Anti-Inflammatory Effects and Their Correlation with Microbial Community of Shindari, a Traditional Jeju Beverage

Jeju Inside Agency & Cosmetic Science Center, Department of Chemistry and Cosmetics, Jeju National University, Jeju 63243, Korea
*
Author to whom correspondence should be addressed.
Fermentation 2020, 6(3), 87; https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030087
Submission received: 15 August 2020
/
Revised: 2 September 2020
/
Accepted: 4 September 2020
/
Published: 7 September 2020
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:In this study, we investigated the microbial community and its correlation with anti-inflammatory effects during the fermentation procedure of shindari. Since shindari is traditionally manufactured through a fermentation process of 3–4 days in summer or 5–6 days in winter, the shindari materials applied in this study were fermented for 1, 3, and 5 days, respectively. Microbial community structure in the shindari manufacturing process was analyzed by Illumina MiSeq sequencing. Results revealed different fermentation time had different influences on the community structure and microbial diversity in the shindari process. Ultimately, we found that pathogenic bacteria such as Enterococcus durans (9.3%) and Enterobacter asburiae (4.5%) dominate the shindari fermented for 1 day, while samples with a fermentation period of more than 3 days occupied lactic acid bacteria such as Pediococcus sp. (97.1%). In addition, to study the relationship between fermentation period and anti-inflammatory effect in the shindari manufacturing process, we applied RAW264.7 cells, a classic cell model for inflammation studies. First, we prepared an ethyl acetate extract of shindari fermented for 1 (S1), 3 (S3), or 5 days (S5), and then it was confirmed that all of these extracts inhibited the nitric oxide (NO) production in a concentration-dependent manner. In addition, these inhibitory effects were correlated with the suppressive effect of shindari extracts against overexpression of inducible nitric oxide synthase (iNOS). Furthermore, S3 and S5 also inhibited the production of pro-inflammatory cytokines such as tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), and IL-6. Overall, the anti-inflammatory effect of S3 was suggested to be mediated through the negative regulation of mitogen-activated protein kinase signaling (MAPKs) and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling pathways.
1. Introduction
Jeju Island, a volcanic island, has little fertile farmland, and transportation to the mainland is inconvenient, so people have to rely on food ingredients produced on the island itself [1]. Food availability is a major problem, especially if the ship has been cut off for a long time on the mainland due to storms or famines, and for this reason, most of the rice and food ingredients purchased on the mainland are considered very valuable [2]. In addition, in the past, it was difficult to store food in hot and humid summer due to the lack of refrigeration for storing food, and the remaining food was often stored using fermentation or drying methods.
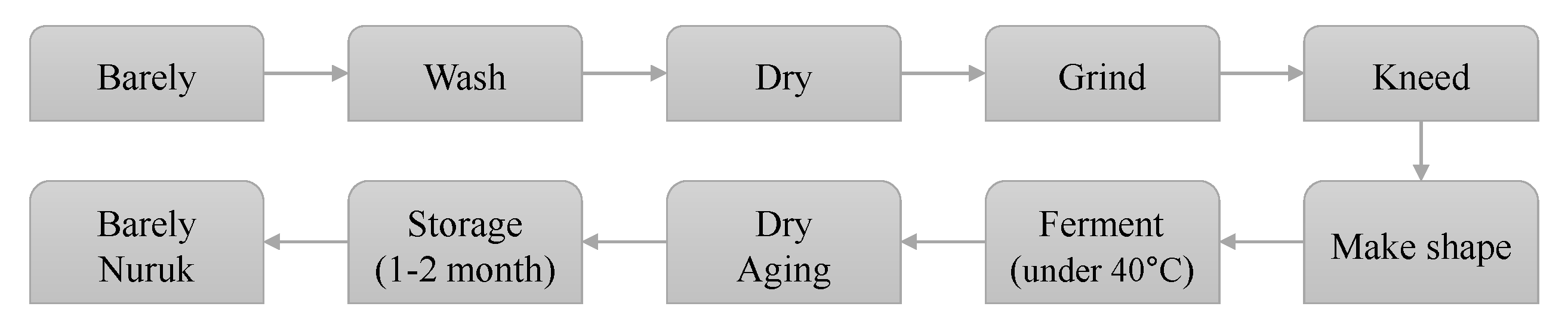
Shindari is a traditional fermented grain beverage called dansool (i.e., sweet liquor), enjoyed by indigenous people who lived in Jeju Island, located on the southernmost point of the Korean Peninsula. Shindari can be considered a Jeju yogurt or low alcohol wine. The word “Shin” means that the food is bad and “dari” means to boil it carefully. The barley nuruk, which is used as a starter in the manufacture of shindari, is made from cleaned and dried barley, finely ground using a millstone. Water is then added—equivalent to about 25% by volume. The mixture is kneaded and molded into a circular shape of the desired size. In other words, shindari is a traditional fermented food of Jeju native people who ferment and boil the food of the spoiled grains with barley nuruk, a fermenting starter. Figure 1 is a schematic diagram of the manufacturing process for the traditional barley nuruk commonly used by Jeju indigenous people [3].
Newly developed sequencing strategies have exploded in recent years, which are commonly referred to as next-generation sequencing (NGS) technologies. NGS technology has high throughput and produces thousands or millions of sequences at the same time. These sequences allow for the precise identification of microbial taxa, including non-culturing organisms and a small number of organisms. The development of NGS technology has enabled researchers to study and understand the microbial world from a broader and deeper perspective. Many studies have emphasized the use of NGS technology to investigate microbial communities in fermented foods, such as cheese, kimchi, and sausage. The technology is developing at a rapid pace, constantly improving quality and cost, and is having a major impact on food microbiology [4].
Nitric oxide synthase (iNOS) is an inflammatory condition that produces excessive amounts of NO (nitric oxide). Consequently, NO and iNOS can be important targets for the treatment of inflammatory diseases. On the other hand, nuclear factor kappa B (NF-κB) is a transcription factor that plays a fundamental role in inflammation and acute reactions. Generally, NF-κB subunits are inactive, bind to IκB, and phosphorylation of IκB activates NF-κB into the nucleus to activate gene expression. NF-κB is one of the main regulators of pre-inflammatory gene expression and mediates the synthesis of cytokines such as tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-6, and IL-8. It also regulates the transcription of other inflammatory mediators, such as cyclooxygenase (COX)-2. It also regulates the transcription of other inflammatory mediators, such as COX-2 or iNOS. Therefore, NF-κB is an essential target for the treatment of inflammatory diseases. MAPK is a protein chain that regulates NF-κB activation. These MAPKs are extracellular signal regulatory kinase 1/2 (ERK 1/2), c-Jun amino-terminal kinase (JNK) and [5,6,7] (p. 38).
This study aimed to analyze the microbial communities of shindari, a traditional Jeju beverage. Additionally, we tried to evaluate whether the microbial community generated during the fermentation process has anti-inflammatory efficacy.
2. Materials and Methods
2.1. Preparation of Extract
The rice used in this experiment was purchased from CJ CheilJedang Co., Ltd. (Seoul, Korea), and nuruk was purchased from Jeju Dongmun Market. After mixing 50 g of cooked rice and 50 g of barley nuruk in each culture container, 750 mL of water was added and left for 24 h, 72 h, or 120 h in a 30 °C incubator, instead of the traditional, 1–2 month, room temperature fermentation conditions. The three shindari samples (24, 72, and 120 h were filtered, and 99% ethanol was added to adjust the final concentration to 70%. The mixture was stirred at 25 °C for 24 h, extracted three times repeatedly, concentrated under reduced pressure with a rotary vacuum concentrator, and lyophilized in a freezer dryer.
2.2. Preparation of Ethyl Acetate (EtOAc) Fraction
After adding and mixing a predetermined amount of distilled water to the obtained shindari fermentation extract, EtOAc was added using a separatory funnel to extract the fraction. In the separatory funnel, equal amounts of distilled water and EtOAc were added, fractionated for 2 h, and shaken every 30 min. When the layers were separated, the extraction solvent was removed and the remaining filtrate was extracted three more times in the same manner, concentrated under reduced pressure with a rotary vacuum concentrator, and lyophilized in a freezer dryer. The resulting substances were named EtOAc fr. of 1-day shindari fermentation (S1), EtOAc fr. of 3-day shindari fermentation (S3), and EtOAc fr. of 5-day shindari fermentation (S5).
2.3. PCR Amplification and Sequencing
Total DNA was extracted from the shindari sample using a FastDNA® SPIN Kit for Soil (MP Biomedicals, Solon, OH, USA) according to the manufacturer’s instructions. To confirm the microbial diversity, DNA obtained from the sample was amplified using fusion primers that amplify the V3-V4 portion of the 16S rRNA gene. For bacterial amplification, the fusion primers 341F (5′-AATGATACGGCGACCACCGAGATCTACAC-XXXXXXXX-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG-CCTACGGGNGGCWGCAG-3′; underlining sequence indicates the target region primer) and 805R (5′-CAAGCAGAAGACGGCATACGAGAT-XXXXXXXX-GTCTCGTGGGCTCGG-AGATGTGTATAAGAGACAG GACTACHVGGGTATCTAATCC-3′). PCR reaction conditions were as follows: denaturation at 95 °C for 3 min, followed by 25 cycles denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, extension at 72 °C for 30 s, and final elongation at 72 °C for 5 min. The PCR amplified product was loaded on 1% agarose gel and purified using CleanPCR (CleanNA). The quality and product size were assessed on a Bioanalyzer 2100 (Agilent, Palo Alto, CA, USA) using a DNA 7500 chip. Mixed amplicons were pooled and the sequencing was carried out at Chunlab, Inc. (Seoul, Korea), with Illumina MiSeq Sequencing System (Illumina, CA, USA) according to the manufacturer’s instructions.
2.4. Cell Culture
The murine macrophage cell line RAW264.7 was purchased from the Korea Cell Line Bank (Seoul, Korea). The medium for cell culture contained fetal bovine serum (FBS, Merck, Darmstadt, Germany), penicillin, and streptomycin in Dulbecco’s Modified Eagle Medium (DMEM, Gibco). Cells were cultured every 2 days and incubated in a 37 °C, 5% CO2 incubator.
2.5. Measurement of Cell Viability
Cell viability was measured using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromi-de (MTT, Sigma). RAW264.7 cells were seeded in a 24-well plate to generate 1.5 × 105 cells/well and incubated for 24 h. The final concentration of the sample was treated to 31.25, 62.5, 125, 250, and 500 µg/mL, and then incubated for 24 h. The culture medium was removed and MTT (0.4 mg/mL) reagent was added and treated for 4 h. After dissolving the crystal in 800 µL DMSO in each well, the absorbance was measured at 570 nm. Cell viability was expressed as a percentage (%) compared to the control group not treated with lipopolysaccharides (LPS).
2.6. Measurement of Nitric Oxide Production
To produce NO, Griess reagent was used to measure the amount of nitrite, a metabolite of NO. After seeding and treatment in the same way as the MTT assay, 100 µL of each cell culture supernatant was taken and transferred to a 96-well plate, and 100 µL of Griess reagent was mixed to measure the absorbance at 540 nm. NO production was expressed as a percentage (%) compared to the control group treated with LPS alone.
2.7. Measurement of Cytokine Production
Mouse IL-6 ELISA kit (BD bioscience, San Diego, CA, USA), mouse TNF-α ELISA kit (BD bioscience, USA), mouse IL-1β ELISA kit (R & D Systems, Minneapolis, MN, USA), and mouse prostaglandin E2 (PGE2) ELISA kit (BD bioscience, USA) were used to measure cytokine expression in the culture medium. After seeding and treating in the same way as in the MTT assay, the supernatant was collected in each well. After centrifugation at 15,000 rpm for 3 min, the supernatant was used for the experiment. Experiments were conducted according to the protocol provided by each manufacturer, and the amount of cytokine produced was expressed as a percentage (%) compared to the control group treated with LPS alone.
2.8. Western Blot Assay
Western blotting was used to check the expression level of the inflammation-related protein to determine whether the sample was an anti-inflammatory substance. RAW264.7 cells were seeded in a 60-mm dish at 8 × 105 cells/dish and incubated for 24 h. After removing the medium, the medium was treated with LPS (Sigma, St. Louis, MO, USA) and the sample was processed at the same time and incubated according to conditions from a minimum of 15 min to 24 h. The medium was removed and washed with cold phosphate buffered saline (PBS; Biosesang, Korea). Each dish was incubated with 150 µL of RIPA lysis buffer (150 mM sodium chloride, 1% Triton X-100, 1% sodium deoxycholate, 0.1% SDS, 50 mM Tris-HCl, pH 7.5, and 2 mM EDTA, protease inhibitor cocktail) and scraped with a scraper. The supernatant obtained by centrifugation for 15 min at 4 °C and 15,000 rpm was quantified using the BCA Protein Assay Kit (Pierce, Madison, WI, USA). After electrophoresis of 30 µg protein using 10% SDS-PAGE, the separated protein was transblotted with a PVDF membrane (Bio-Rad, Hercules, CA, USA). The cells were blocked with 3% skimmed milk for 4 h, washed thrice at 10 min intervals with Tris-buffered saline Tween-20 (TBST), and the primary antibody was incubated overnight. After washing six times at 10 min intervals, the secondary antibody was incubated for 2 h. After washing again six times, the result was confirmed by X-ray film using enhanced chemiluminescence (ECL; Biosesang, Korea) method.
3. Results
3.1. Microbial Community Analysis
We first analyzed the microbial community in the purchased barley nuruk by 16S rRNA amplicon sequencing using the MiSeq system. The taxonomic results obtained using BaseSpace (Illumina) and MEGAN were then compared. The two analytic methods agreed that Bacillus spp. dominated the samples and the predominant species was B. amyloliquefaciens (75.4%), while Ralstonia pickettii (14.4%) and Paraburkholderia kururiensis (1.1%) were the next in abundance. At the phylum level, Firmicutes was the most dominant, followed by Proteobacteria (Supplementary Materials).
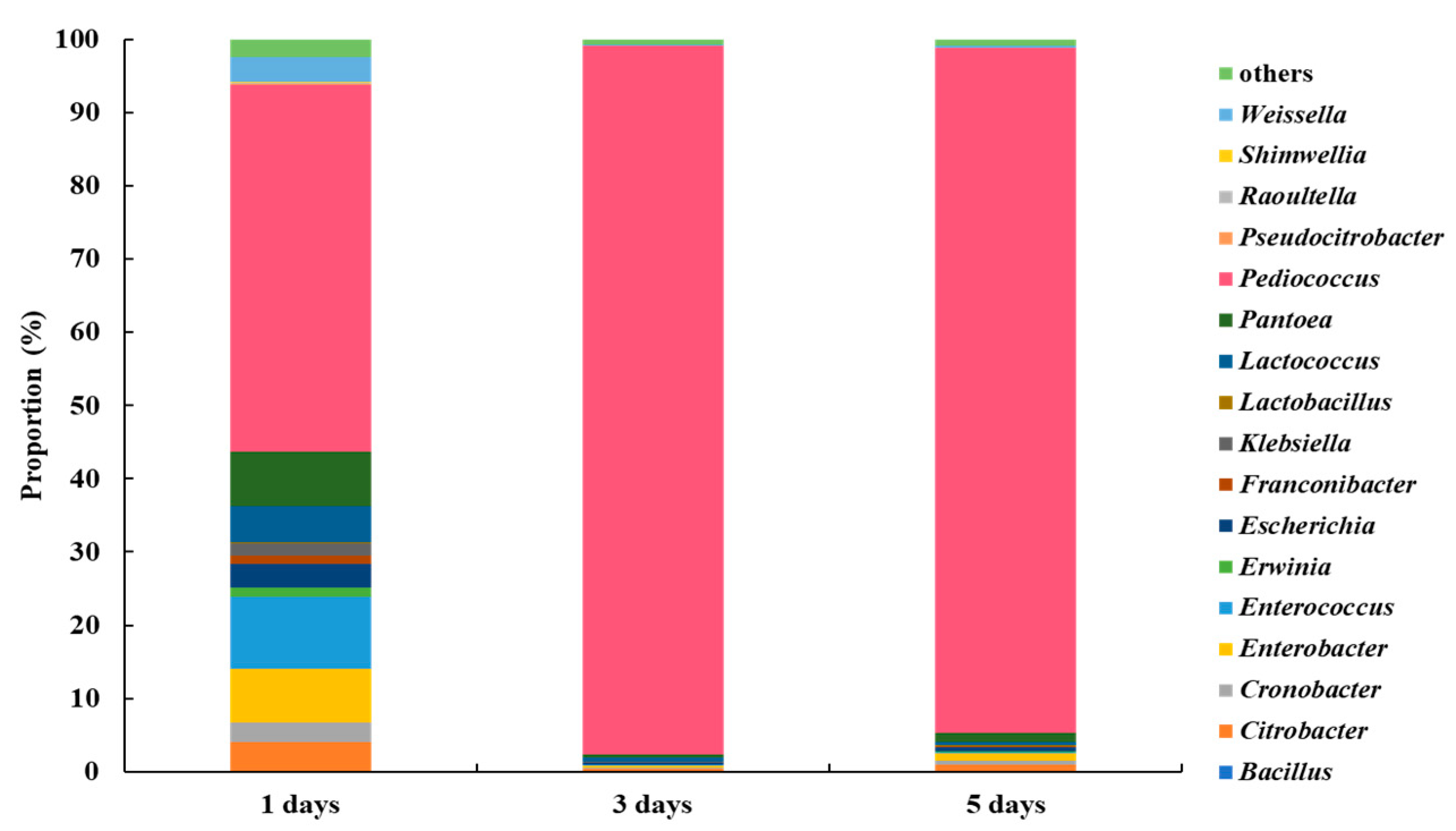
Figure 2 shows the results of the investigation at the genus level to investigate changes in the microbial community with fermentation time. Pediococcus predominated with 50.04% after 1 day; by day 3 and 5, this rose to 96.75% and 93.51%, respectively. Enterobacter are bacteria that cause pyogenic abscess in the urogenital system [8,9]. When shindari was fermented for day 1, 9.87% Enterobacter was present, but on day 3, 0.15%, and on day 5, 0.11% were present, which decreased as the fermentation time increased. The genus Erwinia is also a phytopathogenic bacterium [10]. When fermented for day 1, 1.21% was present, but for day 3 and 5, 0.06% and 0.14% were present, respectively. This was the lowest on day 3. In addition, the genus Klebsiella, known as the causative agent of community pneumonia [11], was 1.75% when fermented on day 1, but was 0.08% and 0.09% on day 3 and 5, respectively, and was the lowest on day 3. Therefore, on day 3 of fermentation, the most beneficial bacteria and the least harmful bacteria were present.
On the other hand, at the phylum level, Firmicutes was the most dominant in 3-day shindari compared to 1-day, followed by Proteobacteria. The Bacilli ratio at the Class level, Lactobacillales at the Order level, and Lactobacillaceae at the Family level dramatically increased with the shindari fermentation time (1-day to 3-day), forming the dominant group (Supplementary Materials).
3.2. Measurement of Cell Viability
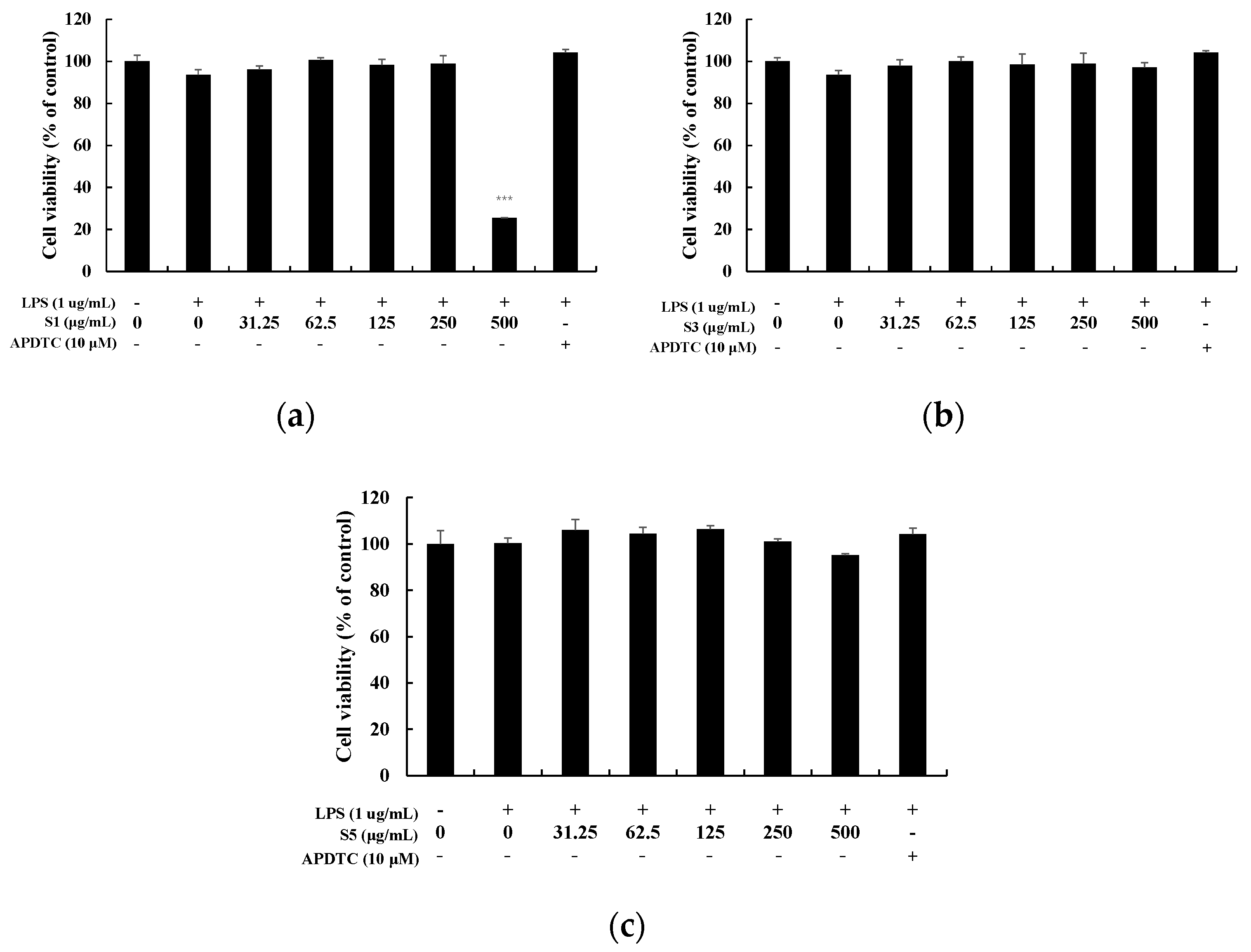
To investigate the cytotoxicity of EtOAc fr. of shindari fermentation, MTT assay was performed by treating extracts of various concentrations (31.25–500 µg/mL) to RAW264.7 cells (Figure 3). The cell viability of the extract was based on the cell viability (100%) of the untreated group. At S1, the cell viability was 25% at a concentration of 500 µg/mL. At 31.25–250 µg/mL, cell viability was over 90%. In the case of S3 or S5 days, cell viability was 90% or more at all concentrations, and did not show toxicity. Therefore, the NO production measurement experiment was conducted at 31.25–250 µg/mL for S1, and 31.25–500 µg/mL for S3 and S5.
3.3. Measurement of Nitric Oxide Production
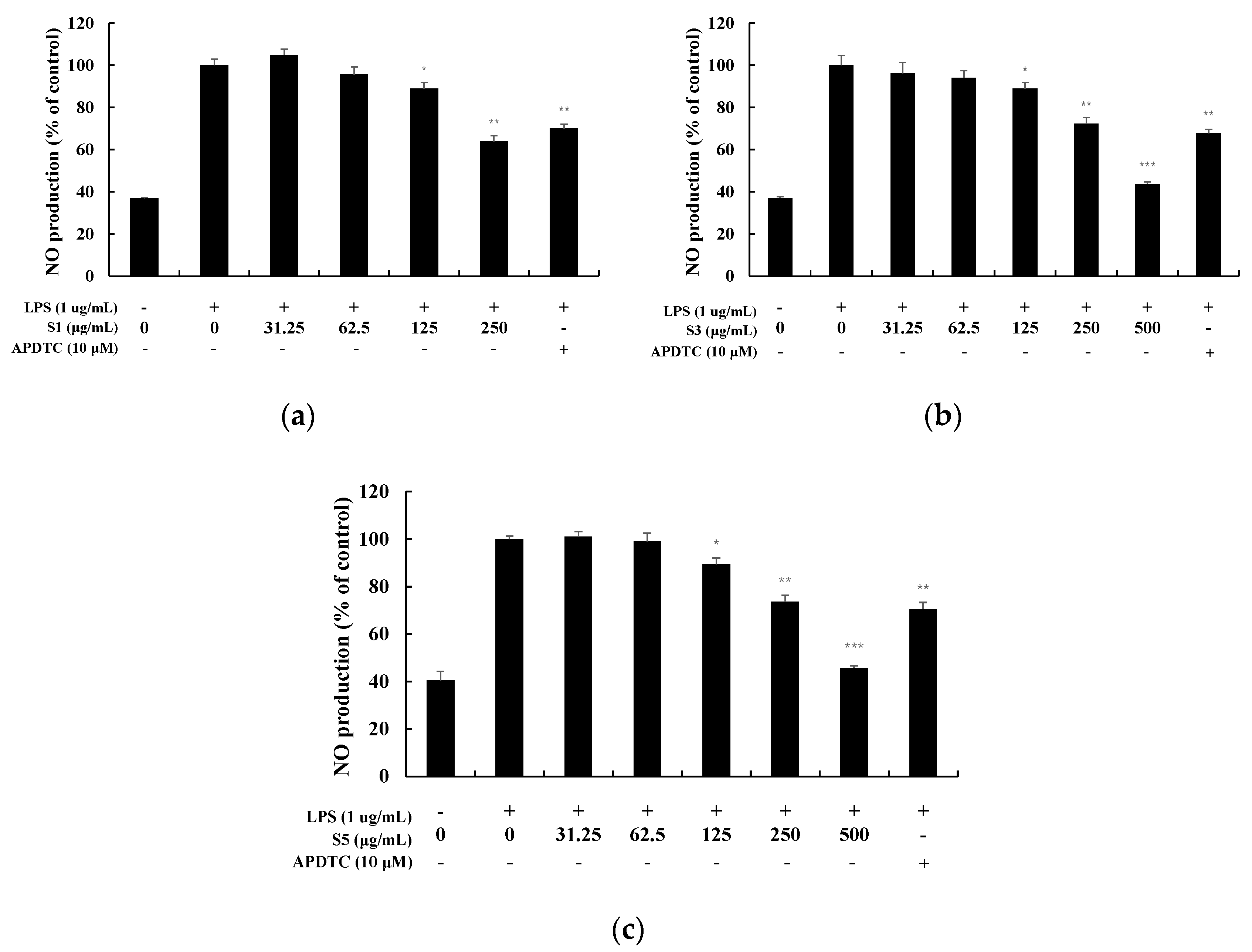
NO, an inflammatory mediator produced by activating RAW264.7 cells with LPS, was measured using Griess reagent in the form of nitrite ions (NO2−) in the culture medium (Figure 4) [12]. At 1 day, NO production was suppressed in a concentration-dependent manner, and at a high concentration of 250 µg/mL, 36% was suppressed. The S3 and S5 also decreased in a concentration-dependent manner, and were suppressed by 56% and 54%, respectively, at a high concentration of 500 µg/mL. Therefore, additional experiments were conducted for S3 and S5 when they exhibited excellent effects in the range where no toxicity appeared.
3.4. Measurement of Cytokine Production
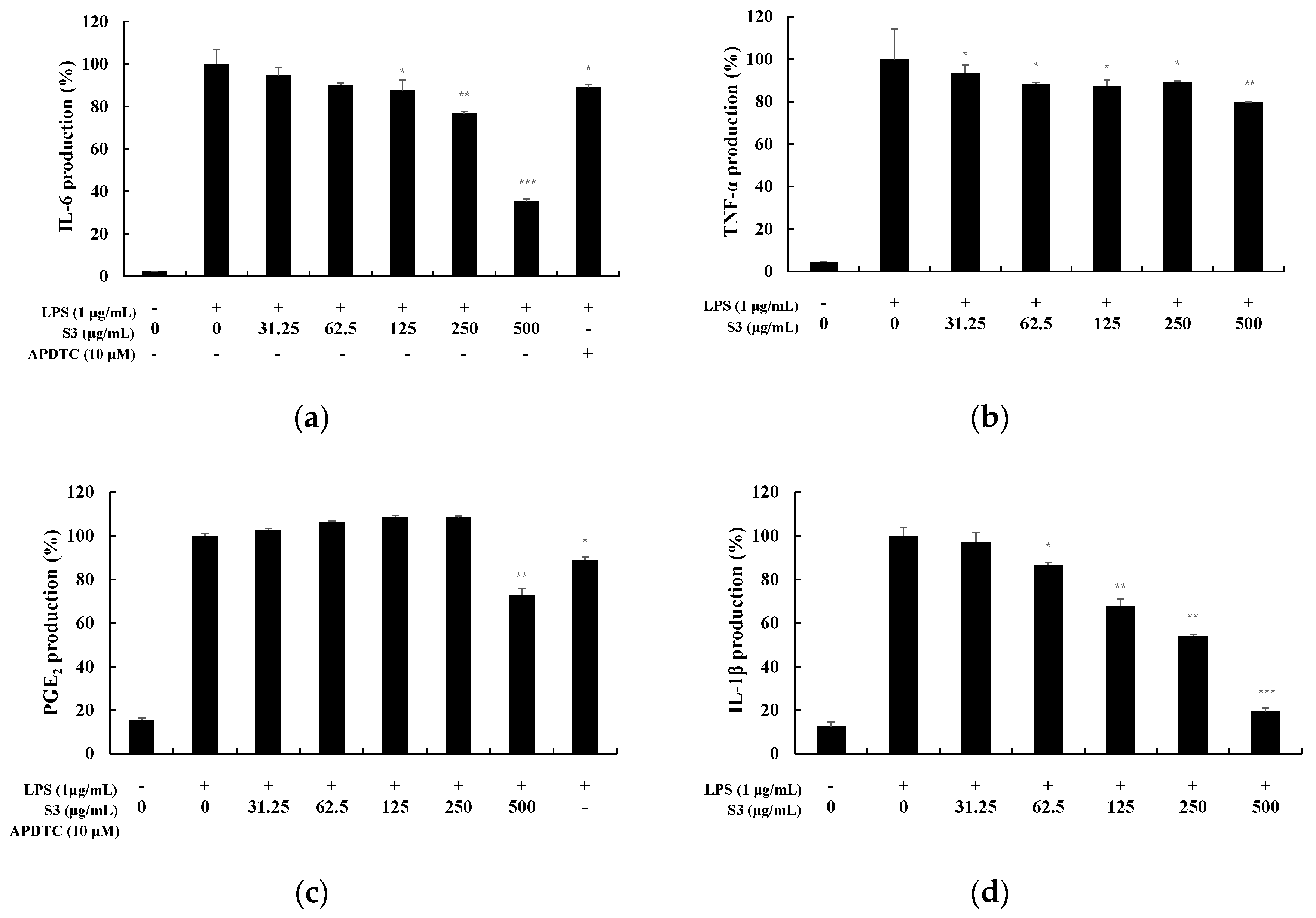
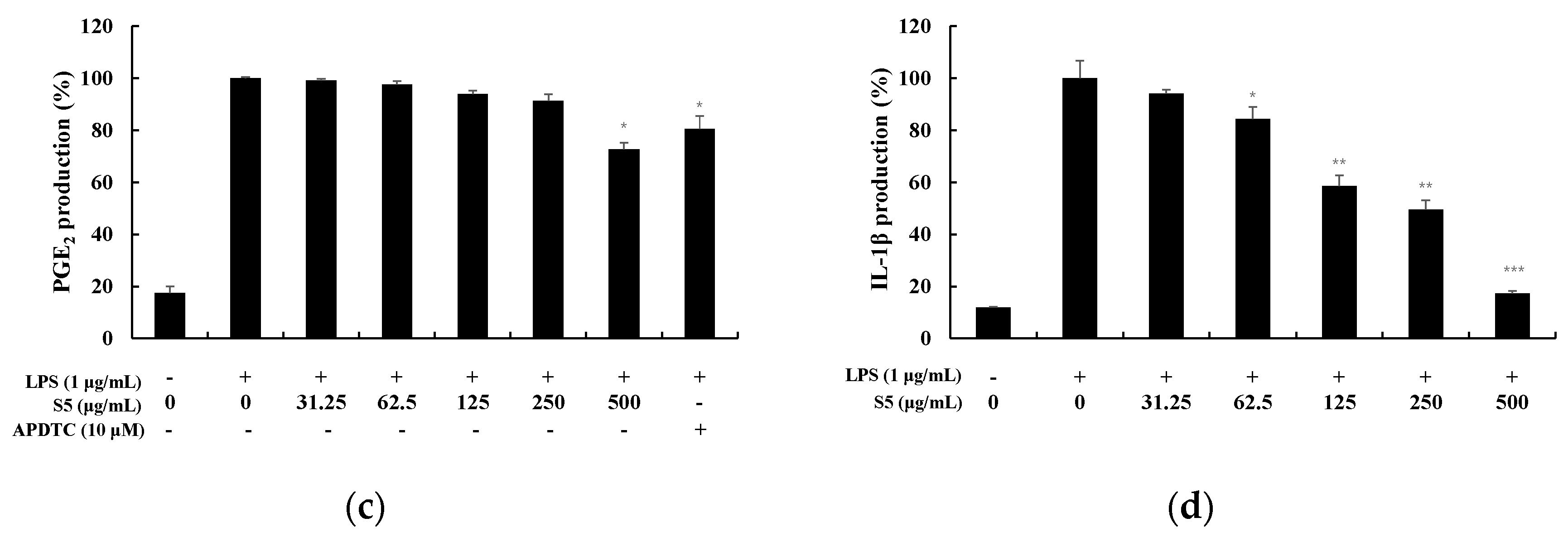
TNF-α is a cytokine that mediates acute inflammatory responses and is known as an early cytokine secreted by macrophages stimulated by LPS. In addition, it stimulates the secretion of IL-6, IL-1β, etc., thereby causing an inflammatory response [13]. COX-2 produced by these pro-inflammatory stimuli causes the secretion of PGE2, an inflammation-causing substance [14]. PGE2 causes inflammatory responses and vasodilation, and may also produce tumors when overexpressed [15]. Therefore, the expression level of cytokines was measured to examine the anti-inflammatory effect of S3 and S5 (Figure 5 and Figure 6). At S3, IL-6 was inhibited by 65%, TNF-α by 20%, PGE2 by 34%, and IL-1β by 81% at 500 µg/mL. In particular, it was observed that the production of IL-6 and IL-1β was significantly inhibited.
3.5. iNOS, COX-2 Expression in LPS-Induced RAW264.7
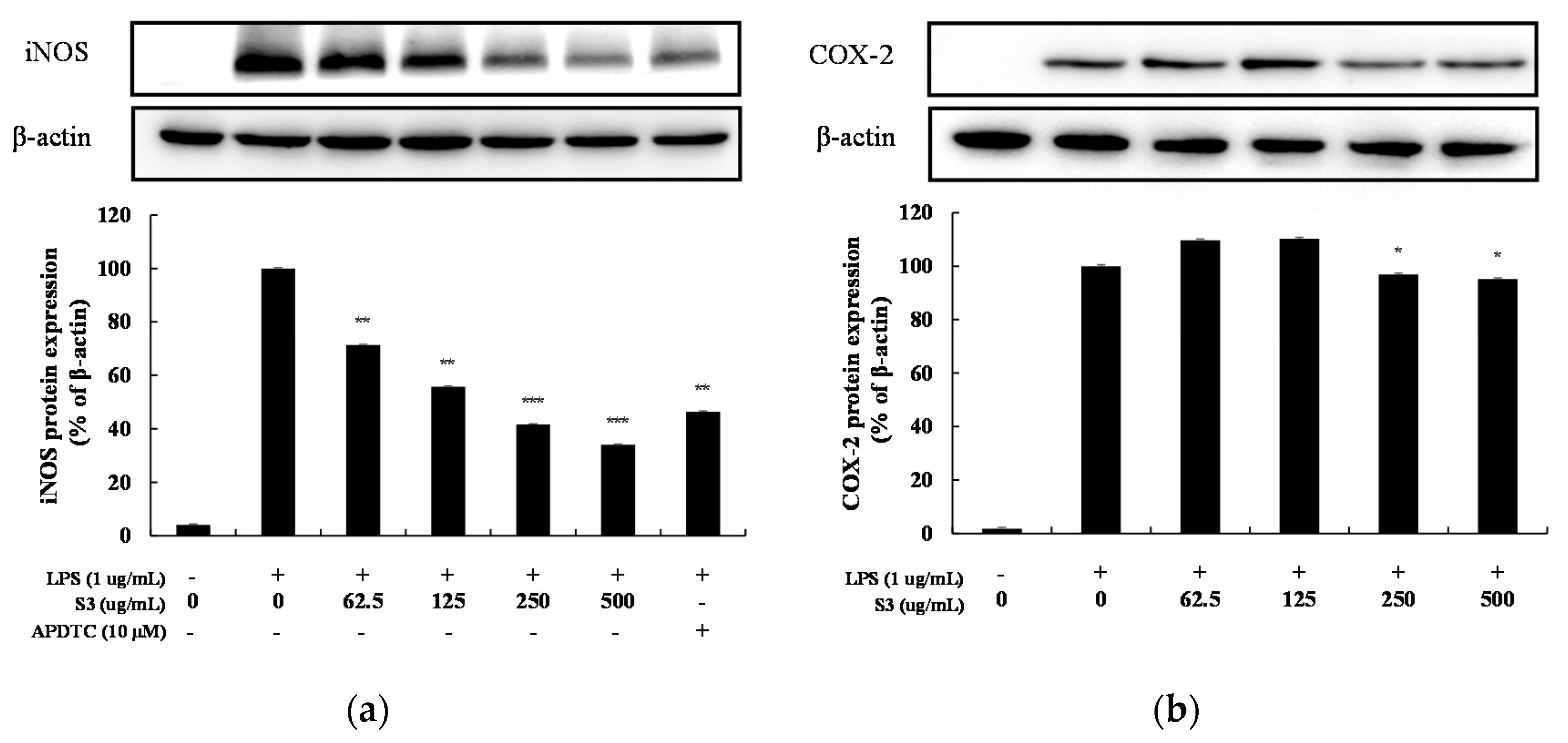
Western blotting was performed to measure the inhibitory effect of inflammatory inducers iNOS and COX-2 protein expression. At this time, the housekeeping gene β-actin, which has little difference in expression level even under various conditions of cells, was used as a positive control [15]. When treated with 62.5–500 µg/mL of S3 in RAW264.7 cells increased by LPS, COX-2 tended to increase and then decrease slightly, but the expression level of iNOS decreased in a concentration-dependent manner (Figure 7). At a high concentration of 500 µg/mL, iNOS expression was inhibited by 66%.
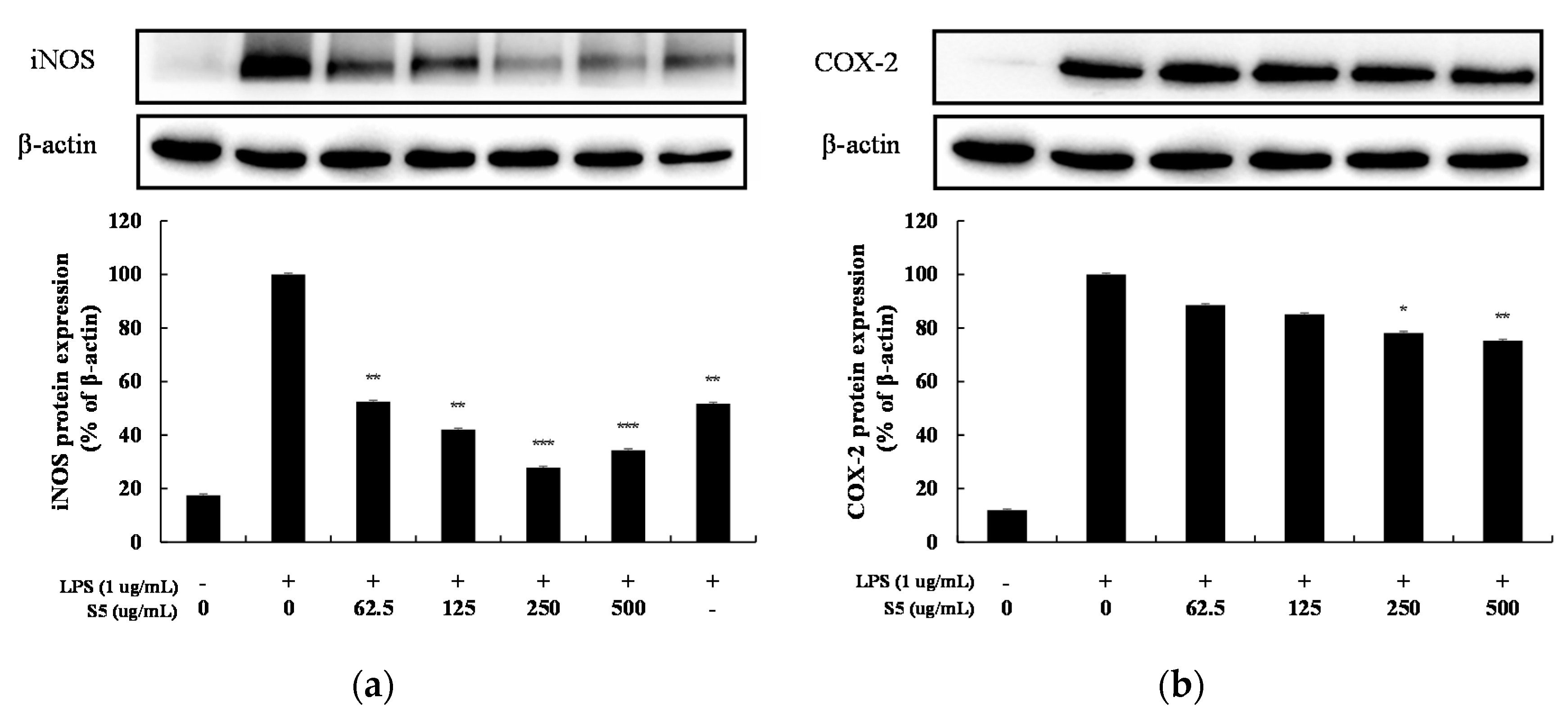
As a result of treatment with 62.5–500 µg/mL of S5 on RAW264.7 cells increased by LPS, the expression levels of iNOS and COX-2 all decreased in a concentration-dependent manner (Figure 8). At a high concentration of 500 µg/mL, iNOS expression was suppressed by 66% and COX-2 expression was suppressed by 25%.
3.6. MAPK and NF-κB Signaling Activation in LPS-Induced RAW264.7
MAPKs are representative intracellular inflammatory signaling factors expressed by inflammatory stimuli, such as LPS. There are three types of inflammation: JNK, ERK, and p38, and phosphorylation increases when stimulated, causing inflammation. NF-κB is a transcription factor that regulates the synthesis of various inflammation-related factors by external stimulation. NF-κB binds to IκB-α in the cytoplasm and remains inactive, but when stimulation is transmitted by an inflammatory substance, IκB-α is phosphorylated and separated to change into an activated form [16].
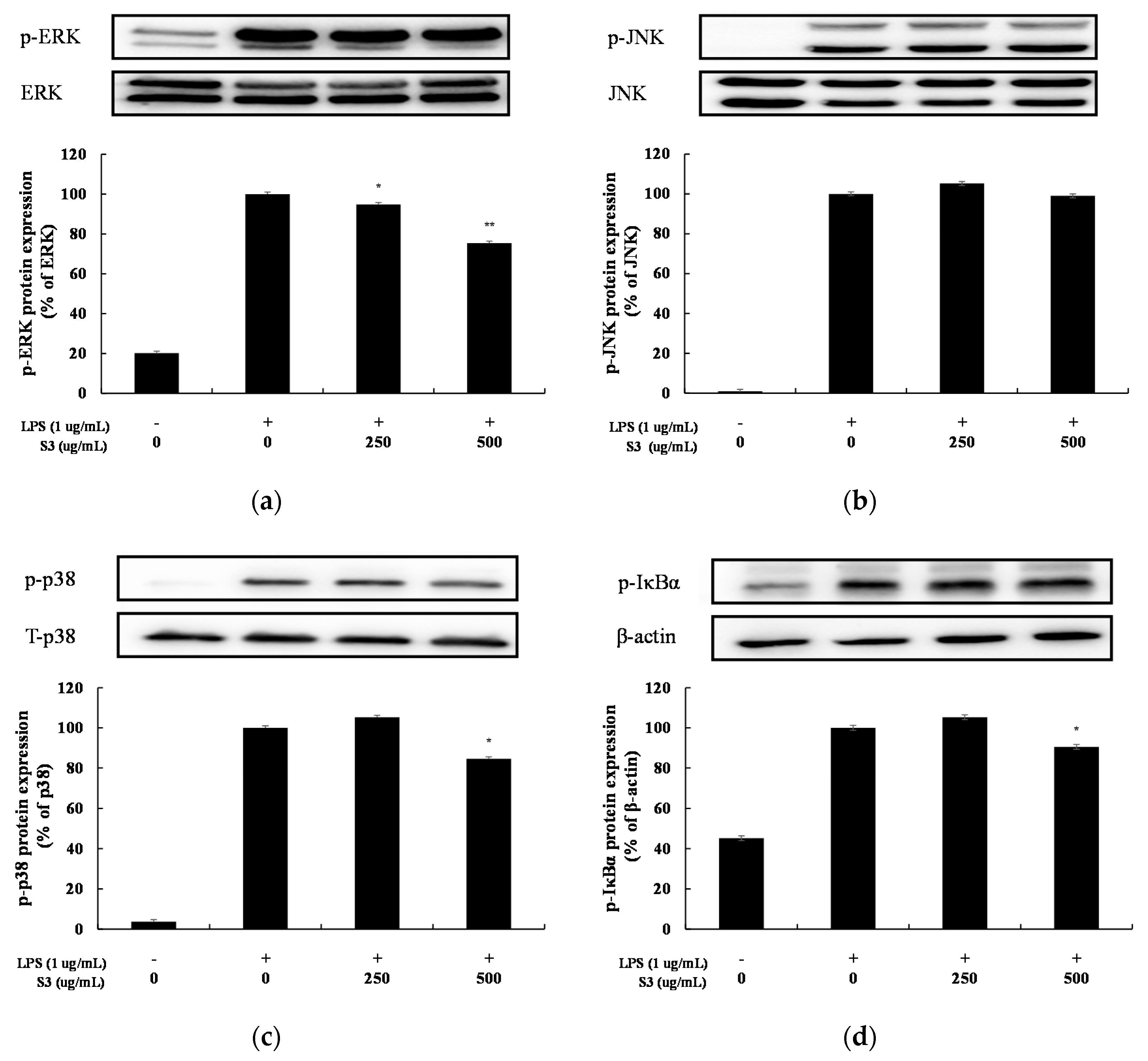
In this study, we investigated the anti-inflammatory effect of shindari fermentation EtOAc fr., the phosphorylation inhibitory effects of ERK, JNK, and IκB-α were examined. Phosphorylation of ERK was inhibited by 25% at 500 µg/mL and phosphorylation of p38 was inhibited by 15% at 500 µg/mL. Phosphorylation of JNK was not observed. Phosphorylation of IκBα also decreased in a concentration-dependent manner and was inhibited by 10% at 500 µg/mL (Figure 9). Therefore, the anti-inflammatory mechanism according to the treatment of S3 has been shown to suppress the MAPKs and NF-κB signaling system inside macrophages.
4. Discussion
Throughout the world, there are traditional fermented foods that represent the country and region that have long been handed down with the wisdom of human ancestors. These traditional fermented foods include Chinese condiments [17], “douch”, an Indonesian témpé made by fermenting soybeans [18], Sweden’s surströmming, famous for the world’s most odorless food [19], and German sauerkraut made by fermenting vegetables [20] like Korean kimchi. In addition, narezushi, a Japanese fermented natural mixture of salted fish meat mixed with rice, is a fermented food that is the beginning of today’s sushi, and in Thailand, there is a traditional fermented food called nan pla fermented with salt in fish [21]. Of course, Jeju Island, the southernmost island on the Korean Peninsula, has a fermented drink called shindari that has been handed down for hundreds of years. Jeju Island, which is composed of barren basalt soil, is a geographical environment where rice farming is not possible, and traditional fermented foods are made using barley. Shindari is also made by adding spoiled rice as a sugar source using barley nuruk and fermenting it for three to four days.
In the real environment, microorganisms adapt to the surrounding environment and form a community through interactions with other microorganisms, and the entire microbial community has various expression characteristics through interaction between individual microorganisms. Therefore, it is very important to understand the microbial community rather than the individual organisms. As a method for investigating the structure of the microbial community, molecular genetic techniques through Illumina MiSeq sequencing have been widely used in recent years. In this study, we attempted to primarily observe changes in the microbial community at each stage of the fermentation process, which is traditionally manufactured through fermentation for 3 to 4 days. As a result of analyzing the microbial community for each sample after fermentation in a microbial incubator for 1, 3, and 5 days using barley nuruk and rice, we found a very interesting result that different fermentation time had different influences on the community structure and microbial diversity in shindari manufacturing process. Ultimately, we found that samples with a fermentation period of more than 3 days contained over 97% lactic acid bacteria, such as Pediococcus, an alcohol-tolerant lactic acid bacteria. This is the fact that Saccharomyces cerevisiae is involved in the Shindari fermentation process within 24 h of fermentation, using the sugars in the rice to make ethanol, carbon dioxide, and various volatile compounds, and then following Saccharomyces cerevisiae in the 48 to 72 h of fermentation, the Pediococcus sp. is rapidly dominant. The Pediococcus genus is one of the representative lactic acid bacteria and has a growth inhibitory effect on several spoilage and pathogenic microorganisms. It has been shown to enhance immunity and reduces blood cholesterol [8].
Foods fermented by lactic acid bacteria have been reported to have various health-promoting effects, such as anti-microbial, immune-promoting, and anti-tumor effects [22]. These beneficial lactic acid bacteria were most present when fermented on day 3. Ancestors consumed shindari after 3–4 days of fermentation; therefore compared with the analysis results, the wisdom of the ancestors was evident.
In this study, we further identified microbial community and its correlation of anti-inflammatory effect in the shindari fermentation process in mouse macrophage RAW264.7 cells. As a result, S3 and S5 extracts had the best inhibitory effect on NO production. This is in line with the tendency of various studies that the anti-inflammatory activity of the extract was increased by the fermentation of lactic acid bacteria [23,24,25]. Therefore, subsequent experiments were conducted only on S3 and S5 and the expression levels of cytokines (IL-6, IL-1β, TNF-α, and PGE2) were measured. On S3 and S5 extracts, cytokines were reduced in a concentration-dependent manner, particularly IL-6 and IL-1β.
In addition, S3 and S5 extracts decreased the expression of iNOS which were related to NO production in a dose-concentration pattern. In the case of S5 extract, the expression levels of the COX-2 which were related to PGE2 production gradually decreased. Furthermore, the present study investigated whether MAPK and/or NF-kB signaling pathways were associated with the macrophage inhibition by S3 treatment. The protein expression levels of ERK, JNK. p38 and IκB-α, and their phosphorylated counterparts, were determined in the S3-treated cells via Western blotting. Compared with the untreated control group, the phosphorylated levels of ERK, p38 and IκB-α, in the S3-treated cells were significantly inhibit. Therefore, the anti-inflammatory mechanism according to the treatment of S3 extract is thought to suppress the expression of NO, cytokines, iNOS, and COX-2 by inhibiting the MAPKs and NF-κB signaling pathways in mouse macrophage RAW264.7 cells. In summary, we confirmed in this paper that shindari, a fermented food that Jeju natives have been drinking for hundreds of years, is a treasure trove of useful microorganisms, while also having anti-inflammatory effects. Through this, we are confident that we have opened a new horizon in the study of fermented foods handed down by our ancestor.
5. Conclusions
In this study, we investigated the microbial community generated during the fermentation process in the manufacture of shindari using Illumina MiSeq sequencing. Ultimately, we confirmed that shindari is a treasure trove of useful microorganisms. In addition, we tried to evaluate whether this microbial community has anti-inflammatory efficacy. We identified that anti-inflammatory effect of Shindari was mediated through the suppression of NO, PGE2, IL-6, IL-1β, and TNF-α production via the NF-κB and MAPK signaling pathways.
In conclusion, this study is, to the best of our knowledge, the first to elucidate the microbial communities and anti-inflammatory activity of shindari, a traditional fermented food that Jeju natives have been drinking for hundreds of years. Through this, we are confident that we have opened a new pathway in the study of the health benefits of traditional fermented foods.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2311-5637/6/3/87/s1.
Author Contributions
Conceptualization, C.-G.H.; software, S.B.H.; validation, S.B.H.; formal analysis, S.B.H.; investigation, C.-G.H.; writing—original draft preparation, S.B.H.; writing—review and editing, C.-G.H.; project administration, C.-G.H.; funding acquisition, C.-G.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Korea Institute for Advancement of Technology (KIAT) grant funded by the Korea Government (MOTIE) (N0002327, The Establishment Project of Industry-University Fusion District).
Acknowledgments
The authors thank all the students in our research group for their helpful cooperation and discussions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lee, J. Research of the Food Culture Comparison between the Tamra & Mongolia. Trans Humanit. 2012, 5, 211–243. [Google Scholar]
- Ahn, S.L.; Yoon, J.H. Development of Jeju Local Food Tour Package and Economic Value Estimation based on Contingent Valuation Method. J. East Asian Soc. Diet. Life 2016, 26, 346–358. [Google Scholar] [CrossRef]
- Kim, S.Y.; Park, E.J. Fermentation Characteristics of Shindari Added with Carrot. Korean J. Food Cook Sci. 2015, 3, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Fanning, S.; Proos, S.; Jordan, K.; Srikumar, S. A Review on the Applications of Next Generation Sequencing Technologies as Applied to Food-Related Microbiome Studies. Front. Microbiol. 2017, 8, 1829. [Google Scholar] [CrossRef]
- Sarkar, D.; Fisher, P.B. Molecular Mechanisms of Aging-associated Inflammation. Cancer Lett. 2006, 236, 13–23. [Google Scholar] [CrossRef]
- Jeong, D.H.; Kang, B.K.; Kim, K.B.W.R.; Kim, M.J.; Ahn, D.H. Anti-inflammatory Activity of Sargassum micracanthum Water Extract. J. Appl. Biol. Chem. 2014, 57, 227–234. [Google Scholar] [CrossRef]
- Choi, S.B.; Bae, G.S.; Jo, I.J.; Park, K.C.; Seo, S.H.; Kim, D.G.; Shin, J.Y.; Gwak, T.S.; Lee, J.H.; Lee, G.S.; et al. The Anti-inflammatory Effect of Lithospermum erythrorhizon on Lipopolysaccharide-induced Inflammatory Response in RAW 264.7 Cells. Korea J. Herbol. 2013, 28, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Ryu, B.H.; Sim, G.S.; Choi, H.Y.; Ha, W.K. A Study on the Natural Preservative(Lactobacillus-fermented antimicrobial solution), Fermented with Plant Originated Lactic Acid Bacteria. Food Sci. Ind. 2011, 44, 45–51. [Google Scholar]
- Moon, Y.L.; You, J.W.; Shim, H.J. Arthroscopic Management of The Septic Gonitis. Knee Surg. Relat. Res. 2001, 13, 114–116. [Google Scholar]
- Seo, S.T.; Lee, J.S.; Park, J.H.; Han, K.S.; Jang, H.I. Investigation of Antibiotic Susceptibility of Some Plant Pathogenic Bacteria. Korean J. Hort. Sci. Technol. 2005, 23, 495–498. [Google Scholar]
- Jeong, I.; Heo, E.Y.; Lee, J.S.; Kim, H.R.; Lee, E.Y.; Kim, Y.W.; Yang, S.C. Streptococcus salivarius Pneumonia with Pulmonary Nocardiosis in a Rheumatoid Arthritis Patient Treated with Immunosuppressants. Korean J. Med. 2009, 77, 128–132. [Google Scholar]
- Lee, J.Y.; Yoo, D.H.; Joo, D.H.; Kim, S.R.; Jo, H.S.; Joo, S.H.; Chae, J.W. Anti-inflammatory Effects of Amelanchier asiatica Fruits Ethanol Extract. J. Soc. Cosmet. Sci. Korea 2017, 43, 19–26. [Google Scholar]
- Blaser, H.; Dostert, C.; Mak, T.W.; Brenner, D. TNF and ROS Crosstalk in Inflammation. Trends Cell Biol. 2016, 26, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Bolli, R.; Shinmura, K.; Tang, X.L.; Kodani, E.; Xuan, Y.T.; Guo, Y.; Dawn, B. Discovery of a New Function of Cyclooxygenase (COX)-2: COX-2 is a Cardioprotective Protein that Alleviates Ischemia/Reperfusion Injury and Mediates the Late Phase of Preconditioning. Cardiovasc. Res. 2002, 55, 506–519. [Google Scholar] [CrossRef]
- Patrono, C. Cardiovascular Effects of Cyclooxygenase-2 Inhibitors: A Mechanistic and Clinical Perspective. Br. J. Clin. Pharmacol. 2016, 82, 957–964. [Google Scholar] [CrossRef]
- Jang, H.H. A Study on the Popularization Process of Chinese Tea. J. Chin. Hum. 2018, 8, 509–535. [Google Scholar]
- Kim, H.S.; Ham, J.S. Antioxidative Ability of Lactic Acid Bacteria. Korean Food Sci. Anim. Resour. 2003, 23, 186–192. [Google Scholar]
- Kobayashi, T.; Kimura, B.; Fujii, T. Strictly Anaerobic Halophiles Isolated from Canned Swedish Fermented Herrings (Surströmming). Int. J. Food Microbiol. 2000, 54, 81–89. [Google Scholar] [CrossRef]
- Barrangou, R.; Yoon, S.S.; Breidt, F., Jr.; Fleming, H.P.; Klaenhammer, T.R. Characterization of Six Leuconostoc fallax Bacteriophages Isolated from an Industrial Sauerkraut Fermentation. Appl. Environ. Microbiol. 2002, 68, 5452–5458. [Google Scholar] [CrossRef] [Green Version]
- Shurtleff, W.; Aoyagi, A. History of Tempeh; Soyinfo Center: Lafayette, CA, USA, 2007; p. 85. [Google Scholar]
- Feng, C.H. The Tale of Sushi: History and Regulations. Compr. Rev. Food Sci. Food Saf. 2012, 11, 205–220. [Google Scholar] [CrossRef]
- Choi, W.S.; Kwon, H.S.; No, R.H.; Choi, G.P.; Lee, H.Y. Enhancement of Anti-inflammatory Activities of Fermented Scutellaria baicalensis Extracts using Lactobacillus rhamnosus. J. Soc. Cosmet. Sci. Korea 2013, 39, 303–311. [Google Scholar]
- Song, H.S.; Eom, S.H.; Kang, Y.M.; Choi, J.D.; Kim, Y.M. Enhancement of the Antioxidant and Anti-inflammatory Activity of Hizikia fusiforme Water Extract by Lactic Acid Bacteria Fermentation. Korean J. Fish Aquat. Sci. 2011, 44, 111–117. [Google Scholar]
- Lim, H.W.; Lee, Y.J.; Huang, Y.H.; Yoon, J.Y.; Lee, S.H.; Kim, K.H.; Lim, C.J. Enhancement of Skin Antioxidant and Anti-Inflammatory Potentials of Agastache rugosa Leaf Extract by Probiotic Bacterial Fermentation in Human Epidermal Keratinocytes. Microbiol. Biotechnol. Lett. 2017, 45, 35–42. [Google Scholar] [CrossRef]
- Kim, S.Y.; Park, Y.K. Ethanol Extract of Forsythiae fructus Inhibits the Production of Inflammatory Mediators in LPS-stimulated BV-2 Microglial Cells. Korea J. Herbol. 2008, 23, 93–102. [Google Scholar]
Figure 1.
Barley nuruk manufacturing process.

Figure 2.
Composition of microbial communities in shindari fermentation.

Figure 3.
Cell viability in LPS-induced RAW264.7 macrophage cells. Cells were each treated with (a) EtOAc fr. of 1 day of shindari fermentation (S1; 31.25, 62.5, 125, 250, and 500 µg/mL), (b) EtOAc fr. of 3 days of shindari fermentation (S3; 31.25, 62.5, 125, 250, and 500 µg/mL), and (c) EtOAc fr. of 5 days of shindari fermentation (S5; 31.25, 62.5, 125, 250, and 500 µg/mL) for 24 h. Cell viability was determined by MTT assay. Data are presented as the mean ± standard deviation (SD) of at least three independent experiments (n = 3). *** p < 0.001.
Figure 3.
Cell viability in LPS-induced RAW264.7 macrophage cells. Cells were each treated with (a) EtOAc fr. of 1 day of shindari fermentation (S1; 31.25, 62.5, 125, 250, and 500 µg/mL), (b) EtOAc fr. of 3 days of shindari fermentation (S3; 31.25, 62.5, 125, 250, and 500 µg/mL), and (c) EtOAc fr. of 5 days of shindari fermentation (S5; 31.25, 62.5, 125, 250, and 500 µg/mL) for 24 h. Cell viability was determined by MTT assay. Data are presented as the mean ± standard deviation (SD) of at least three independent experiments (n = 3). *** p < 0.001.

Figure 4.
Nitric oxide production in LPS-induced RAW 264.7 macrophage cells. macrophages Cells were each treated with (a) EtOAc fr. of 1-day shindari fermentation (S1; 31.25, 62.5, 125, and 250 µg/mL), (b) EtOAc fr. of 3-day shindari fermentation (S3; 31.25, 62.5, 125, 250, and 500 µg/mL) and (c) EtOAc fr. of 5-day shindari fermentation (S5; 31.25, 62.5, 125, 250, and 500 µg/mL) for 24 h. Data are presented as mean ± standard deviation (SD) of at least four independent experiments (n = 3). * p < 0.05, ** p < 0.01, and *** p < 0.001 versus treated control.
Figure 4.
Nitric oxide production in LPS-induced RAW 264.7 macrophage cells. macrophages Cells were each treated with (a) EtOAc fr. of 1-day shindari fermentation (S1; 31.25, 62.5, 125, and 250 µg/mL), (b) EtOAc fr. of 3-day shindari fermentation (S3; 31.25, 62.5, 125, 250, and 500 µg/mL) and (c) EtOAc fr. of 5-day shindari fermentation (S5; 31.25, 62.5, 125, 250, and 500 µg/mL) for 24 h. Data are presented as mean ± standard deviation (SD) of at least four independent experiments (n = 3). * p < 0.05, ** p < 0.01, and *** p < 0.001 versus treated control.

Figure 5.
Effect of EtOAc fr. of 3 day shindari fermentation (S3) on LPS-induced (a) interleukin-6 (IL-6), (b) tumor necrosis factor-α (TNF-α), (c) PGE2 and (d) IL-1β production in macrophages. RAW264.7 cells incubated with LPS (1 μg/mL) for 24 h in the presence or absence of EtOAc fraction extraction. of 3 day shindari fermentation (3 days; 31.25, 62.5, 125, 250, and 500 µg/mL). Culture supernatants were collected and analyzed for the presence of induced IL-6, TNF-α, IL-1β, and PGE2 using enzyme-linked immunosorbent assays. Data are presented as the mean ± standard deviation (SD) of at least three independent experiments (n = 3). * p < 0.05, ** p < 0.01, and *** p < 0.001 versus treated control.
Figure 5.
Effect of EtOAc fr. of 3 day shindari fermentation (S3) on LPS-induced (a) interleukin-6 (IL-6), (b) tumor necrosis factor-α (TNF-α), (c) PGE2 and (d) IL-1β production in macrophages. RAW264.7 cells incubated with LPS (1 μg/mL) for 24 h in the presence or absence of EtOAc fraction extraction. of 3 day shindari fermentation (3 days; 31.25, 62.5, 125, 250, and 500 µg/mL). Culture supernatants were collected and analyzed for the presence of induced IL-6, TNF-α, IL-1β, and PGE2 using enzyme-linked immunosorbent assays. Data are presented as the mean ± standard deviation (SD) of at least three independent experiments (n = 3). * p < 0.05, ** p < 0.01, and *** p < 0.001 versus treated control.

Figure 6.
Effect of EtOAc fr. of 5 day shindari fermentation (S5) on LPS-induced (a) IL-6, (b) TNF-α, (c) PGE2 and (d) IL-1β production in macrophages. RAW264.7 cells incubated with LPS (1 μg/mL) for 24 h in the presence or absence of EtOAc fr. Extraction of 5 day shindari fermentation (S5; 31.25, 62.5, 125, 250, and 500 µg/mL). Culture supernatants were collected and analyzed for the presence of induced IL-6, TNF-α, IL-1β, and PGE2 using enzyme-linked immunosorbent assays. Data are presented as the mean ± standard deviation (SD) of at least three independent experiments (n = 3). * p < 0.05, ** p < 0.01, and *** p < 0.001 versus treated control.
Figure 6.
Effect of EtOAc fr. of 5 day shindari fermentation (S5) on LPS-induced (a) IL-6, (b) TNF-α, (c) PGE2 and (d) IL-1β production in macrophages. RAW264.7 cells incubated with LPS (1 μg/mL) for 24 h in the presence or absence of EtOAc fr. Extraction of 5 day shindari fermentation (S5; 31.25, 62.5, 125, 250, and 500 µg/mL). Culture supernatants were collected and analyzed for the presence of induced IL-6, TNF-α, IL-1β, and PGE2 using enzyme-linked immunosorbent assays. Data are presented as the mean ± standard deviation (SD) of at least three independent experiments (n = 3). * p < 0.05, ** p < 0.01, and *** p < 0.001 versus treated control.


Figure 7.
Effect of EtOAc fr. of 3 days of shindari fermentation on the level of inducible nitric oxide synthase (iNOS) and COX-2 protein in LPS-induced RAW264.7 macrophage cells. RAW264.7 cells incubated with LPS (1 μg/mL) for 24 h in the presence or absence of EtOAc fraction extraction of 3 days of shindari fermentation (S3; 62.5, 125, 250, and 500 µg/mL). (a) iNOS protein level and (b) COX-2 protein level were validated by Image J. β-actin was used as an internal control. Data are presented as the mean ± standard deviation (SD) of at least three independent experiments (n = 3). * p < 0.05, ** p < 0.01, and *** p < 0.001 versus treated control.
Figure 7.
Effect of EtOAc fr. of 3 days of shindari fermentation on the level of inducible nitric oxide synthase (iNOS) and COX-2 protein in LPS-induced RAW264.7 macrophage cells. RAW264.7 cells incubated with LPS (1 μg/mL) for 24 h in the presence or absence of EtOAc fraction extraction of 3 days of shindari fermentation (S3; 62.5, 125, 250, and 500 µg/mL). (a) iNOS protein level and (b) COX-2 protein level were validated by Image J. β-actin was used as an internal control. Data are presented as the mean ± standard deviation (SD) of at least three independent experiments (n = 3). * p < 0.05, ** p < 0.01, and *** p < 0.001 versus treated control.

Figure 8.
Effect of extraction of EtOAc fr. of 5 days of shindari fermentation on the level of iNOS and COX-2 protein in LPS-induced RAW264.7 macrophage cells. RAW264.7 cells incubated with LPS (1 μg/mL) for 24 h in the presence or absence of EtOAc fr. extraction of 5 day shindari fermentation (S5; 62.5, 125, 250, and 500 µg/mL). (a) iNOS protein level and (b) COX-2 protein level were validated by Image J. β-actin was used as an internal control. Data are presented as the mean ± standard deviation (SD) of at least three independent experiments (n = 3). * p < 0.05, ** p < 0.01, and *** p < 0.001 versus treated control.
Figure 8.
Effect of extraction of EtOAc fr. of 5 days of shindari fermentation on the level of iNOS and COX-2 protein in LPS-induced RAW264.7 macrophage cells. RAW264.7 cells incubated with LPS (1 μg/mL) for 24 h in the presence or absence of EtOAc fr. extraction of 5 day shindari fermentation (S5; 62.5, 125, 250, and 500 µg/mL). (a) iNOS protein level and (b) COX-2 protein level were validated by Image J. β-actin was used as an internal control. Data are presented as the mean ± standard deviation (SD) of at least three independent experiments (n = 3). * p < 0.05, ** p < 0.01, and *** p < 0.001 versus treated control.

Figure 9.
Effect of EtOAc fr. of 3 days of Shindari fermentation on the level of phosphorylation of IκBα, and activation of extracellular signal regulatory kinase 1/2 (ERK ½), c-Jun amino-terminal kinase (JNK) 1/2 protein in LPS-induced RAW264.7 macrophage cells. RAW264.7 cells incubated with LPS (1 μg/mL) for 24 h in the presence or absence of EtOAc fr. of the shindari fermentation. (a) Protein levels of p-ERK, (b) p-JNK, (c) p-p38 and (d) p-IκBα were validated by Image J. β-actin was used as an internal control. Data are presented as the mean ± standard deviation (SD) of at least three independent experiments (n = 3). * p < 0.05, ** p < 0.01 versus treated control.
Figure 9.
Effect of EtOAc fr. of 3 days of Shindari fermentation on the level of phosphorylation of IκBα, and activation of extracellular signal regulatory kinase 1/2 (ERK ½), c-Jun amino-terminal kinase (JNK) 1/2 protein in LPS-induced RAW264.7 macrophage cells. RAW264.7 cells incubated with LPS (1 μg/mL) for 24 h in the presence or absence of EtOAc fr. of the shindari fermentation. (a) Protein levels of p-ERK, (b) p-JNK, (c) p-p38 and (d) p-IκBα were validated by Image J. β-actin was used as an internal control. Data are presented as the mean ± standard deviation (SD) of at least three independent experiments (n = 3). * p < 0.05, ** p < 0.01 versus treated control.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hyun, S.B.; Hyun, C.-G. Anti-Inflammatory Effects and Their Correlation with Microbial Community of Shindari, a Traditional Jeju Beverage. Fermentation 2020, 6, 87. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030087
AMA Style
Hyun SB, Hyun C-G. Anti-Inflammatory Effects and Their Correlation with Microbial Community of Shindari, a Traditional Jeju Beverage. Fermentation. 2020; 6(3):87. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030087
Chicago/Turabian StyleHyun, Su Bin, and Chang-Gu Hyun. 2020. "Anti-Inflammatory Effects and Their Correlation with Microbial Community of Shindari, a Traditional Jeju Beverage" Fermentation 6, no. 3: 87. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030087
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.