
Manuka Honey with Varying Levels of Active Manuka Factor (AMF) Ratings as an Anaerobic Fermentation Substrate for Limosilactobacillus reuteri DPC16


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Strains, Media and Chemicals
2.2. Anaerobic Batch Fermentation
2.3. Biomass Growth
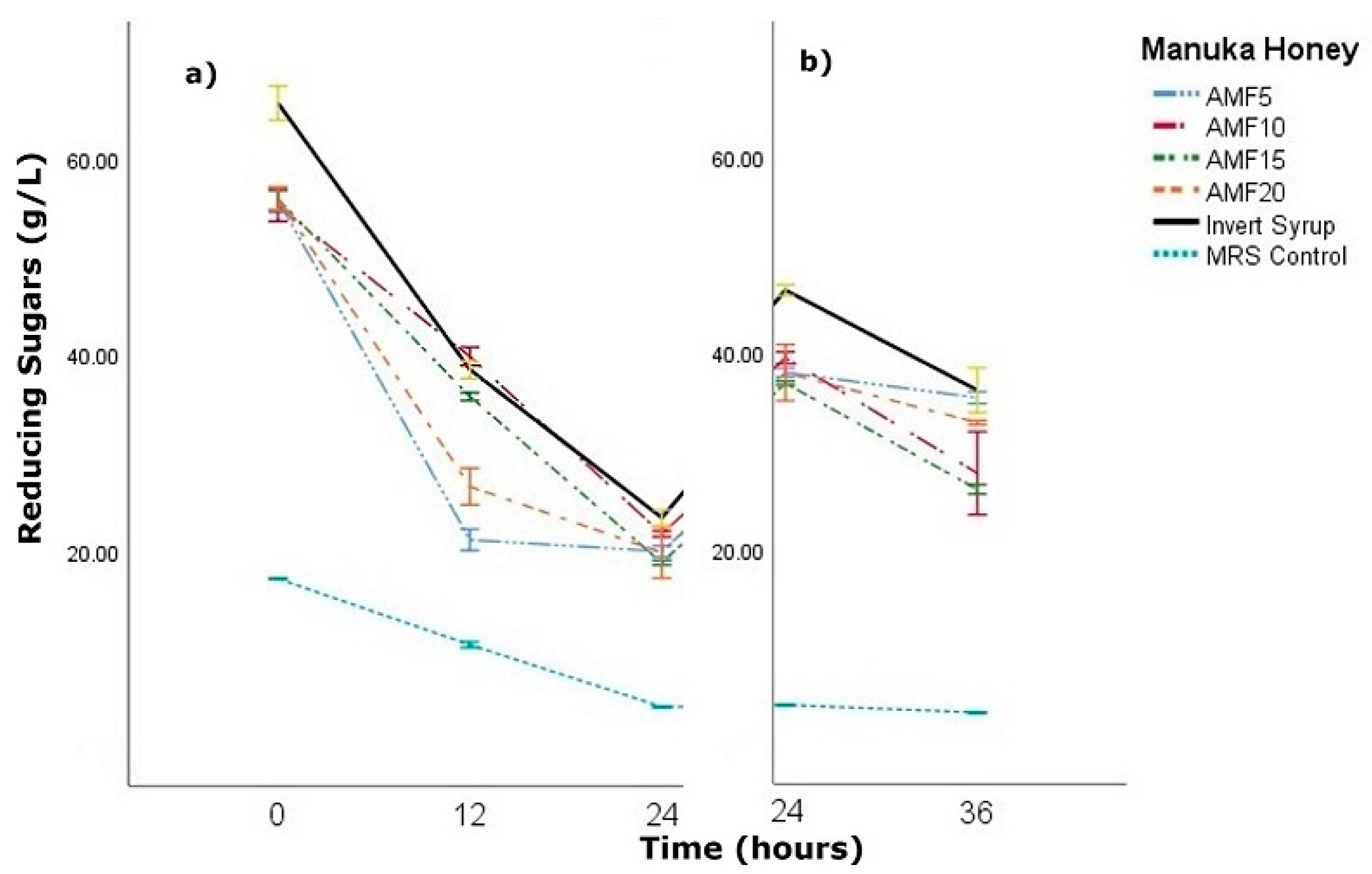
2.4. Reducing Sugar Quantification
2.5. Quantification of Organic Acid Metabolites and Honey Sugars
2.6. Statistical Analysis
3. Results and Discussion
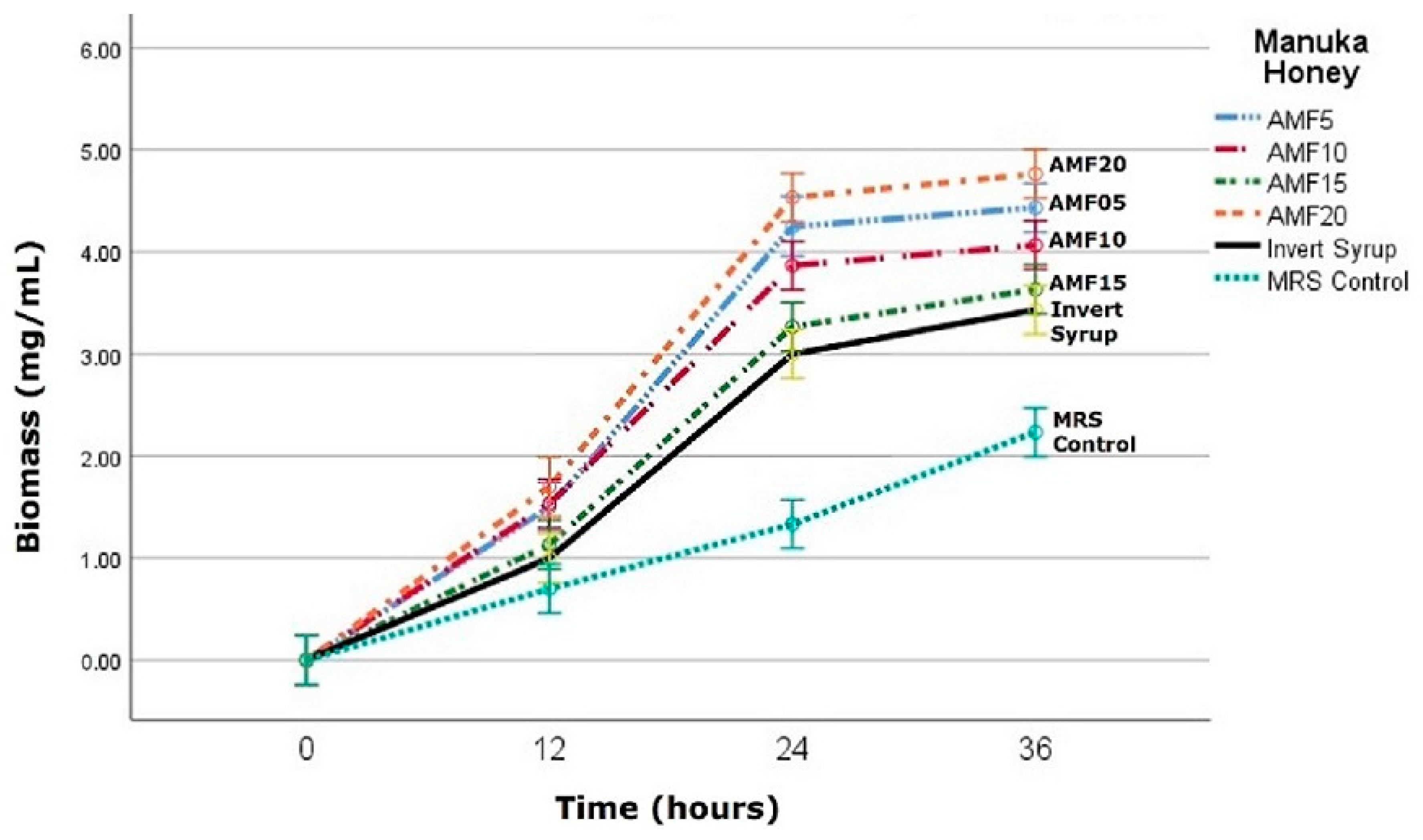
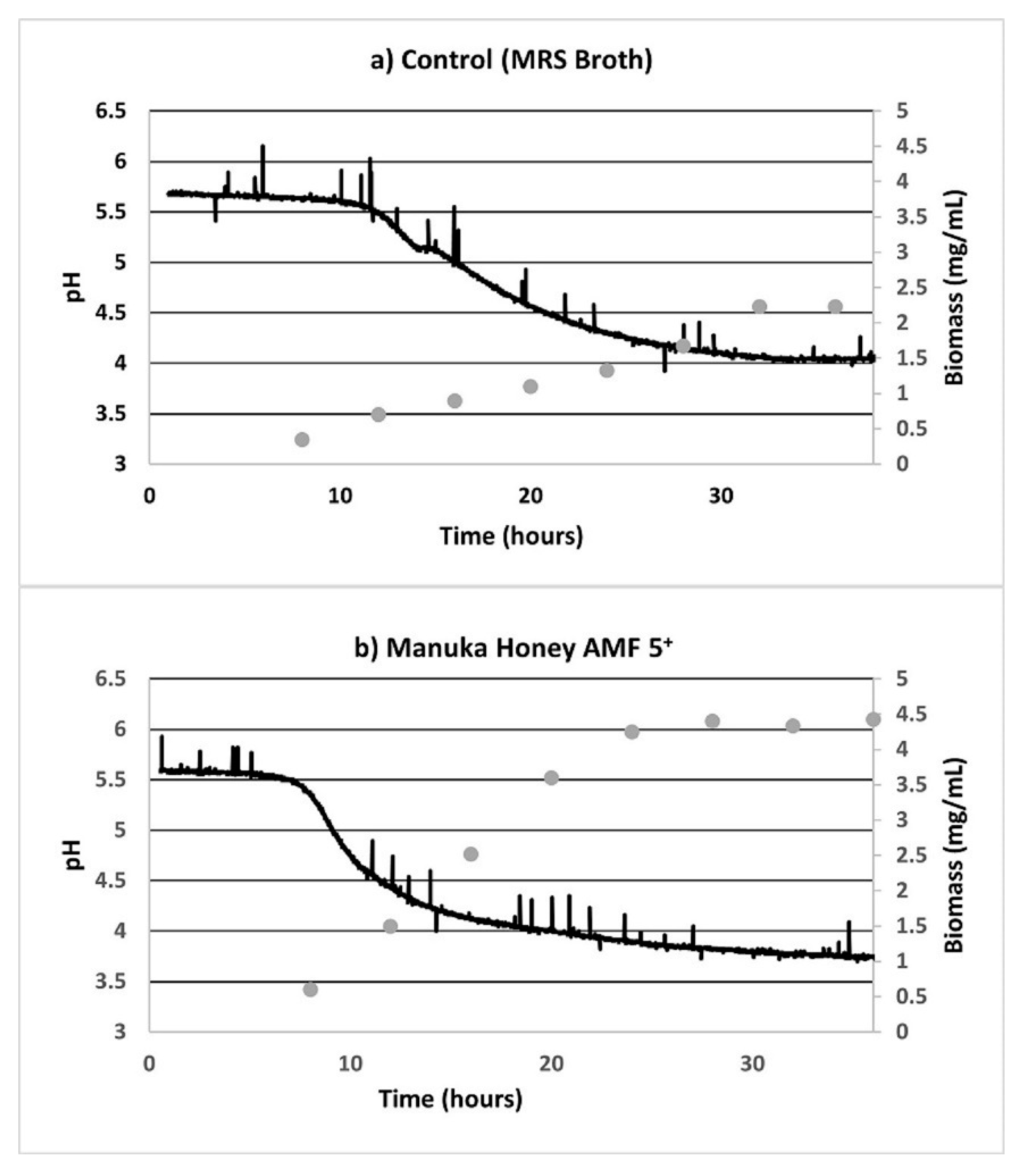
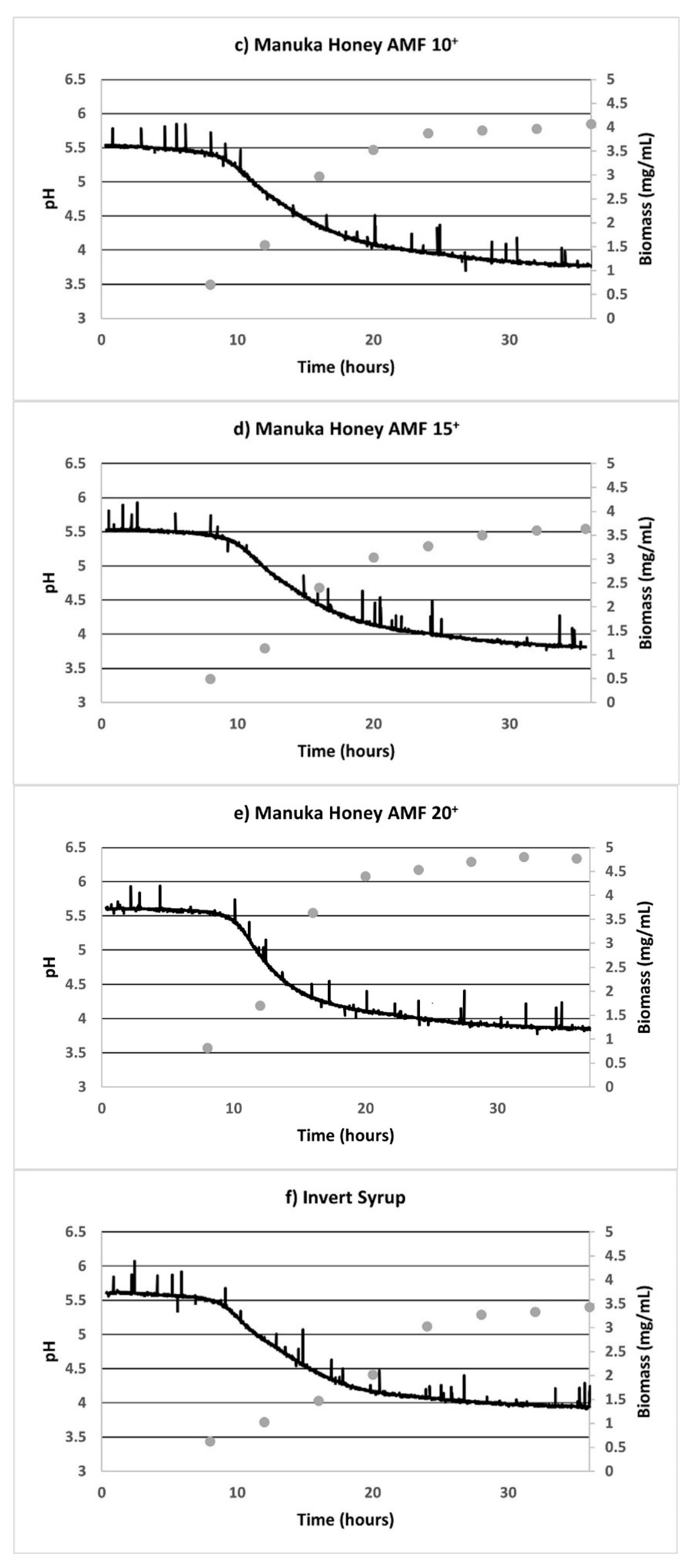
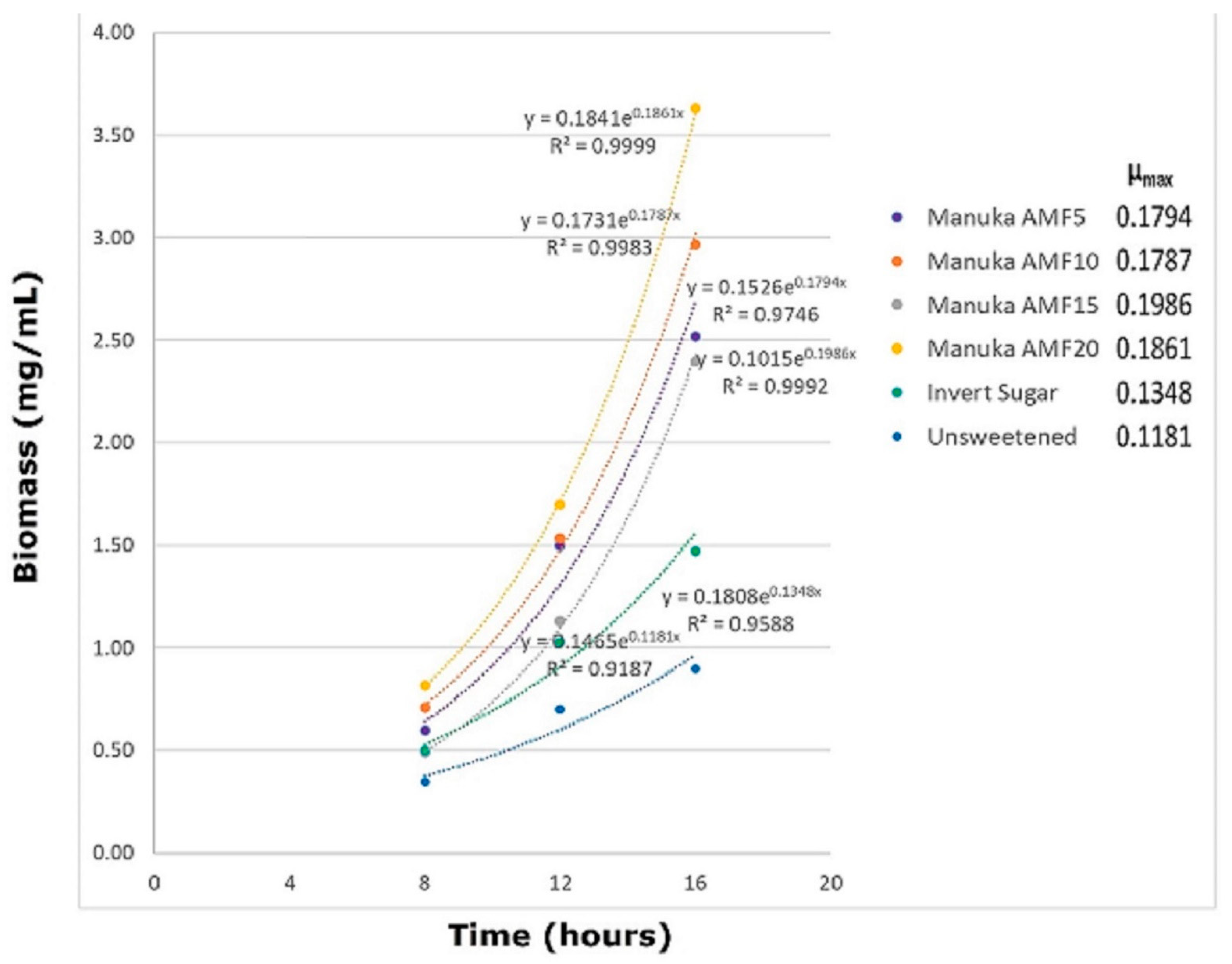
3.1. Substrate Consumption and Biomass Growth
3.2. Fermentation Plots
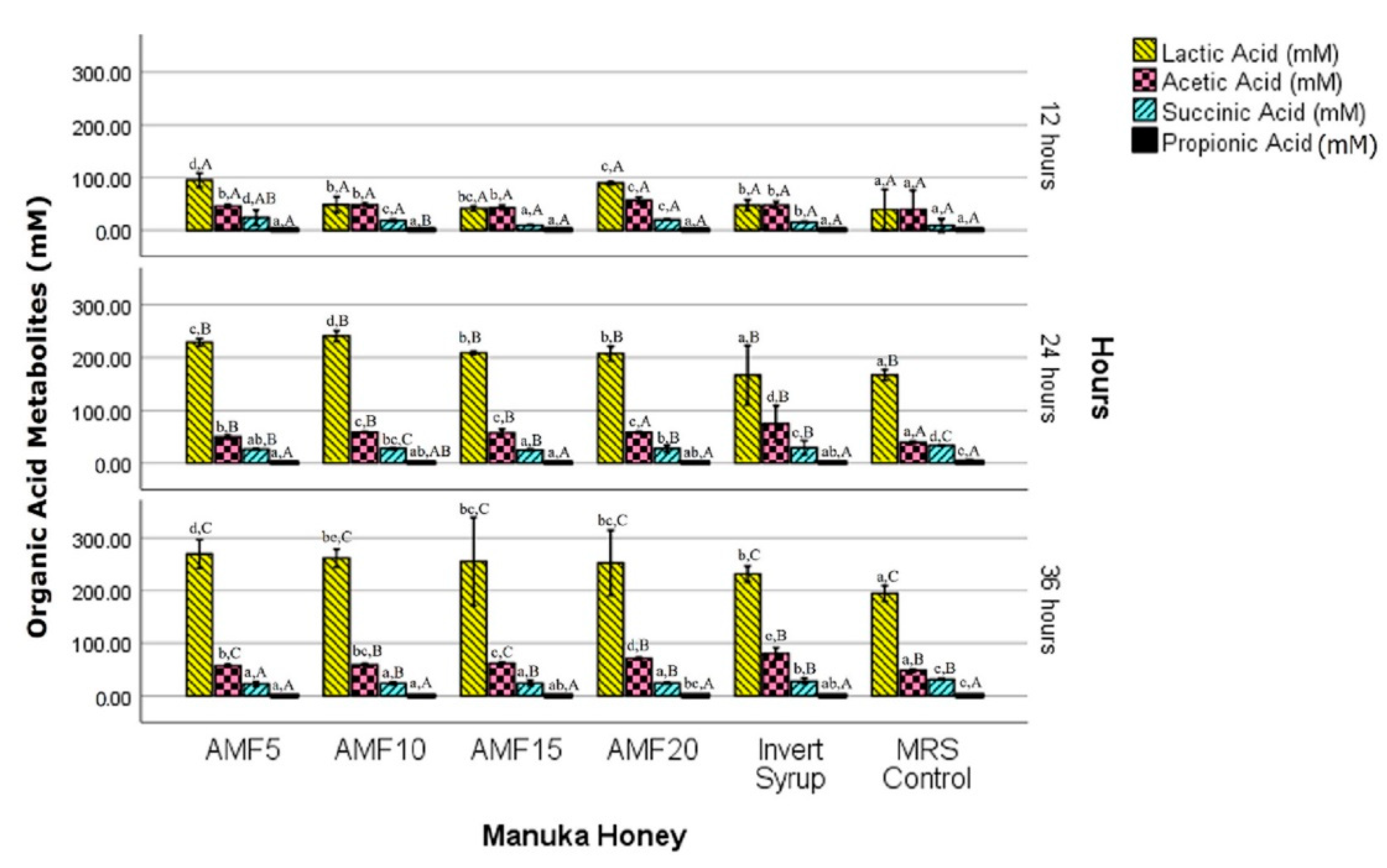
3.3. Beneficial Fermentation Metabolites
3.3.1. Lactic Acid
3.3.2. Acetic Acid
3.3.3. Succinic Acid
3.3.4. Propionic Acid
3.3.5. Butyric Acid
3.4. Honey Sugars Quantification
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wichienchot, S.; He, X.; Fu, X.; Huang, Q.; Zhang, B. In Vitro colonic fermentation of dietary fibers: Fermentation rate, short-chain fatty acid production and changes in microbiota. Trends Food Sci. Technol. 2019, 88, 1–9. [Google Scholar] [CrossRef]
- Asarat, M.; Vasiljevic, T.; Ravikumar, M.; Apostolopoulos, V.; Donkor, O. Extraction and Purification of Short-chain Fatty Acids from Fermented Reconstituted Skim Milk Supplemented with Inulin. Food Anal. Methods. 2016, 9, 3069–3079. [Google Scholar] [CrossRef]
- Wong, J.M.W.; De Souza, R.; Kendall, C.W.C.; Emam, A.; Jenkins, D.J.A. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gibson, G.R. Effects of the In Vitro fermentation of oligofructose and inulin by bacteria growing in the human large intestine. J. Appl. Bacteriol. 1993, 75, 373–380. [Google Scholar] [CrossRef]
- Shamala, T.R.; Shri Jyothi, Y.; Saibaba, P. Stimulatory effect of honey on multiplication of lactic acid bacteria under in vitro and in vivo conditions. Lett. Appl. Microbiol. 2000, 30, 453–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalán, Z.; Hudáček, J.; Štětina, J.; Chumchalová, J.; Halász, A. Production of organic acids by Lactobacillus strains in three different media. Eur. Food Res. Technol. 2010, 230, 395–404. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.M.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Bian, L.; Molan, A.L.; Maddox, I.; Shu, Q. Antimicrobial activity of Lactobacillus reuteri DPC16 supernatants against selected food borne pathogens. World J. Microbiol. Biotechnol. 2011, 27, 991–998. [Google Scholar] [CrossRef]
- Jan Mei, S.; Mohd Nordin, M.S.; Norrakiah, A.S. Fructooligosaccharides in honey and effects of honey on growth of Bifidobacterium longum BB 536. Int. Food Res. J. 2010, 17, 557–561. [Google Scholar]
- Mohan, A.; Quek, S.Y.; Gutierrez-Maddox, N.; Gao, Y.; Shu, Q. Effect of honey in improving the gut microbial balance. Food Qual. Saf. 2017, 1, 107–115. [Google Scholar] [CrossRef]
- Sanz, M.L.; Polemis, N.; Morales, V.; Corzo, N.; Drakoularakou, A.; Gibson, G.R.; Rastall, R.A. In Vitro investigation into the potential prebiotic activity of honey oligosaccharides. J. Agric. Food Chem. 2005, 53, 2914–2921. [Google Scholar] [CrossRef]
- Weston, R.J.; Brocklebank, L.K. The oligosaccharide composition of some New Zealand honeys. Food Chem. 1999, 64, 33–37. [Google Scholar] [CrossRef]
- Molan, P.C.; Russell, K.M. Non-peroxide antibacterial activity in some New Zealand honeys. J. Apic. Res. 1988, 27, 62–67. [Google Scholar] [CrossRef]
- Girma, A.; Seo, W.; SheI, R.C. Antibacterial activity of varying UMF-graded Manuka honeys. PLoS ONE 2019, 14, e0224495. [Google Scholar] [CrossRef] [Green Version]
- Mohan, A.; Hadi, J.; Gutierrez-Maddox, N.; Li, Y.; Leung, I.K.H.; Gao, Y.; Shu, Q.; Quek, S. Sensory, Microbiological and Physicochemical Characterisation of Functional Manuka Honey Yogurts Containing Probiotic Lactobacillus reuteri DPC16. Foods 2020, 9, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drapac Drapac DrKiwi AMF Grading System. Available online: http://www.drapac.co.nz/News/Cover?categoryid=37 (accessed on 3 January 2020).
- Cai, J.; Zhang, J.; Tian, Y.; Zhang, L.; Hatzakis, E.; Krausz, K.W.; Smith, P.B.; Gonzalez, F.J.; Patterson, A.D. Orthogonal Comparison of GC-MS and 1H NMR Spectroscopy for Short Chain Fatty Acid Quantitation. Anal. Chem. 2017, 89, 7900–7906. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Furihata, K.; Ito-Ishida, M.; Kaminogawa, S.; Tanokura, M. Nondestructive observation of bovine milk by NMR spectroscopy: Analysis of existing states of compounds and detection of new compounds. J. Agric. Food Chem. 2004, 52, 4969–4974. [Google Scholar] [CrossRef]
- Schievano, E.; Tonoli, M.; Rastrelli, F. NMR Quantification of Carbohydrates in Complex Mixtures. A Challenge on Honey. Anal. Chem. 2017, 89, 13405–13414. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Furihata, K.; Kato, Y.; Tanokura, M. Nondestructive quantification of organic compounds in whole milk without pretreatment by two-dimensional NMR spectroscopy. J. Agric. Food Chem. 2007, 55, 4307–4311. [Google Scholar] [CrossRef] [PubMed]
- Petersen, B.O.; Hindsgaul, O.; Meier, S. Profiling of carbohydrate mixtures at unprecedented resolution using high-precision 1H-13C chemical shift measurements and a reference library. Analyst 2013, 139, 401–406. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Wu, P.S.C.; Otting, G. Rapid pulse length determination in high-resolution NMR. J. Magn. Reson. 2005, 176, 115–119. [Google Scholar] [CrossRef] [PubMed]
- De Melo, F.H.C.; Menezes, F.N.D.D.; de Sousa, J.M.B.; dos Santos Lima, M.; da Silva Campelo Borges, G.; de Souza, E.L.; Magnani, M. Prebiotic activity of monofloral honeys produced by stingless bees in the semi-arid region of Brazilian Northeastern toward Lactobacillus acidophilus LA-05 and Bifidobacterium lactis BB-12. Food Res. Int. 2020, 128, 108809. [Google Scholar] [CrossRef] [PubMed]
- Nutter, J.; Fritz, R.; Iurlina, M.O.; Saiz, A.I. Effect of Prosopis sp. honey on the growth and fermentative ability of Pediococcus pentosaceus and Lactobacillus fermentum. LWT Food Sci. Technol. 2016, 70, 309–314. [Google Scholar] [CrossRef]
- Nsogning, S.D.; Fischer, S.; Becker, T. Investigating on the fermentation behavior of six lactic acid bacteria strains in barley malt wort reveals limitation in key amino acids and buffer capacity. Food Microbiol. 2018, 73, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Valdés-Varela, L.; Ruas-Madiedo, P.; Gueimonde, M. In Vitro fermentation of different fructo-oligosaccharides by Bifidobacterium strains for the selection of synbiotic combinations. Int. J. Food Microbiol. 2017, 242, 19–23. [Google Scholar] [CrossRef]
- Bourriaud, C.; Robins, R.J.; Martin, L.; Kozlowski, F.; Tenailleau, E.; Cherbut, C.; Michel, C. Lactate is mainly fermented to butyrate by human intestinal microfloras but inter-individual variation is evident. J. Appl. Microbiol. 2005, 99, 201–212. [Google Scholar] [CrossRef]
- Bujna, E.; Farkas, N.A.; Tran, A.M.; Dam, M.S.; Nguyen, Q.D. Lactic acid fermentation of apricot juice by mono- and mixed cultures of probiotic Lactobacillus and Bifidobacterium strains. Food Sci. Biotechnol. 2018, 27, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, D.; Mital, B.K.; Garg, S.K. Utilization of sugars by Lactobacillus acidophilus strains. Int. J. Food Microbiol. 1990, 10, 51–57. [Google Scholar] [CrossRef]
- Hughes, S.A.; Shewry, P.R.; Gibson, G.R.; McCleary, B.V.; Rastall, R.A. In Vitro fermentation of oat and barley derived β-glucans by human faecal microbiota. FEMS Microbiol. Ecol. 2008, 64, 482–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Invert Syrup | Manuka AMF 5+ | Manuka AMF 10+ | Manuka AMF 15+ | Manuka AMF 20+ | |
|---|---|---|---|---|---|
| Fructose | 35.55 ± 0.67 b | 31.66 ± 0.74 a | 32.73 ± 0.68 a | 32.2 ± 0.51 a | 34.86 ± 0.48 b |
| Glucose | 27.05 ± 1.05 a | 25.94 ± 0.88 a | 26.84 ± 0.71 a | 26.38 ± 0.52 a | 30.79 ± 0.59 b |
| Sucrose | 2.33 ± 0.11 b | 1.45 ± 0.01 a | 1.5 ± 0.01 a | 1.47 ± 0.01 a | 1.43 ± 0.03 a |
| Maltose | n.d. | 2.48 ± 0.61 a | 2.65 ± 0.73 a | 2.52 ± 0.41 a | 2.35 ± 0.39 a |
| Nigerose | n.d. | 2.87 ± 0.54 a | 2.45 ± 0.64 a | 2.53 ± 0.28 a | 2.43 ± 0.15 a |
| Kojibiose | n.d. | 1.44 ± 0.21 a | 1.57 ± 0.14 a | 1.74 ± 0.22 a | 1.57 ± 0.04 a |
| Panose | n.d. | 1.29 ± 0.36 a | 1.41 ± 0.72 a | 1.39 ± 0.59 a | 1.66 ± 0.81 a |
| Total Di- and Oligosaccharides | 2.33 ± 0.11 | 9.59 ± 2.75 | 9.88 ± 2.85 | 9.75 ± 2.79 | 10.73 ± 3.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohan, A.; Gutierrez-Maddox, N.; Meng, T.; He, N.; Gao, Y.; Shu, Q.; Quek, S.Y. Manuka Honey with Varying Levels of Active Manuka Factor (AMF) Ratings as an Anaerobic Fermentation Substrate for Limosilactobacillus reuteri DPC16. Fermentation 2021, 7, 128. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030128
Mohan A, Gutierrez-Maddox N, Meng T, He N, Gao Y, Shu Q, Quek SY. Manuka Honey with Varying Levels of Active Manuka Factor (AMF) Ratings as an Anaerobic Fermentation Substrate for Limosilactobacillus reuteri DPC16. Fermentation. 2021; 7(3):128. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030128
Chicago/Turabian StyleMohan, Anand, Noemi Gutierrez-Maddox, Tong Meng, Ning He, Yihuai Gao, Quan Shu, and Siew Young Quek. 2021. "Manuka Honey with Varying Levels of Active Manuka Factor (AMF) Ratings as an Anaerobic Fermentation Substrate for Limosilactobacillus reuteri DPC16" Fermentation 7, no. 3: 128. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030128