695–704. [Google Scholar] [CrossRef]
- Guimarães, P.M.R.; Teixeira, J.A.; Domingues, L. Fermentation of lactose to bio-ethanol by yeasts as part of integrated solutions for the valorisation of cheese whey. Biotechnol. Adv. 2010, 28, 375–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murani, C.S.; da Silva, D.C.M.N.; Schuina, G.L.; Mosinahti, E.F.; Del Bianchi, V.L. Bioethanol production from dairy industrial coproducts. Bioenergy Res. 2019, 12, 112–122. [Google Scholar]
- Prazeres, A.R.; Carvalho, F.; Rivas, J. Cheese whey management: A review. J. Environ. Manag. 2012, 110, 48–68. [Google Scholar] [CrossRef]
- Božanić, R.; Barukčić, I.; Jakopović, K.L.; Tratnik, L. Possibilities of whey utilization. Austin J. Nutr. Food Sci. 2014, 2, 1036–1042. [Google Scholar]
- Pescuma, M.; de Valdez, G.F.; Mozzi, F. Whey-derived valuable products obtained by microbial fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 6183–6196. [Google Scholar] [CrossRef]
- El-Samragy, Y.A.; Khorshid, M.A.; Foda, M.I.; Shehata, A.E. Effect of fermentation conditions on the production of citric acid from cheese whey by Aspergillus niger. Int. J. Food Microbiol. 1996, 29, 411–416. [Google Scholar] [CrossRef]
- Liu, J.; Dantoft, S.H.; Würtz, A.; Jensen, P.R.; Solem, C. A novel cell factory for efficient production of ethanol from dairy waste. Biotechnol. Biofuels 2016, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, W.V.; Dudey, G.L.; Ingram, L.O. Fermentation of sweet whey by ethanologenic Escherichia coli. Biotechnol. Bioeng. 1992, 40, 41–45. [Google Scholar] [CrossRef]
- Leite, A.R.; Guimaraes, W.V.; de Araujo, E.F.; Silva, D.O. Fermentation of sweet whey by recombinant Escherichia coli KO11. Braz. J. Microbiol. 2000, 31, 212–215. [Google Scholar] [CrossRef]
- Pasotti, L.; Zucca, S.; Casanova, M.; Micoli, G.; De Angelis, M.G.C.; Magni, P. Fermentation of lactose to ethanol in cheese whey permeate and concentrated permeate by engineered Escherichia coli. BMC Biotechnol. 2017, 17, 48. [Google Scholar] [CrossRef] [PubMed]
- Kamini, N.R.; Gunasekaran, P. Simultaneous ethanol production from lactose by Kluyveromyces fragilis and Zymomonas mobilis. Curr. Microbiol. 1987, 16, 153–157. [Google Scholar] [CrossRef]
- Guimarães, P.M.R.; François, J.; Parrou, J.L.; Teixeira, J.A.; Domingues, L. Adaptive evolution of a lactose-consuming Saccharomyces cerevisiae recombinant. Appl. Environ. Microbiol. 2008, 74, 1748–1756. [Google Scholar] [CrossRef]
- Kumar, V.; Ramakrishnan, S.; Teeri, T.T.; Knowles, J.K.C.; Hartley, B.S. Saccharomyces cerevisiae cells secreting an Aspergillus niger β-galactosidase grow on whey permeate. Bio/Technology 1992, 10, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, S.; Hartley, B.S. Fermentation of lactose by yeast cells secreting recombinant fungal lactase. Appl. Environ. Microbiol. 1993, 59, 4230–4235. [Google Scholar] [Green Version]
- Domingues, L.; Dantas, M.M.; Lima, N.; Teixeira, J.A. Continuous ethanol fermentation of lactose by a recombinant flocculating Saccharomyces cerevisiae strain. Biotechnol. Bioeng. 1999, 64, 692–697. [Google Scholar] [CrossRef]
- Guimarães, P.M.R.; Teixeira, J.A.; Dominguez, L. Fermentation of high concentrations of lactose to ethanol by engineered flocculent Saccharomyces cerevisiae. Biotechnol. Lett. 2008, 30, 1953–1958. [Google Scholar] [CrossRef]
- Zafar, S.; Owais, M. Ethanol production from crude whey by Kluyveromyces marxianus. Biochem. Eng. J. 2006, 27, 295–298. [Google Scholar] [CrossRef]
- Ozmihci, S.; Kargi, F. Ethanol fermentation of cheese whey powder solution by repeated fed-batch operation. Enzym. Microb. Technol. 2007, 41, 169–174. [Google Scholar] [CrossRef]
- Sansonetti, S.; Curcio, S.; Calabrò, V.; Iorio, G. Bio-ethanol production by fermentation of ricotta cheese whey as an effective alternative non-vegetable source. Biomass Bioenergy 2009, 33, 1687–1692. [Google Scholar] [CrossRef]
- Guo, X.; Zhou, J.; Xiao, D. Improved ethanol production by mixed immobilized cells of Kluyveromyces marxianus and Saccharomyces cerevisiae from cheese whey powder solution fermentation. Appl. Biochem. Biotechnol. 2010, 160, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.D.; Kádár, Z.; Oleskowicz-Popiel, P.; Thomsen, M.H. Production of bioethanol from organic whey using Kluyveromyces marxianus. J. Ind. Microbiol. Biotechnol. 2011, 38, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Zoppellari, F.; Bardi, L. Production of bioethanol from effluents of the dairy industry by Kluyveromyces marxianus. New Biotechnol. 2013, 30, 607–613. [Google Scholar] [CrossRef]
- Seiboth, B.; Pakdaman, B.S.; Hartl, L.; Kubicek, C.P. Lactose metabolism in filamentous fungi: How to deal with an unknown substrate. Fungal Biol. Rev. 2007, 21, 42–48. [Google Scholar] [CrossRef]
- Okamoto, K.; Imashiro, K.; Akizawa, Y.; Onimura, A.; Yoneda, M.; Nitta, Y.; Maekawa, N.; Yanase, H. Production of ethanol by the white-rot basidiomycetes Peniophora cinerea and Trametes suaveolens. Biotechnol. Lett. 2010, 32, 909–913. [Google Scholar] [CrossRef]
- Okamoto, K.; Nitta, Y.; Maekawa, N.; Yanase, H. Direct ethanol production from starch, wheat bran and rice straw by the white rot fungus Trametes hirsuta. Enzym. Microb. Technol. 2011, 48, 273–277. [Google Scholar] [CrossRef]
- Okamoto, K.; Sugita, Y.; Nishikori, N.; Nitta, Y.; Yanase, H. Characterization of two acidic β-glucosidases and ethanol fermentation in the brown rot fungus Fomitopsis palustris. Enzym. Microb. Technol. 2011, 48, 359–364. [Google Scholar] [CrossRef]
- Okamoto, K.; Kanawaku, R.; Masumoto, M.; Yanase, H. Efficient xylose fermentation by the brown rot fungus Neolentinus lepideus. Enzym. Microb. Technol. 2012, 50, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Uchii, A.; Kanawaku, R.; Yanase, H. Bioconversion of xylose, hexoses and biomass to ethanol by a new isolate of the white rot basidiomycete Trametes versicolor. Springerplus 2014, 3, 121. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Yoneda, M.; Fumioka, T. Isolation of a Peniophora strain capable of producing ethanol from starch and kitchen waste. Ferment. Technol. 2017, 6, 1. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, F.; Hou, Z.; Yuan, C.; Zhu, X. High level production of β-galactosidase exhibiting excellent milk-lactose degradation ability from Aspergillus oryzae by codon and fermentation optimization. Appl. Biochem. Biotechnol. 2014, 172, 2787–2799. [Google Scholar] [CrossRef] [PubMed]
- Németh, Z.; Molnár, Á.P.; Fejes, B.; Novák, L.; Karaffa, L.; Keller, N.P.; Fekete, E. Growth-phase sterigmatocystin formation on lactose is mediated via low specific growth rates in Aspergillus nidulans. Toxins 2016, 8, 354. [Google Scholar] [CrossRef] [PubMed]
- Rojas, A.L.; Nagem, R.A.P.; Neustroev, K.N.; Arand, M.; Adamska, M.; Eneyskaya, E.V.; Kulminskaya, A.A.; Garratt, R.C.; Golubev, A.M.; Polikarpov, I. Crystal structures of β-galactosidase from Penicillium sp. and its complex with galactose. J. Mol. Biol. 2004, 343, 1281–1292. [Google Scholar] [CrossRef] [PubMed]
- Jónás, Á.; Fekete, E.; Flipphi, M.; Sándor, E.; Jäger, S.; Molnár, Á.P.; Szentirmai, A.; Karaffa, L. Extra- and intracellular lactose catabolism in Penicillium chrysogenum: Phylogenetic and expression analysis of the putative permease and hydrolase genes. J. Antibiot. 2014, 67, 489–497. [Google Scholar] [CrossRef]
- Rubio-Texeira, M. Endless versatility in the biotechnological applications of Kluyveromyces LAC genes. Biotechnol. Adv. 2006, 24, 212–225. [Google Scholar] [CrossRef]
- Ovington, J.D. The calcium and magnesium contents of tree species grown in close stand. New Phytol. 1959, 58, 164–175. [Google Scholar] [CrossRef]
- Patil, S.G.; Gokhale, D.V.; Patil, B.G. Enhancement in ethanol production from cane molasses by skim milk supplementation. Enzym. Microb. Technol. 1986, 8, 481–484. [Google Scholar] [CrossRef]
Figure 1.
Ethanol production from lactose by N. lepideus RS1911. Cells were cultured statically in T medium supplemented with 20 g/L (a), 60 g/L (b), or 100 g/L (c) of lactose. Symbols represent lactose (filled diamonds), liberated glucose (filled triangles), liberated galactose (asterisks), ethanol (open circles), and pH (open triangles). Error bars indicate the standard deviation of the mean from three independent experiments.
Figure 1.
Ethanol production from lactose by N. lepideus RS1911. Cells were cultured statically in T medium supplemented with 20 g/L (a), 60 g/L (b), or 100 g/L (c) of lactose. Symbols represent lactose (filled diamonds), liberated glucose (filled triangles), liberated galactose (asterisks), ethanol (open circles), and pH (open triangles). Error bars indicate the standard deviation of the mean from three independent experiments.

Figure 2.
Ethanol production from a mixture of lactose and glucose by N. lepideus RS1911. Cells were cultured statically in T medium supplemented with 20 g/L (a), 40 g/L (b), or 60 g/L (c) of glucose. Symbols represent lactose (filled diamonds), glucose (filled triangles), liberated galactose (asterisks), ethanol (open circles), and pH (open triangles). Error bars indicate the standard deviation of the mean from three independent experiments.
Figure 2.
Ethanol production from a mixture of lactose and glucose by N. lepideus RS1911. Cells were cultured statically in T medium supplemented with 20 g/L (a), 40 g/L (b), or 60 g/L (c) of glucose. Symbols represent lactose (filled diamonds), glucose (filled triangles), liberated galactose (asterisks), ethanol (open circles), and pH (open triangles). Error bars indicate the standard deviation of the mean from three independent experiments.

Figure 3.
Ethanol production from lactose supplemented with calcium by N. lepideus RS1911. Cells were cultured statically in T medium containing 60 g/L of lactose supplemented with 500 mg/L (a), 750 mg/L (b), or 1000 mg/L (c) of CaCl2. Symbols represent lactose (filled diamonds), glucose (filled triangles), liberated galactose (asterisks), ethanol (open circles), and pH (open triangles). Error bars indicate the standard deviation of the mean from three independent experiments.
Figure 3.
Ethanol production from lactose supplemented with calcium by N. lepideus RS1911. Cells were cultured statically in T medium containing 60 g/L of lactose supplemented with 500 mg/L (a), 750 mg/L (b), or 1000 mg/L (c) of CaCl2. Symbols represent lactose (filled diamonds), glucose (filled triangles), liberated galactose (asterisks), ethanol (open circles), and pH (open triangles). Error bars indicate the standard deviation of the mean from three independent experiments.

Figure 4.
Ethanol production from whey by N. lepideus RS1911. Cells were cultured statically in whey (a), whey diluted 2-fold (b), or whey condensed 2-fold (c). Symbols represent lactose (filled diamonds), liberated glucose (filled triangles), liberated galactose (asterisks), ethanol (open circles), and pH (open triangles). Error bars indicate the standard deviation of the mean from three independent experiments.
Figure 4.
Ethanol production from whey by N. lepideus RS1911. Cells were cultured statically in whey (a), whey diluted 2-fold (b), or whey condensed 2-fold (c). Symbols represent lactose (filled diamonds), liberated glucose (filled triangles), liberated galactose (asterisks), ethanol (open circles), and pH (open triangles). Error bars indicate the standard deviation of the mean from three independent experiments.

Figure 5.
Ethanol production from expired milk by N. lepideus RS1911. Cells were cultured statically in whole milk (a), low-fat milk (b), or high-fat milk (c). Symbols represent lactose (filled diamonds), liberated glucose (filled triangles), liberated galactose (asterisks), ethanol (open circles), and pH (open triangles). Error bars indicate the standard deviation of the mean from three independent experiments.
Figure 5.
Ethanol production from expired milk by N. lepideus RS1911. Cells were cultured statically in whole milk (a), low-fat milk (b), or high-fat milk (c). Symbols represent lactose (filled diamonds), liberated glucose (filled triangles), liberated galactose (asterisks), ethanol (open circles), and pH (open triangles). Error bars indicate the standard deviation of the mean from three independent experiments.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Comparison of ethanol production by microorganisms from cheese whey.
| Strain | Maximum Ethanol Concentration (g/L) | Fermentation Time (h) | Ethanol Yield (g/g) | Initial Lactose (g/L) | Ethanol Productivity (g/L/h) | Reference |
|---|---|---|---|---|---|---|
| N. lepideus RS1911 | 9 | 48 | 0.38 | 27 | 0.19 | This study |
| 17 | 96 | 0.35 | 54 | 0.18 | ||
| 33 | 192 | 0.32 | 108 | 0.17 | ||
| Recombinant E. coli | 26 | 144 | 0.44 | 60 | 0.18 | Guimaraes et al. [12] |
| Recombinant S. cerevisiae | 55 | 120 | 0.37 | 150 | 0.46 | Guimarães et al. [20] |
| K. marxianus | 30 | 96 | 0.32 | 100 | 0.31 | Guo et al. [24] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Okamoto, K.; Nakagawa, S.; Kanawaku, R.; Kawamura, S. Ethanol Production from Cheese Whey and Expired Milk by the Brown Rot Fungus Neolentinus lepideus. Fermentation 2019, 5, 49. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation5020049
AMA Style
Okamoto K, Nakagawa S, Kanawaku R, Kawamura S. Ethanol Production from Cheese Whey and Expired Milk by the Brown Rot Fungus Neolentinus lepideus. Fermentation. 2019; 5(2):49. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation5020049
Chicago/Turabian StyleOkamoto, Kenji, Saki Nakagawa, Ryuichi Kanawaku, and Sayo Kawamura. 2019. "Ethanol Production from Cheese Whey and Expired Milk by the Brown Rot Fungus Neolentinus lepideus" Fermentation 5, no. 2: 49. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation5020049
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.