
Expression Analysis and Interaction Protein Screening of CRY1 in Strawberry

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
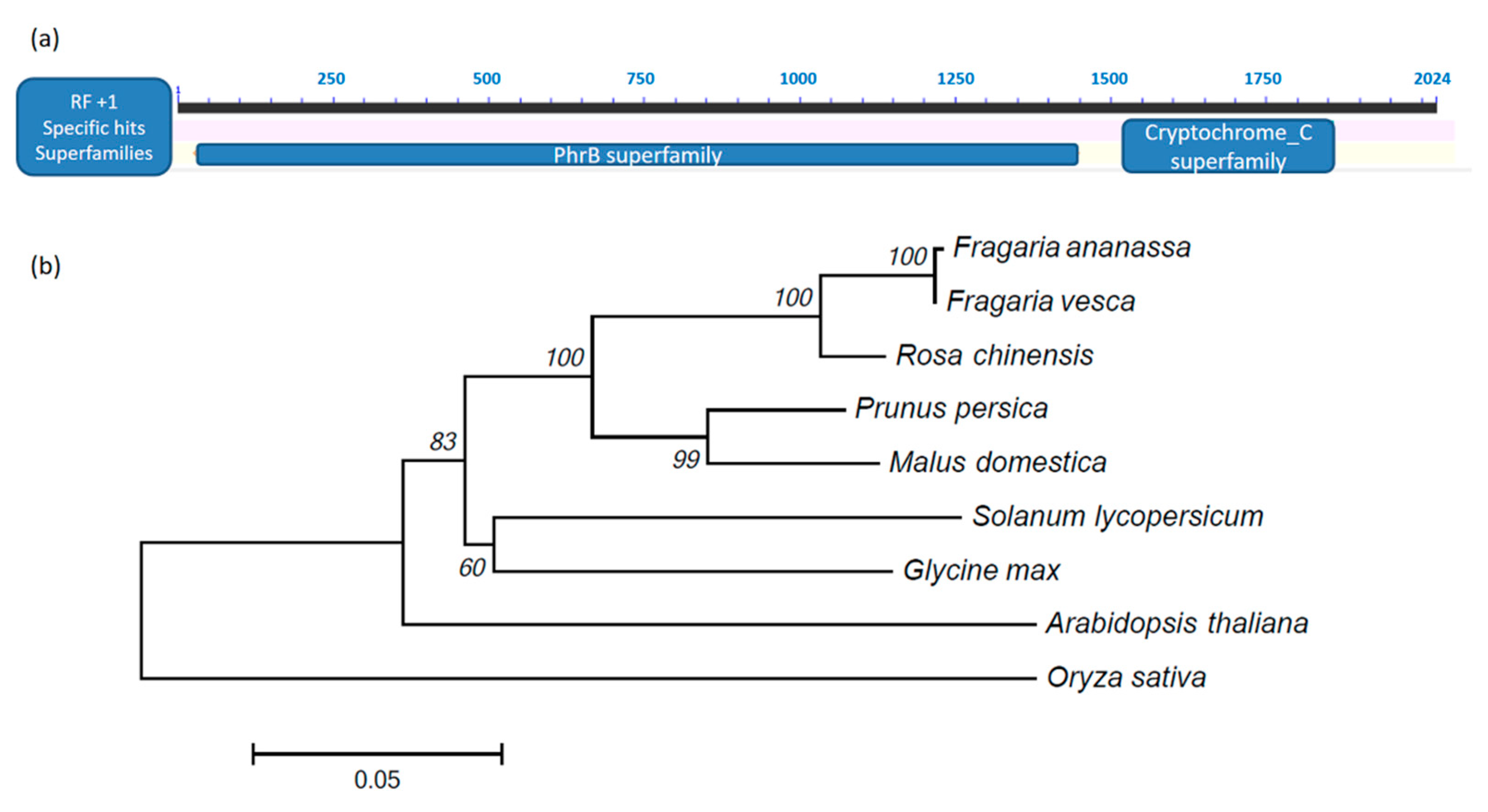
2.2. Gene Cloning, Sequence Analysis and Phylogenetic Tree Construction
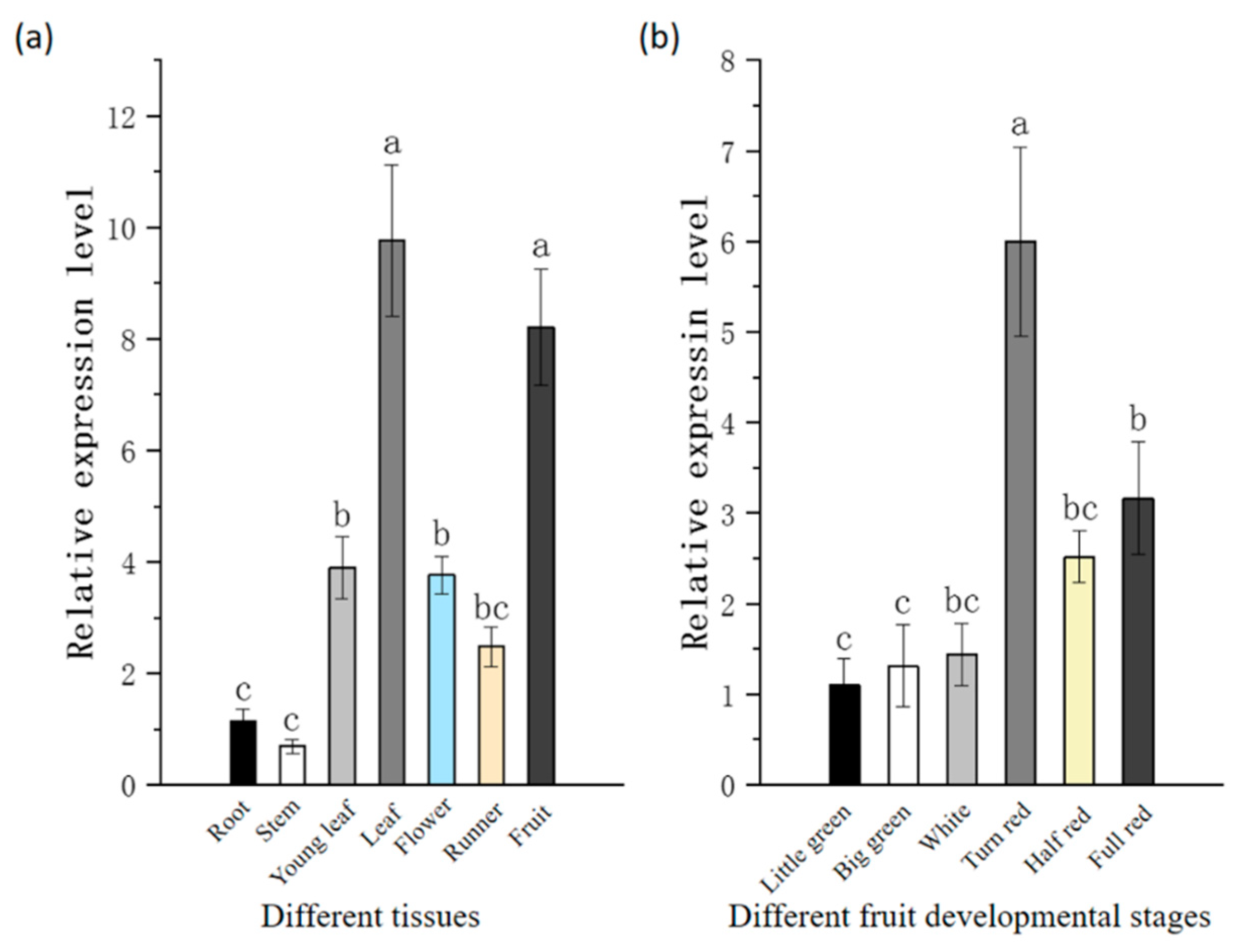
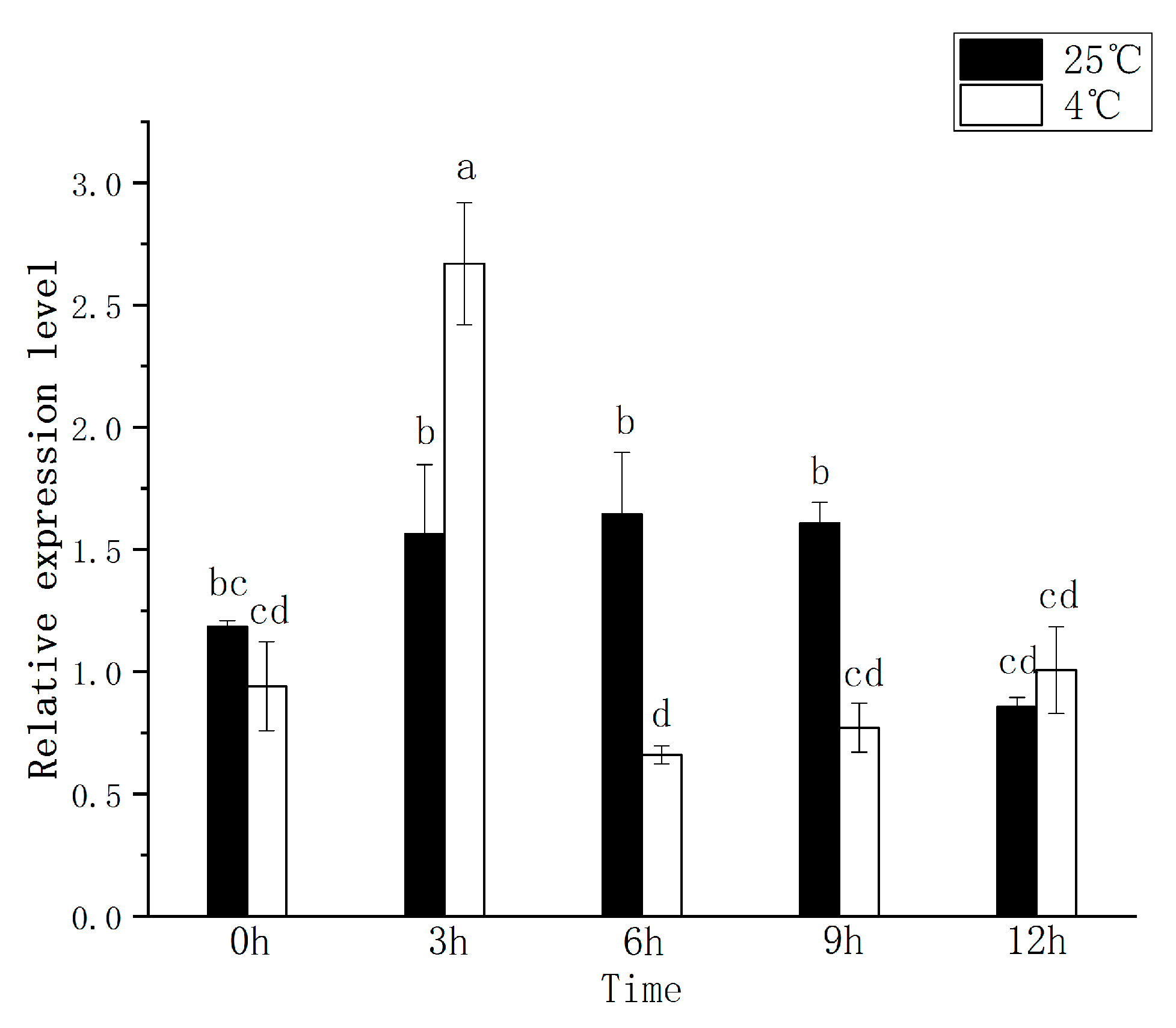
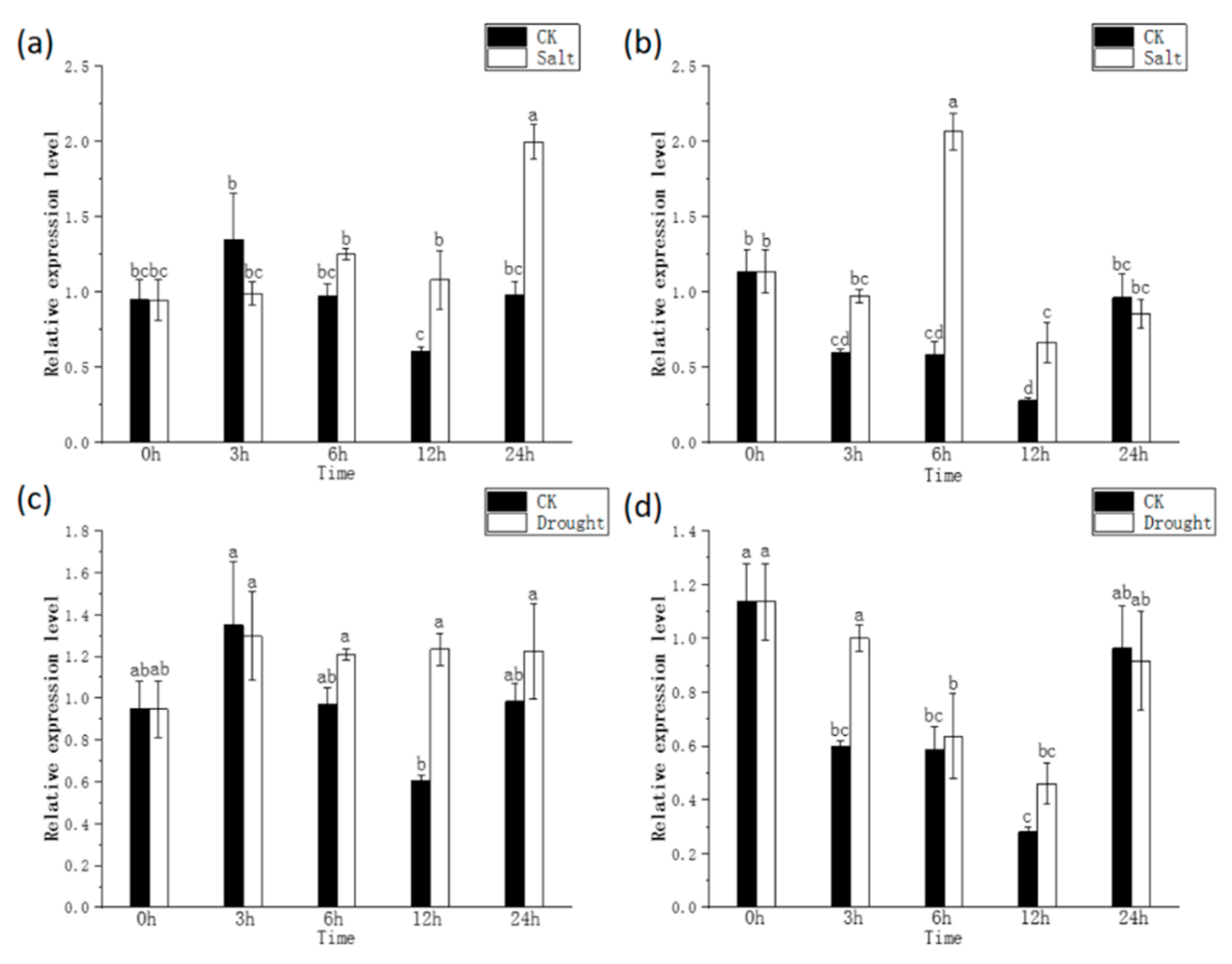
2.3. The Expression Pattern of CRY1 Analyzed with qRT-PCR
2.4. Subcellular Localization Analysis
2.5. Yeast Two-Hybrid Screening Assay
2.6. Agrobacterium-Mediated Heterologous Expression of the FaCRY1 in Tobacco
3. Results
3.1. Gene Cloning, Protein Property and Phylogenetic Analysis of FaCRY1
3.2. Cis-Reglatory Elements in the Promoters of FaCRY1 Genes
3.3. The Expression Pattern of CRY1 in Strawberry
3.4. Effects of Abiotic Stress on CRY1 Expression
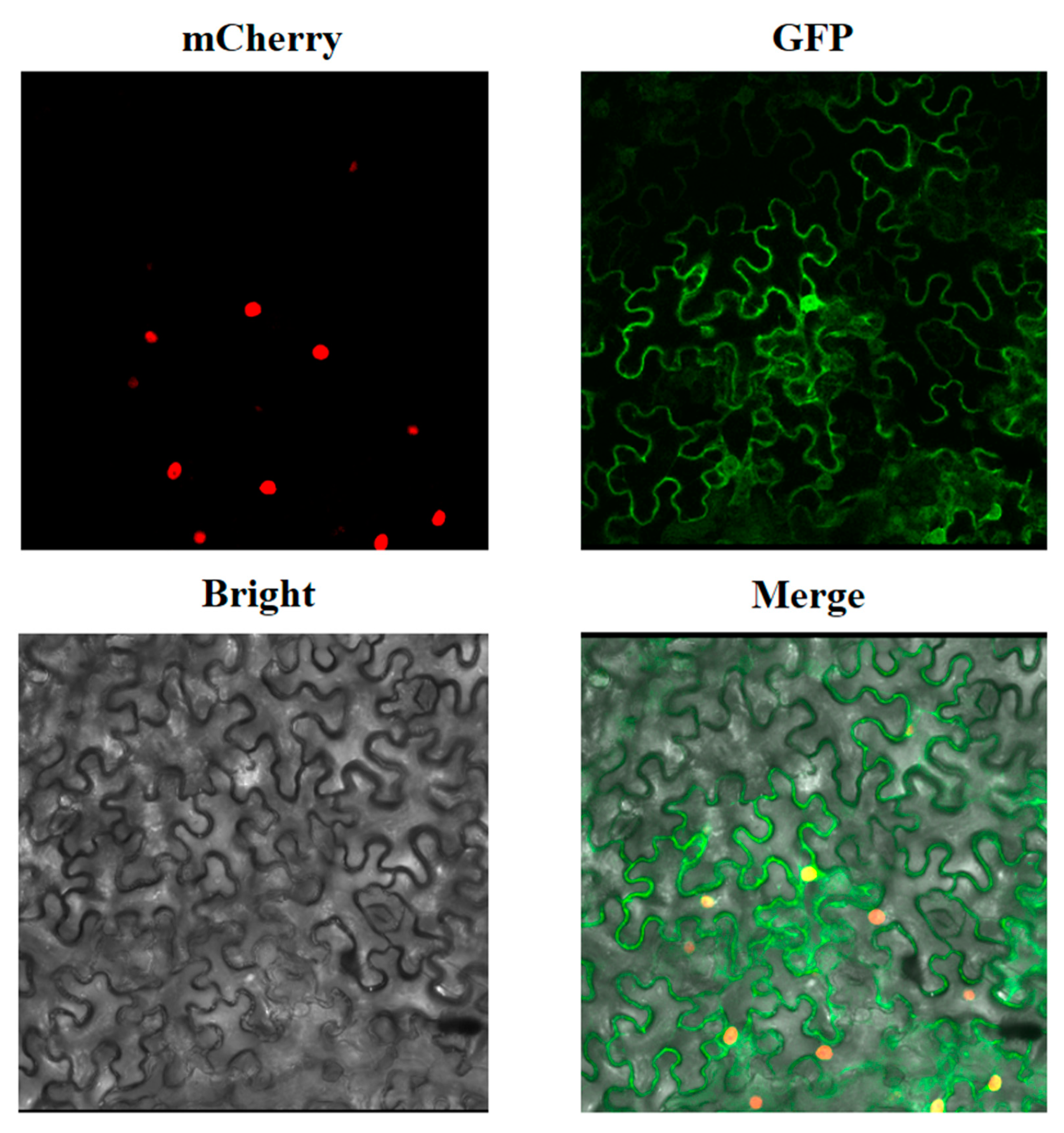
3.5. Subcellular Localization of FaCRY1
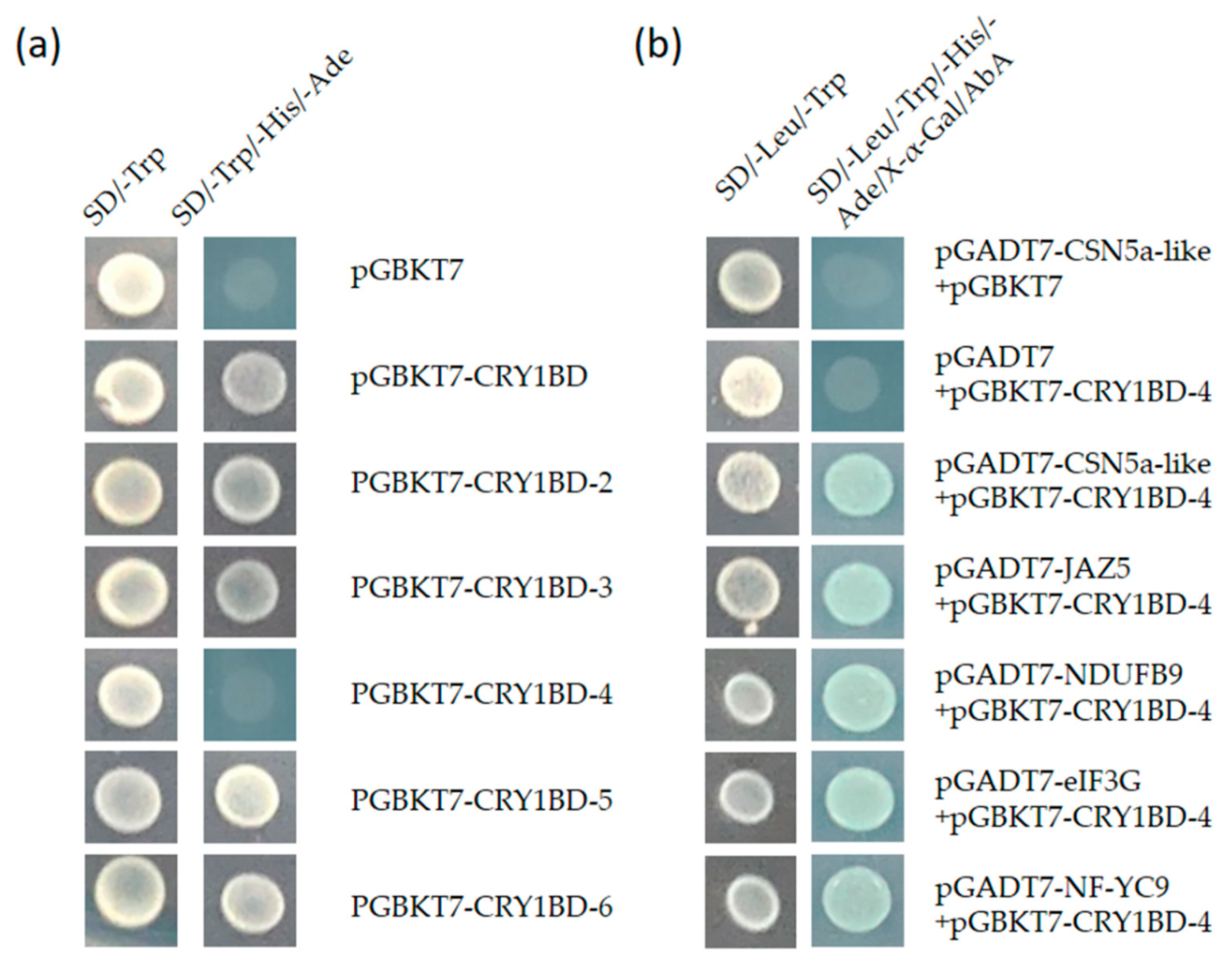
3.6. Yeast Two-Hybrid Library Screening Assay
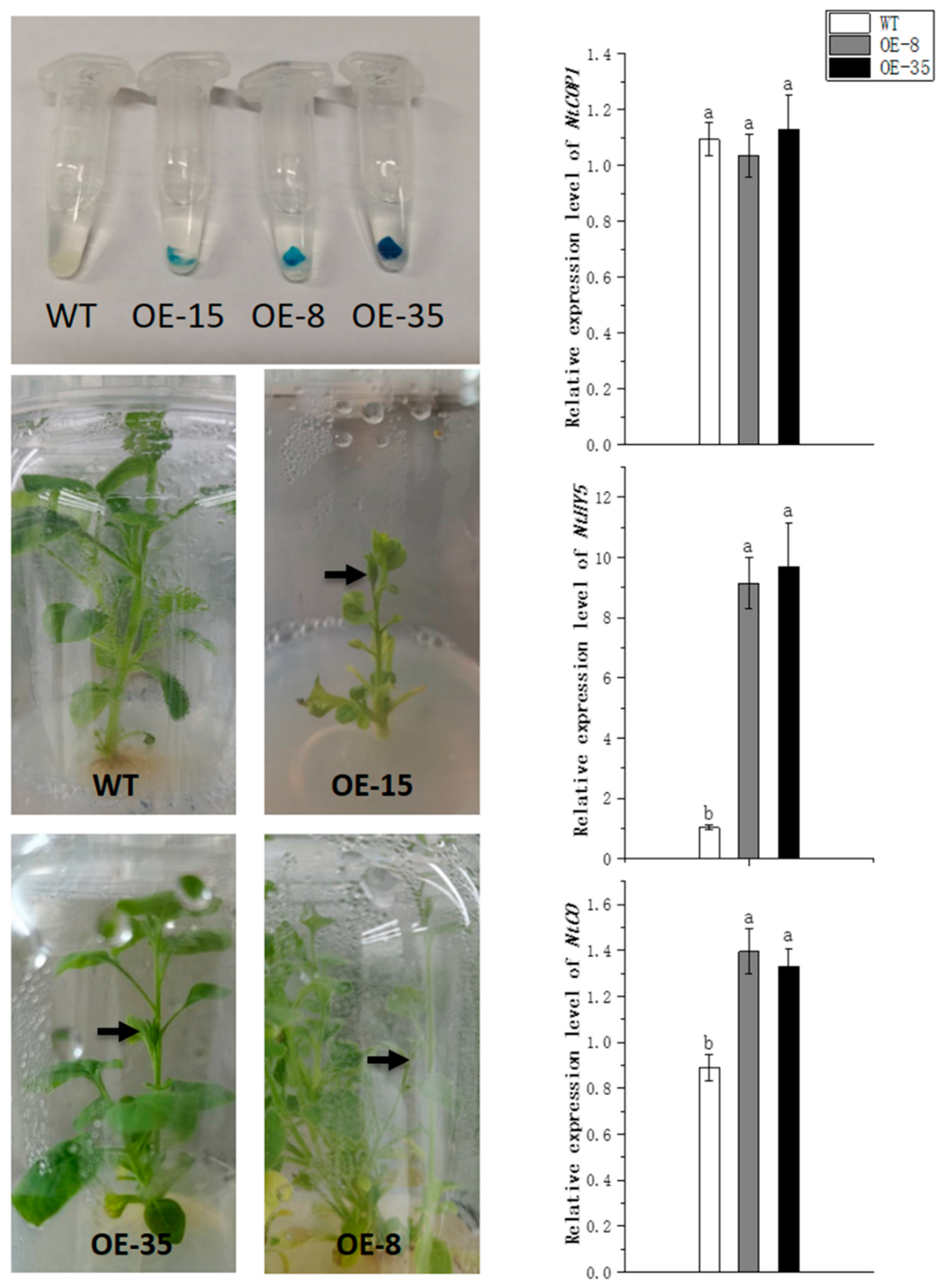
3.7. Heterologous Overexpression of Strawberry CRY1 in Tobacco
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giampieri, F.; Tulipani, S.; Alvarez-Suarez, J.M.; Quiles, J.L.; Mezzetti, B.; Battino, M. The strawberry: Composition, nutritional quality, and impact on human health. Nutrition 2012, 28, 9–19. [Google Scholar] [CrossRef]
- Choi, H.G.; Moon, B.Y.; Kang, N.J. Effects of LED Light on the Production of Strawberry during Cultivation in a Plastic Greenhouse and in a Growth Chamber. Sci. Hortic. 2015, 189, 22–31. [Google Scholar] [CrossRef]
- Hanenberg, M.A.A.; Janse, J.; Verkerke, W. LED light to improve strawberry flavour, quality and production. Acta Hortic. 2016, 1137, 207–212. [Google Scholar] [CrossRef]
- Zheng, J.F.; Ji, F.; He, D.X.; Niu, G.H. Effect of Light Intensity on Rooting and Growth of Hydroponic Strawberry Runner Plants in a LED Plant Factory. Agronomy 2019, 9, 875. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, S.; Chen, Y.; Liu, Y.; Lin, Y.; Li, M.; Wang, Y.; He, W.; Chen, Q.; Zhang, Y.; et al. Heterologous overexpression of strawberry bZIP11 induces sugar accumulation and inhibits plant growth of tomato. Sci. Hortic. 2022, 292, 110634. [Google Scholar] [CrossRef]
- Kolle, S.R.; Shankarappa, T.H.; Rahimi, B.A.; Satish, M.V. Review of Trends in Strawberry Research from 1960 to 2016. J. Agric. Food Inf. 2019, 20, 25–38. [Google Scholar] [CrossRef]
- Ye, Y.; Liu, Y.; Li, X.; Wang, G.; Zhou, Q.; Chen, Q.; Li, J.; Wang, X.; Tang, H. An Evolutionary Analysis of B-Box Transcription Factors in Strawberry Reveals the Role of FaBBx28c1 in the Regulation of Flowering Time. Int. J. Mol. Sci. 2021, 22, 11766. [Google Scholar] [CrossRef]
- Avestan, S.; Ghasemnezhad, M.; Esfahani, M.; Byrt, C.S. Application of Nano-Silicon Dioxide Improves Salt Stress Tolerance in Strawberry Plants. Agronomy 2019, 9, 246. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 17672. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, L.; Gao, L.; Chen, Z.; Bie, Y.; Zhao, Q.; Zhang, S.; Hu, X.; Liu, Q.; Wang, X.; et al. Differential photoregulation of the nuclear and cytoplasmic CRY1 in Arabidopsis. New Phytol. 2022, 234, 1332–1346. [Google Scholar] [CrossRef]
- Bhatnagar, A.; Singh, S.; Khurana, J.P.; Burman, N. HY5-COP1: The Central Module of Light Signaling Pathway. J. Plant Biochem. Biotechnol. 2020, 29, 590–610. [Google Scholar] [CrossRef]
- Xu, P.B.; Lian, H.L.; Wang, W.X.; Xu, F.; Yang, H.Q. Pivotal Roles of the Phytochrome-Interacting Factors in Cryptochrome Signaling. Mol. Plant 2016, 9, 496–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Z.; Wei, X.; Li, L.; Xu, P.; Zhang, J.; Wang, W.; Guo, T.; Kou, S.; Wang, W.; Miao, L.; et al. Arabidopsis cryptochrome 1 controls photomorphogenesis through regulation of H2A.Z deposition. Plant Cell 2021, 33, 1961–1979. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Zhao, J.; Yang, G.; Xu, P.; Cao, X.; Du, S.; Xu, F.; Jiang, L.; Zhang, S.; Wei, X.; et al. Arabidopsis cryptochrome 1 undergoes COP1 and LRBs-dependent degradation in response to high blue light. New Phytol. 2021, 234, 1347–1362. [Google Scholar] [CrossRef]
- Prerostova, S.; Dobrev, P.I.; Knirsch, V.; Jarosova, J.; Gaudinova, A.; Zupkova, B.; Prasil, I.T.; Janda, T.; Brzobohaty, B.; Skalak, J.; et al. Light Quality and Intensity Modulate Cold Acclimation in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 2736. [Google Scholar] [CrossRef]
- Hamidi, H.; Masoudian, N.; Ebadi, M.; Roudi, B.; Khajehzadeh, M.H. How Nitric Oxide Down-Regulates Cryptochrome 1 from Canola in Blue but Not Red Light, and Reduces Hypocotyl Dwarfism in Lead and Drought Stress. Pol. J. Environ. Stud. 2019, 29, 1161–1167. [Google Scholar] [CrossRef]
- D’Amico-Damiao, V.; Carvalho, R.F. Cryptochrome-Related Abiotic Stress Responses in Plants. Front. Plant Sci. 2018, 9, 1897. [Google Scholar] [CrossRef]
- Hwang, O.J.; Back, K. Suppression of Rice Cryptochrome 1b Decreases Both Melatonin and Expression of Brassinosteroid Biosynthetic Genes Resulting in Salt Tolerance. Molecules 2021, 26, 1075. [Google Scholar] [CrossRef]
- Xu, P.; Xiang, Y.; Zhu, H.; Xu, H.; Zhang, Z.; Zhang, C.; Zhang, L.; Ma, Z. Wheat cryptochromes: Subcellular localization and involvement in photomorphogenesis and osmotic stress responses. Plant Physiol. 2009, 149, 760–774. [Google Scholar] [CrossRef] [Green Version]
- Mao, J.; Zhang, Y.C.; Sang, Y.; Li, Q.H.; Yang, H.Q. A role for Arabidopsis cryptochromes and COP1 in the regulation of stomatal opening. Proc. Natl. Acad. Sci. USA 2005, 102, 12270–12275. [Google Scholar] [CrossRef] [Green Version]
- Pedmale, U.V.; Huang, S.C.; Zander, M.; Cole, B.J.; Hetzel, J.; Ljung, K.; Reis, P.A.B.; Sridevi, P.; Nito, K.; Nery, J.R.; et al. Cryptochromes Interact Directly with PIFs to Control Plant Growth in Limiting Blue Light. Cell 2016, 164, 233–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Lu, X.; Li, L.; Lian, H.; Mao, Z.; Xu, P.; Guo, T.; Xu, F.; Du, S.; Cao, X.; et al. Photoexcited CRYPTOCHROME1 Interacts with Dephosphorylated BES1 to Regulate Brassinosteroid Signaling and Photomorphogenesis in Arabidopsis. Plant Cell 2018, 30, 1989–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Chen, H.; Li, T.; Xu, F.; Mao, Z.; Cao, X.; Miao, L.; Du, S.; Hua, J.; Zhao, J.; et al. Blue light-dependent interactions of CRY1 with GID1 and DELLA proteins regulate gibberellin signaling and photomorphogenesis in Arabidopsis. Plant Cell 2021, 33, 2375–2394. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Zeng, B.; Tang, D.; Yang, J.; Qu, L.; Yan, J.; Wang, X.; Li, X.; Liu, X.; Zhao, X. The blue light receptor CRY1 interacts with GID1 and DELLA proteins to repress GA signaling during photomorphogenesis in Arabidopsis. Mol. Plant 2021, 14, 1328–1342. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Q.; Yu, H.; Zhou, S.; Tang, H. Expression Analysis of the ANS Gene in Fragaria × ananassa cv. Toyonaka. J. Life Sci. 2012, 6, 1185–1189. [Google Scholar]
- Llop-Tous, I.; DomõÂnguez-Puigjaner, E.; Vendrell, M. Characterization of a strawberry cDNA clone homologous to calciumdependent protein kinases that is expressed during fruit ripening and affected by low temperature. J. Exp. Bot. 2002, 53, 2283–2285. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, Y.; Hu, W.; Sun, B.; Chen, Q.; Tang, H. Anthocyanin accumulation and related gene expression affected by low temperature during strawberry coloration. Acta Physiol. Plant. 2018, 40, 192. [Google Scholar] [CrossRef]
- Liang, J.; Zheng, J.; Wu, Z.; Wang, H. Strawberry FaNAC2 Enhances Tolerance to Abiotic Stress by Regulating Proline Metabolism. Plants 2020, 9, 1417. [Google Scholar] [CrossRef]
- Mozafari, A.A.; Ghaderi, N.; Havas, F.; Dedejani, S. Comparative investigation of structural relationships among morphophysiological and biochemical properties of strawberry (Fragaria × ananassa Duch.) under drought and salinity stresses: A study based on in vitro culture. Sci. Hortic. 2019, 256, 108601. [Google Scholar] [CrossRef]
- Mozafari, A.A.; Dedejani, S.; Ghaderi, N. Positive responses of strawberry (Fragaria×ananassa Duch.) explants to salicylic and iron nanoparticle application under salinity conditions. Plant Cell Tissue Organ Cult. 2018, 134, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Tang, H.; Zhang, Y.; Du, Z.; Yu, H.; Chen, Q. Comparison and improvement of different methods of RNA isolation from strawberry (Fragria x ananassa). J. Agric. Sci. 2012, 4, 51. [Google Scholar] [CrossRef]
- Li, Y.; Xu, P.; Chen, G.; Wu, J.; Liu, Z.; Lian, H. FvbHLH9 Functions as a Positive Regulator of Anthocyanin Biosynthesis by Forming a HY5-bHLH9 Transcription Complex in Strawberry Fruits. Plant Cell Physiol. 2020, 61, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Goodin, M.M.; Dietzgen, R.G.; Schichnes, D.; Ruzin, S.; Jackson, A. PGD Vectors: Versatile Tools for the Expression of Green and Red Fluorescent Protein Fusions in Agroinfiltrated Plant Leaves. Plant J. 2002, 31, 375–383. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Zhao, X.; Kong, F.; Zuo, Z.; Liu, X. TCP2 positively regulates HY5/HYH and photomorphogenesis in Arabidopsis. J. Exp. Bot. 2016, 67, 775–785. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Zuo, Z.; Liu, H.; Liu, X.; Lin, C. Arabidopsis cryptochrome 1 interacts with SPA1 to suppress COP1 activity in response to blue light. Genes Dev. 2011, 25, 1029–1034. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.; Chattopadhyaya, B.; Koduru, L.; Das, N.; Maiti, M.K. Bran-specific expression of Brassica juncea microsomal ω-3 desaturase gene (BjFad3) improves the nutritionally desirable ω-6:ω-3 fatty acid ratio in rice bran oil. Plant Cell Tiss. Organ. Cult. 2014, 119, 117–129. [Google Scholar] [CrossRef]
- Zhang, Y.; Ye, Y.; Jiang, L.; Lin, Y.; Gu, X.; Chen, Q.; Sun, B.; Zhang, Y.; Luo, Y.; Wang, Y.; et al. Genome-Wide Characterization of Snf1-Related Protein Kinases (SnRKs) and Expression Analysis of SnRK1.1 in Strawberry. Genes 2020, 11, 427. [Google Scholar] [CrossRef]
- Jin, X.Q.; Chen, Z.W.; Tan, R.H.; Zhao, S.J.; Hu, Z.B. Isolation and functional analysis of 4-coumarate:coenzyme A ligase gene promoters from Salvia miltiorrhiza. Biol. Plant. 2012, 56, 261–268. [Google Scholar] [CrossRef]
- Kaur, A.; Pati, P.K.; Pati, A.M.; Nagpal, A.K. In-silico analysis of cis-acting regulatory elements of pathogenesis-related proteins of Arabidopsis thaliana and Oryza sativa. PLoS ONE 2017, 12, e0184523. [Google Scholar] [CrossRef] [Green Version]
- Kummari, D.; Bhatnagar-Mathur, P.; Sharma, K.K.; Vadez, V.; Palakolanu, S.R. Functional characterization of the promoter of pearl millet heat shock protein 10 (PgHsp10) in response to abiotic stresses in transgenic tobacco plants. Int. J. Biol. Macromol. 2020, 156, 103–110. [Google Scholar] [CrossRef]
- Ponnu, J.; Hoecker, U. Illuminating the COP1/SPA Ubiquitin Ligase: Fresh Insights Into Its Structure and Functions During Plant Photomorphogenesis. Front. Plant Sci. 2021, 12, 662793. [Google Scholar] [CrossRef] [PubMed]
- Marzi, D.; Brunetti, P.; Mele, G.; Napoli, N.; Calo, L.; Spaziani, E.; Matsui, M.; De Panfilis, S.; Costantino, P.; Serino, G.; et al. Light controls stamen elongation via cryptochromes, phytochromes and COP1 through HY5 and HYH. Plant J. Cell Mol. Biol. 2020, 103, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.; Liu, H.; Liu, B.; Liu, X.; Lin, C. Blue light-dependent interaction of CRY2 with SPA1 regulates COP1 activity and floral initiation in Arabidopsis. Curr. Biol. CB 2011, 21, 841–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancar, A. Structure and Function of DNA Photolyase and Cryptochrome Blue-light Photoreceptors. Chem. Rev. 2003, 103, 2203–2237. [Google Scholar] [CrossRef]
- Liscum, E.; Hodgson, D.W.; Campbell, T.J. Blue light signaling through the cryptochromes and phototropins. So that’s what the blues is all about. Plant Physiol. 2003, 133, 1429–1436. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, N.; Hirano, T.; Iwasaki, T.; Yamamoto, N. Functional analysis and intracellular localization of rice cryptochromes. Plant Physiol. 2003, 133, 1494–1503. [Google Scholar] [CrossRef] [Green Version]
- Zeugner, A.; Byrdin, M.; Bouly, J.-P.; Bakrim, N.; Giovani, B.; Brettel, K.; Ahmad, M. Light-induced Electron Transfer in Arabidopsis Cryptochrome-1 Correlates with in Vivo Function. J. Biol. Chem. 2005, 280, 19437–19440. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Chatterjee, M.; Burman, N.; Khurana, J.P. Cryptochrome 1 regulates growth and development in Brassica through alteration in the expression of genes involved in light, phytohormone and stress signalling. Plant Cell Environ. 2014, 37, 961–977. [Google Scholar] [CrossRef]
- Facella, P.; Daddiego, L.; Giuliano, G.; Perrotta, G. Gibberellin and auxin influence the diurnal transcription pattern of photoreceptor genes via CRY1a in tomato. PLoS ONE 2012, 7, e30121. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Liu, H.; Klejnot, J.; Lin, C. The Cryptochrome Blue Light Receptors. Arab. Book 2010, 8, e0135. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.C.; Ahammed, G.J.; Wang, G.T.; Xu, C.J.; Chen, K.S.; Zhou, Y.H.; Yu, J.Q. Tomato CRY1a plays a critical role in the regulation of phytohormone homeostasis, plant development, and carotenoid metabolism in fruits. Plant Cell Environ. 2018, 41, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Fantini, E.; Sulli, M.; Zhang, L.; Aprea, G.; Jimenez-Gomez, J.M.; Bendahmane, A.; Perrotta, G.; Giuliano, G.; Facella, P. Pivotal Roles of Cryptochromes 1a and 2 in Tomato Development and Physiology. Plant Physiol. 2019, 179, 732–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.M.; Wei, X.R.; Li, Z.; Jia, G.X.; Chen, J.R.; Chen, H.X.; Cao, F.X.; Zheng, S.X.; Li, J.H.; Li, Y.F. Molecular cloning of cryptochrome 1 from Liliumxformolongi and the characterization of its photoperiodic flowering function in Arabidopsis. Plant Sci. Int. J. Exp. Plant Biol. 2022, 316, 111164. [Google Scholar]
- Yuan, S.; Zhang, Z.W.; Zheng, C.; Zhao, Z.Y.; Wang, Y.; Feng, L.Y.; Niu, G.; Wang, C.Q.; Wang, J.H.; Feng, H.; et al. Arabidopsis cryptochrome 1 functions in nitrogen regulation of flowering. Proc. Natl. Acad. Sci. USA 2016, 113, 7661–7666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perin, E.C.; da Silva Messias, R.; Borowski, J.M.; Crizel, R.L.; Schott, I.B.; Carvalho, I.R.; Rombaldi, C.V.; Galli, V. ABA-dependent salt and drought stress improve strawberry fruit quality. Food Chem. 2019, 271, 516–526. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Q.; Han, Y.J.; Liu, Q.; Gu, L.; Yang, Z.; Su, J.; Liu, B.; Zuo, Z.; He, W.; et al. A CRY-BIC negative-feedback circuitry regulating blue light sensitivity of Arabidopsis. Plant J. Cell Mol. Biol. 2017, 92, 426–436. [Google Scholar] [CrossRef]
- Yang, H.Q.; Wu, Y.J.; Tang, R.H.; Liu, D.M.; Liu, Y.; Cashmore, A.R. The C termini of Arabidopsis cryptochromes mediate a constitutive light response. Cell 2000, 103, 815–827. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Klejnot, J.; Zhao, X.; Shalitin, D.; Maymon, M.; Yang, H.; Lee, J.; Liu, X.; Lopez, J.; Lin, C. Arabidopsis cryptochrome 2 completes its posttranslational life cycle in the nucleus. Plant Cell 2007, 19, 3146–3156. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Du, L.; Li, Q.; Kang, J.; Guo, Q.; Wang, S. AtCRY2 Negatively Regulates the Functions of AtANN2 and AtANN3 in Drought Tolerance by Affecting Their Subcellular Localization and Transmembrane Ca2+ Flow. Front. Plant Sci. 2021, 12, 754567. [Google Scholar] [CrossRef]
- Gao, S.W.; Yang, Y.Y.; Wang, C.F.; Guo, J.L.; Zhou, D.G.; Wu, Q.B.; Su, Y.C.; Xu, L.P.; Que, Y.X. Transgenic Sugarcane with a cry1Ac Gene Exhibited Better Phenotypic Traits and Enhanced Resistance against Sugarcane Borer. PLoS ONE 2016, 11, e0153929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, N.; Deng, X.W. The Role of the COP/DET/FUS Genes in Light Control of Arabidopsis Seedling Development. Plant Physiol. 1996, 112, 871–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwok, F.S.; Solano, R.; Tsuge, T.; Chamovitz, D.A.; Ecker, J.R.; Matsui, M.; Deng, X.W. Arabidopsis Homologs of a c-Jun Coactivator Are Present Both in Monomeric Form and in the COP9 Complex, and Their Abundance Is Differentially Affected by the Pleiotropic cop/det/fus Mutations. Plant Cell 1998, 10, 1779–1790. [Google Scholar] [CrossRef] [Green Version]
- Gusmaroli, G.; Feng, S.; Deng, X.W. The Arabidopsis CSN5A and CSN5B subunits are present in distinct COP9 signalosome complexes, and mutations in their JAMM domains exhibit differential dominant negative effects on development. Plant Cell 2004, 16, 2984–3001. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Stolz, S.; Chetelat, A.; Reymond, P.; Pagni, M.; Dubugnon, L.; Farmer, E.E. A downstream mediator in the growth repression limb of the jasmonate pathway. Plant Cell 2007, 19, 2470–2483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thines, B.; Katsir, L.; Melotto, M.; Niu, Y.; Mandaokar, A.; Liu, G.; Nomura, K.; He, S.Y.; Howe, G.A.; Browse, J. JAZ repressor proteins are targets of the SCF(COI1) complex during jasmonate signalling. Nature 2007, 448, 661–665. [Google Scholar] [CrossRef]
- Demianski, A.J.; Chung, K.M.; Kunkel, B.N. Analysis of Arabidopsis JAZ gene expression during Pseudomonas syringae pathogenesis. Mol. Plant Pathol. 2012, 13, 46–57. [Google Scholar] [CrossRef]
- Huang, H.; Gao, H.; Liu, B.; Qi, T.; Tong, J.; Xiao, L.; Xie, D.; Song, S. Arabidopsis MYB24 Regulates Jasmonate-Mediated Stamen Development. Front. Plant Sci. 2017, 8, 1525. [Google Scholar] [CrossRef]
- Jiang, Y.; Yu, D. The WRKY57 Transcription Factor Affects the Expression of Jasmonate ZIM-Domain Genes Transcriptionally to Compromise Botrytis cinerea Resistance. Plant Physiol. 2016, 171, 2771–2782. [Google Scholar] [CrossRef] [Green Version]
- Hinnebusch, A.G. eIF3: A versatile scaffold for translation initiation complexes. Trends Biochem. Sci. 2006, 31, 553–562. [Google Scholar] [CrossRef]
- Singh, B.; Chauhan, H.; Khurana, J.P.; Khurana, P.; Singh, P. Evidence for the role of wheat eukaryotic translation initiation factor 3 subunit g (TaeIF3g) in abiotic stress tolerance. Gene 2013, 532, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Tuo, D.; Shen, W.; Yan, P.; Li, X.; Zhou, P. NIa-Pro of Papaya ringspot virus interacts with Carica papaya eukaryotic translation initiation factor 3 subunit G (CpeIF3G). Virus Genes 2015, 50, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Thie´beauld, O.; Schepetilnikov, M.; Park, H.; Geldreich, A.l.; Kobayashi, K.; Keller, M.; Hohn, T.; Ryabova, L.A. A new plant protein interacts with eIF3 and 60S to enhance virus-activated translation re-initiation. EMBO J. 2009, 28, 3171–3184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Hu, Y.; Jiang, L.; Rui, P.; Zhao, Q.; Feng, J.; Zuo, D.; Zhou, X.; Jiang, T. Strawberry Vein Banding Virus P6 Protein Is a Translation Trans-Activator and Its Activity Can be Suppressed by FveIF3g. Viruses 2018, 10, 717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumimoto, R.W.; Zhang, Y.; Siefers, N.; Holt, B.F., III. NF-YC3, NF-YC4 and NF-YC9 are required for CONSTANS-mediated, photoperiod-dependent flowering in Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2010, 63, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.; Susila, H.; Nasim, Z.; Jung, J.Y.; Ahn, J.H. Arabidopsis ABF3 and ABF4 Transcription Factors Act with the NF-YC Complex to Regulate SOC1 Expression and Mediate Drought-Accelerated Flowering. Mol. Plant 2019, 12, 489–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, C.; Ma, Y.; Wang, X.F.; Zhang, D.P. Overexpression of the transcription factor NF-YC9 confers abscisic acid hypersensitivity in Arabidopsis. Plant Mol. Biol. 2017, 95, 425–439. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yang, Y.; Hu, Y.; Zhou, L.; Li, Y.; Hou, X. Temporal-Specific Interaction of NF-YC and CURLY LEAF during the Floral Transition Regulates Flowering. Plant Physiol. 2018, 177, 105–114. [Google Scholar] [CrossRef]
- Giliberto, L.; Perrotta, G.; Pallara, P.; Weller, J.L.; Fraser, P.D.; Bramley, P.M.; Fiore, A.; Tavazza, M.; Giuliano, G. Manipulation of the blue light photoreceptor cryptochrome 2 in tomato affects vegetative development, flowering time, and fruit antioxidant content. Plant Physiol. 2005, 137, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.C.; Chi, C.; Jin, L.J.; Zhu, J.; Yu, J.Q.; Zhou, Y.H. The bZip transcription factor HY5 mediates CRY1a-induced anthocyanin biosynthesis in tomato. Plant Cell Environ. 2018, 41, 1762–1775. [Google Scholar] [CrossRef]
- Hou, X.; Zhou, J.; Liu, C.; Liu, L.; Shen, L.; Yu, H. Nuclear factor Y-mediated H3K27me3 demethylation of the SOC1 locus orchestrates flowering responses of Arabidopsis. Nat. Commun. 2014, 5, 4601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Gao, Z.; Wang, Y.; Chen, Z.; Zhang, W.; Huang, J.; Yu, H.; He, Y. The Nuclear Factor-Constans complex antagonizes Polycomb repression to de-repress FLOWERING LOCUS T expression in response to inductive long days in Arabidopsis. Plant J. Cell Mol. Biol. 2018, 95, 17–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classify | Site Name | Numbers | Annotation |
|---|---|---|---|
| Light | Box 4 | 2 | part of a conserved DNA module involved in light responsiveness |
| G-Box | 1 | involved in light responsiveness | |
| G-box | 1 | involved in light responsiveness | |
| GATA-motif | 2 | part of a light responsive element | |
| GT1-motif | 2 | light responsive element | |
| I-box | 1 | part of a light responsive element | |
| MRE | 1 | MYB binding site involved in light responsiveness | |
| TCCC-motif | 1 | part of a light responsive element | |
| TCT-motif | 1 | part of a light responsive element | |
| Hormone | AAGAA-motif | 1 | involved in the abscisic acid responsiveness |
| ABRE | 1 | involved in the abscisic acid responsiveness | |
| AuxRR-core | 1 | involved in auxin responsiveness | |
| GARE-motif | 1 | gibberellin-responsive element | |
| P-box | 2 | gibberellin-responsive element | |
| Stress | STRE | 3 | Osmotic stress response element |
| TC-rich repeats | 1 | involved in defense and stress responsiveness | |
| TCA | 1 | associated with salt stress | |
| W-box | 1 | stress response element | |
| Others | MYB | 4 | N/A |
| MYB-like sequence | 2 | N/A | |
| Myb-binding site | 2 | N/A | |
| CCGTCC motif | 1 | related to meristem-specific activation | |
| CCGTCC-box | 1 | related to meristem-specific activation | |
| MYC | 1 | N/A | |
| Myc | 1 | N/A | |
| ARE | 1 | cis-acting regulatory element essential for the anaerobic induction |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Y.; Li, R.; Pu, W.; Zhang, Y.; Jiang, L.; Li, H.; Liu, Y.; Ye, Y.; Yue, M.; Lin, Y.; et al. Expression Analysis and Interaction Protein Screening of CRY1 in Strawberry. Horticulturae 2022, 8, 460. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8050460
Ye Y, Li R, Pu W, Zhang Y, Jiang L, Li H, Liu Y, Ye Y, Yue M, Lin Y, et al. Expression Analysis and Interaction Protein Screening of CRY1 in Strawberry. Horticulturae. 2022; 8(5):460. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8050460
Chicago/Turabian StyleYe, Yuyun, Ruiling Li, Wenchao Pu, Yunting Zhang, Leiyu Jiang, Hao Li, Yongqiang Liu, Yuntian Ye, Maolan Yue, Yuanxiu Lin, and et al. 2022. "Expression Analysis and Interaction Protein Screening of CRY1 in Strawberry" Horticulturae 8, no. 5: 460. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8050460