
Production of Exopolysaccharides by Cultivation of Halotolerant Bacillus atrophaeus BU4 in Glucose- and Xylose-Based Synthetic Media and in Hydrolysates of Quinoa Stalks

 ,
,  , , , , , ,
, , , , , , 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Characterization of the Bacterial Isolate
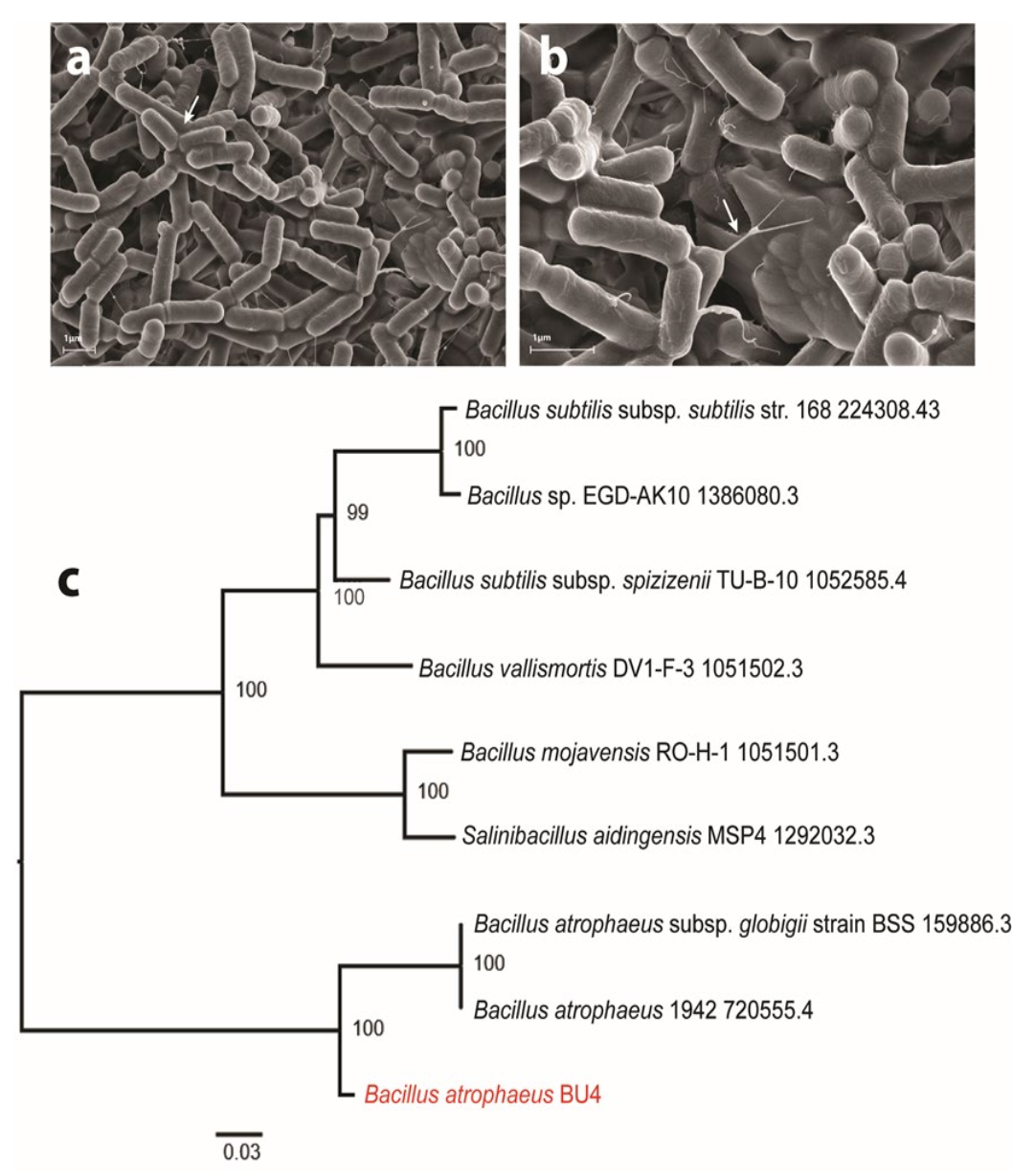
2.2.1. SEM Analysis
2.2.2. Genome Sequencing
2.3. Raw Material
2.4. Preparation of the Hydrolysates
2.4.1. Hemicellulosic Hydrolysate
2.4.2. Cellulosic Hydrolysate
2.5. Cultivation of Bacteria for EPS Production
2.5.1. Media
2.5.2. Cultivation
2.5.3. Analysis of the Cultivation Samples
2.6. Isolation and Purification of the EPS
2.7. EPS Characterization
2.7.1. Determination of the Monosaccharide Composition
2.7.2. High-Performance Size Exclusion Chromatography (HPSEC)
2.7.3. Fourier-Transform Infrared (FTIR) Spectroscopy
2.7.4. Two-Dimensional 1H-13C Heteronuclear Single-Quantum Coherence (HSQC) Nuclear Magnetic Resonance (NMR) Spectroscopy
2.7.5. Scanning Electron Microscopy (SEM)
2.7.6. X-ray Diffraction (XRD)
2.7.7. Thermogravimetric Analysis (TGA)
3. Results and Discussion
3.1. Characterization of the Bacterial Isolate
3.2. Production of Exopolysaccharides
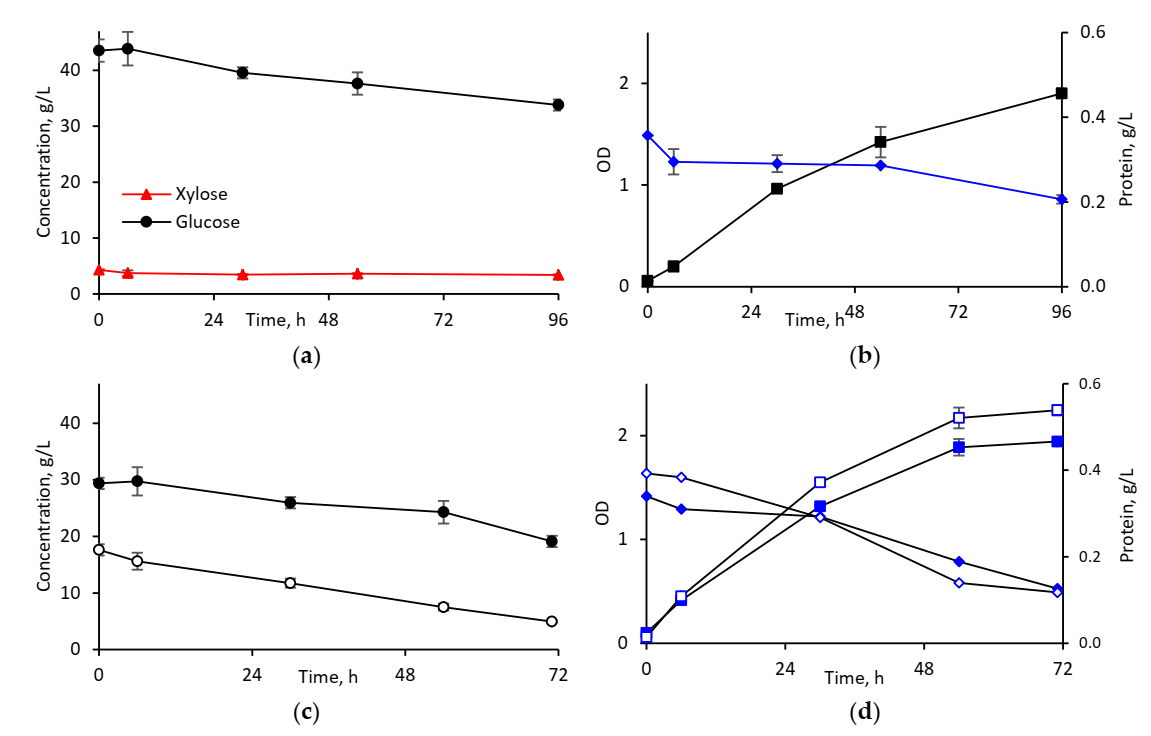
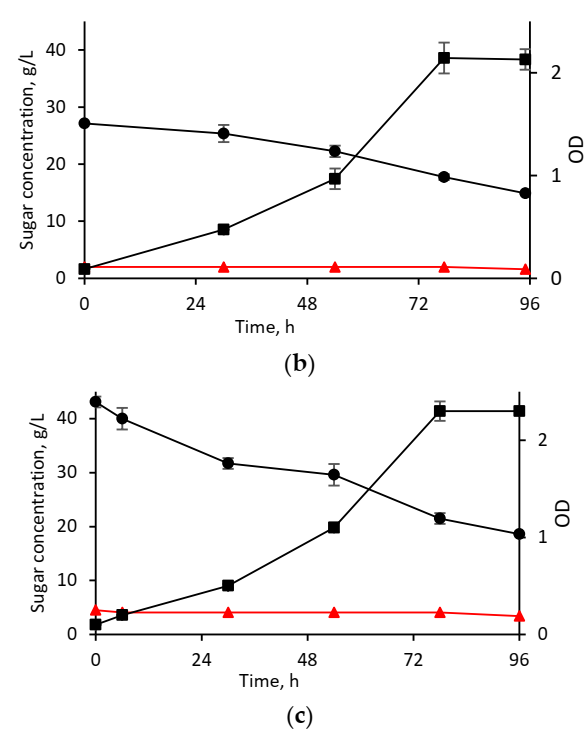
3.2.1. Cultivation in Synthetic Media
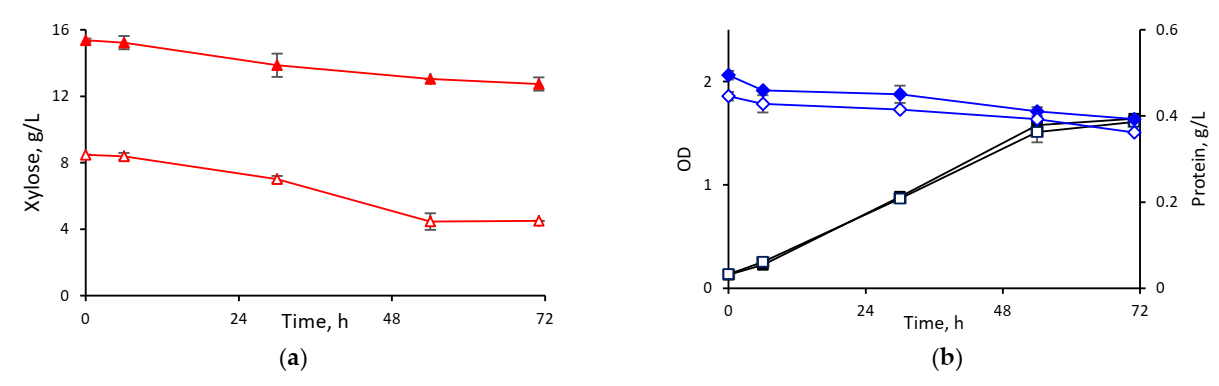
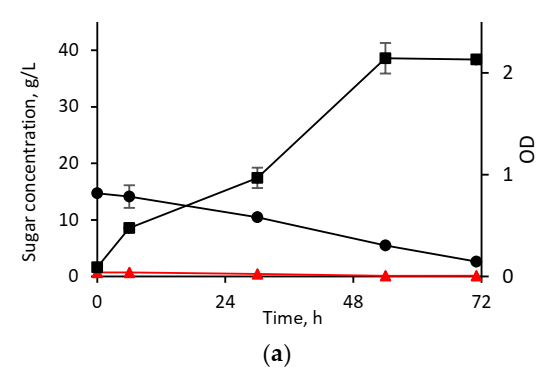
3.2.2. Cultivation in Hydrolysates
3.3. EPS Characterization
3.3.1. Determination of the Monosaccharide Composition
3.3.2. High-Performance Size Exclusion Chromatography (HPSEC)
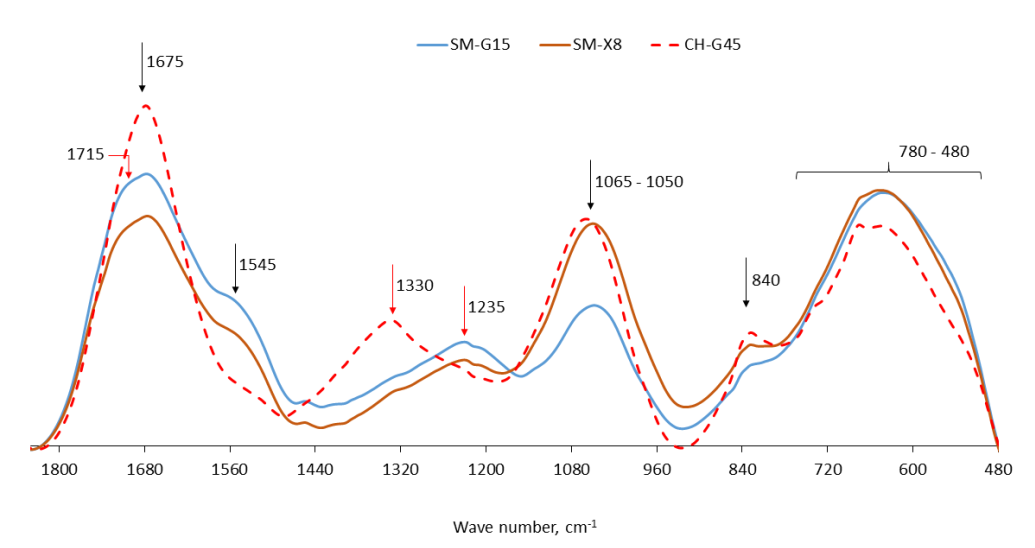
3.3.3. FTIR Spectra
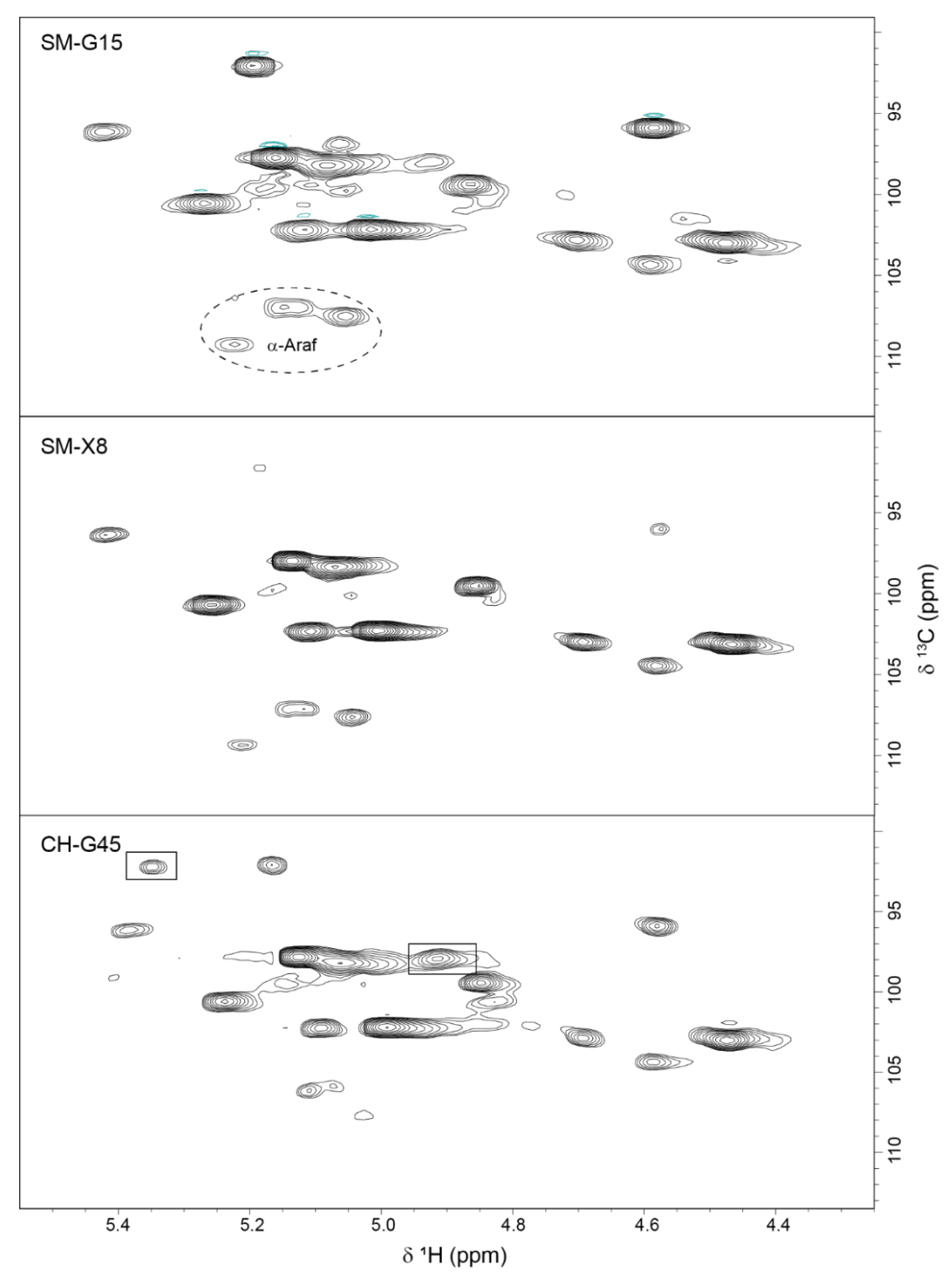
3.3.4. 2D 1H-13C HSQC NMR Spectra
3.3.5. SEM Imaging
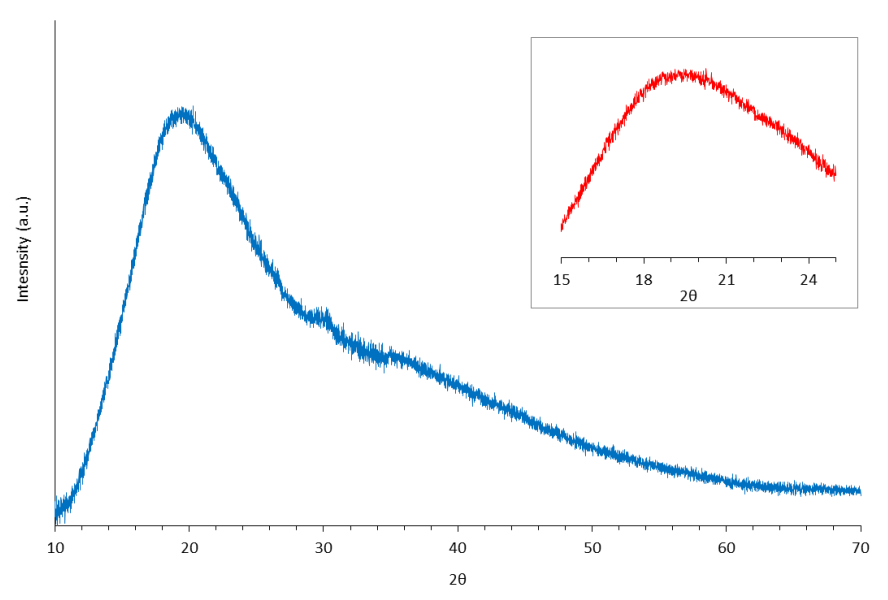
3.3.6. X-ray Diffraction
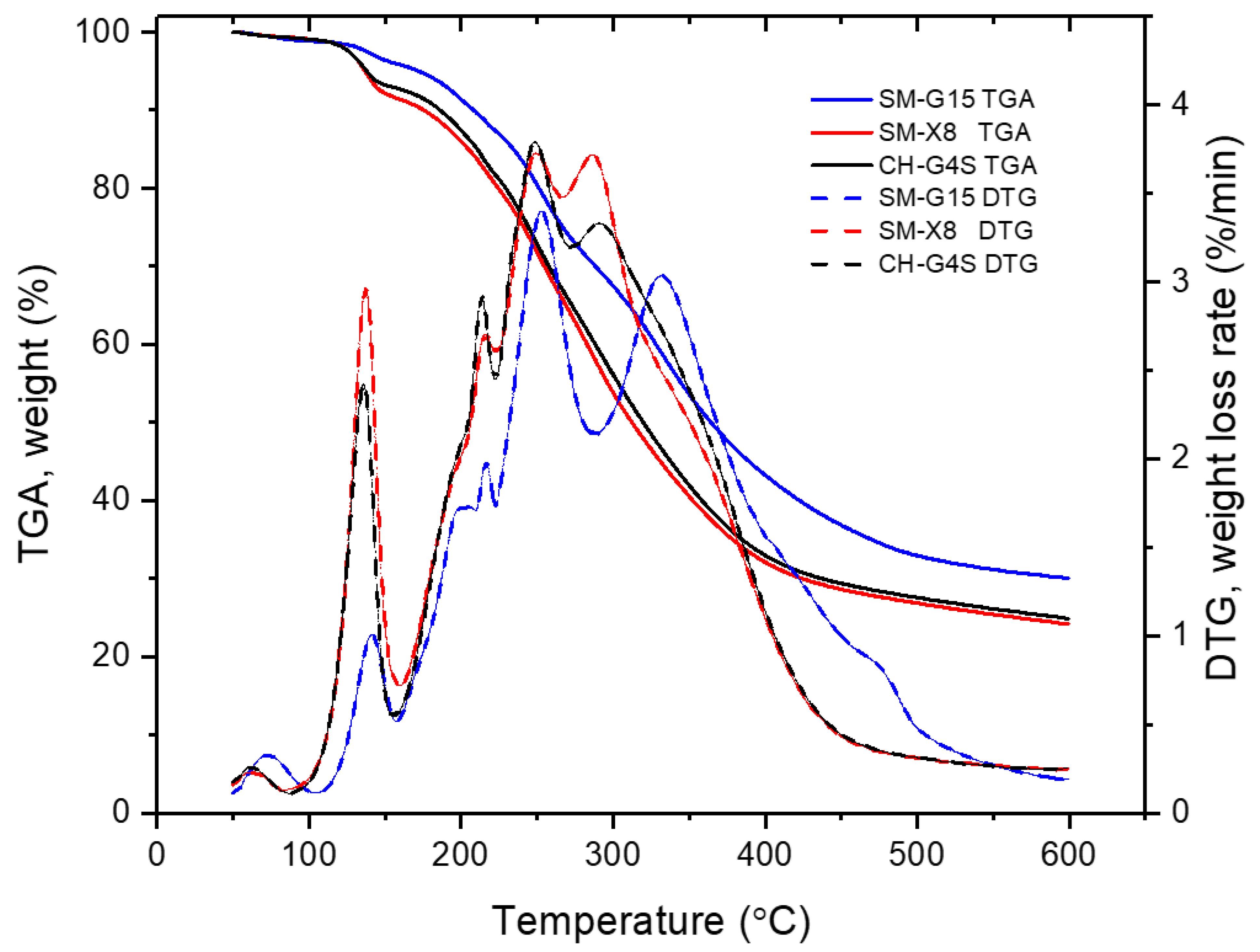
3.3.7. TGA
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Poli, A.; Finore, I.; Romano, I.; Gioiello, A.; Lama, L.; Nicolaus, B. Microbial diversity in extreme marine habitats and their biomolecules. Microorganisms 2017, 5, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.-Q.; Jiang, X.-R. Next generation industrial biotechnology based on extremophilic bacteria. Curr. Opin. Biotechnol. 2018, 50, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Kushner, D.J. Life in high salt and solute concentrations: Halophilic bacteria. In Microbial Life in Extreme Environments; Kushner, D.J., Ed.; Academic Press: London, UK, 1978; pp. 317–368. [Google Scholar]
- Poli, A.; Anzelmo, G.; Nicolaus, B. Bacterial exopolysaccharides from extreme marine habitats: Production, characterization and biological activities. Mar. Drugs 2010, 8, 1779–1802. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Sarkar, S.; Govil, T.; González-Faune, P.; Cabrera-Barjas, G.; Bandopadhyay, R.; Salem, D.R.; Sani, R.K. Extremophilic exopolysaccharides: Biotechnologies and wastewater remediation. Front. Microbiol. 2021, 12, 2349. [Google Scholar] [CrossRef] [PubMed]
- Daba, G.M.; Elnahas, M.O.; Elkhateeb, W.A. Contributions of exopolysaccharides from lactic acid bacteria as biotechnological tools in food, pharmaceutical, and medical applications. Int. J. Biol. Macromol. 2021, 173, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Angelin, J.; Kavitha, M. Exopolysaccharides from probiotic bacteria and their health potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Potnis, A.A.; Raghavan, P.S.; Rajaram, H. Overview on cyanobacterial exopolysaccharides and biofilms: Role in bioremediation. Rev. Environ. Sci. Bio/Technol. 2021, 20, 781–794. [Google Scholar] [CrossRef]
- Govarthanan, M.; Jeon, C.-H.; Jeon, Y.-H.; Kwon, J.-H.; Bae, H.; Kim, W. Non-toxic nano approach for wastewater treatment using Chlorella vulgaris exopolysaccharides immobilized in iron-magnetic nanoparticles. Int. J. Biol. Macromol. 2020, 162, 1241–1249. [Google Scholar] [CrossRef]
- Quillaguamán, J.; Delgado, O.; Mattiasson, B.; Hatti-Kaul, R. Chromohalobacter sarecensis sp. nov., a psychrotolerant moderate halophile isolated from the saline Andean region of Bolivia. Int. J. Syst. Evol. Microbiol. 2004, 54, 1921–1926. [Google Scholar] [CrossRef]
- Guzmán, H.; Van-Thuoc, D.; Martín, J.; Hatti-Kaul, R.; Quillaguamán, J. A process for the production of ectoine and poly(3-hydroxybutyrate) by Halomonas boliviensis. Appl. Microbiol. Biotechnol. 2009, 84, 1069–1077. [Google Scholar] [CrossRef]
- Guzmán, D.; Quillaguamán, J.; Muñoz, M.; Hatti-Kaul, R. Halomonas andesensis sp. nov., a moderate halophile isolated from the saline lake Laguna Colorada in Bolivia. Int. J. Syst. Evol. Microbiol. 2010, 60, 749–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambi, D.; Romero-Soto, L.; Villca, R.; Orozco-Gutiérrez, F.; Vega-Baudrit, J.; Quillaguamán, J.; Hatti-Kaul, R.; Martín, C.; Carrasco, C. Exopolysaccharides production by cultivating a bacterial isolate from the hypersaline environment of Salar de Uyuni (Bolivia) in pretreatment liquids of steam-exploded quinoa stalks and enzymatic hydrolysates of curupaú sawdust. Fermentation 2021, 7, 33. [Google Scholar] [CrossRef]
- Alandia, G.; Rodriguez, J.P.; Jacobsen, S.-E.; Bazile, D.; Condori, B. Global expansion of quinoa and challenges for the Andean region. Glob. Food Secur. 2020, 26, 100429. [Google Scholar] [CrossRef]
- Gil-Ramirez, A.; Salas-Veizaga, D.M.; Grey, C.; Karlsson, E.N.; Rodriguez-Meizoso, I.; Linares-Pastén, J.A. Integrated process for sequential extraction of saponins, xylan and cellulose from quinoa stalks (Chenopodium quinoa Willd). Ind. Crops Prod. 2018, 121, 54–65. [Google Scholar] [CrossRef]
- Carrasco, C.; Cuno, D.; Carlqvist, K.; Galbe, M.; Lidén, G. SO2-catalysed steam pretreatment of quinoa stalks. J. Chem. Technol. Biotechnol. 2015, 90, 64–71. [Google Scholar] [CrossRef]
- Carrasco, C.; Jönsson, L.J.; Martín, C. Hydrothermal pretreatment of water-extracted and aqueous ethanol-extracted quinoa stalks for enzymatic saccharification of cellulose. Energies 2021, 14, 4102. [Google Scholar] [CrossRef]
- Fendrihan, S.; Dornmayr-Pfaffenhuemer, M.; Gerbl, F.W.; Holzinger, A.; Grösbacher, M.; Briza, P.; Erler, A.; Gruber, C.; Plätzer, K.; Stan-Lotter, H. Spherical particles of halophilic archaea correlate with exposure to low water activity—Implications for microbial survival in fluid inclusions of ancient halite. Geobiology 2012, 10, 424–433. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC bioinformatics resource center: Expanding data and analysis capabilities. Nucleic Acids Res. 2019, 48, D606–D612. [Google Scholar] [CrossRef] [Green Version]
- Ondov, B.D.; Treangen, T.J.; Melsted, P.; Mallonee, A.B.; Bergman, N.H.; Koren, S.; Phillippy, A.M. Mash: Fast genome and metagenome distance estimation using MinHash. Genome Biol. 2016, 17, 132. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-R, L.M.; Gunturu, S.; Harvey, W.T.; Rosselló-Mora, R.; Tiedje, J.M.; Cole, J.R.; Konstantinidis, K.T. The microbial genomes atlas (MiGA) Webserver: Taxonomic and gene diversity analysis of archaea and bacteria at the whole genome level. Nucleic Acids Res. 2018, 46, W282–W288. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kanamarlapudi, S.L.R.K.; Muddada, S. Characterization of exopolysaccharide produced by Streptococcus thermophilus CC30. Biomed. Res. Int. 2017, 2017, 4201809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sardari, R.R.R.; Kulcinskaja, E.; Ron, E.Y.C.; Björnsdóttir, S.; Friðjónsson, Ó.H.; Hreggviðsson, G.Ó.; Karlsson, E.N. Evaluation of the production of exopolysaccharides by two strains of the thermophilic bacterium Rhodothermus marinus. Carbohydr. Polym. 2017, 156, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jönsson, L.J.; Martín, C. Pretreatment of lignocellulose: Formation of inhibitory by-products and strategies for minimizing their effects. Bioresour. Technol. 2016, 199, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Seo, W.T.; Kahng, G.G.; Nam, S.H.; Choi, S.D.; Suh, H.H.; Kim, S.W.; Park, Y.H. Isolation and characterization of a novel exopolysaccharide producing Paenibacillus sp. WN9 KCTC 8951P. J. Microbiol. Biotechnol. 1999, 9, 820–825. [Google Scholar]
- Han, Y.; Liu, E.; Liu, L.; Zhang, B.; Wang, Y.; Gui, M.; Wu, R.; Li, P. Rheological, emulsifying and thermostability properties of two exopolysaccharides produced by Bacillus amyloliquefaciens LPL061. Carbohydr. Polym. 2015, 115, 230–237. [Google Scholar] [CrossRef]
- Chikkanna, A.; Ghosh, D.; Kishore, A. Expression and characterization of a potential exopolysaccharide from a newly isolated halophilic thermotolerant bacteria Halomonas nitroreducens strain WB1. PeerJ 2018, 6, e4684. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Wang, Y.; Yan, F.; Song, R.; Li, Z.; Li, Y.; Song, B. Physical and chemical properties, percutaneous absorption-promoting effects of exopolysaccharide produced by Bacillus atrophaeus WYZ strain. Carbohydr. Polym. 2018, 192, 52–60. [Google Scholar] [CrossRef]
- Čopíková, J.; Barros, A.S.; Šmídová, I.; Černá, M.; Teixeira, D.H.; Delgadillo, I.; Synytsya, A.; Coimbra, M.A. Influence of hydration of food additive polysaccharides on FT-IR spectra distinction. Carbohydr. Polym. 2006, 63, 355–359. [Google Scholar] [CrossRef]
- Naumann, D. FT-Infrared and FT-Raman Spectroscopy in Biomedical Research. In Infrared and Raman Spectroscopy of Biological Materials; Practical Spectroscopy Series; Gremlich, H.-U., Yan, B., Eds.; Marcel Dekker Inc.: New York, NY, USA, 2001; Volume 24, pp. 323–377. [Google Scholar]
- Hounsell, E.F. 1H NMR in the structural and conformational analysis of oligosaccharides and glycoconjugates. Prog. Nucl. Magn. Reson. Spectrosc. 1995, 27, 445–474. [Google Scholar] [CrossRef]
- Beier, R.C.; Mundy, B.P.; Strobel, G.A. Assignment of anomeric configuration and identification of carbohydrate residues by 13C NMR. 1. galacto- and glucopyranosides and furanosides. Can. J. Chem. 1980, 58, 2800–2804. [Google Scholar] [CrossRef]
- Cantu-Jungles, T.M.; Iacomini, M.; Cipriani, T.R.; Cordeiro, L.M.C. Extraction and characterization of pectins from primary cell walls of edible açaí (Euterpe oleraceae) berries, fruits of a monocotyledon palm. Carbohydr. Polym. 2017, 158, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Capek, P.; Matulová, M.; Navarini, L.; Suggi-Liverani, F. Structural features of an arabinogalactan-protein isolated from instant coffee powder of coffea arabica beans. Carbohydr. Polym. 2010, 80, 180–185. [Google Scholar] [CrossRef]
- Park, S.; Baker, J.O.; Himmel, M.E.; Parilla, P.A.; Johnson, D.K. Cellulose crystallinity index: Measurement techniques and their impact on interpreting cellulase performance. Biotechnol. Biofuels 2010, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Sardjono, S.A.; Suryanto, H.; Aminnudin; Muhajir, M. Crystallinity and morphology of the bacterial nanocellulose membrane extracted from pineapple peel waste using high-pressure homogenizer. AIP Conf. Proc. 2019, 2120, 080015. [Google Scholar] [CrossRef]
- Wingender, J.; Neu, T.R.; Flemming, H.-C. Microbial Extracellular Polymeric Substances—Characterization, Structure and Function; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Spanò, A.; Gugliandolo, C.; Lentini, V.; Maugeri, T.; Anzelmo, G.; Poli, A.; Nicolaus, B. A novel EPS-producing strain of Bacillus licheniformis isolated from a shallow vent off Panarea Island (Italy). Curr. Microbiol. 2013, 67, 21–29. [Google Scholar] [CrossRef]
- Castellane, T.C.L.; Campanharo, J.C.; Colnago, L.A.; Coutinho, I.D.; Lopes, É.M.; Lemos, M.V.F.; de Macedo Lemos, E.G. Characterization of new exopolysaccharide production by Rhizobium tropici during growth on hydrocarbon substrate. Int. J. Biol. Macromol. 2017, 96, 361–369. [Google Scholar] [CrossRef]
- Rani, R.P.; Anandharaj, M.; Sabhapathy, P.; Ravindran, A.D. Physiochemical and biological characterization of novel exopolysaccharide produced by Bacillus tequilensis FR9 isolated from chicken. Int. J. Biol. Macromol. 2017, 96, 1–10. [Google Scholar] [CrossRef]
- Hu, X.; Li, D.; Qiao, Y.; Wang, X.; Zhang, Q.; Zhao, W.; Huang, L. Purification, characterization and anticancer activities of exopolysaccharide produced by Rhodococcus erythropolis HX-2. Int. J. Biol. Macromol. 2020, 145, 646–654. [Google Scholar] [CrossRef]
- Zamora, F.; González, M.C.; Dueñas, M.T.; Irastorza, A.; Velasco, S.; Ibarburu, I. Thermodegradation and thermal transitions of an exopolysaccharide produced by Pediococcus damnosus 2.6. J. Macromol. Sci. Part B 2002, 41, 473–486. [Google Scholar] [CrossRef]
- Werner, K.; Pommer, L.; Broström, M. Thermal decomposition of hemicelluloses. J. Anal. Appl. Pyrolysis 2014, 110, 130–137. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | AAI (%) | ANI (%) | Fraction of Genome Shared (%) |
|---|---|---|---|
| B. atrophaeus NZ CP021500 | 95 | 97.01 | 91.85 |
| B. atrophaeus NZ CP024051 | 95 | 96.96 | 90.96 |
| B. atrophaeus 1942 NC 014639 | 95 | 96.91 | 90.89 |
| Cellulosic Hydrolysate | Hemicellulosic Hydrolysate | |
|---|---|---|
| Glucose | 48.0 (1.3) | 1.5 (0.1) |
| Xylose | 4.9 (0.5) | 16.3 (0.7) |
| Mannose | 0.9 (<0.1) | 2.4 (0.1) |
| Galactose | 0.4 (<0.1) | 1.5 (<0.1) |
| Arabinose | 0.3 (<0.1) | 1.1 (<0.1) |
| Furfural | N.D. 1 | 4.7 (0.1) |
| HMF | N.D. 1 | 0.5 (<0.1) |
| Acetic acid | 1.0 (<0.1) | 5.6 (0.1) |
| Formic acid | N.D. 1 | 1.4 (<0.1) |
| Levulinic acid | N.D. 1 | 1.1 (0.1) |
| Medium | Carbon Source/Initial Concentration, g/L | Nomenclature | YInit 1, g/g | YCons. 2, g/g |
|---|---|---|---|---|
| Glucose-based synthetic medium | Glucose/45, xylose/5 | SM-G45 | 0.018 3 | 0.104 3 |
| Glucose-based synthetic medium | Glucose/30 | SM-G30 | 0.033 3 | 0.097 3 |
| Glucose-based synthetic medium | Glucose/15 | SM-G15 | 0.064 | 0.075 |
| Xylose-based synthetic medium | Xylose/16 | SM-X16 | 0.022 | 0.135 |
| Xylose-based synthetic medium | Xylose/8 | SM-X8 | 0.047 | 0.126 |
| Cellulosic hydrolysate | Glucose/45, xylose/5 | CH-G45 | 0.018 | 0.043 |
| Cellulosic hydrolysate | Glucose/30 | CH-G30 | 0.031 3 | 0.111 3 |
| Hemicellulosic hydrolysate | Xylose/16 | HcH-X16 | - | - |
| Hemicellulosic hydrolysate | Xylose/8 | HcH-X8 | - | - |
| EPS Preparation | Glucose | Mannose | Galactose | Arabinose | Xylose |
|---|---|---|---|---|---|
| SM-G15 | 49.6 (1.0) | 35.5 (0.9) | 8.2 (0.1) | 4.6 (<0.1) | 2.0 (0.1) |
| SM-X8 | 42.9 (4.5) | 36.2 (2.7) | 10.7 (1.2) | 6.5 (0.5) | 3.6 (0.3) |
| CH-G45 | 41.4 (<0.1) | 40.2 (0.4) | 9.9 (<0.1) | 3.8 (0.2) | 4.7 (0.2) |
| Peak 1 | Peak 2 | Peak 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Mw, g/mol | Mn, g/mol | PDI 1 | Mw, g/mol | Mn, g/mol | PDI 1 | Mw, g/mol | Mn, g/mol | PDI 1 | |
| SM-G15 | 9.1 × 105 | 8.9 × 105 | 1.0 | 4.7 × 104 | 4.4 × 104 | 1.1 | |||
| 9.2 × 105 | 8.2 × 105 | 1.1 | 6.1 × 104 | 4.7 × 104 | 1.3 | ||||
| SM-X8 | 8.7 × 105 | 7.9 × 104 | 1.1 | 6.1 × 104 | 4.9 × 104 | 1.2 | 2.2 × 104 | 2.2 × 104 | 1.0 |
| CH-G15 | 4.7×105 | 4.1×105 | 1.2 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chambi, D.; Lundqvist, J.; Nygren, E.; Romero-Soto, L.; Marin, K.; Gorzsás, A.; Hedenström, M.; Carlborg, M.; Broström, M.; Sundman, O.; et al. Production of Exopolysaccharides by Cultivation of Halotolerant Bacillus atrophaeus BU4 in Glucose- and Xylose-Based Synthetic Media and in Hydrolysates of Quinoa Stalks. Fermentation 2022, 8, 79. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8020079
Chambi D, Lundqvist J, Nygren E, Romero-Soto L, Marin K, Gorzsás A, Hedenström M, Carlborg M, Broström M, Sundman O, et al. Production of Exopolysaccharides by Cultivation of Halotolerant Bacillus atrophaeus BU4 in Glucose- and Xylose-Based Synthetic Media and in Hydrolysates of Quinoa Stalks. Fermentation. 2022; 8(2):79. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8020079
Chicago/Turabian StyleChambi, Diego, Jenny Lundqvist, Erik Nygren, Luis Romero-Soto, Katherine Marin, András Gorzsás, Mattias Hedenström, Markus Carlborg, Markus Broström, Ola Sundman, and et al. 2022. "Production of Exopolysaccharides by Cultivation of Halotolerant Bacillus atrophaeus BU4 in Glucose- and Xylose-Based Synthetic Media and in Hydrolysates of Quinoa Stalks" Fermentation 8, no. 2: 79. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8020079