
Brassinosteroid Applications Enhance the Tolerance to Abiotic Stresses, Production and Quality of Strawberry Fruits

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Treatments with BRs
2.3. Growth Promotion
2.4. Abiotic Stress
2.5. Fruit Quality Measurements
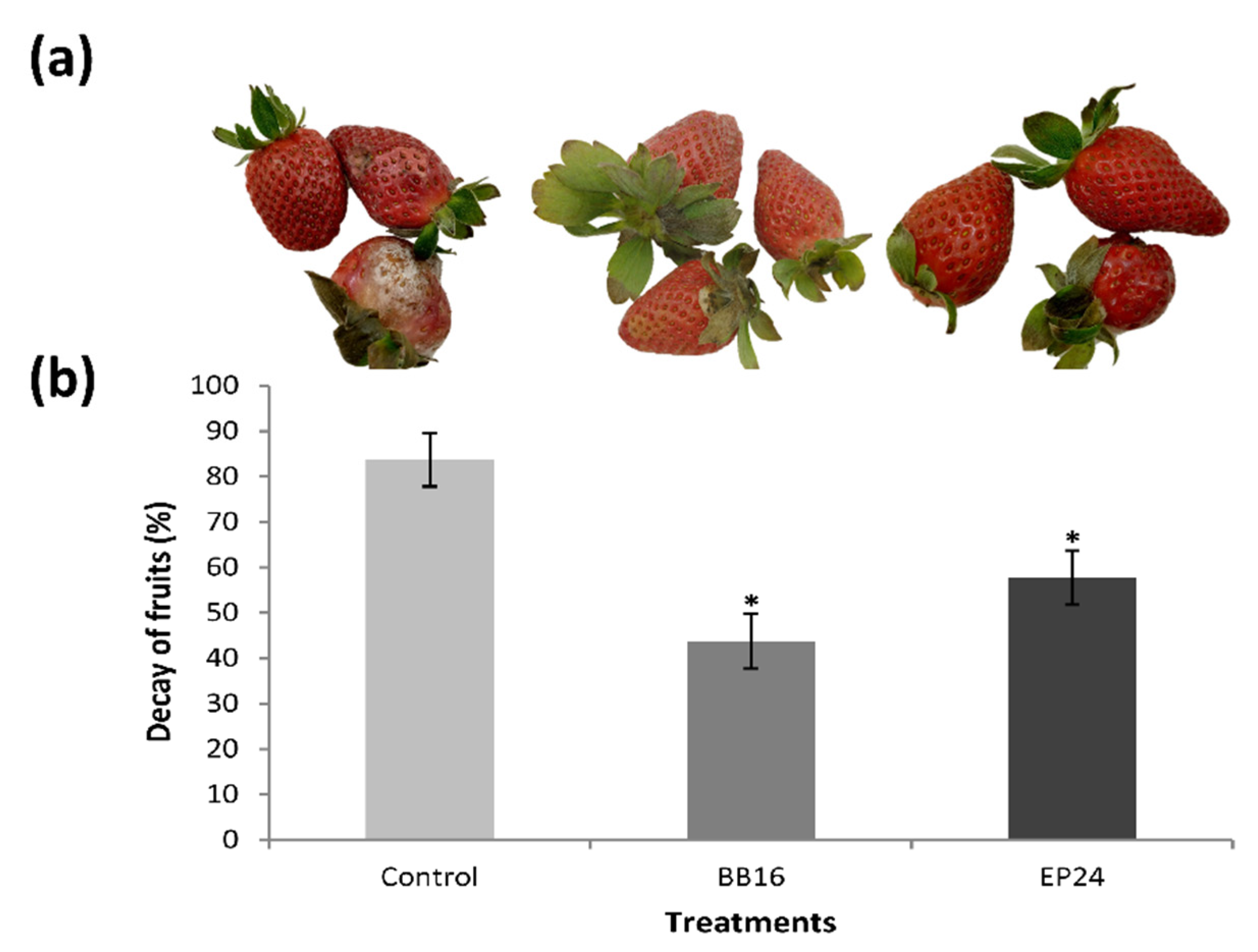
2.6. Incidence of Postharvest Diseases in Fruits
2.7. Statistical Analysis
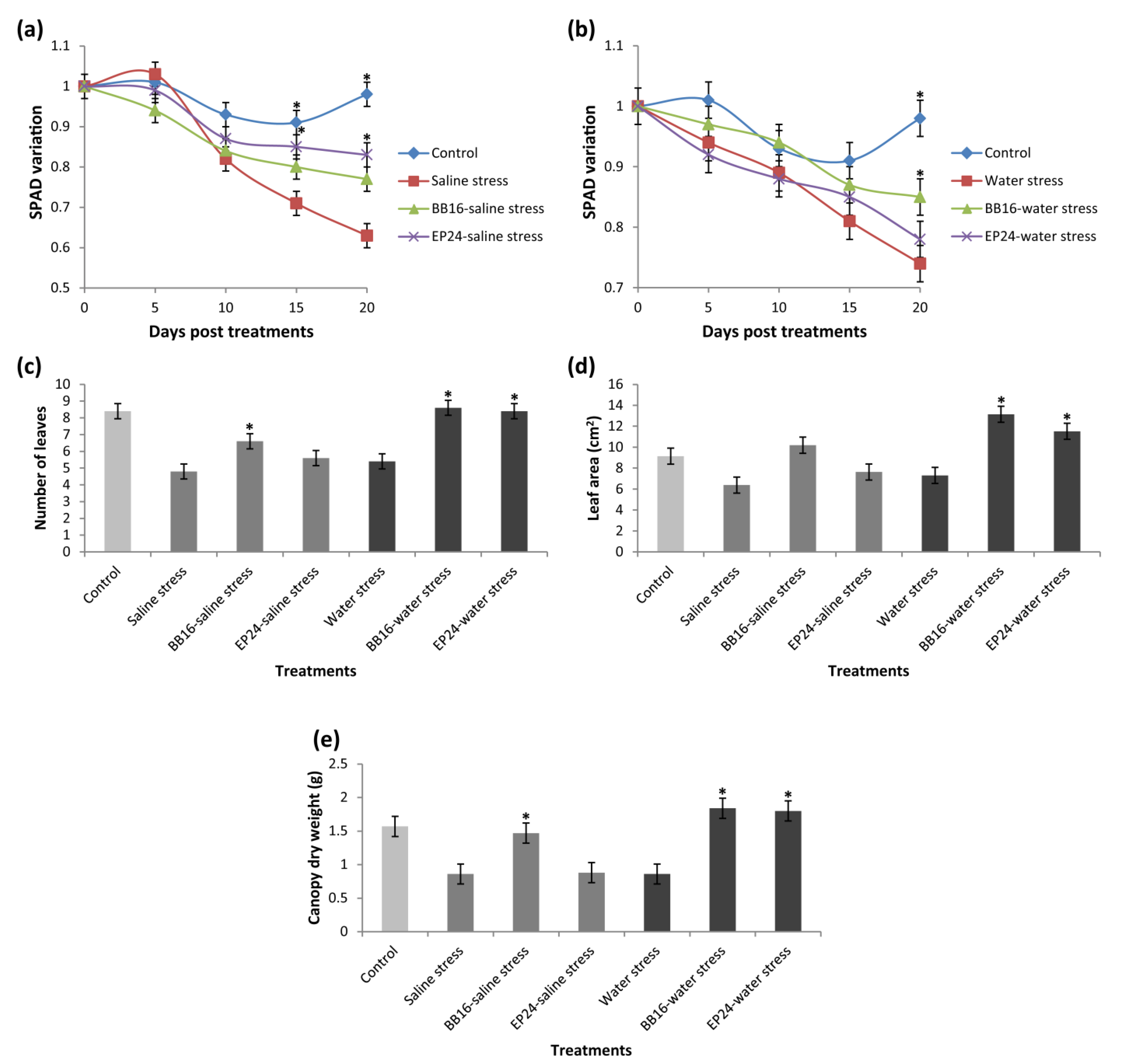
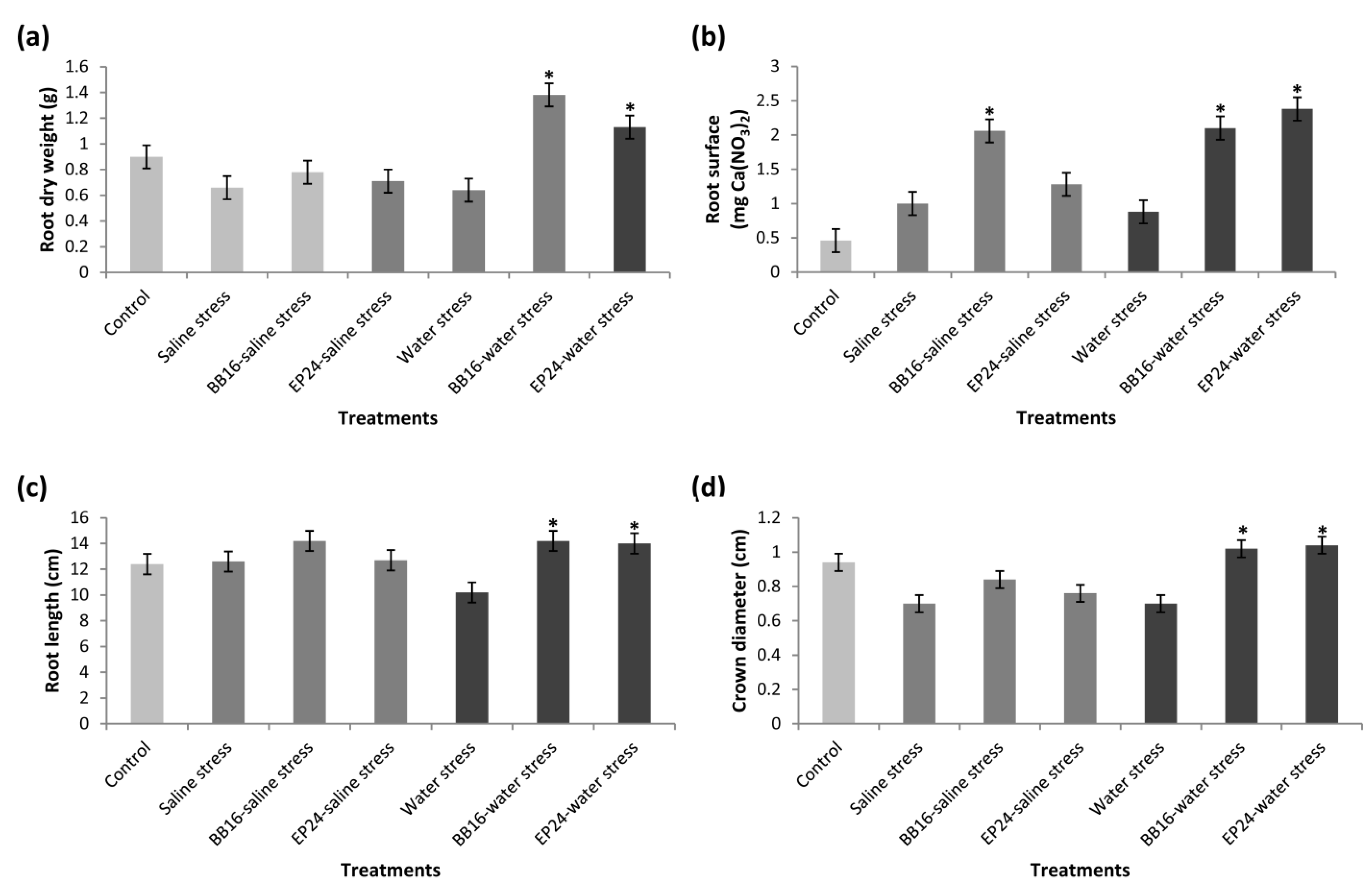
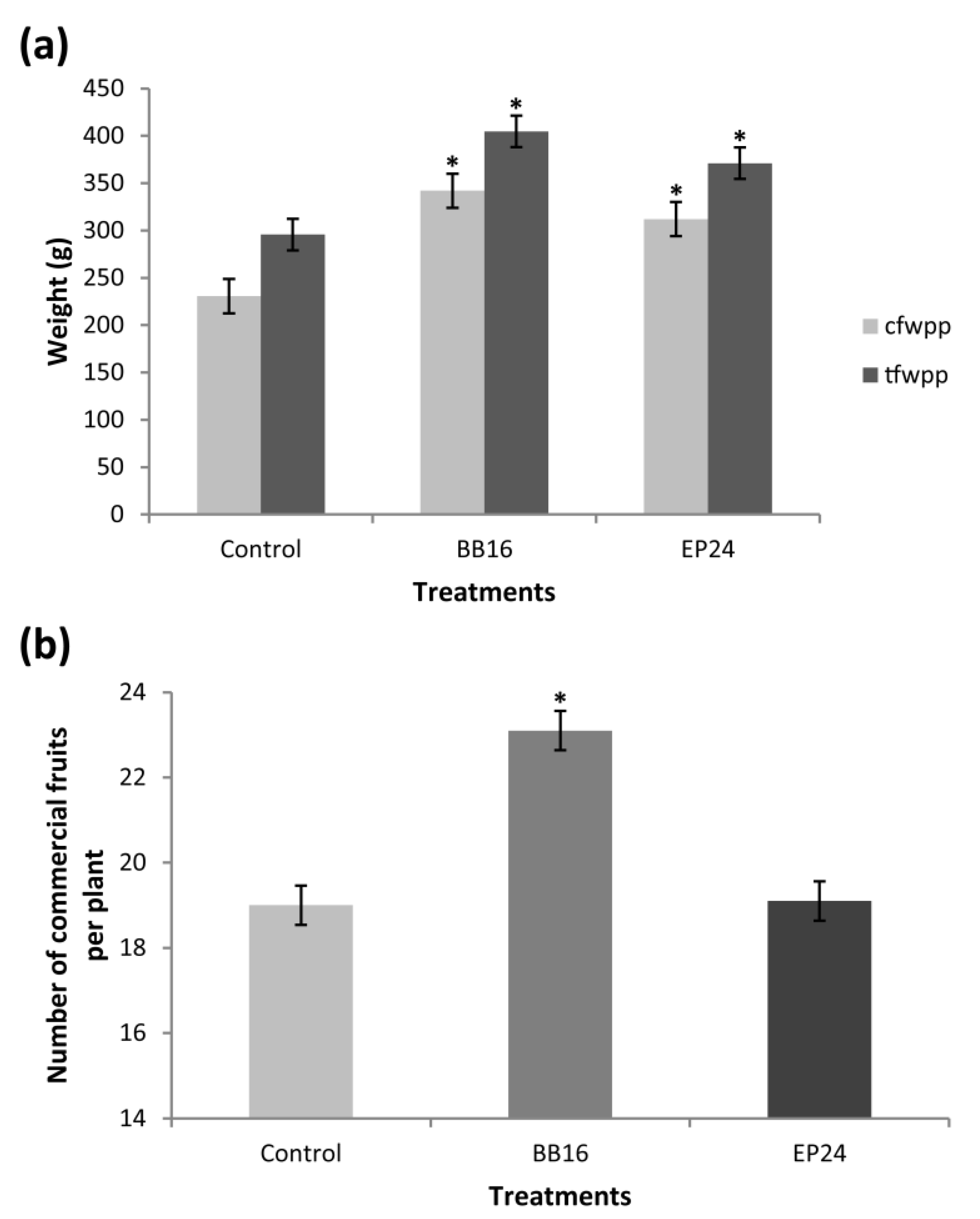
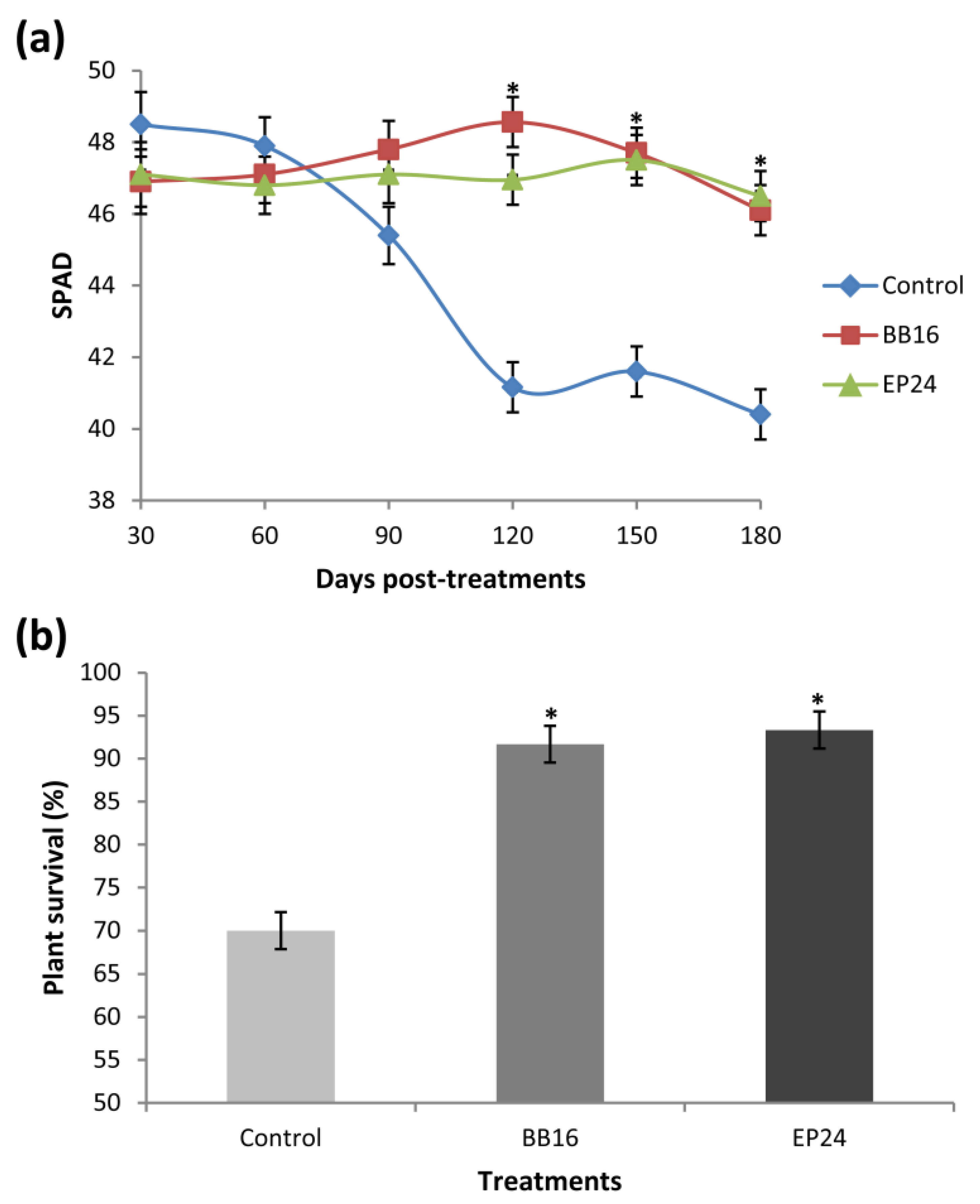
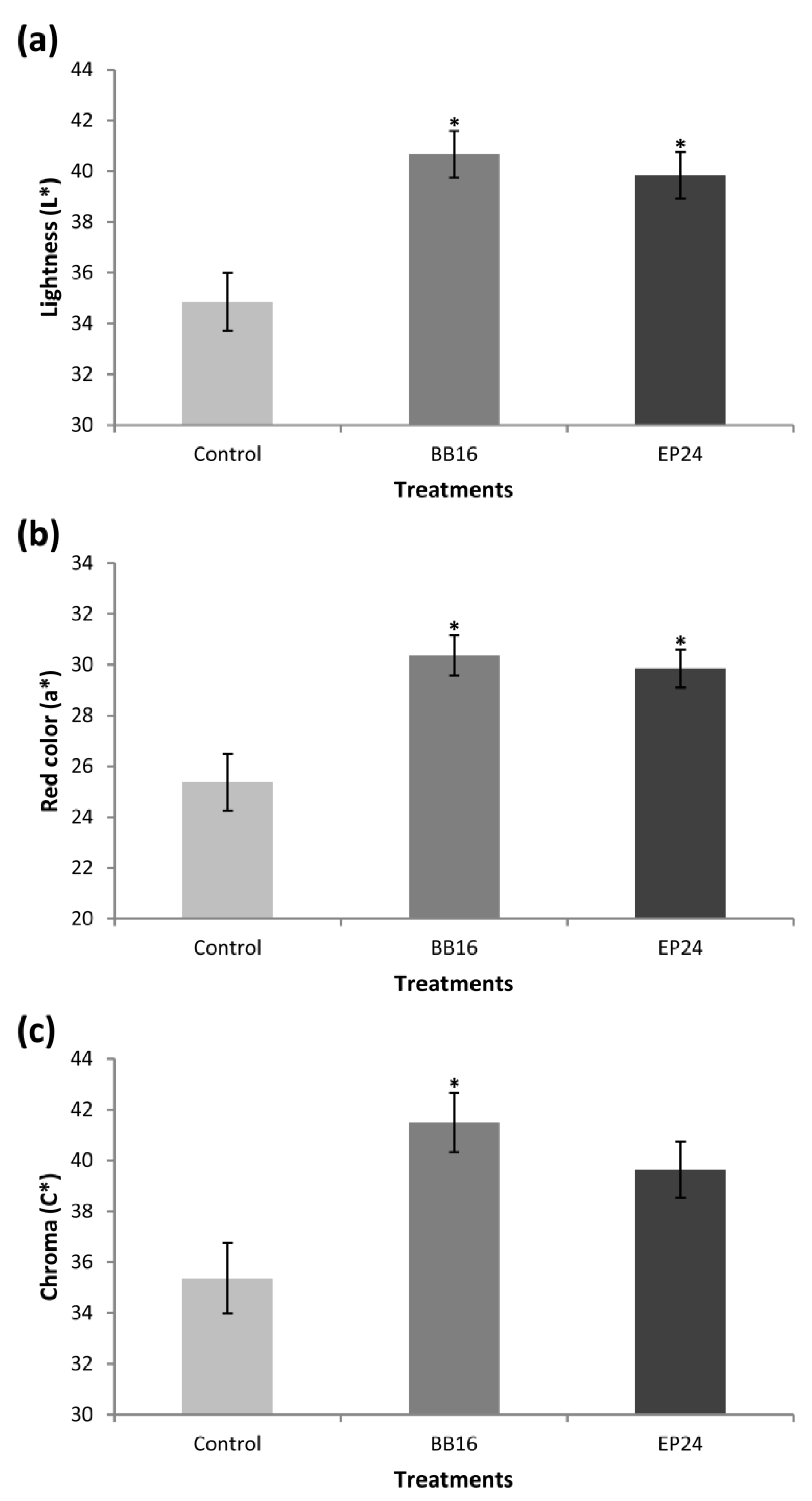
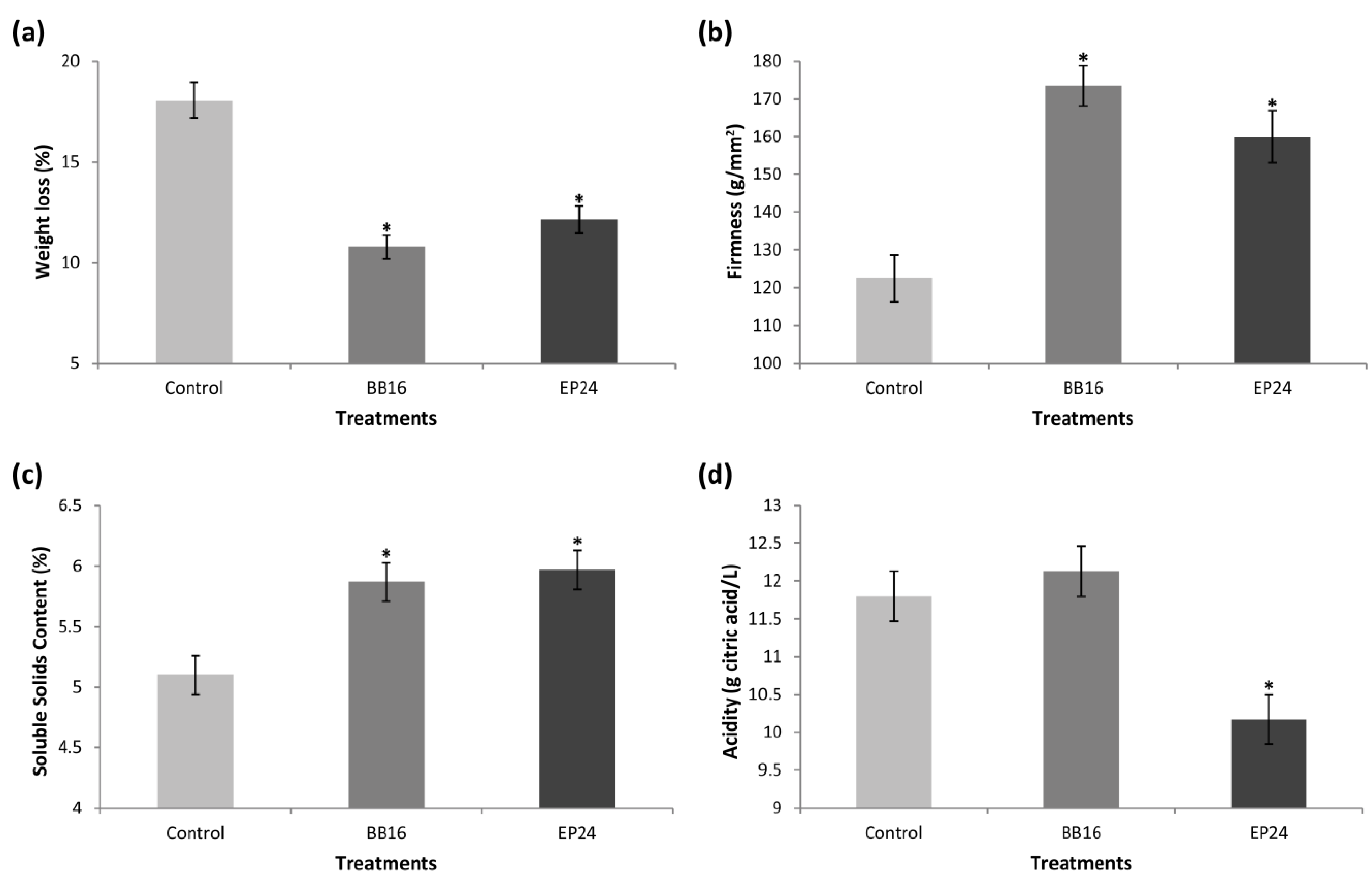
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mitchell, J.W.; Mandava, N.B.; Worley, J.F.; Plimmer, J.R.; Smith, M.V. Brassins: A new family of plant hormones from rape pollen. Nature 1970, 225, 1065–1066. [Google Scholar] [CrossRef] [PubMed]
- Mandava, N.B. Plant growth-promoting brassinosteroids. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1988, 39, 23–52. [Google Scholar] [CrossRef]
- Clouse, S.D. Brassinosteroid signal transduction: From receptor kinase activation to transcriptional networks regulating plant development. Plant Cell 2011, 23, 1219–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clouse, S.D.; Sasse, J.M. Brassinosteroids: Essential regulators of plant growth and development. Annu. Rev. Plant Biol. 1998, 49, 427–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Núñez, M.; Torres, W.; Coll, F. Effectiveness of a synthetic brassinosteroid on potato and tomato yields. Cultiv. Trop. 1995, 16, 26–27. [Google Scholar]
- Torres, W.; Núñez, M. The application of biobras–6 and its effect on potato (Solanum tuberosum L.) yields. Cultiv. Trop. 1997, 18, 8–10. [Google Scholar]
- Cortes, P.A.; Terrazas, T.; León, T.; Larqué-Saavedra, A. Brassinosteroid effects on the precocity and yield of cladodes of cactus pear (Opuntiaficus-indica (L.) Mill.). Sci. Hortic. 2003, 97, 65–73. [Google Scholar] [CrossRef]
- Gomes, M.M.A.; Compostrini, E.; Rocha, N.; Pio, A.; Massi, T.; Siqueira, L. Brassinosteroid analogue effects on the yield of yellow passion fruit plants (Passiflora edulis f. flavicarpa). Sci. Hortic. 2006, 110, 235–240. [Google Scholar] [CrossRef]
- Wu, C.Y.; Trieu, A.; Radhakrishnan, P.; Kwok, S.F.; Harris, S.; Zhang, K.; Wang, J.L.; Wan, J.; Zhai, H.; Takatsuto, S.; et al. Brassinosteroids regulate grain filling in rice. Plant Cell 2008, 20, 2130–2145. [Google Scholar] [CrossRef] [Green Version]
- Stepien, P.; Klobus, G. Water relations and photosynthesis in Cucumis sativus L. leaves under salt stress. Biol. Plant. 2006, 50, 610–616. [Google Scholar] [CrossRef]
- Türkan, I.; Demiral, T. Recent developments in understanding salinity tolerance. Environ. Exp. Bot. 2009, 67, 2–9. [Google Scholar] [CrossRef]
- Rady, M.M. Effect of 24-epibrassinolide on growth, yield, antioxidant system and cadmium content of bean (Phaseolus vulgaris L.) plants under salinity and cadmium stress. Sci. Hortic. 2011, 129, 232–237. [Google Scholar] [CrossRef]
- Chandler, C.K.; Ferree, D.C. Response of ‘Raritan’ and ‘Surecrop’ strawberry plants to drought stress. Fruit Var. J. 1990, 44, 183–184. [Google Scholar]
- Klamkowski, K.; Treder, W. Morphological and physiological responses of strawberry plants to water stress. Agric. Conspec. Sci. 2006, 71, 159–165. [Google Scholar]
- Manghwar, H.; Hussain, A.; Ali, Q.; Liu, F. Brassinosteroids (BRs) Role in Plant Development and Coping with Different Stresses. Int. J. Mol. Sci. 2022, 23, 1012. [Google Scholar] [CrossRef]
- Serna, M.; Coll, Y.; Zapata, P.J.; Botella, M.Á.; Pretel, M.T.; Amorós, A. A brassinosteroid analogue prevented the effect of salt stress on ethylene synthesis and polyamines in lettuce plants. Sci. Hortic. 2015, 185, 105–112. [Google Scholar] [CrossRef]
- Núñez-Vázquez, M.; Pérez-Domínguez, G.; Martínez-González, L.; Reyes-Guerrero, Y.; Coll-García, Y. Spirostanic analogues of brassinosteroids enhance the rice (Oryza sativa L.) cv. INCA LP-7 seedling growth under NaCl stress. Cultiv. Trop. 2016, 37, 152–159. [Google Scholar]
- Yuan, G.F.; Jia, C.G.; Li, Z.; Sun, B.; Zhang, L.P.; Li, N.; Wang, Q.M. Effect of brassinosteroids on drought resistance and abscisic acid concentration in tomato underwater stress. Sci. Hortic. 2010, 126, 103–108. [Google Scholar] [CrossRef]
- Furio, R.N.; Albornoz, P.L.; Coll, Y.; Martinez Zamora, G.M.; Salazar, S.M.; Martos, G.G.; Díaz Ricci, J.C. Effect of natural and synthetic Brassinosteroids on strawberry immune response against Colletotrichum acutatum. Eur. J. Plant Pathol. 2019, 153, 167–181. [Google Scholar] [CrossRef]
- Furio, R.N.; Salazar, S.M.; Martínez-Zamora, G.M.; Coll, Y.; Hael-Conrad, V.; Díaz-Ricci, J.C. Brassinosteroids promote growth, fruit quality and protection against Botrytis on Fragaria x ananassa. Eur. J. Plant Pathol. 2019, 154, 801–810. [Google Scholar] [CrossRef]
- de Carvalho, R.O.; Machado, M.B.; Göebel, J.T.S.; Lang, G.H.; da Luz, M.L.G.S.; Gadotti, G.I.; Silveira da Luz, C.A.; Gomes, M.C. Economical Feasibility of Strawberry Production in a Semi-Hydroponic System and Agroindustry of Jelly on a Small Property . Agric. Eng. Int. CIGR J. 2015, Special Issue, pp. 173–176. Available online: http://library.tuit.uz/knigiPDF/Ebsco/8-112.pdf (accessed on 14 March 2021).
- Treftz, C.; Omaye, S.T. Comparision between hydroponic and soil systems for growing strawberries in a greenhouse. Int. J. Agric. Ext. 2016, 3, 195–200. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Carley, H.E.; Watson, T.W. A new gravimetric method for estimating root-surface areas. Soil Sci. 1966, 102, 289–291. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Montenegro-Gonzalez, H.; Malagón-Castro, D.; Guerrero, I. Propiedades Físicas de Los Suelos; CO-BAC: Bogotá, Colombia, 1990. [Google Scholar]
- Barnes, R.B.; Richardson, D.; Berry, J.W.; Hood, R.L. Flame photometry a rapid analytical procedure. Ind. Eng. Chem. Anal. Ed. 1945, 17, 605–611. [Google Scholar] [CrossRef]
- Kafkafi, U.; Xu, G.; Imas, P.; Magen, H.; Tarchitzky, J. Potassium and Chloride in Crops and Soils: The Role of potassium Chloride Fertilizer in Crop Nutrition; International Potash Institute: Basel, Switzerland, 2001. [Google Scholar]
- Sapers, G.M. Color characteristics and stability of nonbleeding cocktail cherries dyed with carotenoid pigments. J. Food Sci. 1994, 59, 135–138. [Google Scholar] [CrossRef]
- Agüero, J.J.; Salazar, S.M.; Kirschbaum, D.S.; Jerez, E.F. Factors affecting fruit quality in strawberries grown in a subtropical environment. Int. J. Fruit Sci. 2015, 15, 223–234. [Google Scholar] [CrossRef]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Version 2013. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Available online: http://www.infostat.com.ar (accessed on 8 November 2013).
- Salazar, S.M.; Coll, Y.; Viejobueno, J.; Coll, F. Response of Strawberry Plants to the Application of Brassinosteroid Under Field Conditions. Rev. Agronómica Del Noroeste Argent. 2016, 36, 37–41. Available online: https://ranar.faz.unt.edu.ar/index.php/ranar/article/view/68 (accessed on 13 July 2021).
- Xia, X.J.; Huang, L.F.; Zhou, Y.H.; Mao, W.H.; Shi, K.; Wu, J.X.; Asami, T.; Chen, Z.; Yu, J.Q. Brassinosteroids promote photosynthesis and growth by enhancing activation of Rubisco and expression of photosynthetic genes in Cucumis sativus. Planta 2009, 230, 1185. [Google Scholar] [CrossRef]
- Oswald, O.; Martin, T.; Dominy, P.J.; Graham, I.A. Plastid redox state and sugars: Interactive regulators of nuclear-encoded photosynthetic gene expression. Proc. Natl. Acad. Sci. USA 2001, 98, 2047–2052. [Google Scholar] [CrossRef]
- Pfannschmidt, T.; Allen, J.F.; Oelmüller, R. Principles of redox control in photosynthesis gene expression. Physiol. Plant. 2001, 112, 1–9. [Google Scholar] [CrossRef]
- Zhang, N.; Kallis, R.P.; Ewy, R.G.; Portis, A.R. Light modulation of Rubisco in Arabidopsis requires a capacity for redox regulation of the larger Rubisco activase isoform. Proc. Natl. Acad. Sci. USA 2002, 99, 3330–3334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, S.A.; Hayat, S.; Ahmad, A. Brassinosteroids protect photosynthetic machinery against the cadmium induced oxidative stress in two tomato cultivars. Chemosphere 2011, 84, 1446–1451. [Google Scholar] [CrossRef]
- Neill, S.; Desikan, R.; Hancock, J. Hydrogen peroxide signalling. Curr. Opin. Plant Biol. 2002, 5, 388–395. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.J.; Wang, Y.J.; Zhou, Y.H.; Tao, Y.; Mao, W.H.; Shi, K.; Asami, T.; Chen, Z.; Yu, J.Q. Reactive oxygen species are involved in brassinosteroid-induced stress tolerance in cucumber. Plant Physiol. 2009, 150, 801–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokota, T.; Sato, T.; Takeuchi, Y.; Nomura, T.; Uno, K.; Watanabe, T.; Takatsuto, S. Roots and shoots of tomato produce 6-deoxo-28-norcathasterone, 6-deoxo-28-nortyphasterol and 6-deoxo-28-norcastasterone, possible precursors of 28-norcastasterone. Phytochemistry 2001, 58, 233–238. [Google Scholar] [CrossRef]
- Chaiwanon, J.; Wang, Z.Y. Spatiotemporal brassinosteroid signaling and antagonism with auxin pattern stem cell dynamics in Arabidopsis roots. Curr. Biol. 2015, 25, 1031–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-García, M.P.; Vilarrasa-Blasi, J.; Zhiponova, M.; Divol, F.; Mora-García, S.; Russinova, E.; Caño-Delgado, A.I. Brassinosteroids control meristem size by promoting cell cycle progression in Arabidopsis roots. Development 2011, 138, 849–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mussig, C.; Fischer, S.; Altmann, T. Brassinosteroid-regulated gene expression. Plant Physiol. 2002, 129, 1241–1251. [Google Scholar] [CrossRef] [Green Version]
- Grant, O.M.; Johnson, A.W.; Davies, M.J.; James, C.M.; Simpson, D.W. Physiological and morphological diversity of cultivated strawberry (Fragaria × ananassa) in response to water deficit. Environ. Exp. Bot. 2010, 68, 264–272. [Google Scholar] [CrossRef]
- Arkin, G.F.; Taylor, H.M. Modifying the Root Environment to Reduce Crop Stress; American Society of Agricultural Engineers: St. Joseph, MI, USA, 1981; pp. 305–343. [Google Scholar]
- Saied, A.S.; Keutgen, A.J.; Noga, G. The influence of NaCl salinity on growth, yield and fruit quality of strawberry cvs ‘Elsanta’ and ‘Korona’. Sci. Hortic. 2005, 103, 289–303. [Google Scholar] [CrossRef]
- Arreola, J.; Martínez-Sánchez, J.J.; Conesa, E.; Franco, J.A. Effect of pre-conditioning water regimes during nursery production on seedling root system characteristics of Silene vulgaris. Acta Hortic. 2008, 782, 287–292. [Google Scholar] [CrossRef]
- Franco, J.A.; Arreola, J.; Vicente, M.J.; Martínez-Sánchez, J.J. Nursery irrigation regimes affect the seedling characteristics of Silene vulgaris as they relate to potential performance following transplanting into semi-arid conditions. J. Hortic. Sci. Biotechnol. 2008, 83, 15–22. [Google Scholar] [CrossRef]
- Teale, W.D.; Ditengou, F.A.; Dovzhenko, A.D.; Li, X.; Molendijk, A.M.; Ruperti, B.; Paponov, I.; Palme, K. Auxin as a model for the integration of hormonal signal processing and transduction. Mol. Plant. 2008, 1, 229–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirlak, L.; Esitken, A. Salinity effects on growth, proline and ion accumulation in strawberry plants. Acta Agric. Scand.-B Soil Plant Sci. 2004, 54, 189–192. [Google Scholar] [CrossRef]
- Maas, E.V.; Hoffman, G.J. Crop salt tolerance–current assessment. J. Irrig. Drain. Div. 1977, 103, 115–134. [Google Scholar] [CrossRef]
- Bould, C.; Hewitt, E.J.; Needham, P. The occurence and treatment of mineral disorders in the field. Diagn. Miner. Disord. Plants 1983, 1, 139–155. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Plant response to salt stress and role of exogenous protectants to mitigate salt-induced damages. In Ecophysiology and Responses of Plants under Salt Stress; Springer: New York, NY, USA, 2013. [Google Scholar] [CrossRef]
- Makale, P.; Kontturi, M.; Pehu, E.; Somersalo, S. Photosynthetic response of drought and salt-stressed tomato and turnip rape plants to foliar applied glycinebetaine. Physiol. Plant. 1999, 105, 45–50. [Google Scholar] [CrossRef]
- Wise, R.R.; Naylor, A.W. Chilling-enhanced photooxidation: Evidence for the role of singlet oxygen and Superoxide in the Breakdown of Pigments and endogenous antioxidant. Plant Physiol. 1987, 83, 278–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zapata, P.J.; Serrano, M.; Petrel, M.T.; Botella, M.A. Changes in free polyamine concentration induced by salt stress in seedling of different species. Plant Growth Regul. 2008, 56, 167–177. [Google Scholar] [CrossRef]
- Lovisolo, C.; Perrone, I.; Carra, A.; Ferrandino, A.; Flexas, J.; Medrano, H.; Schubert, A. Drought-induced changes in development and function of grapevine (Vitis spp.) organs and in their hydraulic and non-hydraulic interactions at the whole-plant level: A physiological and molecular update. Funct. Plant Biol. 2010, 37, 98–116. [Google Scholar] [CrossRef]
- Meggio, F.; Prinsi, B.; Negri, A.S.; Simone Di Lorenzo, G.; Lucchini, G.; Pitacco, A.; Espen, L. Biochemical and physiological responses of two grapevine rootstock genotypes to drought and salt treatments. Aust. J. Grape Wine Res. 2014, 20, 310–323. [Google Scholar] [CrossRef]
- Zeng, H.; Tang, Q.; Hua, X. Arabidopsis brassinosteroid mutants det2-1 and bin2-1 display altered salt tolerance. J. Plant Growth Regul. 2010, 29, 44–52. [Google Scholar] [CrossRef]
- Shahid, M.A.; Balal, R.M.; Pervez, M.A.; Garcia-Sanchez, F.; Gimeno, V.; Abbas, T.; Mattson, N.S.; Riaz, A. Treatment with 24-epibrassinolide mitigates NaCl-induced toxicity by enhancing carbohydrate metabolism, osmolyte accumulation, and antioxidant activity in Pisum sativum. Turk. J. Bot. 2014, 38, 511–525. [Google Scholar] [CrossRef]
- Nunes, M.C.N.; Emond, J.P.; Rauth, M.; Dea, S.; Chau, K.V. Environmental conditions encountered during typical consumer retail display affect fruit and vegetable quality and waste. Postharvest Biol. Technol. 2009, 51, 232–241. [Google Scholar] [CrossRef]
- Velickova, E.; Winkelhausen, E.; Kuzmanova, S.; Alves, V.D.; Moldão-Martins, M. Impact of chitosan-beeswax edible coatings on the quality of fresh strawberries (Fragaria ananassa cv Camarosa) under commercial storage conditions. LWT 2013, 52, 80–92. [Google Scholar] [CrossRef]
- Liu, L.; Jia, C.; Zhang, M.; Chen, D.; Chen, S.; Guo, R.; Wang, Q. Ectopic expression of a BZR1-1D transcription factor in brassinosteroidsignalling enhances carotenoid accumulation and fruit quality attributes in tomato. Plant Biotechnol. J. 2014, 12, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Roghabadi, M.A.; Pakkish, Z.A.H.R.A. Role of brassinosteroid on yield, fruit quality and postharvest storage of ‘Tak Danehe Mashhad’ sweet cherry (Prunus avium L.). Agric. Commun. 2014, 2, 49–56. [Google Scholar]
- Pakkish, Z.; Ghorbani, B.; Najafzadeh, R. Fruit quality and shelf life improvement of grape cv. Rish Baba using Brassinosteroid during cold storage. J. Food Meas. Charact. 2019, 13, 967–975. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Canopy Parameters | Control | BB16 | EP24 |
|---|---|---|---|
| Fresh weight (g) | 21.9 ± 2.49 a | 29.73 ± 2.49 ab | 36.95 ± 2.49 b |
| Dry weight (g) | 8.93 ± 0.65 a | 12.4 ± 0.65 bc | 14.95 ± 0.65 c |
| Number of leaves | 11.75 ± 0.93 a | 17.5 ± 0.93 bc | 19.75 ± 0.93 c |
| Leaf area (cm2) | 11.63 ± 1.6 a | 28.18 ± 1.6 b | 27.77 ± 1.6 b |
| Number of stolons | 4.98 ± 0.56 a | 6.50 ± 0.56 a | 9.08 ± 0.56 a |
| Weight of total fruits per plant (g) | 49.09 ± 1.56 a | 59.27 ± 1.56 b | 56.67 ± 1.56 b |
| Commercial quality fruit (%) | 18.15 ± 1.57 a | 25.46 ± 1.57 b | 28.15 ± 1.57 b |
| SPAD | 32.92 ±- 0.74 a | 40.34 ± 0.74 b | 40.56 ± 0.74 b |
| Root Parameters | Control | BB16 | EP24 |
|---|---|---|---|
| Fresh weight (g) | 23.5 ± 4.33 a | 48.23 ± 4.33 b | 43.1 ± 4.33 b |
| Dry weight (g) | 4.6 ± 0.58 a | 12.83 ± 0.58 b | 11.88 ± 0.58 b |
| Root lenght (cm) | 29.04 ± 1.26 a | 34.75 ± 1.26 b | 32.25 ± 1.26 ab |
| Root surface (mg Ca(NO3)2) | 3.13 ± 0.72 a | 13.13 ± 0.72 b | 14.1 ± 0.72 b |
| Crown diameter (cm) | 9.38 ± 0.55 a | 12.98 ± 0.55 bc | 11.95 ± 0.55 bc |
| Soil Analysis | |||
|---|---|---|---|
| Control | Saline Stress | Water stress | |
| pH (1:2,5) | 5.78 ± 0.18 b | 5.73 ± 0.18 a | 5.72 ± 0.18 ab |
| CE (dS/m) | 2.66 ± 0.17 a | 35.66 ± 2.35 b | 2.73 ± 0.17 a |
| H2O (%) | 240.67 ± 4.9 c | 101.33 ± 4.9 b | 68.33 ± 4.9 a |
| Na (meq/l) | 17.23 ± 1.05 a | 339.13 ± 10.92 b | 16.26 ± 1.05 a |
| Cl (meq/l) | 17.8 ± 0.48 b | 333.33 ± 15.04 c | 15.77 ± 0.48 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furio, R.N.; Salazar, S.M.; Mariotti-Martínez, J.A.; Martínez-Zamora, G.M.; Coll, Y.; Díaz-Ricci, J.C. Brassinosteroid Applications Enhance the Tolerance to Abiotic Stresses, Production and Quality of Strawberry Fruits. Horticulturae 2022, 8, 572. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8070572
Furio RN, Salazar SM, Mariotti-Martínez JA, Martínez-Zamora GM, Coll Y, Díaz-Ricci JC. Brassinosteroid Applications Enhance the Tolerance to Abiotic Stresses, Production and Quality of Strawberry Fruits. Horticulturae. 2022; 8(7):572. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8070572
Chicago/Turabian StyleFurio, Ramiro N., Sergio M. Salazar, Jorge A. Mariotti-Martínez, Gustavo M. Martínez-Zamora, Yamilet Coll, and Juan C. Díaz-Ricci. 2022. "Brassinosteroid Applications Enhance the Tolerance to Abiotic Stresses, Production and Quality of Strawberry Fruits" Horticulturae 8, no. 7: 572. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8070572