
Lychee-Derived, Thermotolerant Yeasts for Second-Generation Bioethanol Production
 ,
,  and
and
Abstract
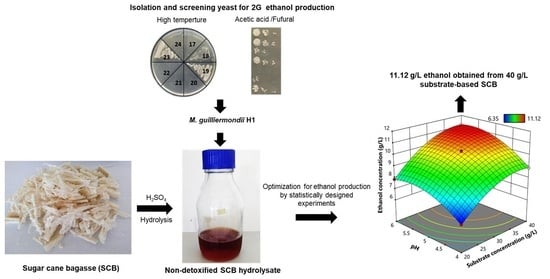
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Collection, Yeast Isolation and Thermotolerant Yeasts Screening
2.3. Glucose-Based Bioethanol Production by Selected Thermotolerant Yeasts
2.4. DNA Sequencing and Yeast Identification
2.5. Screening of the Thermotolerant Yeasts for Second-Generation Bioethanol Production
2.5.1. Tolerance of Thermotolerant Yeasts to Individual Stress Factors
2.5.2. Fermentation of Sugars
2.6. Optimization for Bioethanol Production from SCB Hydrolysate by Statistical Experimental Design
2.6.1. Plackett–Burman (PB) Design
2.6.2. Response Surface Methodology
2.7. Analytical Methods
3. Results and Discussion
3.1. Yeast Isolation and Thermotolerant Screening
3.2. Ethanol Fermentation Using Thermotolerant Yeasts
3.3. Identification of Selected Thermotolerant Yeasts
3.4. Selection of Superior Yeast Strains for Second-Generation Bioethanol Production
3.4.1. Tolerance of Thermotolerant Yeasts to Individual Stress Factors
3.4.2. Selected Yeast Fermenting Different Sugars
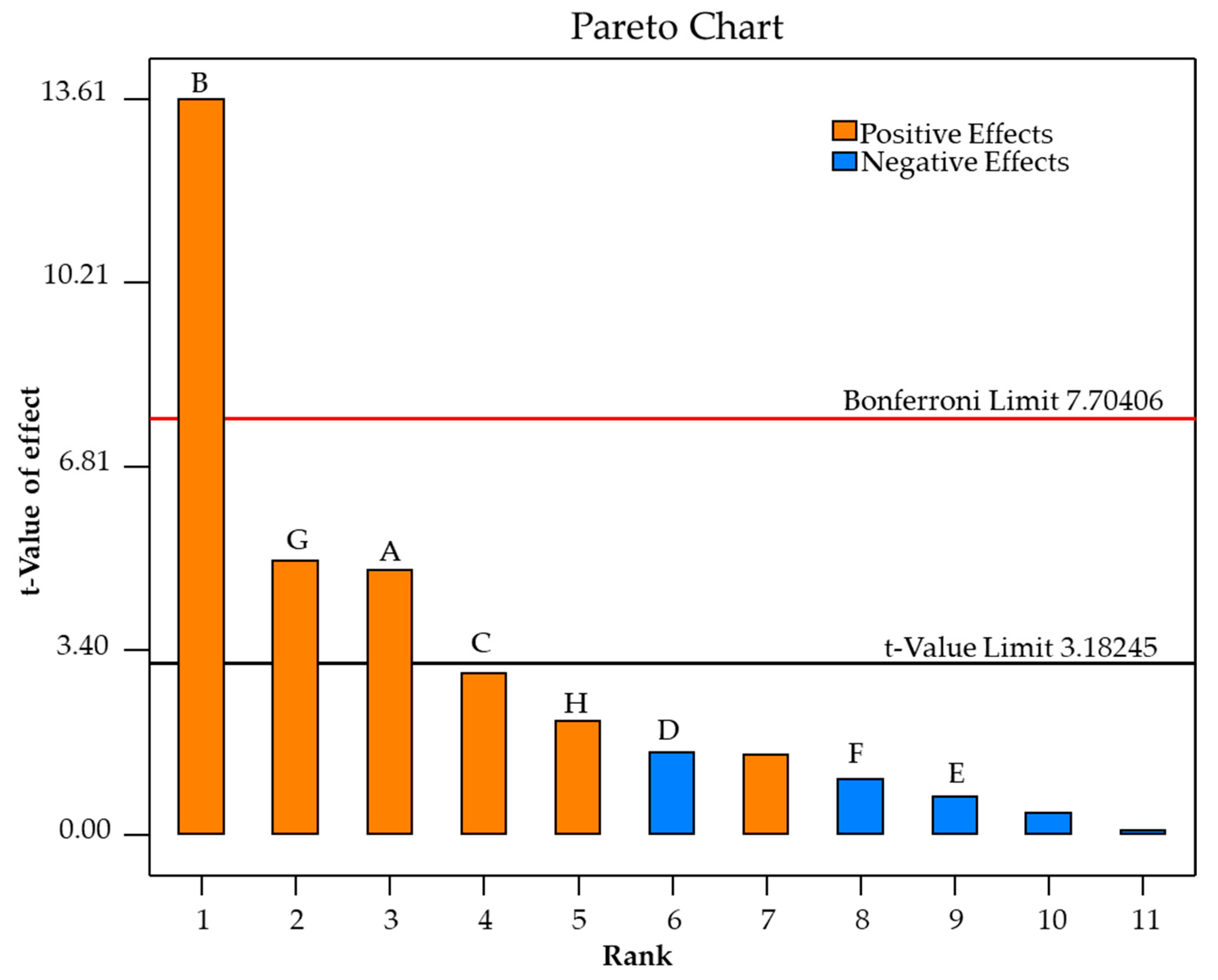
3.5. Screening Factors Affecting Ethanol Production from SCB Hydrolysate Using M. guilliermondii H1 Strain by Plackett–Burman Design
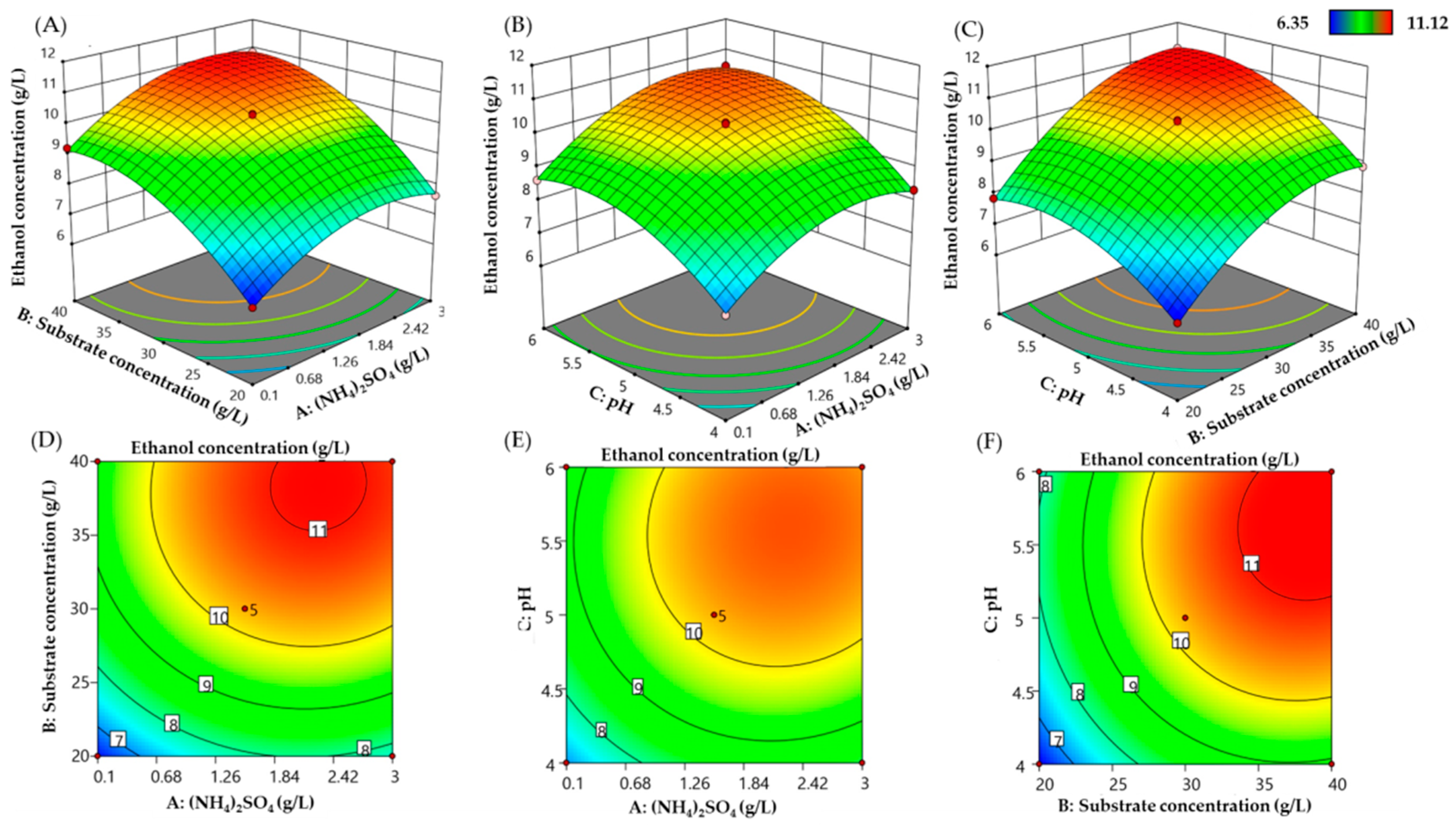
3.6. Optimization of Ethanol Production from SCB Hydrolysate by M. guilliermondii H1 Strain Using Box–Behnken Design
3.7. Validation of the Model
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bai, F.W.; Anderson, W.A.; Moo-Young, M. Ethanol Fermentation Technologies from Sugar and Starch Feedstocks. Biotechnol. Adv. 2008, 26, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Farrell, A.E.; Plevin, R.J.; Turner, B.T.; Jones, A.D.; O’hare, M.; Kammen, D.M. Ethanol Can Contribute to Energy and Environmental Goals. Science 2006, 311, 506–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harun, R.; Danquah, M.K.; Forde, G.M. Microalgal Biomass as a Fermentation Feedstock for Bioethanol Production. J. Chem. Technol. Biotechnol. 2010, 85, 199–203. [Google Scholar] [CrossRef]
- Christensen, E.; Yanowitz, J.; Ratcliff, M.; McCormick, R.L. Renewable Oxygenate Blending Effects on Gasoline Properties. Energy Fuels 2011, 25, 4723–4733. [Google Scholar] [CrossRef]
- Månsson, A.; Johansson, B.; Nilsson, L.J. Assessing Energy Security: An Overview of Commonly Used Methodologies. Energy 2014, 73, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Carrillo-Nieves, D.; Alanís, M.J.R.; de la Cruz Quiroz, R.; Ruiz, H.A.; Iqbal, H.M.N.; Parra-Saldívar, R. Current Status and Future Trends of Bioethanol Production from Agro-Industrial Wastes in Mexico. Renew. Sustain. Energy Rev. 2019, 102, 63–74. [Google Scholar] [CrossRef]
- Ho, D.P.; Ngo, H.H.; Guo, W. A Mini Review on Renewable Sources for Biofuel. Bioresour. Technol. 2014, 169, 742–749. [Google Scholar] [CrossRef] [Green Version]
- Canilha, L.; Chandel, A.K.; Suzane dos Santos Milessi, T.; Antunes, F.A.F.; Luiz da Costa Freitas, W.; das Graças Almeida Felipe, M.; da Silva, S.S. Bioconversion of Sugarcane Biomass into Ethanol: An Overview about Composition, Pretreatment Methods, Detoxification of Hydrolysates, Enzymatic Saccharification, and Ethanol Fermentation. J. Biomed. Biotechnol. 2012, 2012, 989572. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, B.G.; Moutta, R.d.O.; Ferraz, F.d.O.; Vieira, E.R.; Nogueira, A.S.; Baratella, B.F.; Rodrigues, L.C.; Hou-Rui, Z.; Da Silva, S.S. Biological Detoxification of Different Hemicellulosic Hydrolysates Using Issatchenkia Occidentalis CCTCC M 206097 Yeast. J. Ind. Microbiol. Biotechnol. 2011, 38, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qin, Y.; Xu, J.; Zeng, W.; Zhang, Y.; Wang, W.; Wang, P. Crack Resistance Investigation of Mixtures with Reclaimed SBS Modified Asphalt Pavement Using the SCB and DSCT Tests. Constr. Build. Mater. 2020, 265, 120365. [Google Scholar] [CrossRef]
- Abdel-Banat, B.; Hoshida, H.; Ano, A.; Nonklang, S.; Akada, R. High-Temperature Fermentation: How Can Processes for Ethanol Production at High Temperatures Become Superior to the Traditional Process Using Mesophilic Yeast? Appl. Microbiol. Biotechnol. 2010, 85, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Ryabova, O.B.; Chmil, O.M.; Sibirny, A.A. Xylose and Cellobiose Fermentation to Ethanol by the Thermotolerant Methylotrophic Yeast Hansenula Polymorpha. FEMS Yeast Res. 2003, 4, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Urbina, H.; Blackwell, M. Multilocus Phylogenetic Study of the Scheffersomyces Yeast Clade and Characterization of the N-Terminal Region of Xylose Reductase Gene. PLoS ONE 2012, 7, e39128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouri, H.; Azin, M.; Mousavi, M.L. Xylan-Hydrolyzing Thermotolerant Candida tropicalis HNMA-1 for Bioethanol Production from Sugarcane Bagasse Hydrolysate. Ann. Microbiol. 2017, 67, 633–641. [Google Scholar] [CrossRef]
- Steensels, J.; Snoek, T.; Meersman, E.; Nicolino, M.P.; Voordeckers, K.; Verstrepen, K.J. Improving Industrial Yeast Strains: Exploiting Natural and Artificial Diversity. FEMS Microbiol. Rev. 2014, 38, 947–995. [Google Scholar] [CrossRef] [Green Version]
- Tanimura, A.; Kikukawa, M.; Yamaguchi, S.; Kishino, S.; Ogawa, J.; Shima, J. Direct Ethanol Production from Starch Using a Natural Isolate, Scheffersomyces Shehatae: Toward Consolidated Bioprocessing. Sci. Rep. 2015, 5, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sree, N.K.; Sridhar, M.; Suresh, K.; Banat, I.M.; Rao, L.V. Isolation of Thermotolerant, Osmotolerant, Flocculating Saccharomyces cerevisiae for Ethanol Production. Bioresour. Technol. 2000, 72, 43–46. [Google Scholar] [CrossRef]
- Techaparin, A.; Thanonkeo, P.; Klanrit, P. High-Temperature Ethanol Production Using Thermotolerant Yeast Newly Isolated from Greater Mekong Subregion. Braz. J. Microbiol. 2017, 48, 461–475. [Google Scholar] [CrossRef]
- Pongcharoen, P.; Chawneua, J.; Tawong, W. High Temperature Alcoholic Fermentation by New Thermotolerant Yeast Strains Pichia Kudriavzevii Isolated from Sugarcane Field Soil. Agric. Nat. Resour. 2018, 52, 511–518. [Google Scholar] [CrossRef]
- Milessi, T.S.S.; Antunes, F.A.F.; Chandel, A.K.; Silva, S.S. Rice Bran Extract: An Inexpensive Nitrogen Source for the Production of 2G Ethanol from Sugarcane Bagasse Hydrolysate. 3 Biotech 2013, 3, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.C.; Van Nguyen, P.; Truong, H. Heavy Metal Tolerance of Novel Papiliotrema Yeast Isolated from Vietnamese Mangosteen. Mycobiology 2020, 48, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Chamnipa, N.; Thanonkeo, S.; Klanrit, P.; Thanonkeo, P. The Potential of the Newly Isolated Thermotolerant Yeast Pichia Kudriavzevii RZ8-1 for High-Temperature Ethanol Production. Braz. J. Microbiol. 2018, 49, 378–391. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Robnett, C.J. Identification and Phylogeny of Ascomycetous Yeasts from Analysis of Nuclear Large Subunit (26S) Ribosomal DNA Partial Sequences. Antonie Van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef]
- Li, H.; Wu, M.; Xu, L.; Hou, J.; Guo, T.; Bao, X.; Shen, Y. Evaluation of Industrial Saccharomyces cerevisiae Strains as the Chassis Cell for Second-generation Bioethanol Production. Microb. Biotechnol. 2015, 8, 266–274. [Google Scholar] [CrossRef]
- Limtong, S.; Sringiew, C.; Yongmanitchai, W. Production of Fuel Ethanol at High Temperature from Sugar Cane Juice by a Newly Isolated Kluyveromyces Marxianus. Bioresour. Technol. 2007, 98, 3367–3374. [Google Scholar] [CrossRef] [PubMed]
- Pongcharoen, P. The Ability of Pichia Kudriavzevii to Tolerate Multiple Stresses Makes It Promising for Developing Improved Bioethanol Production Processes. Lett. Appl. Microbiol. 2022, 75, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Phong, H.X.; Klanrit, P.; Dung, N.T.P.; Yamada, M.; Thanonkeo, P. Isolation and Characterization of Thermotolerant Yeasts for the Production of Second-Generation Bioethanol. Ann. Microbiol. 2019, 69, 765–776. [Google Scholar] [CrossRef]
- Talukder, A.A.; Easmin, F.; Mahmud, S.A.; Yamada, M. Thermotolerant Yeasts Capable of Producing Bioethanol: Isolation from Natural Fermented Sources, Identification and Characterization. Biotechnol. Biotechnol. Equip. 2016, 30, 1106–1114. [Google Scholar] [CrossRef] [Green Version]
- Nuanpeng, S.; Thanonkeo, S.; Yamada, M.; Thanonkeo, P. Ethanol Production from Sweet Sorghum Juice at High Temperatures Using a Newly Isolated Thermotolerant Yeast Saccharomyces cerevisiae DBKKU Y-53. Energies 2016, 9, 253. [Google Scholar] [CrossRef] [Green Version]
- Koutinas, M.; Patsalou, M.; Stavrinou, S.; Vyrides, I. High Temperature Alcoholic Fermentation of Orange Peel by the Newly Isolated Thermotolerant Pichia Kudriavzevii KVMP 10. Lett. Appl. Microbiol. 2016, 62, 75–83. [Google Scholar] [CrossRef]
- Matos, I.T.S.R.; Cassa-Barbosa, L.A.; Galvão, R.; Nunes-Silva, C.G.; Astolfi Filho, S. Isolation, Taxonomic Identification and Investigation of the Biotechnological Potential of Wild-Type Meyerozyma guilliermondii Associated with Amazonian Termites Able to Ferment D-Xylose. Biosci. J. 2014, 30, 260–266. [Google Scholar]
- Martini, C.; Tauk-Tornisielo, S.M.; Codato, C.B.; Bastos, R.G.; Ceccato-Antonini, S.R. A Strain of Meyerozyma guilliermondii Isolated from Sugarcane Juice Is Able to Grow and Ferment Pentoses in Synthetic and Bagasse Hydrolysate Media. World J. Microbiol. Biotechnol. 2016, 32, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Gibson, B.R.; Lawrence, S.J.; Leclaire, J.P.R.; Powell, C.D.; Smart, K.A. Yeast Responses to Stresses Associated with Industrial Brewery Handling. FEMS Microbiol. Rev. 2007, 31, 535–569. [Google Scholar] [CrossRef] [Green Version]
- Favaro, L.; Basaglia, M.; Trento, A.; Van Rensburg, E.; García-Aparicio, M.; Van Zyl, W.H.; Casella, S. Exploring Grape Marc as Trove for New Thermotolerant and Inhibitor-Tolerant Saccharomyces cerevisiae Strains for Second-Generation Bioethanol Production. Biotechnol. Biofuels 2013, 6, 168. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T.; Robert, V. Methods for Isolation, Phenotypic Characterization and Maintenance of Yeasts. In The Yeasts; Elsevier: Amsterdam, The Netherlands, 2011; pp. 87–110. [Google Scholar]
- Myers, R.H.; Montgomery, D.C.; Anderson-Cook, C.M. Response Surface Methodology: Process and Product Optimization Using Designed Experiments; John Wiley & Sons: New York, NY, USA, 2016; ISBN 1118916034. [Google Scholar]
- Nissen, T.L.; Kielland-Brandt, M.C.; Nielsen, J.; Villadsen, J. Optimization of Ethanol Production in Saccharomyces cerevisiae by Metabolic Engineering of the Ammonium Assimilation. Metab. Eng. 2000, 2, 69–77. [Google Scholar] [CrossRef]
- Li, Z.; Wang, D.; Shi, Y.-C. Effects of Nitrogen Source on Ethanol Production in Very High Gravity Fermentation of Corn Starch. J. Taiwan Inst. Chem. Eng. 2017, 70, 229–235. [Google Scholar] [CrossRef]
- Yue, G.; Yu, J.; Zhang, X.; Tan, T. The Influence of Nitrogen Sources on Ethanol Production by Yeast from Concentrated Sweet Sorghum Juice. Biomass Bioenergy 2012, 39, 48–52. [Google Scholar] [CrossRef]
- Izmirlioglu, G.; Demirci, A. Enhanced Bio-Ethanol Production from Industrial Potato Waste by Statistical Medium Optimization. Int. J. Mol. Sci. 2015, 16, 24490–24505. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, D.; Suman, S.K.; Pandey, D.; Ghosh, D.; Khan, R.; Agrawal, D.; Jain, R.K.; Vadde, V.T.; Adhikari, D.K. Design and Optimization of Ethanol Production from Bagasse Pith Hydrolysate by a Thermotolerant Yeast Kluyveromyces Sp. IIPE453 Using Response Surface Methodology. SpringerPlus 2013, 2, 159. [Google Scholar] [CrossRef] [Green Version]
- Martiniano, S.E.; Chandel, A.K.; Soares, L.C.S.R.; Pagnocca, F.C.; da Silva, S.S. Evaluation of Novel Xylose-Fermenting Yeast Strains from Brazilian Forests for Hemicellulosic Ethanol Production from Sugarcane Bagasse. 3 Biotech 2013, 3, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.D.; Mussatto, S.I.; Cadete, R.M.; Rosa, C.A.; Silva, S.S. Ethanol Production by a New Pentose-fermenting Yeast Strain, Scheffersomyces Stipitis UFMG-IMH 43.2, Isolated from the Brazilian Forest. Yeast 2011, 28, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Canilha, L.; Carvalho, W.; de Almeida Felipe, M.d.G.; de Almeida e Silva, J.B.; Giulietti, M. Ethanol Production from Sugarcane Bagasse Hydrolysate Using Pichia Stipitis. Appl. Biochem. Biotechnol. 2010, 161, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Moodley, P.; Kana, E.B.G. Bioethanol Production from Sugarcane Leaf Waste: Effect of Various Optimized Pretreatments and Fermentation Conditions on Process Kinetics. Biotechnol. Rep. 2019, 22, e00329. [Google Scholar] [CrossRef] [PubMed]
- Narendranath, N.V.; Power, R. Relationship between PH and Medium Dissolved Solids in Terms of Growth and Metabolism of Lactobacilli and Saccharomyces cerevisiae during Ethanol Production. Appl. Environ. Microbiol. 2005, 71, 2239–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jargalsaikhan, O.; Saraçoğlu, N. Application of Experimental Design Method for Ethanol Production by Fermentation of Sunflower Seed Hull Hydrolysate Using Pichia Stipitis NRRL-124. Chem. Eng. Commun. 2008, 196, 93–103. [Google Scholar] [CrossRef]
- Cadete, R.M.; Melo, M.A.; Dussan, K.J.; Rodrigues, R.C.L.B.; Silva, S.S.; Zilli, J.E.; Vital, M.J.S.; Gomes, F.C.O.; Lachance, M.-A.; Rosa, C.A. Diversity and Physiological Characterization of D-Xylose-Fermenting Yeasts Isolated from the Brazilian Amazonian Forest. PLoS ONE 2012, 7, e43135. [Google Scholar] [CrossRef]
- Sasikumar, E.; Viruthagiri, T. Optimization of Process Conditions Using Response Surface Methodology (RSM) for Ethanol Production from Pretreated Sugarcane Bagasse: Kinetics and Modeling. BioEnergy Res. 2008, 1, 239–247. [Google Scholar] [CrossRef]
- Hande, A.; Mahajan, S.; Prabhune, A. Evaluation of Ethanol Production by a New Isolate of Yeast during Fermentation in Synthetic Medium and Sugarcane Bagasse Hemicellulosic Hydrolysate. Ann. Microbiol. 2013, 63, 63–70. [Google Scholar] [CrossRef]
- Kwon, Y.-J.; Ma, A.-Z.; Li, Q.; Wang, F.; Zhuang, G.-Q.; Liu, C.-Z. Effect of Lignocellulosic Inhibitory Compounds on Growth and Ethanol Fermentation of Newly-Isolated Thermotolerant Issatchenkia Orientalis. Bioresour. Technol. 2011, 102, 8099–8104. [Google Scholar] [CrossRef]
- Perna, M.d.S.C.; Bastos, R.G.; Ceccato-Antonini, S.R. Single and Combined Effects of Acetic Acid, Furfural, and Sugars on the Growth of the Pentose-Fermenting Yeast Meyerozyma Guilliermondii. 3 Biotech 2018, 8, 119. [Google Scholar] [CrossRef]
- Trichez, D.; Steindorff, A.S.; Soares, C.E.V.F.; Formighieri, E.F.; Almeida, J.R.M. Physiological and Comparative Genomic Analysis of New Isolated Yeasts Spathaspora Sp. JA1 and Meyerozyma Caribbica JA9 Reveal Insights into Xylitol Production. FEMS Yeast Res. 2019, 19, foz034. [Google Scholar] [CrossRef] [PubMed]
- Levin, L.; Herrmann, C.; Papinutti, V.L. Optimization of Lignocellulolytic Enzyme Production by the White-Rot Fungus Trametes Trogii in Solid-State Fermentation Using Response Surface Methodology. Biochem. Eng. J. 2008, 39, 207–214. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | 37 °C | 40 °C | 45 °C | ||||||
|---|---|---|---|---|---|---|---|---|---|
| P (g/L) | Qp (g/L∙h) | YP/S (g/g) | P (g/L) | Qp (g/L∙h) | YP/S (g/g) | P (g/L) | Qp (g/L∙h) | YP/S (g/g) | |
| H1 | 71.57 ± 0.43 b | 1.49 ± 0.03 a | 0.45 ± 0.02 c | 68.4 ± 0.33 ab | 1.43 ± 0.01 b | 0.43 ± 0.01 a | 48.25 ± 0.22 a | 1.01 ± 0.01 a | 0.30 ± 0.00 c |
| H8 | 62.46 ± 0.24 ac | 1.30 ± 0.04 a | 0.39 ± 0.01 ab | 58.05 ± 0.29 c | 1.21 ± 0.02 d | 0.36 ± 0.00 e | 13.62 ± 0.18 b | 0.28 ± 0.00 ab | 0.09 ± 0.00 ac |
| H10 | 51.33 ± 0.12 d | 1.07 ± 0.01 d | 0.32 ± 0.00 a | 47.96 ± 0.25 a | 1.00 ± 0.02 a | 0.30 ± 0.01 b | 18.07 ± 0.11 b | 0.38 ± 0.01 b | 0.11 ± 0.01 a |
| H12 | 56.14 ± 0.31 ab | 1.36 ± 0.02 bc | 0.35 ± 0.01 b | 53.2 ± 0.15 b | 1.11 ± 0.01 c | 0.33 ± 0.01 bc | 29.73 ± 0.3 de | 0.62 ± 0.02 d | 0.19 ± 0.01 ab |
| H18 | 52.58 ± 0.25 bc | 1.10 ± 0.01 d | 0.33 ± 0.00 bc | 48.62 ± 0.36 e | 0.99 ± 0.02 b | 0.30 ± 0.00 a | 16.28 ± 0.15 a | 0.34 ± 0.00 e | 0.10 ± 0.00 d |
| H19 | 72.19 ± 0.55 c | 1.50 ± 0.04 ab | 0.45 ± 0.01 d | 69.77 ± 0.42 c | 1.45 ± 0.01 d | 0.44 ± 0.01 ac | 50.42 ± 0.34 d | 1.05 ± 0.02 bc | 0.32 ± 0.01 e |
| H23 | 70.35 ± 0.62 ab | 1.47 ± 0.02 e | 0.44 ± 0.01 ab | 70.18 ± 0.37 e | 1.46 ± 0.01 a | 0.44 ± 0.00 ab | 55.83 ± 0.28 c | 1.16 ± 0.03 a | 0.35 ± 0.01 bc |
| Carbon Source | Yeast strain | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| H1 | H12 | M. guilliermondii a | H10 | H18 | S. cerevisiae b | H8 | H19 | H23 | C. tropicalis b | |
| Glucose | + | + | + | + | + | + | + | + | + | + |
| Fructose | + | + | + | + | + | + | + | + | + | + |
| Sucrose | + | + | + | + | + | − | + | + | + | − |
| Galactose | + | + | + | + | + | − | + | + | + | − |
| Maltose | + | + | + | + | + | − | + | + | + | − |
| Lactose | − | − | − | − | − | − | − | − | − | − |
| Xylose | + | + | + | − | − | − | − | − | − | − |
| Arabinose | + | + | + | − | − | − | − | − | − | − |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value | Note |
|---|---|---|---|---|---|---|
| Model | 34.54 | 8 | 4.32 | 31.46 | 0.008 | significant |
| A-(NH4)2SO4 | 3.28 | 1 | 3.28 | 23.87 | 0.016 | significant |
| B-Substrate con | 25.43 | 1 | 25.43 | 185.33 | 0.001 | significant |
| C-ZnSO4·7H2O | 1.22 | 1 | 1.22 | 8.91 | 0.058 | |
| D-MgSO4·7H2O | 0.30 | 1 | 0.30 | 2.22 | 0.233 | |
| E-MnSO4·H2O | 0.02 | 1 | 0.02 | 0.17 | 0.707 | |
| F-KH2PO4 | 0.14 | 1 | 0.14 | 1.04 | 0.382 | |
| G-pH | 3.53 | 1 | 3.53 | 25.74 | 0.015 | significant |
| H-CaCl2·2H2O | 0.60 | 1 | 0.60 | 4.39 | 0.127 | |
| Residual | 0.41 | 3 | 0.14 | |||
| Cor Total | 34.95 | 11 | ||||
| Std. Dev. | 0.370446 | R2 | 0.9882 | |||
| C.V. % | 4.578594 | Adjusted R2 | 0.9568 |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value | Note |
|---|---|---|---|---|---|---|
| Model | 39.77 | 9 | 4.42 | 1085.21 | 0.000 | significant |
| A-(NH4)2SO4 | 4.58 | 1 | 4.58 | 1123.56 | 0.000 | |
| B-Substrate con | 17.43 | 1 | 17.43 | 4281.41 | 0.000 | |
| C-pH | 7.11 | 1 | 7.11 | 1745.14 | 0.000 | |
| AB | 0.04 | 1 | 0.04 | 9.82 | 0.017 | |
| AC | 0.07 | 1 | 0.07 | 15.97 | 0.005 | |
| BC | 0.14 | 1 | 0.14 | 34.53 | 0.001 | |
| A2 | 3.00 | 1 | 3.00 | 736.54 | 0.000 | |
| B2 | 3.44 | 1 | 3.44 | 844.99 | 0.000 | |
| C2 | 2.88 | 1 | 2.88 | 706.32 | 0.000 | |
| Residual | 0.03 | 7 | 0.00 | |||
| Lack of Fit | 0.02 | 3 | 0.01 | 3.37 | 0.1356 | not significant |
| Pure Error | 0.01 | 4 | 0.002 | |||
| Cor Total | 39.80 | 16 | ||||
| Std. Dev. | 0.0638 | R2 | 0.9993 | |||
| C.V. % | 0.7038 | Adjusted R2 | 0.9984 |
| Initial Concentration (g/L) | Final Concentration (g/L) | Ethanol Concentration (g/L) | Product Yield (YP/S, g/g) | |
|---|---|---|---|---|
| Total sugar | 40 ± 0.5 a | 2.83 ± 0.1 bc | 11.13 ± 0.062 b | 0.345 |
| Xylose | 32 | 2.45 ± 0.03 c | ||
| Glucose | 5 | nd | ||
| Arabinose | 3.14 ± 0.094 b | 0.28 ± 0.03 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, P.V.; Nguyen, K.H.V.; Nguyen, N.L.; Ho, X.T.T.; Truong, P.H.; Thi Nguyen, K.C. Lychee-Derived, Thermotolerant Yeasts for Second-Generation Bioethanol Production. Fermentation 2022, 8, 515. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8100515
Nguyen PV, Nguyen KHV, Nguyen NL, Ho XTT, Truong PH, Thi Nguyen KC. Lychee-Derived, Thermotolerant Yeasts for Second-Generation Bioethanol Production. Fermentation. 2022; 8(10):515. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8100515
Chicago/Turabian StyleNguyen, Phu Van, Khanh Hoang Viet Nguyen, Ngoc Linh Nguyen, Xuan Tuy Thi Ho, Phuc Hung Truong, and Kim Cuc Thi Nguyen. 2022. "Lychee-Derived, Thermotolerant Yeasts for Second-Generation Bioethanol Production" Fermentation 8, no. 10: 515. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8100515