
Enhancing Ethanol Tolerance via the Mutational Breeding of Pichia terricola H5 to Improve the Flavor Profiles of Wine


Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Growth Conditions
2.2. Screening the Non-Saccharomyces Yeasts for Ethanol Tolerance
2.2.1. High-Throughput 2,3,5-Triphenyltetrazolium Chloride (TTC) Medium Method for First Stage Selection
2.2.2. Detecting Non-Saccharomyces Yeast Activity under Ethanol Stress
2.2.3. The Influence of Ethanol Addition on Yeast Cell Growth
2.3. Mutation Procedure of Non-Saccharomyces Yeast by DES and UV
2.4. Identification of Yeast
2.5. The Practical Fermentation of Selected Strains
2.6. Determining the Physicochemical Indexes via HPLC
2.7. Determining the Aroma Compounds via HS-SPME/GC-MS-FID
2.8. Sensory Evaluation
2.9. Statistical Analysis
3. Results
3.1. Selecting a Non-Saccharomyces Yeast with High-Alcoholicity Endurance
3.2. Screening for Non-Saccharomyces Yeasts with High Ethanol Tolerance Using DES and UV
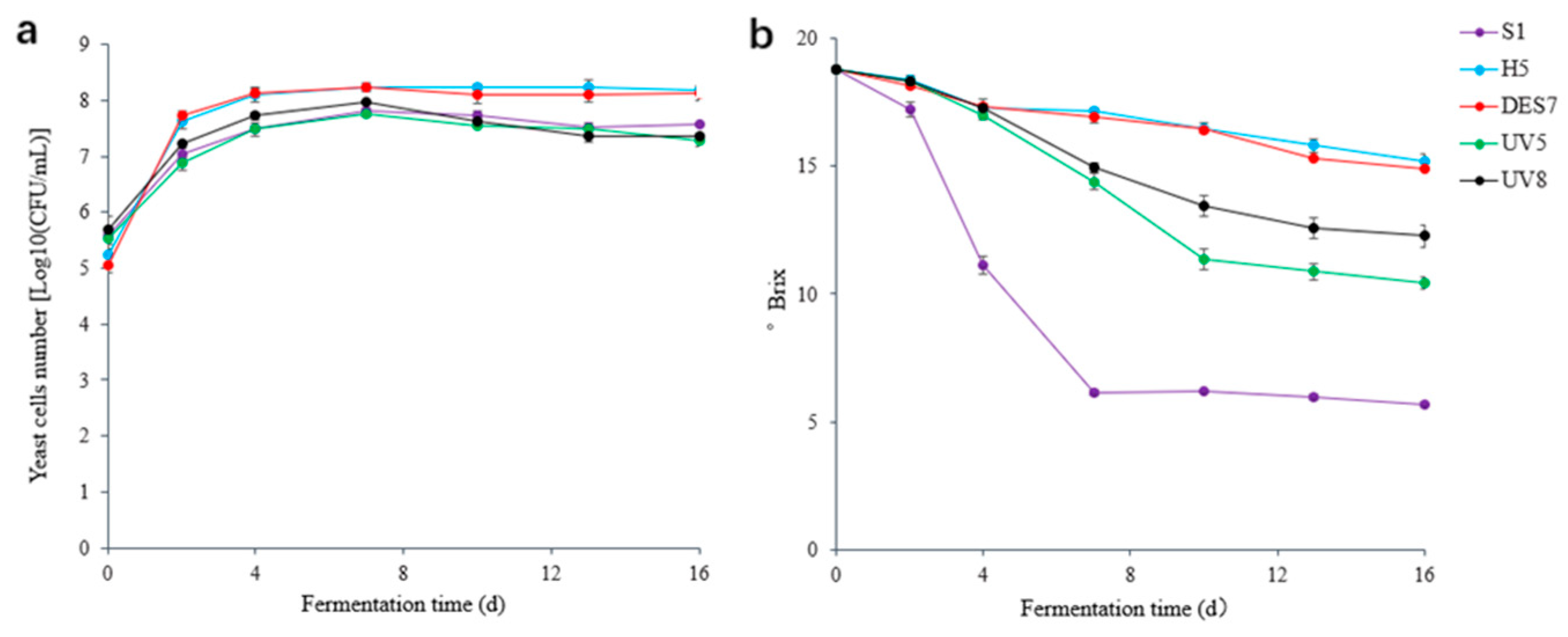
3.3. Evaluation of the Fermentation Ability of the Mutagenized Strains
3.3.1. The Performance of the Mutants during Fermentation
3.3.2. The Influence of Mutants on the Volatile Aroma Compounds
3.4. Identification of Strain UV5
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gobbi, M.; De Vero, L.; Solieri, L.; Comitini, F.; Oro, L.; Giudici, P.; Ciani, M. Fermentative Aptitude of Non-Saccharomyces Wine Yeast for Reduction in the Ethanol Content in Wine. Eur. Food Res. Technol. 2014, 239, 41–48. [Google Scholar] [CrossRef]
- Prior, K.J.; Bauer, F.F.; Divol, B. The Utilisation of Nitrogenous Compounds by Commercial Non-Saccharomyces Yeasts Associated with Wine. Food Microbiol. 2019, 79, 75–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled Mixed Culture Fermentation: A New Perspective on the Use of Non-Saccharomyces Yeasts in Winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not Your Ordinary Yeast: Non-Saccharomyces Yeasts in Wine Production Uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Sadoudi, M.; Tourdot-Marechal, R.; Rousseaux, S.; Steyer, D.; Gallardo-Chacon, J.J.; Ballester, J.; Vichi, S.; Guerin-Schneider, R.; Caixach, J.; Alexandre, H. Yeast-Yeast Interactions Revealed by Aromatic Profile Analysis of Sauvignon Blanc Wine Fermented by Single or Co-Culture of Non-Saccharomyces and Saccharomyces Yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef]
- Cordero-Bueso, G.; Esteve-Zarzoso, B.; Cabellos, J.M.; Gil-Diaz, M.; Arroyo, T. Biotechnological Potential of Non-Saccharomyces Yeasts Isolated During Spontaneous Fermentations of Malvar (Vitis Vinifera Cv. L.). Eur. Food Res. Technol. 2013, 236, 193–207. [Google Scholar] [CrossRef]
- Giorello, F.; Valera, M.J.; Martin, V.; Parada, A.; Salzman, V.; Camesasca, L.; Farina, L.; Boido, E.; Medina, K.; Dellacassa, E.; et al. Genomic and Transcriptomic Basis of Hanseniaspora vineae’s Impact on Flavor Diversity and Wine Quality. Appl. Environ. Microb. 2019, 85, e01959-18. [Google Scholar] [CrossRef] [Green Version]
- Whitener, M.E.B.; Carlin, S.; Jacobson, D.; Weighill, D.; Divol, B.; Conterno, L.; Du Toit, M.; Vrhovsek, U. Early Fermentation Volatile Metabolite Profile of Non-Saccharomyces Yeasts in Red and White Grape Must: A Targeted Approach. LWT Food Sci. Technol. 2015, 64, 412–422. [Google Scholar] [CrossRef]
- Anfang, N.; Brajkovich, M.; Goddard, M.R. Co-Fermentation with Pichia Kluyveri Increases Varietal Thiol Concentrations in Sauvignon Blanc. Aust. J. Grape Wine Res. 2009, 15, 1–8. [Google Scholar] [CrossRef]
- Liu, P.T.; Lu, L.; Duan, C.Q.; Yan, G.L. The Contribution of Indigenous Non-Saccharomyces Wine Yeast to Improved Aromatic Quality of Cabernet Sauvignon Wines by Spontaneous Fermentation. LWT Food Sci. Technol. 2016, 71, 356–363. [Google Scholar]
- Heard, G.M.; Fleet, G.H. The Effects of Temperature and Ph on the Growth of Yeast Species During the Fermentation of Grape Juice. J. Appl. Bacteriol. 1988, 65, 23–28. [Google Scholar] [CrossRef]
- Addo-Quaye, C.; Tuinstra, M.; Carraro, N.; Weil, C.; Dilkes, B.P. Whole-Genome Sequence Accuracy Is Improved by Replication in a Population of Mutagenized Sorghum. G3 Genes Genomes Genet. 2018, 8, 1079–1094. [Google Scholar]
- Zhang, G.Q.; Lin, Y.P.; Qi, X.N.; Wang, L.X.; He, P.; Wang, Q.H.; Ma, Y.H. Genome Shuffling of the Nonconventional Yeast Pichia anomala for Improved Sugar Alcohol Production. Microb. Cell Fact. 2015, 14, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, L.J.; Lee, P.H.; Small, E.M.; Hillyer, C.; Guo, Y.; Osley, M.A. Regulation of UV Damage Repair in Quiescent Yeast Cells. DNA Repair 2020, 90, 102861. [Google Scholar] [CrossRef] [PubMed]
- Ikehata, H.; Ono, T. The Mechanisms of UV Mutagenesis. J. Radiat. Res. 2011, 52, 115–125. [Google Scholar] [PubMed] [Green Version]
- Hoffmann, G.R. Genetic Effects of Dimethyl Sulfate, Diethyl Sulfate, and Related-Compounds. Mutat. Res. 1980, 75, 63–129. [Google Scholar] [CrossRef]
- Abrol, V.; Kushwaha, M.; Arora, D.; Mallubhotla, S.; Jaglan, S. Mutation, Chemoprofiling, Dereplication, and Isolation of Natural Products from Penicillium oxalicum. ACS Omega 2021, 6, 16266–16272. [Google Scholar] [CrossRef]
- Pang, Z.W.; Liang, J.J.; Qin, X.J.; Wang, J.R.; Feng, J.X.; Huang, R.B. Multiple Induced Mutagenesis for Improvement of Ethanol Production by Kluyveromyces marxianus. Biotechnol. Lett. 2010, 32, 1847–1851. [Google Scholar] [CrossRef]
- Yi, S.; Zhang, X.; Li, H.X.; Du, X.X.; Liang, S.W.; Zhao, X.H. Screening and Mutation of Saccharomyces cerevisiae UV-20 with a High Yield of Second Generation Bioethanol and High Tolerance of Temperature, Glucose and Ethanol. Indian J. Microbiol. 2018, 58, 440–447. [Google Scholar] [CrossRef]
- Kang, L.Z.; Han, F.; Lin, J.F.; Guo, L.Q.; Bai, W.F. Breeding of New High-Temperature-Tolerant Strains of Flammulina velutipes. Sci. Hortic. 2013, 151, 97–102. [Google Scholar]
- Hesham, A.E.L.; Wambui, V.; Jo, H.O.; Maina, J.M. Phylogenetic Analysis of Isolated Biofuel Yeasts Based on 5.8S-ITS rDNA and D1/D2 26S rDNA Sequences. J. Genet. Eng. Biotechnol. 2014, 12, 37–43. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T.; Robert, V. Methods for Isolation, Phenotypic Characterization and Maintenance of Yeasts. In The Yeasts, a Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 87–110. [Google Scholar]
- Sun, X.Y.; Zhao, Y.; Liu, L.L.; Jia, B.; Zhao, F.; Huang, W.D.; Zhan, J.C. Copper Tolerance and Biosorption of Saccharomyces cerevisiae During Alcoholic Fermentation. PLoS ONE 2015, 10, e0128611. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Chia, J.Y.; Liu, S.Q. Impact of Addition of Aromatic Amino Acids on Non-Volatile and Volatile Compounds in Lychee Wine Fermented with Saccharomyces cerevisiae Merit.Ferm. Int. J. Food Microbiol. 2014, 170, 12–20. [Google Scholar] [CrossRef]
- Volschenk, H.; Van Vuuren, H.J.J.; Viljoen-Bloom, M. Malic Acid in Wine: Origin, Function and Metabolism During Vinification. S. Afr. J. Enol. Vitic. 2006, 27, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Peinado, R.A.; Mauricio, J.C.; Moreno, J. Aromatic Series in Sherry Wines with Gluconic Acid Subjected to Different Biological Aging Conditions by Saccharomyces cerevisiae Var. Capensis. Food Chem. 2006, 94, 232–239. [Google Scholar] [CrossRef]
- Lelova, Z.; Ivanova-Petropulos, V.; Masar, M.; Lisjak, K.; Bodor, R. Optimization and Validation of a New Capillary Electrophoresis Method with Conductivity Detection for Determination of Small Anions in Red Wines. Food Anal. Methods 2018, 11, 1457–1466. [Google Scholar] [CrossRef]
- Redzepovic, S.; Orlic, S.; Majdak, A.; Kozina, B.; Volschenk, H.; Viljoen-Bloom, M. Differential Malic Acid Degradation by Selected Strains of Saccharomyces During Alcoholic Fermentation. Int. J. Food Microbiol. 2003, 83, 49–61. [Google Scholar] [CrossRef]
- Ferreira, A.M.; Mendes-Faia, A. The Role of Yeasts and Lactic Acid Bacteria on the Metabolism of Organic Acids During Winemaking. Foods 2019, 9, 1231. [Google Scholar] [CrossRef]
- Younis, O.S.; Stewart, G.G. Sugar Uptake and Subsequent Ester and Higher Alcohol Production by Saccharomyces cerevisiae. J. Inst. Brew. 1998, 104, 255–264. [Google Scholar] [CrossRef]
- Lambrechts, M.; Pretorius, I. Yeast and Its Importance to Wine Aroma. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial Modulation of Aromatic Esters in Wine: Current Knowledge and Future Prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Pérez-Martín, F.; Seseña, S.; Izquierdo, P.M.; Palop, M.L. Esterase Activity of Lactic Acid Bacteria Isolated from Malolactic Fermentation of Red Wines. Int. J. Food Microbiol. 2013, 163, 153–158. [Google Scholar] [CrossRef]
- Lilly, M.; Styger, G.; Bauer, F.F.; Lambrechts, M.G.; Pretorius, I.S. The Effect of Increased Branched-Chain Amino Acid Transaminase Activity in Yeast on the Production of Higher Alcohols and on the Flavour Profiles of Wine and Distillates. FEMS Yeast Res. 2006, 6, 726–743. [Google Scholar] [PubMed] [Green Version]
- Swiegers, J.; Bartowsky, E.; Henschke, P.; Pretorius, I. Yeast and Bacterial Modulation of Wine Aroma and Flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- De Ovalle, S.; Cavello, I.; Brena, B.M.; Cavalitto, S.; González-Pombo, P. Production and Characterization of a β-Glucosidase from Issatchenkia terricola and Its Use for Hydrolysis of Aromatic Precursors in Cabernet Sauvignon Wine. LWT Food Sci. Technol. 2018, 87, 515–522. [Google Scholar] [CrossRef] [Green Version]
- Mauriello, G.; Capece, A.; D’Auria, M.; Garde-Cerdán, T.; Romano, P. SPME-GC Method as a Tool to Differentiate VOC Profiles in Saccharomyces cerevisiae Wine Yeasts. Food Microbiol. 2009, 26, 246–252. [Google Scholar] [CrossRef]
- Dubourdieu, D.; Torninaga, T.; Masneuf, I.; Des Gachons, C.P.; Murat, M.L. The Role of Yeasts in Grape Flavor Development During Fermentation: The Example of Sauvignon Blanc. Am. J. Enol. Vitic. 2006, 57, 81–88. [Google Scholar]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of Yeast Species and Strains in Wine Flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Renault, P.; Miot-Sertier, C.; Marullo, P.; Hernández-Orte, P.; Lagarrigue, L.; Lonvaud-Funel, A.; Bely, M. Genetic Characterization and Phenotypic Variability in Torulaspora delbrueckii Species: Potential Applications in the Wine Industry. Int. J. Food Microbiol. 2009, 134, 201–210. [Google Scholar] [CrossRef]
- Clemente-Jiménez, J.M.; Mingorance-Cazorla, L.; Martinez-Rodríguez, S.; Las Heras-Vázquez, F.J.; Rodríguez-Vico, F. Molecular Characterization and Oenological Properties of Wine Yeasts Isolated During Spontaneous Fermentation of Six Varieties of Grape Must. Food Microbiol. 2004, 21, 149–155. [Google Scholar] [CrossRef]
- Liu, Y.Z.; Rousseaux, S.; Tourdot-Marechal, R.; Sadoudi, M.; Gougeon, R.; Schmitt-Kopplin, P.; Alexandre, H. Wine Microbiome: A Dynamic World of Microbial Interactions. Crit. Rev. Food Sci. 2017, 57, 856–873. [Google Scholar]
- Bai, F.W.; Chen, L.J.; Zhang, Z.; Anderson, W.A.; Moo-Young, M. Continuous Ethanol Production and Evaluation of Yeast Cell Lysis and Viability Loss under Very High Gravity Medium Conditions. J. Biotechnol. 2004, 110, 287–293. [Google Scholar] [CrossRef]
- Teixeira, M.C.; Raposo, L.R.; Mira, N.P.; Lourenco, A.B.; Sa-Correia, I. Genome-Wide Identification of Saccharomyces cerevisiae Genes Required for Maximal Tolerance to Ethanol. Appl. Environ. Microbiol. 2009, 75, 5761–5772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archana, K.M.; Ravi, R.; Anu-Appaiah, K.A. Correlation between Ethanol Stress and Cellular Fatty Acid Composition of Alcohol Producing Non-Saccharomyces in Comparison with Saccharomyces cerevisiae by Multivariate Techniques. J. Food Sci. Technol. 2015, 52, 6770–6776. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Lee, S.B.; Jeon, J.Y.; Park, H.D. Development of Air-Blast Dried Non-Saccharomyces Yeast Starter for Improving Quality of Korean Persimmon Wine and Apple Cider. Int. J. Food Microbiol. 2019, 290, 193–204. [Google Scholar] [PubMed]
- Pérez-Torrado, R.; Querol, A.; Guillamón, J.M. Genetic Improvement of non-GMO Wine Yeasts: Strategies, Advantages and Safety. Trends Food Sci. Technol. 2015, 45, 1–11. [Google Scholar] [CrossRef]
- Farahnak, F.; Seki, T.; Ryu, D.D.Y.; Ogrydziak, D. Construction of Lactose-Assimilating and High-Ethanol-Producing Yeasts by Protoplast Fusion. Appl. Environ. Microbiol. 1986, 51, 362–367. [Google Scholar] [CrossRef] [Green Version]
- Pattanakittivorakul, S.; Lertwattanasakul, N.; Yamada, M.; Limtong, S. Selection of Thermotolerant Saccharomyces cerevisiae for High Temperature Ethanol Production from Molasses and Increasing Ethanol Production by Strain Improvement. Antonie Leeuwenhoek 2019, 112, 975–990. [Google Scholar] [CrossRef]
- Watanabe, T.; Watanabe, I.; Yamamoto, M.; Ando, A.; Nakamura, T. A UV-Induced Mutant of Pichia stipitis with Increased Ethanol Production from Xylose and Selection of a Spontaneous Mutant with Increased Ethanol Tolerance. Bioresour. Technol. 2011, 102, 1844–1848. [Google Scholar]
- Dong, Y.; Zhang, N.; Lu, J.H.; Lin, F.; Teng, L.R. Improvement and Optimization of the Media of Saccharomyces cerevisiae Strain for High Tolerance and High Yield of Ethanol. Afr. J. Microbiol. Res. 2012, 6, 2357–2366. [Google Scholar]
- Liang, M.H.; Liang, Y.J.; Wu, X.N.; Zhou, S.S.; Jiang, J.G. Mutation Breeding of Saccharomyces cerevisiae with Lower Methanol Content and the Effects of Pectinase, Cellulase and Glycine in Sugar Cane Spirits. J. Sci. Food Agric. 2015, 95, 1949–1955. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, M.; Kiran Sree, N.; Venkateswar, R.L. Effect of UV Radiation on Thermotolerance, Ethanol Tolerance and Osmotolerance of Saccharomyces cerevisiae VS1 and VS3 Strains. Bioresour. Technol. 2002, 83, 199–202. [Google Scholar] [CrossRef]
- Wen, L.K.; Wang, L.F.; Wang, G.Z. Degradation of L-Malic and Critic Acids by Issatchenkia terricola. Food Sci. 2011, 32, 220–223. [Google Scholar]
- Yoshikawa, K.; Tanaka, T.; Furusawa, C.; Nagahisa, K.; Hirasawa, T.; Shimizu, H. Comprehensive Phenotypic Analysis for Identification of Genes Affecting Growth under Ethanol Stress in Saccharomyces cerevisiae. FEMS Yeast Res. 2009, 9, 32–44. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Ethanol Concentration (% (v/v)) | ||||
|---|---|---|---|---|---|
| 4 | 6 | 8 | 10 | 12 | |
| D1 | + | - | - | - | - |
| D6 | + | + | + | - | - |
| B21 | + | + | - | - | - |
| E31 | + | + | - | - | - |
| H5 | + | + | + | - | - |
| F26 | + | + | + | - | - |
| Strain | Ethanol Concentration (% (v/v)) | ||||
|---|---|---|---|---|---|
| 8 | 9 | 10 | 11 | 12 | |
| S1 | +++ | +++ | ++ | ++ | ++ |
| H5 | + | - | - | - | - |
| UV1 | - | - | - | - | - |
| UV2 | - | - | - | - | - |
| UV3 | + | - | - | - | - |
| UV4 | - | - | - | - | - |
| UV5 | +++ | ++ | ++ | + | - |
| UV6 | - | - | - | - | - |
| UV7 | + | - | - | - | - |
| UV8 | +++ | ++ | ++ | - | - |
| DES1 | - | - | - | - | - |
| DES2 | - | - | - | - | - |
| DES3 | - | - | - | - | - |
| DES4 | + | - | - | - | - |
| DES5 | + | - | - | - | - |
| DES6 | - | - | - | - | - |
| DES7 | ++ | - | - | - | - |
| Grape Must (Day 0) | Wine (Day 16) | |||||
|---|---|---|---|---|---|---|
| S1 | H5 | DES7 | UV5 | UV8 | ||
| °Brix | 18.77 ± 0.05 a | 5.67 ± 0.09 e | 15.20 ± 0.28 b | 14.90 ± 0.08 b | 10.43 ± 0.25 d | 12.27 ± 0.42 c |
| Ethanol (% v/v) | N.D. | 11.32 ± 0.12 a | 2.05 ± 0.10 d | 2.28 ± 0.07 cd | 6.55 ± 0.06 b | 2.44 ± 0.17 c |
| Glycerol (g/L) | N.D. | 10.45 ± 0.08 a | 7.07 ± 0.18 c | 7.62 ± 0.31 bc | 8.22 ± 0.51 b | 6.81 ± 0.75 c |
| Sugar (g/L) | ||||||
| Glucose | 105.08 ± 0.76 a | N.D. | 61.18 ± 2.53 b | 62.78 ± 1.26 b | 15.17 ± 2.34 d | 31.83 ± 3.58 c |
| Fructose | 106.05 ± 1.00 a | 2.16 ± 0.08 e | 63.39 ± 2.14 c | 69.12 ± 2.82 b | 40.10 ± 1.59 d | 68.59 ± 2.80 b |
| Acid (g/L) | ||||||
| Citric acid | 0.81 ± 0.02 b | 0.75 ± 0.02 c | 0.67 ± 0.02 d | 0.58 ± 0.02 e | 0.77 ± 0.01 bc | 0.88 ± 0.03 a |
| Tartaric acid | 3.00 ± 0.05 a | 2.28 ± 0.08 c | 3.06 ± 0.00 a | 2.96 ± 0.02 a | 1.87 ± 0.03 d | 2.51 ± 0.11 b |
| Malic acid | 4.29 ± 0.08 a | 2.86 ± 0.07 d | 2.29 ± 0.03 f | 2.59 ± 0.03 e | 3.53 ± 0.07 b | 3.30 ± 0.08 c |
| Succinic acid | 3.59 ± 0.12 c | 4.42 ± 0.13 a | 3.82 ± 0.31 bc | 3.66 ± 0.09 c | 4.15 ± 0.03 ab | 2.94 ± 0.21 d |
| Lactic acid | N.D. | N.D. | 1.05 ± 0.22 a | 0.86 ± 0.14 a | N.D. | N.D. |
| Acetic acid | 0.06 ± 0.01 c | 0.11 ± 0.00 b | 0.15 ± 0.02 a | 0.11 ± 0.01 b | 0.09 ± 0.00 b | 0.15 ± 0.01 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, J.; He, X.; Huang, W.; You, Y.; Zhan, J. Enhancing Ethanol Tolerance via the Mutational Breeding of Pichia terricola H5 to Improve the Flavor Profiles of Wine. Fermentation 2022, 8, 149. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8040149
Gao J, He X, Huang W, You Y, Zhan J. Enhancing Ethanol Tolerance via the Mutational Breeding of Pichia terricola H5 to Improve the Flavor Profiles of Wine. Fermentation. 2022; 8(4):149. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8040149
Chicago/Turabian StyleGao, Jie, Xiuli He, Weidong Huang, Yilin You, and Jicheng Zhan. 2022. "Enhancing Ethanol Tolerance via the Mutational Breeding of Pichia terricola H5 to Improve the Flavor Profiles of Wine" Fermentation 8, no. 4: 149. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8040149