
Influence of Light Conditions and Medium Composition on Morphophysiological Characteristics of Stevia rebaudiana Bertoni In Vitro and In Vivo


 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Influence of PGRs on S. rebaudiana Morphogenesis In Vitro
2.3. Influence of Light Factors on the Morphophysiological Parameters of S. rebaudiana In Vitro
2.4. The Combined Effect of Physical and PGRs Factors on the Morphophysiological Parameters of In Vitro S. rebaudiana Microshoot Growth
2.5. Phenotypic Analysis of Plants
2.6. Quantitative Determination of Photosynthetic Pigments of S. rebaudiana In Vivo
2.7. Qualitative and Semi-Quantitative Determination of Diterpene Glycosides in Dry Leaves of S. rebaudiana In Vivo
2.8. Quantitative Determination of Steviosides by High-Performance Liquid Chromatography (HPLC) of Dry Leaves of S. rebaudiana In Vivo
2.9. Statistical Analysis of Experimental Data
3. Results and Discussion
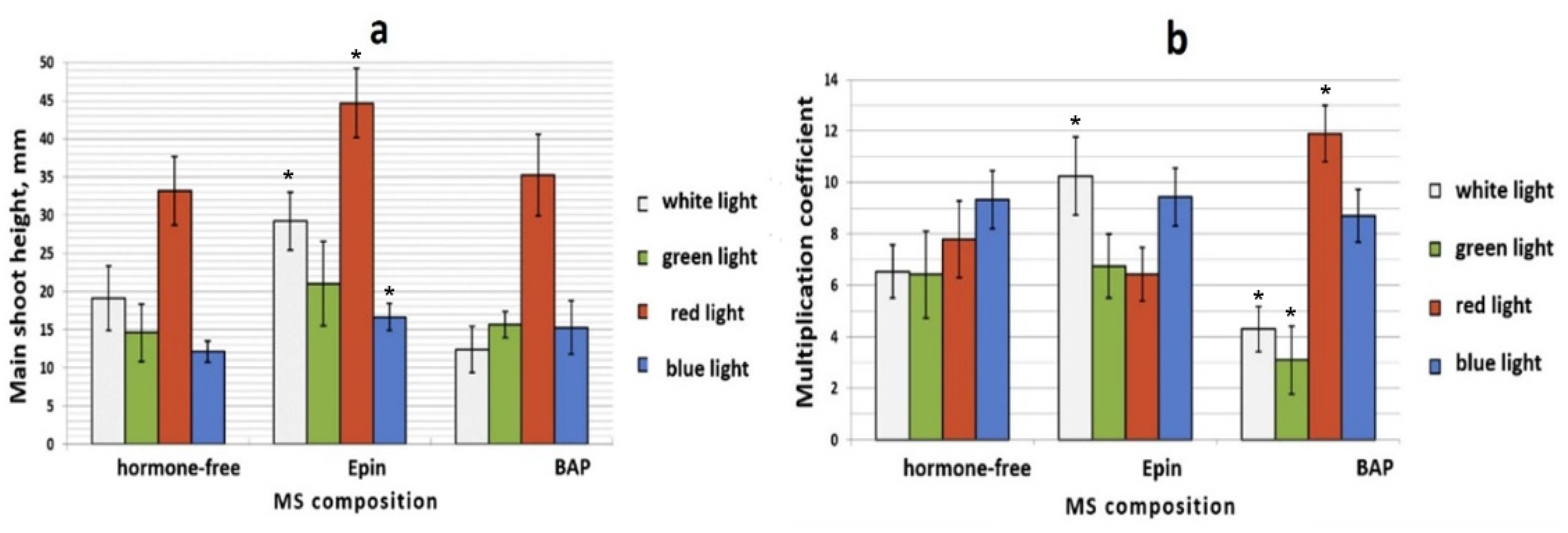
3.1. Influence of PGRs on In Vitro S. rebaudiana Clonal Propagation and Microshoot Morphology
3.2. Influence of Physical Factors on the Morphophysiological Parameters of S. rebaudiana In Vitro
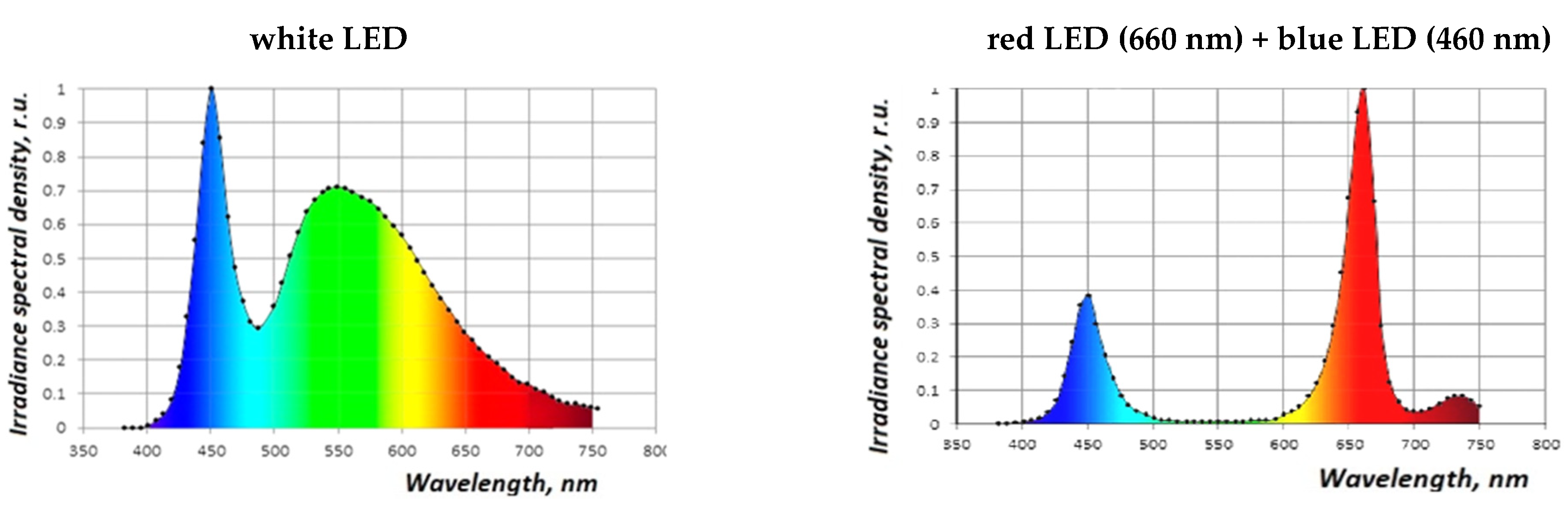
3.2.1. Effect of Red-Blue LED Lighting on Morphophysiological Parameters of S. rebaudiana Microshoots
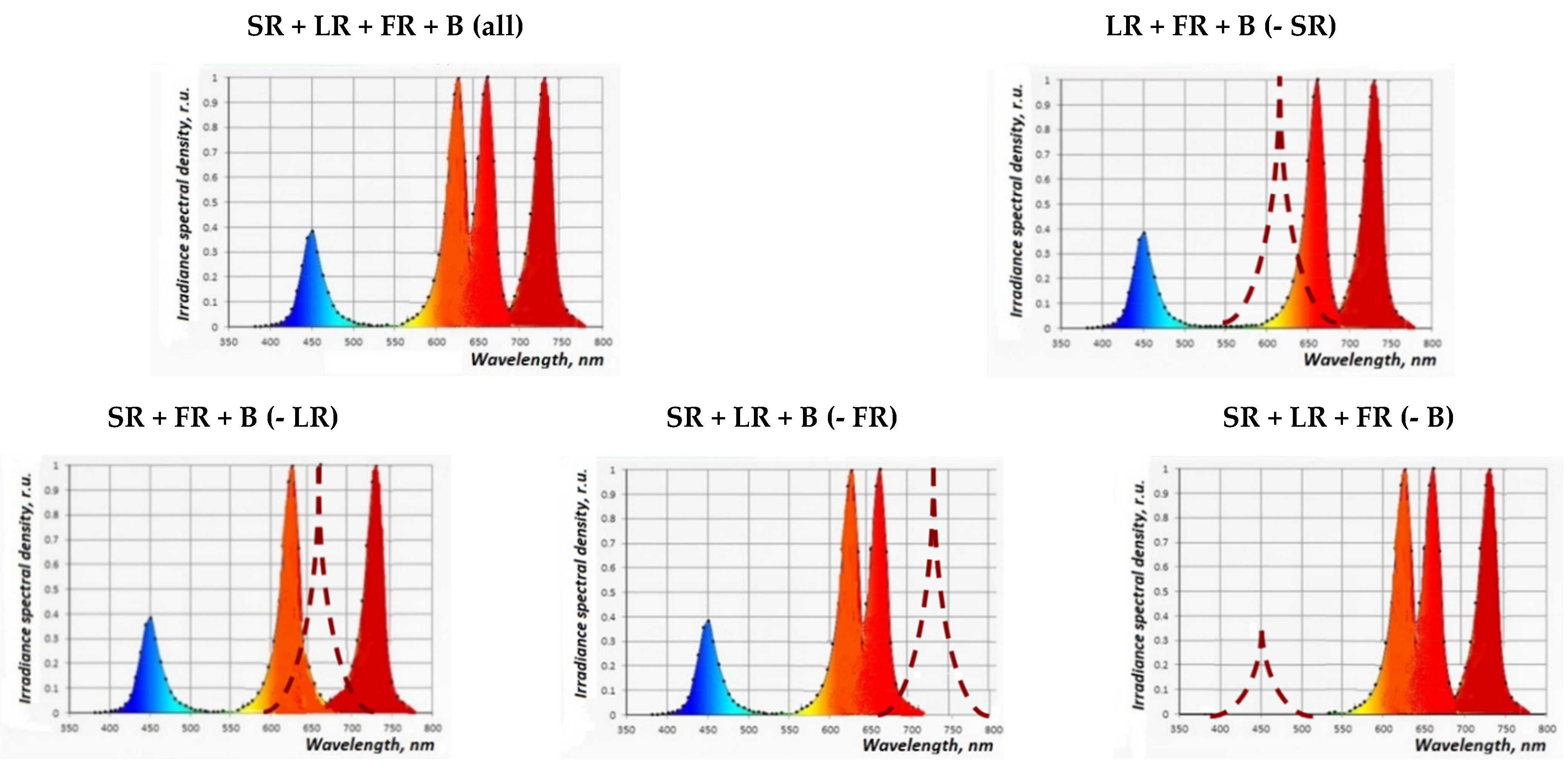
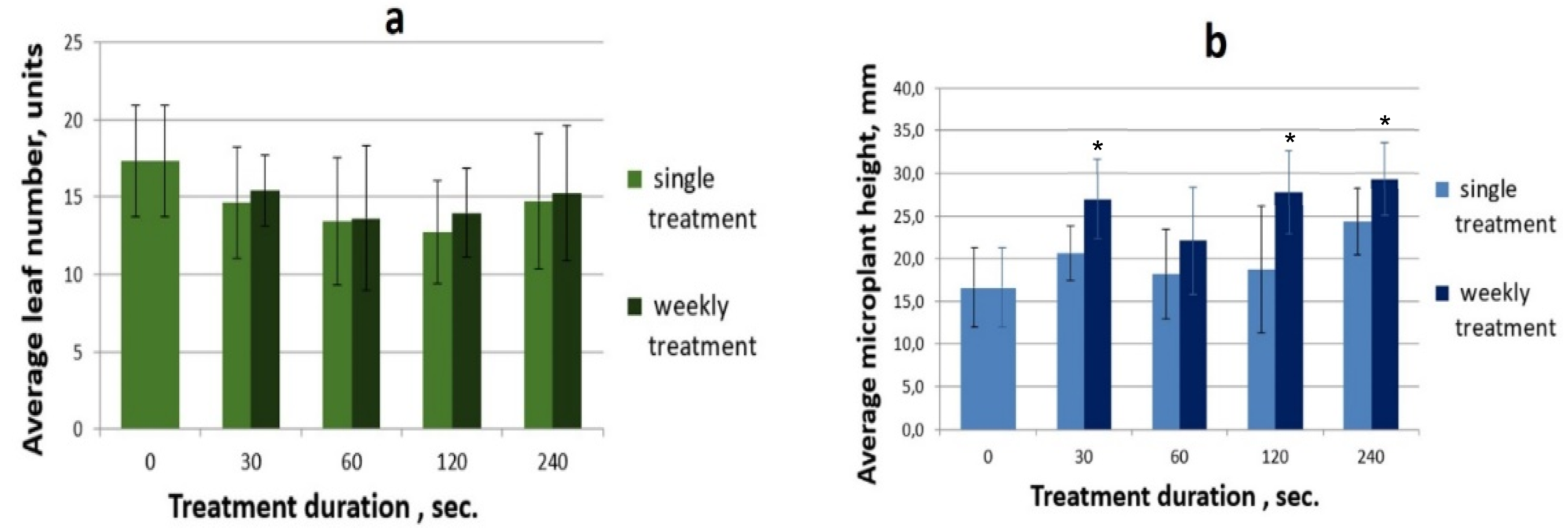
3.2.2. The Effect of Coherent Light Treatment on the Morphophysiological Characteristics of S. rebaudiana Microshoots In Vitro
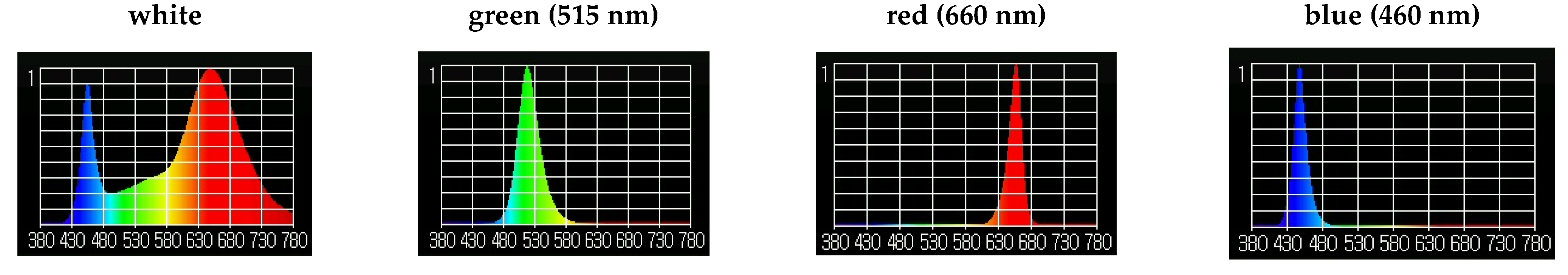
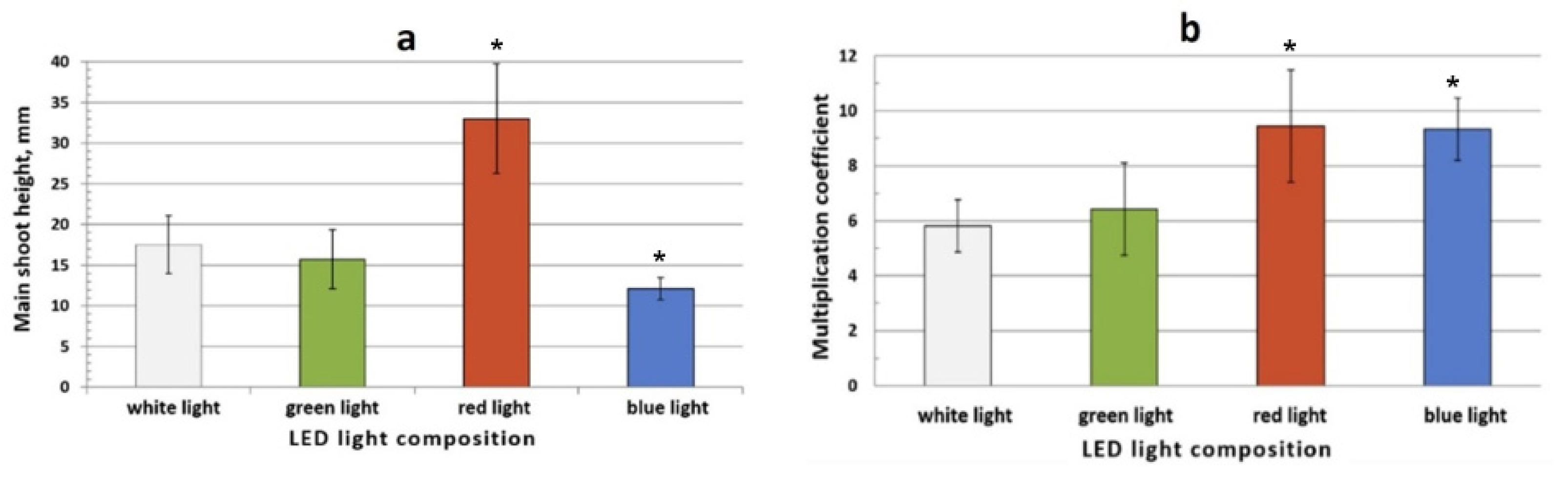
3.2.3. The Effect of Monochrome Light Treatment on the Morphophysiological Characteristics of S. rebaudiana Microshoots In Vitro
3.2.4. The Combined Effect of Physical and PGRs Factors on the Morphogenesis of S. rebaudiana Microshoots Culture In Vitro
Monochrome Light Treatment in Combination with PGRs under In Vitro Conditions
Coherent Light Treatment in Combination with PGRs under In Vitro Conditions
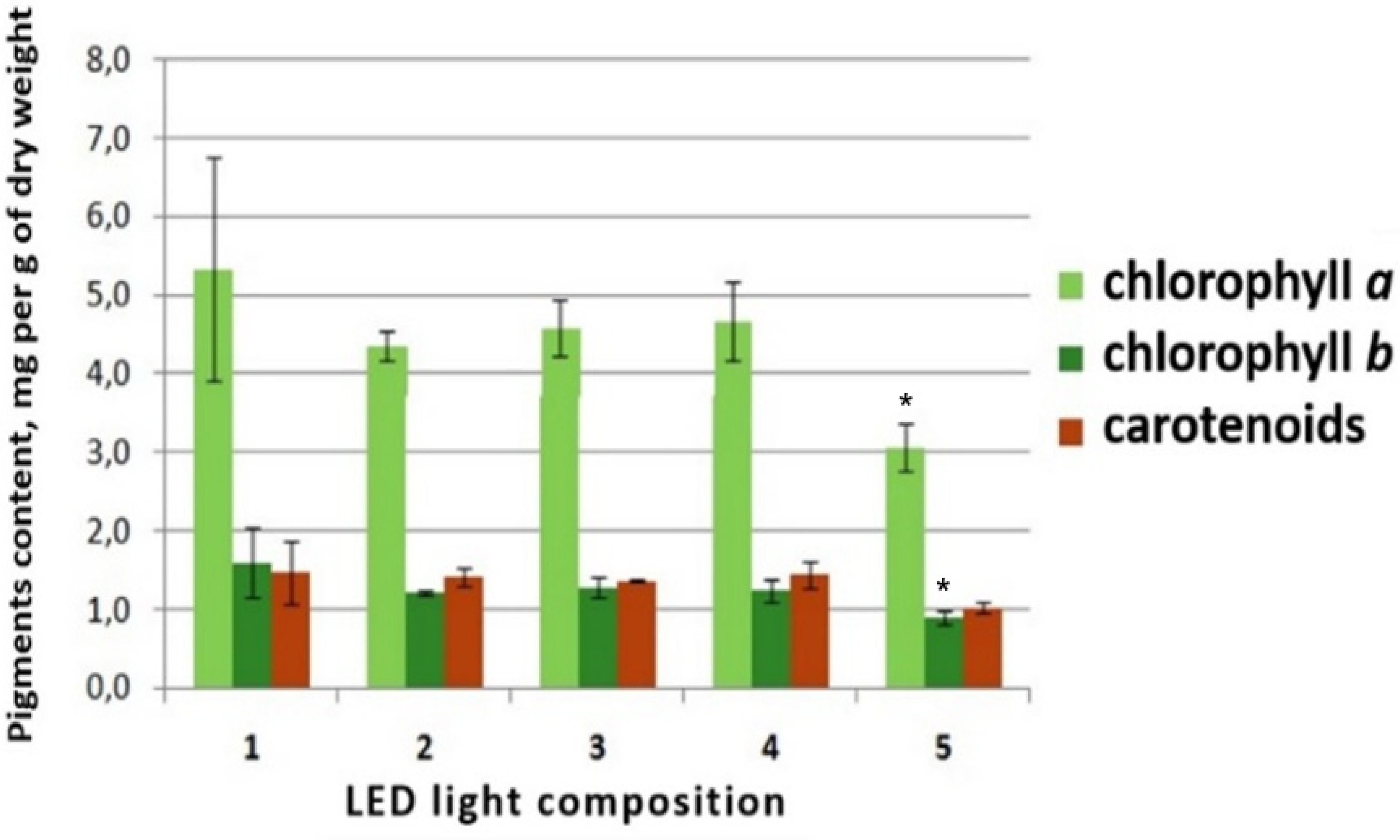
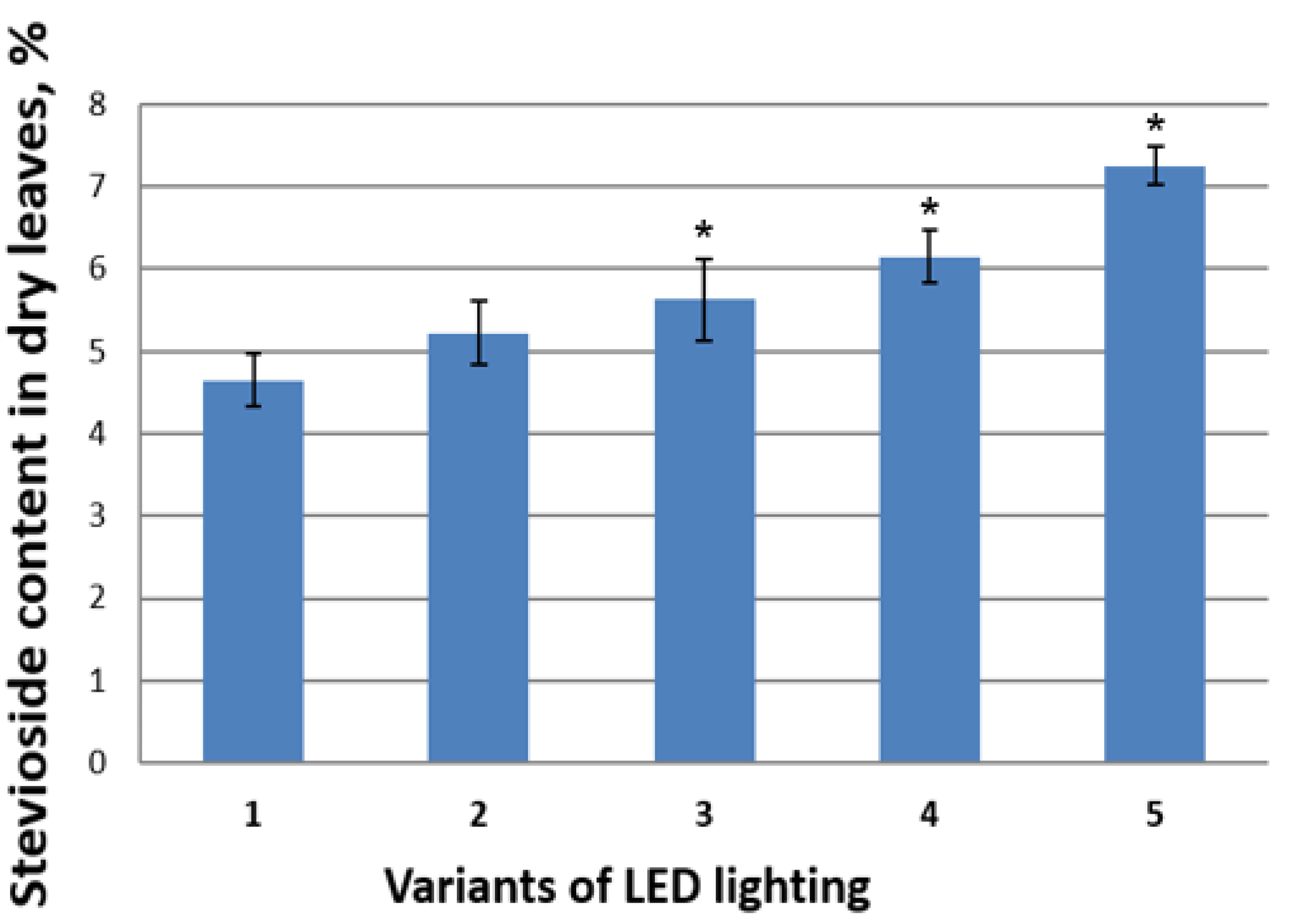
3.3. Effects of Red-Blue LED Lighting on the Biochemical Characteristics of S. rebaudiana Plants In Vivo
3.3.1. Quantitative Determination of Photosynthetic Pigments
3.3.2. Qualitative and Semi-Quantitative Determination of Diterpene Glycosides by TLC
3.3.3. Quantitative Determination of Stevioside by HPLC
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saraiva, A.; Carrascosa, C.; Raheem, D.; Ramos, F.; Raposo, A. Natural sweeteners: The relevance of food naturalness for consumers, food security aspects, sustainability and health impacts. Int. J. Environ. Res. Public Health 2020, 17, 6285. [Google Scholar] [CrossRef]
- Bergman, M.E.; Davis, B.; Phillips, M.A. Medically useful plant terpenoids: Biosynthesis, occurrence, and mechanism of action. Molecules 2019, 24, 3961. [Google Scholar] [CrossRef] [Green Version]
- Lanzotti, V. Diterpenes for Therapeutic Use, in Natural Products; Springer: Berlin/Heidelberg, Germany, 2013; p. 3173. [Google Scholar]
- Goel, M.K.; Kukreja, A.K.; Singh, A.K.; Khanuja, S.P.S. In vitro plant growth promoting activity of phyllocladane diterpenoids isolated from Callicarpa macrophylla Vahl. in shoot cultures of Rauwolfia serpentina. Nat. Prod. Commun. 2007, 2, 1934578X0700200802. [Google Scholar] [CrossRef]
- Abbaszadeh, G.; Srivastava, C.; Walia, S. Insecticidal and antifeedant activities of clerodane diterpenoids isolated from the Indian bhant tree, Clerodendron infortunatum, against the cotton bollworm, Helicoverpa armigera. J. Insect Sci. 2014, 14, 1. [Google Scholar] [CrossRef]
- Urzúa, A.; Rezende, M.C.; Mascayano, C.; Vásquez, L. A structure-activity study of antibacterial diterpenoids. Molecules 2008, 13, 882–891. [Google Scholar] [CrossRef] [Green Version]
- Bozov, P.; Girova, T.; Prisadova, N.; Hristova, Y.; Gochev, V. Antimicrobial activity of neo-clerodane diterpenoids isolated from Lamiaceae species against pathogenic and food spoilage microorganisms. Nat. Prod. Commun. 2015, 10, 1934578X1501001101. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.T. Diterpenes and their derivatives as potential anticancer agents. Phytother. Res. 2017, 31, 691–712. [Google Scholar] [CrossRef]
- Jian, B.; Zhang, H.; Han, C.; Liu, J. Anti-cancer activities of diterpenoids derived from Euphorbia fischeriana Steud. Molecules 2018, 23, 387. [Google Scholar] [CrossRef] [Green Version]
- Koyama, E.; Kitazawa, K.; Ohori, Y.; Izawa, O.; Kakegawa, K.; Fujino, A.; Ui, M. In vitro metabolism of the glycosidic sweeteners, stevia mixture and enzymatically modified stevia in human intestinal microflora. Food Chem. Toxicol. 2003, 41, 359–374. [Google Scholar] [CrossRef]
- Bondarev, N.I.; Sukhanova, M.A.; Reshetnyak, O.V.; Nosov, A.M. Steviol glycoside content in different organs of Stevia rebaudiana and its dynamics during ontogeny. Biol. Plant. 2003, 47, 261–264. [Google Scholar] [CrossRef]
- Muanda, F.N.; Soulimani, R.; Diop, B.; Dicko, A. Study on chemical composition and biological activities of essential oil and extracts from Stevia rebaudiana Bertoni leaves. LWT 2011, 44, 1865–1872. [Google Scholar] [CrossRef]
- Chen, T.H.; Chen, S.C.; Chan, P.; Chu, Y.; Yang, H.Y.; Cheng, J.T. Mechanism of the hypoglycemic effect of stevioside, a glycoside of Stevia rebaudiana. Planta Med. 2005, 71, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Megeji, N.W.; Kumar, J.K.; Singh, V.; Kaul, V.K.; Ahuja, P.S. Introducing Stevia rebaudiana, a natural zero-calorie sweetener. Curr. Sci. 2005, 88, 801–804. [Google Scholar]
- Gupta, E.; Purwar, S.; Sundaram, S.; Rai, G.K. Nutritional and therapeutic values of Stevia rebaudiana: A review. J. Med. Plants Res. 2013, 7, 3343–3353. [Google Scholar]
- Jeppesen, P.B.; Gregersen, S.; Alstrup, K.K.; Hermansen, K. Stevioside induces antihyperglycaemic, insulinotropic and glucagonostatic effects in vivo: Studies in the diabetic Goto-Kakizaki (GK) rats. Phytomedicine 2002, 9, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Starratt, A.N.; Kirby, C.W.; Pocs, R.; Brandle, J.E.; Rebaudioside, F. A diterpene glycoside from Stevia rebaudiana. Phytochemistry 2002, 59, 367–370. [Google Scholar] [CrossRef]
- Tadhani, M.B.; Patel, V.H.; Subhash, R. In vitro antioxidant activities of Stevia rebaudiana leaves and callus. J. Food Compos. Anal. 2007, 20, 323–329. [Google Scholar] [CrossRef]
- Brandle, J.E.; Starratt, A.N.; Gijzen, M. Stevia rebaudiana: Its agricultural, biological, and chemical properties. Can. J. Plant Sci. 1998, 78, 527–536. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.B.; Salahin, M.; Karim, R.; Razvy, M.A.; Hannan, M.M.; Sultana, R.; Hossain, M.; Islam, R. An efficient method for in vitro clonal propagation of a newly introduced sweetener plant (Stevia rebaudiana Bertoni.) in Bangladesh. Am. Eurasian J. Sci. Res. 2007, 2, 121–125. [Google Scholar]
- Bondarev, N.; Reshetnyak, O.; Nosov, A. Peculiarities of diterpenoid steviol glycoside production in in vitro cultures of Stevia rebaudiana Bertoni. Plant Sci. 2001, 161, 155–163. [Google Scholar] [CrossRef]
- Ferreira, C.M.; Handro, W. Production, maintenance and plant regeneration from cell suspension cultures of Stevia rebaudiana (Bert.) Bertoni. Plant Cell Rep. 1988, 7, 123–126. [Google Scholar] [CrossRef]
- Gantait, S.; Das, A.; Mandal, N. Stevia: A comprehensive review on ethnopharmacological properties and in vitro regeneration. Sugar Tech. 2015, 17, 95–106. [Google Scholar] [CrossRef]
- Giridhar, P.; Sowmya, K.S.; Ramakrishna, A.; Ravishankar, G.A. Rapid clonal propagation and stevioside profiles of Stevia rebaudiana Bertoni. Int. J. Plant Dev. Biol. 2010, 4, 47–52. [Google Scholar]
- Guruchandran, V.; Sasikumar, C. Organogenic plant regeneration via callus induction in Stevia rebaudiana Bert. Int. J. Curr. Microbiol. Appl. Sci. 2013, 2, 56–61. [Google Scholar]
- Jarma-Orozco, A.; Combatt-Caballero, E.; Jaraba-Navas, J. Growth and development of Stevia rebaudiana Bert., in high and low levels of radiation. Curr. Plant Biol. 2020, 22, 100144. [Google Scholar] [CrossRef]
- Kalpana, M.; Anbazhagan, M.; Natarajan, V.; Dhanavel, D. Improved micropropagation method for the enhancement of biomass in Stevia rebaudiana Bertoni. Rec. Res. Sci. Tech. 2010, 2, 8–13. [Google Scholar]
- Karim, M.A.; Jannat, R.; Rahman, M.S.; Haque, M.S. Micropropagation of stevia plant from nodal segments. Prog. Agric. 2008, 19, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Kulchin, Y.N.; Nakonechnaya, O.V.; Gafitskaya, I.V.; Grishchenko, O.V.; Epifanova, T.Y.; Orlovskaya, I.Y.; Zhuravlev, Y.N.; Subbotin, E.P. Plant morphogenesis under different light intensity. Defect Diffus. Forum Trans. Tech. Publ. 2018, 386, 201–206. [Google Scholar] [CrossRef]
- Melviana, A.C.; Esyanti, R.R.; Mel, M.; Setyobudi, R.H. Biomass enhancement of Stevia rebaudiana Bertoni Shoot culture in temporary immersion system (TIS) RITA® bioreactor optimized in two different immersion periods. In Proceedings of the 1st International Conference on Bioenergy and Environmentally Sustainable Agriculture Technology (ICoN BEAT 2019), East Java, Indonesia, 7–8 November 2019; p. 00007. [Google Scholar]
- Razak, U.N.A.A.; Ong, C.B.; Yu, T.S.; Lau, L.K. In vitro micropropagation of Stevia rebaudiana Bertoni in Malaysia. Braz. Arch. Biol. Technol. 2014, 57, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Muthu, S.; Schuurmans, F.J.; Pashley, M.D. Red, green, and blue LEDs for white light illumination. IEEE J. Sel. Top. Quantum Electron. 2002, 8, 333–338. [Google Scholar] [CrossRef] [Green Version]
- Sivaram, L.; Mukundan, U. In vitro culture studies on Stevia rebaudiana. Vitro Cell. Dev. Biol. Plant 2003, 39, 520–523. [Google Scholar] [CrossRef]
- Ahmad, A.; Ali, H.; Khan, H.; Begam, A.; Khan, S.; Ali, S.S.; Ahmad, N.; Fazal, H.; Ali, M.; Hano, C.; et al. Effect of Gibberellic Acid on Production of Biomass, Polyphenolics and Steviol Glycosides in Adventitious Root Cultures of Stevia rebaudiana (Bert.). Plants 2020, 9, 420. [Google Scholar] [CrossRef] [Green Version]
- Blinstrubienė, A.; Burbulis, N.; Juškevičiūtė, N.; Vaitkevičienė, N.; Žūkienė, R. Effect of Growth Regulators on Stevia rebaudianaBertoni Callus Genesis and Influence of Auxin and Proline to Steviol Glycosides, Phenols, Flavonoids Accumulation, and Antioxidant Activity In Vitro. Molecules 2020, 25, 2759. [Google Scholar] [CrossRef] [PubMed]
- Bondarev, N.; Reshetnyak, O.; Bondareva, T.; Il’in, M.; Nosov, A. Impact of Cultivation Factors in vitro on the Growth and the Biosynthesis of Steviol Glycosides in Stevia rebaudiana cell Cultures. Physiol. Mol. Biol. Plants 2019, 25, 1091–1096. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Sharma, S.; Saxena, S. Biomass Yield and Steviol Glycoside Production in Callus and Suspension Culture of Stevia rebaudiana Treated with Proline and Polyethylene Glycol. Appl. Biochem. Biotechnol. 2015, 176, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Libik-Konieczny, M.; Capecka, E.; Tuleja, M.; Konieczny, R. Synthesis and production of steviol glycosides: Recent research trends and perspectives. Appl. Microbiol. Biotechnol. 2021, 105, 3883–3900. [Google Scholar] [CrossRef] [PubMed]
- Rameeh, V.; Gerami, M.; Omran, V.G.; Ghavampour, S. Impact of glycine betaine on salinity tolerance of stevia (Stevia rebaudiana Bertoni) under in vitro condition. Cercet. Agron. Mold. 2017, 3, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Sharma, S.; Saxena, S. Effect of abiotic stress on growth parameters and steviol glycoside content in Stevia rebaudiana (Bertoni) raised in vitro. J. Appl. Res. Med. Aromat. Plants 2016, 3, 160–167. [Google Scholar] [CrossRef]
- Lucho, S.R.; do Amaral, M.N.; Auler, P.A.; Bianchi, V.J.; María Ángeles Ferrer, M.A.; Calderón, A.A.; Bolacel Braga, E.J. Salt stress-induced changes in in vitro cultured Stevia rebaudiana Bertoni: Effect on metabolite contents, antioxidant capacity and expression of steviol glycosides-related biosynthetic genes. J. Plant. Growth Regul. 2019, 38, 1341–1353. [Google Scholar] [CrossRef]
- Fallah, F.; Nokhasi, F.; Ghaheri, M.; Kahrizi, D.; Beheshti, A.A.A.; Ghorbani, T.; Kazemi, E.; Ansarypou, Z. Effect of salinity on gene expression, morphological and biochemical characteristics of Stevia rebaudiana Bertoni under in vitro conditions. Cell Mol. Biol. 2017, 63, 102–103. [Google Scholar] [CrossRef]
- Pandey, M.; Chikara, S.K. Effect of salinity and drought stress on growth parameters, glycoside content and expression level of vital genes in steviol glycosides biosynthesis pathway of Stevia rebaudiana (Bertoni). Int. J. Genet. 2015, 7, 153–160. [Google Scholar]
- Bayraktar, M.; Naziri, E.; Akgun, I.H.; Karabey, F.; Ilhan, E.; Akyol, B.; Bedir, E.; Gurel, A. Elicitor induced stevioside production, in vitro shoot growth, and biomass accumulation in micropropagated Stevia rebaudiana. Plant Cell Tissue Organ Cult. 2016, 127, 289–300. [Google Scholar] [CrossRef]
- Ghazal, B.; Saif, S.; Farid, K.; Khan, A.; Rehman, S.; Reshma, A.; Fazal, H.; Ali, M.; Ahmad, A.; Rahman, L.; et al. Stimulation of secondary metabolites by copper and gold nanoparticles in submerge adventitious root cultures of Stevia rebaudiana (Bert.). IET Nanobiotechnol. 2018, 12, 569–573. [Google Scholar] [PubMed]
- Tarakanov, I.G. Light control of growth and development in vegetable plants with various life strategies. Acta Hortic. 2006, 711, 315–321. [Google Scholar] [CrossRef]
- Tarakanov, I.; Yakovleva, O.; Konovalova, I.; Paliutina, G.; Anisimov, A. Light-emitting diodes: On the way to combinatorial lighting technologies for basic research and crop production. Acta Hortic. 2012, 956, 171–178. [Google Scholar] [CrossRef]
- Nakonechnaya, O.V.; Gafitskaya, I.V.; Burkovskaya, E.V.; Khrolenko, Y.A.; Grishchenko, O.V.; Zhuravlev, Y.N.; Sussotin, E.P.; Kulchin, Y.N. Effect of light intensity on the morphogenesis of Stevia rebaudiana under in vitro conditions. Russ. J. Plant Physiol. 2019, 66, 656–663. [Google Scholar] [CrossRef]
- Bondarev, N.I.; Reshetnyak, O.V.; Nosov, A.M. Influence of photoperiod and irradiation intensity on the development of Stevia rebaudiana shoots in vitro and synthesis of steviol glycosides. Izv. TSKhA 2008, 4, 102–107. [Google Scholar]
- Idrees, M.; Sania, B.; Hafsa, B.; Kumari, S.; Khan, H.; Fazal, H.; Ahmad, I.; Akbar, F.; Ahmad, N.; Ali, S.; et al. Spectral lights trigger biomass accumulation and production of antioxidant secondary metabolites in adventitious root cultures of Stevia rebaudiana (Bert.). C. R. Biol. 2018, 341, 334–342. [Google Scholar] [CrossRef]
- Ahmad, N.; Rab, A.; Ahmad, N. Light-induced biochemical variations in secondary metabolite production and antioxidant activity in callus cultures of Stevia rebaudiana (Bert). J. Photochem. Photobiol. B 2016, 154, 6–51. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Jaitak, V.; Gupta, A.P.; Kaul, V.K.; Ahuja, P.S. Validated high-performance thin-layer chromatography method for steviol glycosides in Stevia rebaudiana. J. Pharm. Biomed. Anal. 2008, 47, 790–794. [Google Scholar] [CrossRef]
- Rokosa, M.T.; Kulpa, D. Micropropagation of Stevia rebaudiana plants. Ciência Rural 2020, 50, e20181029. [Google Scholar] [CrossRef] [Green Version]
- Nower, A.A. In vitro propagation and synthetic seeds production: An efficient method for Stevia rebaudiana Bertoni. Sugar Tech. 2014, 16, 100–108. [Google Scholar] [CrossRef]
- Bayraktar, M. Micropropagation of Stevia rebaudiana Bertoni using RITA®® bioreactor. HortScience 2019, 54, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Lata, H.; Chandra, S.; Wang, Y.H.; ElSohly, M.A.; Khan, I.A. Polyhouse cultivation of in vitro raised elite Stevia rebaudiana Bertoni: An assessment of biochemical and photosynthetic characteristics. Int. J. Trop. Agric. 2015, 33, 2381–2389. [Google Scholar]
- Ceunen, S.; Werbrouck, S.; Geuns, J.M. Stimulation of steviol glycoside accumulation in Stevia rebaudiana by red LED light. J. Plant Physiol. 2012, 169, 749–752. [Google Scholar] [CrossRef]
- Nikolova-Damyanova, B.; Bankova, V.; Popov, S. Separation and quantitation of stevioside and rebaudioside a in plant extracts by normal-phase high performance liquid chromatography and thin-layer chromatography: A comparison. Phytochem. Anal. 1994, 5, 81–85. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shulgina, A.A.; Kalashnikova, E.A.; Tarakanov, I.G.; Kirakosyan, R.N.; Cherednichenko, M.Y.; Polivanova, O.B.; Baranova, E.N.; Khaliluev, M.R. Influence of Light Conditions and Medium Composition on Morphophysiological Characteristics of Stevia rebaudiana Bertoni In Vitro and In Vivo. Horticulturae 2021, 7, 195. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7070195
Shulgina AA, Kalashnikova EA, Tarakanov IG, Kirakosyan RN, Cherednichenko MY, Polivanova OB, Baranova EN, Khaliluev MR. Influence of Light Conditions and Medium Composition on Morphophysiological Characteristics of Stevia rebaudiana Bertoni In Vitro and In Vivo. Horticulturae. 2021; 7(7):195. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7070195
Chicago/Turabian StyleShulgina, Alla A., Elena A. Kalashnikova, Ivan G. Tarakanov, Rima N. Kirakosyan, Mikhail Yu. Cherednichenko, Oksana B. Polivanova, Ekaterina N. Baranova, and Marat R. Khaliluev. 2021. "Influence of Light Conditions and Medium Composition on Morphophysiological Characteristics of Stevia rebaudiana Bertoni In Vitro and In Vivo" Horticulturae 7, no. 7: 195. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7070195