
Immune Response and Fatty Acid Profile of Eggs from Laying Hens Fed Fermented Feed Rich in Polyunsaturated Fatty Acids

 ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of Fermented Feed
2.3. Analysis of Fatty Acid Profile in Egg Yolk and Feed Mixtures
2.4. Relative Expression of Selected Genes in Quantitative Real-Time PCR (qRT-PCR)
2.4.1. Homogenization of Caecal Samples and Isolation of Total RNA of MUC-2, IgA, and IGF-2 (Growth Factor) Gene
2.4.2. Quantitative Real/Time PCR
2.5. Phagocyte Activity Testing
2.6. Lymphocyte Phenotyping
2.7. Microbiological Screening
2.8. Statistical Analysis
3. Results
3.1. Relative Expression of Selected Parameters
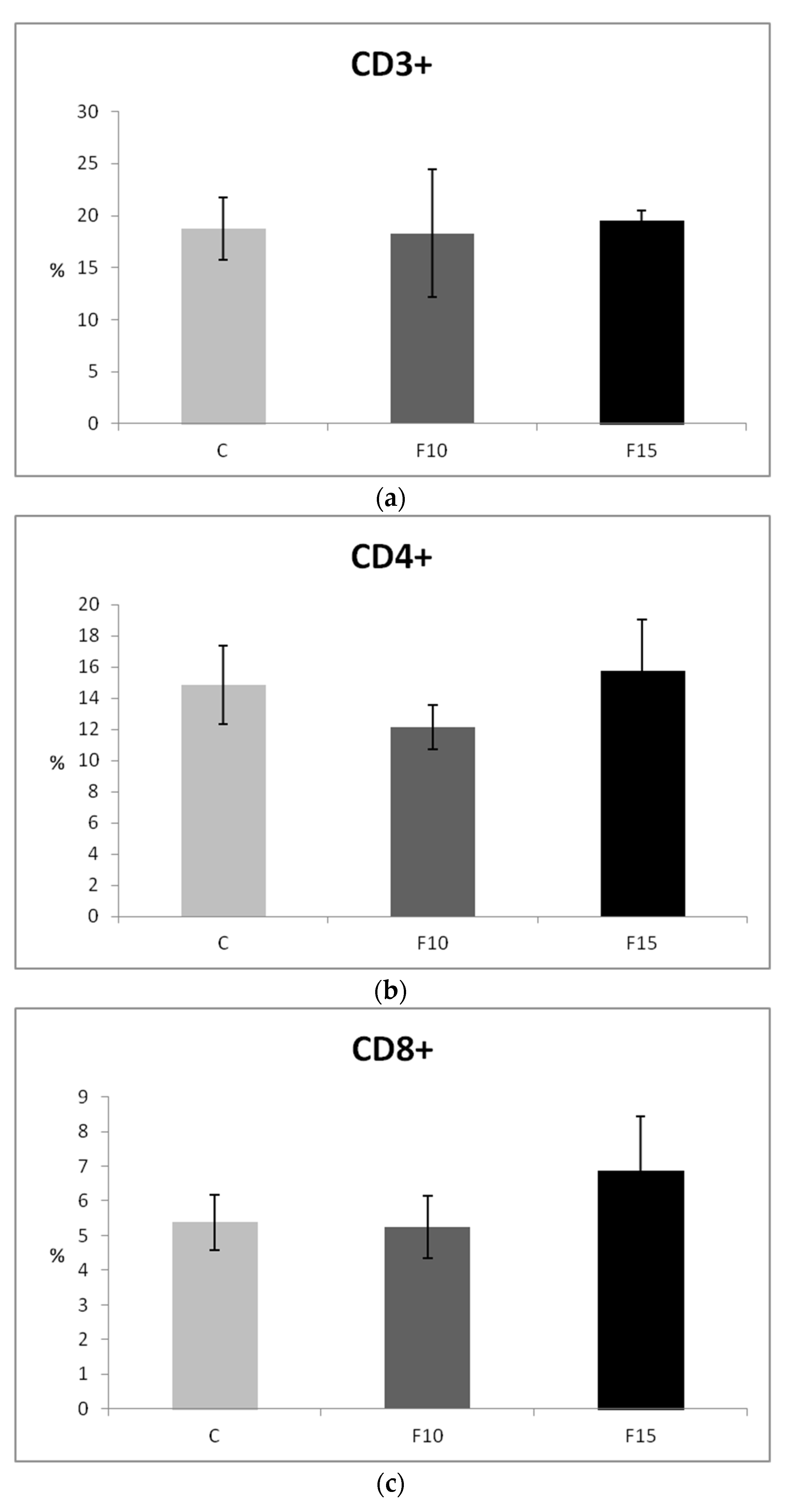
3.2. Cellular Immune Response
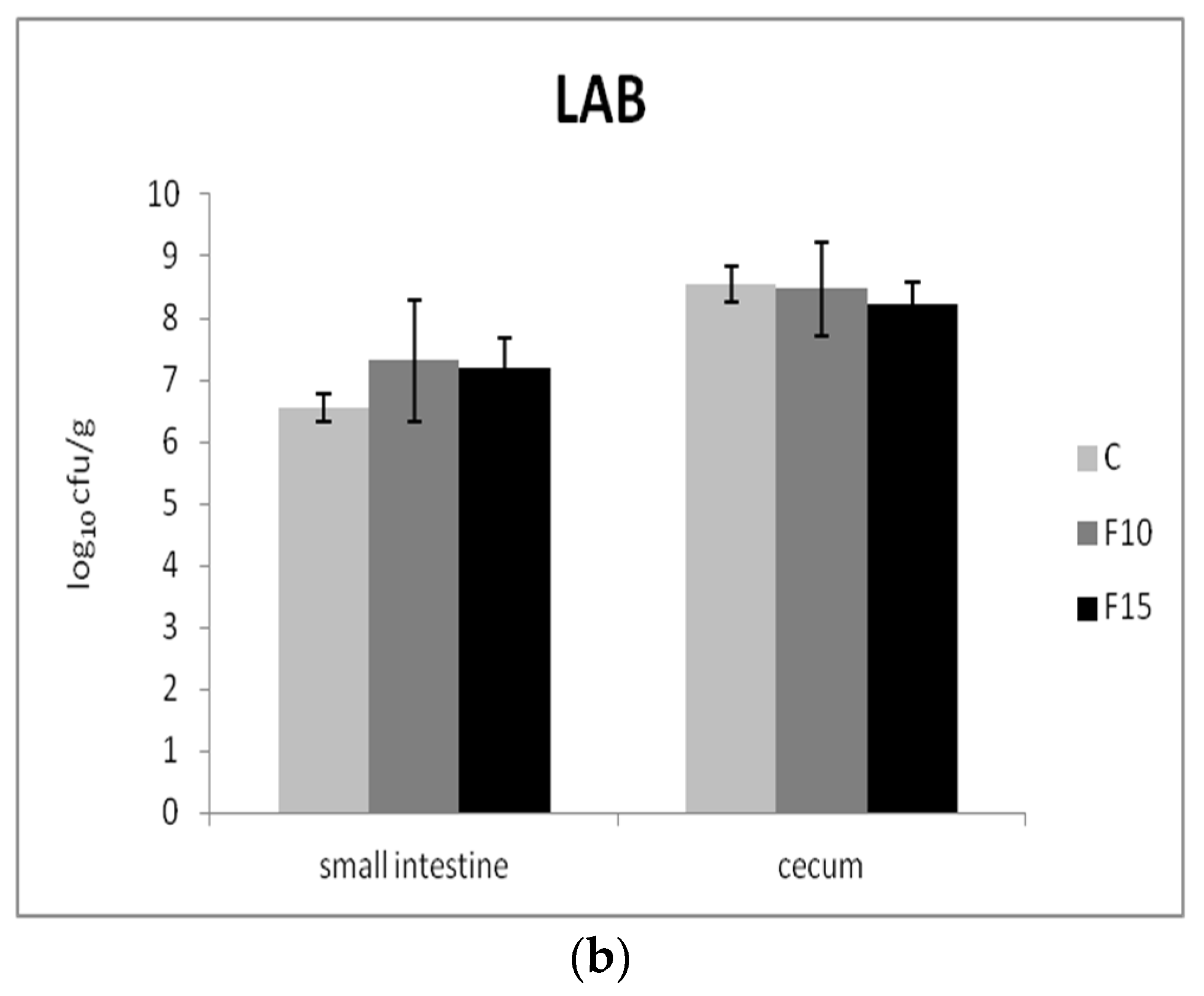
3.3. Intestinal Microbiota
3.4. Fatty Acid Profile in Yolk of Egg
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Ander, B.P.; Dupasquier, C.M.; Prociuk, M.A.; Pierce, G.N. Polyunsaturated fatty acids and their effects on cardiovascular disease. Exp. Clin. Cardiol. 2003, 8, 164–172. [Google Scholar] [PubMed]
- Čertík, M.; Klempová, T.; Guothová, L.; Mihálik, D.; Kraic, J. Biotechnology for the functional improvement of cereal-based materials enriched with PUFA and pigments. Eur. J. Lipid. Sci. Technol. 2013, 115, 1247–1256. [Google Scholar] [CrossRef]
- Sakuradani, E.; Ando, A.; Ogawa, J.; Shimizu, S. Improved production of various polyunsaturated fatty acids through filamentous fungus Mortierella alpina breeding. Appl. Microbiol. Biotechnol. 2009, 84, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Semjon, B.; Bartkovský, M.; Marcinčáková, D.; Klempová, T.; Bujňák, L.; Hudák, M.; Jaďuttová, I.; Čertík, M.; Marcinčák, S. Effect of Solid-State Fermented Wheat Bran Supplemented with Agrimony Extract on Growth Performance, Fatty Acid Profile, and Meat Quality of Broiler Chickens. Animals 2020, 10, 942. [Google Scholar] [CrossRef] [PubMed]
- Directive. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes. Off. J. Eur. Union 2010, 276, 33–79. [Google Scholar]
- Klempová, T.; Slaný, O.; Šišmiš, M.; Marcinčík, S.; Čertík, M. Dual production of polyunsaturated fatty acids and beta-carotene with Mucor wosnessenskii by the process of solid-state fermentation using agro-industrial waste. J. Biotechnol. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lammers, A.; Wieland, W.H.; Kruijt, L.; Jansma, A.; Straetemans, T.; Schots, A.; den Hartog, G.; Parmentier, H.K. Successive immunoglobulin and cytokine expression in the small intestine of juvenile chicken. Dev. Comp. Immunol. 2010, 34, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, A.; Tako, E.; Ferket, P.R.; Uni, Z. Mucin gene expression and mucin content in the chicken intestinal goblet cells are affected by in ovo feeding of carbohydrates. Poult. Sci. 2006, 85, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Mudronová, D.; Karaffová, V.; Košcová, J.; Bartkovský, M.; Marcincáková, D.; Popelka, P.; Klempová, T.; Certík, M.; Macanga, J.; Marcincák, S. Effect of fungal gamma-linolenic acid and beta-carotene containing prefermented feed on immunity and gut of broiler chicken. Poult. Sci. 2018, 97, 4211–4218. [Google Scholar] [CrossRef]
- De Boever, S.; Vangestel, C.; De Backer, P.; Croubels, S.; Sys, S.U. Identification and validation of housekeeping genes as internal control for gene expression in an intravenous LPS inflammation model in chickens. Vet. Immunol. Immunopathol. 2008, 122, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Mudroňová, D.; Karaffová, V.; Semjon, B.; Naď, P.; Koščová, J.; Bartkovský, M.; Makiš, A.; Bujňák, L.; Nagy, J.; Mojžišová, J.; et al. Effects of Dietary Supplementation of Humic Substances on Production Parameters, Immune Status and Gut Microbiota of Laying Hens. Agriculture 2021, 11, 744. [Google Scholar] [CrossRef]
- Bertram, E.M. Characterisation of duck thromhocytes. Res. Vet. Sci. 1998, 64, 267–270. [Google Scholar] [CrossRef]
- Luthala, M. Chicken CD4, CD8αβ, and CD8αα T cell co-receptor molecules. Poult. Sci. 1998, 77, 1858–1873. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Liu, J.; Wang, H.; Luo, J.; Chen, T.; Xi, Q.; Zhang, Y.; Sun, J. Effects of fermented feeds and ginseng polysaccharides on the intestinal morphology and microbiota composition of Xuefeng black-bone chicken. PLoS ONE 2020, 15, e0237357. [Google Scholar] [CrossRef]
- Mantis, N.J.; Rol, N.; Corthésy, B. Secretory IgA’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef]
- Ziegler, A.N.; Feng, Q.; Chidambaram, S.; Testai, J.M.; Kumari, E.; Rothbard, D.E.; Constancia, M.; Sandovici, I.; Cominski, T.; Pang, K.; et al. Insulin-like growth factor ii: An essential adult stem cell niche constituent in brain and intestine. Stem Cell Rep. 2019, 12, 816–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Feng, J.; Wang, Y.; Lv, J.; Li, J.; Guo, L.; Min, Y. Fermented Corn–Soybean Meal Mixed Feed Modulates Intestinal Morphology, Barrier Functions and Cecal Microbiota in Laying Hens. Animals 2021, 11, 3059. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, H.G.; Takeo Sato, F.; Curi, R.; Vinolo, M.A.R. Fatty acids as modulators of neutrophil recruitment, function and survival. Eur. J. Pharmacol. 2016, 785, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Martins de Lima, T.; Gorjão, R.; Hatanaka, E.; Cury-Boaventura, M.F.; Portioli Silva, E.P.; Procopio, J.; Curi, R. Mechanisms by which fatty acids regulate leucocyte function. Clin. Sci. 2007, 113, 65–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calder, P.C. Mechanisms of action of (n-3) fatty acids. J. Nutr. 2012, 142, 592S–599S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelan, J.; Gowdy, K.M.; Shaikh, S.R. N-3 polyunsaturated fatty acids modulate B cell activity in pre-clinical models: Implications for the immune response to infections. Eur. J. Pharmacol. 2016, 785, 10–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geier, M.S.; Torok, V.A.; Allison, G.E.; Ophel-Keller, K.; Gibson, R.A.; Munday, C.; Hughes, R.J. Dietary omega-3 polyunsaturated fatty acid does not influence the intestinal microbial communities of broiler chickens. Poult. Sci. 2009, 88, 2399–2405. [Google Scholar] [CrossRef]
- Kankaanpää, P.E.; Salminen, S.J.; Isolauri, E.; Lee, Y.K. The influence of polyunsaturated fatty acids on probiotic growth and adhesion. FEMS Microbiol. Lett. 2001, 194, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; DeCoffe, D.; Brown, K.; Rajendiran, E.; Estaki, M.; Dai, C.; Yip, A.; Gibson, D.L. Fish oil attenuates omega-6 polyunsaturated fatty acid-induced dysbiosis and infectious colitis but impairs LPS dephosphorylation activity causing sepsis. PLoS ONE 2013, 8, e55468. [Google Scholar] [CrossRef] [PubMed]
- Vlaicu, P.A.; Panaite, T.D.; Turcu, R.P. Enriching laying hens eggs by feeding diets with different fatty acid composition and antioxidants. Sci. Rep. 2021, 11, 20707. [Google Scholar] [CrossRef]
- Alagawany, M.; Elnesr, S.S.; Farag, M.R.; Abd El-Hack, M.E.; Khafaga, A.F.; Taha, A.E.; Tiwari, R.; Yatoo, M.I.; Bhatt, P.; Khurana, S.K.; et al. Omega-3 and Omega-6 fatty acids in poultry nutrition: Effect on production performance and health. Animals 2019, 9, 573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopacz, M.; Drazbo, A.A.; Smiecinska, K.; Ognik, K. Performance and egg quality of laying hens fed diets containing raw, hydrobarothermally-treated and fermented rapeseed cake. Animals 2021, 11, 3083. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, R.; Huang, Y.S. Gamma linolenic acid: An antiinflammatory omega-6 fatty acid. Curr. Pharm. Biotechnol. 2006, 7, 531–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciubota-Rosie, C.; Macoveanu, M.; Maria Fernandez, C.; Jesus Ramos, M.; Perez, A.; Moreno, A. Sinapis alba seed as a prospective biodiesel source. Biomass Bioenergy 2013, 51, 83–90. [Google Scholar] [CrossRef]
- Kikukawa, H.; Sakuradani, E.; Nishibaba, Y.; Okuda, T.; Ando, A.; Shima, J.; Shimizu, S.; Ogawa, J. Production of cis-11-eicosenoic acid by Mortierella fungi. J. Appl. Microbiol. 2015, 118, 641–647. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound (Unit) | FP | C | F10 | F15 |
|---|---|---|---|---|
| ME (MJ/kg) | 6.49 | 11.12 | 10.62 | 10.43 |
| Dry matter (g/kg) | 1000 | 1000 | 1000 | 1000 |
| Protein (g/kg) | 193.4 | 157.5 | 165.6 | 167.1 |
| Fat (g/kg) | 50.1 | 37.2 | 38.7 | 41.7 |
| fibre (g/kg) | 169.1 | 58.9 | 78.4 | 79.2 |
| Ash (g/kg) | 75.8 | 154.3 | 152.2 | 147.4 |
| Starch (g/kg) | 75.5 | 411.8 | 371.8 | 353.0 |
| Ca (g/kg) | 1.63 | 20.0 | 19.75 | 15.45 |
| P (g/kg) | 5.97 | 4.86 | 4.90 | 5.03 |
| Mg (g/kg) | 4.34 | 3.09 | 3.41 | 3.49 |
| Na (g/kg) | 0.22 | 2.21 | 1.99 | 2.01 |
| K (g/kg) | 13.02 | 7.51 | 7.92 | 8.12 |
| Cu (mg/kg) | 36.03 | 44.19 | 41.79 | 43.52 |
| Zn (mg/kg) | 106.78 | 101.63 | 104.89 | 103.57 |
| Mn (mg/kg) | 195.44 | 173.44 | 176.08 | 178.67 |
| Lysine (%) | nt | 0.88 | 0.84 | 0.82 |
| Digestible Lys (%) | nt | 0.73 | 0.69 | 0.66 |
| Methionine (%) | nt | 0.44 | 0.42 | 0.40 |
| Digestible Met (%) | nt | 0.37 | 0.34 | 0.33 |
| Fatty Acid | Fermented Product (FP) | C | F10 | F15 |
|---|---|---|---|---|
| C16:0, PA | 15.70 ± 1.02 | 12.81 ± 0.39 | 13.32 ± 0.89 | 13.62 ± 0.20 |
| C16:1, n-7, POA | 0.59 ± 0.03 | 0.257 ± 0.000 | 0.357 ± 0.034 | 0.403 ± 0.042 |
| C18:0, SA | 1.88 ± 0.09 | 3.12 ± 0.09 | 2.75 ± 0.13 | 2.89 ± 0.17 |
| C18:1, n-9, OA | 17.23 ± 0.62 | 42.59 ± 1.91 | 37.86 ± 0.64 | 36.36 ± 1.65 |
| C18:1, n-7 | 1.21 ± 0.09 | 1.79 ± 0.19 | 1.80 ± 0.12 | 1.71 ± 0.13 |
| C18:2, n-6, LA | 45.51 ± 2.11 | 36.37 ± 2.57 | 36.45 ± 0.72 | 36.14 ± 1.81 |
| C18:3, n-6, GLA | 2.48 ± 0.73 | nd. | 0.73 ± 0.09 | 0.79 ± 0.09 |
| C18:3, n-3, ALA | 6.31 ± 0.72 | 1.69 ± 0.15 | 3.28 ± 0.37 | 3.38 ± 0.09 |
| C20:0 | 0.40 ± 0.04 | 0.48 ± 0.01 | 0.43 ± 0.02 | 0.46 ± 0.03 |
| C20:1, n-9 | 0.661 ± 0.015 | 0.37 ± 0.03 | 0.51 ± 0.01 | 0.45 ± 0.03 |
| C20:3, n-6, DGLA | 0.362 ± 0.095 | nd. | nd. | nd. |
| C20:4, n-6, ARA | 6.51 ± 0.34 | nd. | 1.64 ± 0.07 | 2.82 ± 0.05 |
| C20:5, n-3, EPA | 0.394 ± 0.195 | nd. | nd. | nd. |
| C24:0 | 0.77 ± 0.10 | 0.41 ± 0.02 | 0.49 ± 0.02 | 0.56 ± 0.01 |
| Σ SFA | 18.75 ± 0.31 | 16.82 ± 0.45 | 16.99 ± 0.50 | 17.54 ± 0.39 |
| Σ MUFA | 19.69 ± 0.18 | 45.12 ± 2.27 | 40.90 ± 0.64 | 39.35 ± 1.53 |
| Σ PUFA | 61.56 ± 1.18 | 38.06 ± 2.46 | 42.10 ± 0.61 | 43.11± 1.89 |
| Σ n-6 | 54.86 ± 0.82 | 36.37 ± 2.57 | 37.18 ± 0.65 | 36.91 ± 1.39 |
| Σ n-3 | 6.70 ± 0.46 | 1.69 ± 0.15 | 3.28 ± 0.37 | 3.38 ± 0.09 |
| Σ n-6/Σ n-3 | 8.18 ± 0.43 | 21.52 ± 4.02 | 11.33 ± 0.55 | 10.91 ± 1.28 |
| Primer | Sequence 5′–3′ | Annealing/Temperature Time | References |
|---|---|---|---|
| MUC-2 Fw | GCTGATTGTCACTCACGCCTT | 54 °C/1 min | [9] |
| MUC-2 Rev | ATCTGCCTGAATCACAGGTGC | ||
| IgA Fw | GTCACCGTCACCTGGACTACA | 59 °C/30 s | [8] |
| IgA Rev | ACCGATGGTCTCCTTCACATC | ||
| IGF-2 Fw | CTCTGCTGGAAACCTACTGT | 60 °C/1 min | [10] |
| IGF-2 Rev | GAGTACTTGGCATGAGATGG | ||
| GAPDH Fw | CCTGCATCTGCCCATTT | 59 °C/30 s | [11] |
| GAPDH Rev | GGCACGCCATCACTATC |
| Fatty Acid (%) | C | F10 | F15 |
|---|---|---|---|
| C16:0, PA | 25.98 ± 1.49 | 25.71 ± 0.15 | 26.19 ± 0.38 |
| C16:1, n-9, POA | 3.97 ± 0.97 | 3.82 ± 0.38 | 3.86 ± 0.28 |
| C18:0, SA | 7.19 ± 0.68 | 7.81 ± 0.44 | 7.19 ± 0.16 |
| C18:1, n-9, OA | 39.74 ± 1.91 | 39.49 ± 1.64 | 39.15 ± 0.96 |
| C18:1, n-7 VA | 2.32 ± 0.14 | 2.37 ± 0.28 | 2.31 ± 0.05 |
| C18:2, n-6, LA | 15.88 ± 1.79 | 14.53 ± 1.53 | 15.28 ± 0.49 |
| C18:3, n-6, GLA | 0.12 ± 0.10 b | 0.16 ± 0.04 b | 0.28 ± 0.17 a |
| C18:3, n-3, ALA | 0.22 ± 0.08 c | 0.31 ± 0.04 b | 0.40 ± 0.01 a |
| C20:1, GA | 0.28 ± 0.02 a | 0.19 ± 0.11 c | 0.23 ± 0.01 b |
| C20:3, n-6, DGLA | 0.13 ± 0.08 c | 0.20 ± 0.04 b | 0.24 ± 0.01 a |
| C20:4, n-6, ARA | 2.11 ± 0.15 | 2.34 ± 0.32 | 2.08 ± 0.16 |
| C20:5, n-3, EPA | Nd b | 0.05 ± 0.01 a | 0.06 ± 0.01 a |
| C22:6, n-3, DHA | 0.39 ± 0.01 b | 0.59 ± 0.09 a | 0.59 ± 0.08 a |
| Σ SFA | 33.61 ± 1.29 | 33.81 ± 0.32 | 33.69 ± 0.51 |
| Σ MUFA | 46.70 ± 1.46 | 46.21 ± 1.96 | 46.23 ± 0.76 |
| Σ PUFA | 19.69 ±1.20 | 19.98 ± 0.82 | 20.08 ± 0.36 |
| Σ n-6 | 19.08 ± 1.22 | 19.03 ± 1.08 | 19.03 ± 0.41 |
| Σ n-3 | 0.61 ± 0.08 b | 0.95 ± 0.05 a | 1.05 ± 0.04 a |
| Σ n-6/Σ n-3 | 32.07 ± 1.44 a | 19.97 ± 1.43 b | 18.12 ± 1.28 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karaffová, V.; Mudroňová, D.; Semjon, B.; Klempová, T.; Slaný, O.; Čertík, M.; Nagy, J.; Bartkovský, M.; Marcinčák, S. Immune Response and Fatty Acid Profile of Eggs from Laying Hens Fed Fermented Feed Rich in Polyunsaturated Fatty Acids. Fermentation 2022, 8, 98. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8030098
Karaffová V, Mudroňová D, Semjon B, Klempová T, Slaný O, Čertík M, Nagy J, Bartkovský M, Marcinčák S. Immune Response and Fatty Acid Profile of Eggs from Laying Hens Fed Fermented Feed Rich in Polyunsaturated Fatty Acids. Fermentation. 2022; 8(3):98. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8030098
Chicago/Turabian StyleKaraffová, Viera, Dagmar Mudroňová, Boris Semjon, Tatiana Klempová, Ondrej Slaný, Milan Čertík, Jozef Nagy, Martin Bartkovský, and Slavomír Marcinčák. 2022. "Immune Response and Fatty Acid Profile of Eggs from Laying Hens Fed Fermented Feed Rich in Polyunsaturated Fatty Acids" Fermentation 8, no. 3: 98. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8030098