Enrichment Increases Aggression in Zebrafish


Department of Animal and Plant Sciences, University of Sheffield, Sheffield S10 2TN, UK
*
Author to whom correspondence should be addressed.
Fishes 2019, 4(1), 22; https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4010022
Submission received: 20 December 2018
/
Revised: 11 March 2019
/
Accepted: 12 March 2019
/
Published: 19 March 2019
(This article belongs to the Special Issue Welfare of Cultured and Experimental Fishes)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Environmental enrichment, or the enhancement of an animal’s surroundings when in captivity to maximise its wellbeing, has been increasingly applied to fish species, particularly those used commercially. Laboratory species could also benefit from enrichment, but it is not always clear what constitutes an enriched environment. The zebrafish, Danio rerio, is used widely in research and is one of the most commonly-used laboratory animals. We investigated whether changing the structural complexity of housing tanks altered the behaviour of one strain of zebrafish. Fish were kept in three treatments: (1) very enhanced (VE); (2) mildly enhanced (ME); and (3) control (CT). Level of aggression, fertilisation success, and growth were measured at regular intervals in a subset of fish in each treatment group. The VE fish were more aggressive over time than either ME or CT fish, both in the number of attacks they made against a mirror image and in their tendency to stay close to their reflection rather than avoid it. Furthermore, VE fish were shorter than CT fish by the end of the experiment, though mass was not significantly affected. There was no significant effect of treatment on fertilisation success. These findings suggest that the way in which fish are housed in the laboratory can significantly affect their behaviour, and potentially, their growth. The zebrafish is a shoaling species with a dominance hierarchy, and so may become territorial over objects placed in the tank. The enrichment of laboratory tanks should consider aspects of the species’ behaviour.
1. Introduction
Laboratory animal holding facilities conventionally have been designed on the basis of human requirements, both economic and ergonomic. Therefore, the captive conditions in which animals are kept are vastly different from their natural environment, and so the behaviour of such animals is likely to be unnatural. Indeed, captivity could adversely affect the behaviour and physiology of animals, for example, if their blood chemistry and metabolism are altered due to unnatural stress levels [1]. In an attempt to improve scientific validity, laboratories are beginning to alter housing facilities in order to simulate more natural environments, a concept known as environmental enrichment [2].
Environmental enrichment has traditionally focussed on mammalian species, specifically domestic and farm animals [1]. Although a variety of definitions exist, the emphasis is generally on increasing the amount of stimulation the captive environment provides for the animals [1]. Smith and Taylor [2] state that environmental enrichment should allow an animal to express a more natural behavioural repertoire. However, this highlights a significant problem: how to define the natural or normal behaviour of an animal [3]?
Despite fish being commonly used in laboratories [1], information about how to make them behave naturally or live stress-free lives could be improved. Environmental enrichment has been widely studied in commercially reared species of fish in an attempt to boost populations when they are released into the wild (e.g., [4,5,6]). However, less is known about the environmental and biological requirements of cultured species [7,8], and so improving the current knowledge of the impact of environmental enrichment on different fish species is important (see Reference [9] for a review of this subject).
Environmental enrichment within both commercial and laboratory aquaria often involves increasing the structural complexity of the tank by adding various objects to simulate natural habitats, including gravel, stones, real and artificial plants, plastic tubing, driftwood and upturned plant pots (e.g., [10,11,12]), and so reducing abnormal traits [9]. Another common technique used in fish studies involves altering tank lighting or colour (e.g., [13,14,15,16,17,18,19]) in an attempt to simulate a more natural environment.
Previous studies that compared fish reared in bare aquaria to those in enriched tanks found significant behavioural differences between the two. For example, fish in structured environments have been found to display reduced aggression levels (zebrafish, Danio rerio [20,21], salmonids [22]), possibly due to the structures reducing the visibility between individuals and so reducing aggressive interactions. Several studies have found that fish kept in bare tanks are more exploratory than those in tanks with structures added, which are more cautious in a novel environment (zebrafish [12], Atlantic salmon, Salmo salar [23]). However, Roberts et al. [23] found that when conspecific alarm cues were released into the water, Atlantic salmon housed in structurally complex tanks took significantly less time to leave a shelter.
There are mixed findings regarding the growth of fish in enriched and bare tanks, as some studies have found that fish kept in bare tanks are larger (Chinook salmon, Oncorhynchus tshawytscha [11], zebrafish [12]), whereas others show enriched fish to be larger (common carp, Cyprinus carpio [14], thinlip mullet, Liza ramada [19], Atlantic salmon [24]).
The zebrafish is an important vertebrate model organism in a variety of biological disciplines and is a commonly-used laboratory animal. It has been suggested that enrichment could improve the quality of care for zebrafish [25] and they seem to prefer structure in a tank [26], but there is a lack of specific guidelines regarding enrichment for this species. Indeed, plants, which are commonly used to enrich tanks, can have both positive and negative effects on fish (reduced forebrain, Chinook salmon [11], slower rate of learning, zebrafish [12], reduced aggression, zebrafish [20,21], reduced risk taking, Atlantic salmon [23]).
In this study, we investigated the effect of various types of tank enhancement, feasible in a laboratory setting, on the behaviour (aggression, boldness, activity), reproduction and growth of one strain of zebrafish by manipulating the tank environment. The results showed that aggression can increase in fish from tanks that have a higher structural diversity, and that growth may be affected.
2. Results
2.1. Aggression
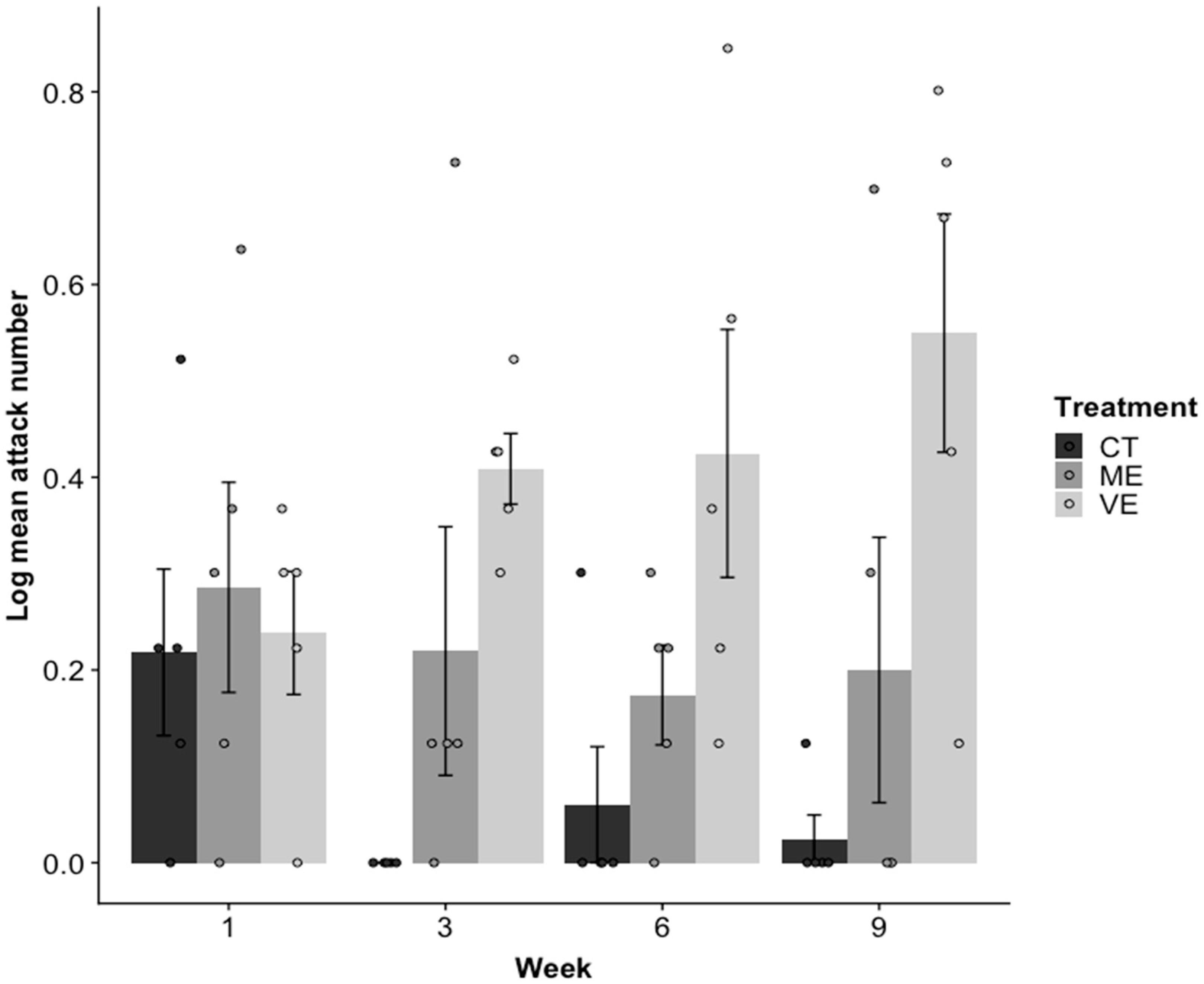
The type of tank enhancement that fish were kept in significantly affected the mean number of aggressive attacks fish made towards their image in the mirror test over the course of the experiment (R2 = 0.40, p = 0.034; Figure 1), with fish from very enhanced (VE) tanks making significantly more attacks than fish from both control (CT) and mildly enhanced (ME) tanks, given the week (CT: p = 0.017, ME: p = 0.036). Week affected the number of attacks differently over time for each enhancement treatment. VE tanks promoted more attacks over time, whereas CT and ME tanks had slightly fewer attacks over time (model estimates in comparison to VE; CT: −0.05, ME: −0.04).
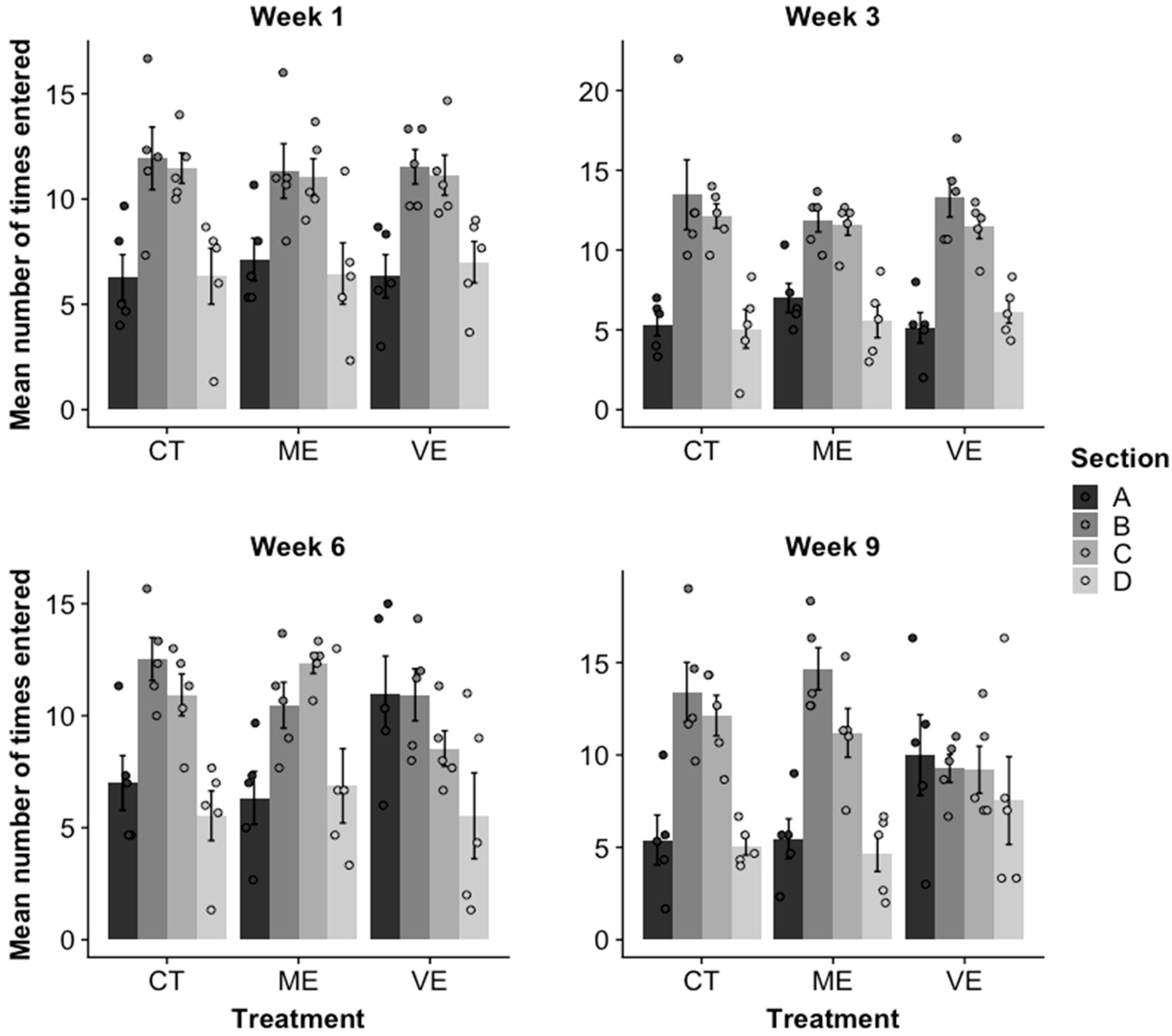
There was a significant interaction between section, treatment, and week for the mean number of times that fish entered a section (R2 = 0.55, p = 0.002; Figure 2). Fish preferred sections B and C throughout the experiment for CT and ME tanks, which suggested they were neither avoiding nor interacting with the mirror image. This was also true of fish in the VE tanks for weeks 1 and 3. However, in week 6, fish spent more time in sections A and B than C and D. This suggests that fish in the VE tank began to interact with the mirror. In week 9, fish in the VE tank entered each section approximately the same number of times.
The number of aggressive attacks conducted by fish was significantly correlated with the time they spent closest to their mirror image (section A) throughout the experiment (Pearson correlation, r = 0.27, df = 58, p < 0.05), suggesting that section preference was an indication of aggressiveness.
2.2. Boldness
Boldness, the propensity of individuals to explore a novel environment, was not significantly affected by type of tank enhancement (p = 0.85) or week (p = 0.43). The mean freezing time, when fish were completely still and often associated with anxiety, did not differ significantly among enhancement treatments (p = 0.58) or between weeks (p = 0.45).
2.3. Activity Level
Activity level, the degree to which fish actively moved in their home tank, was not significantly affected by type of tank enhancement (p = 0.98) or week (p = 0.75). Overall, activity level (Mean ± SE: 88.3 ± 2.1) was significantly lower than boldness (246.6 ± 7.3) (Two-sample t-test: t = 20.75, df = 118, p < 0.001).
2.4. Reproduction
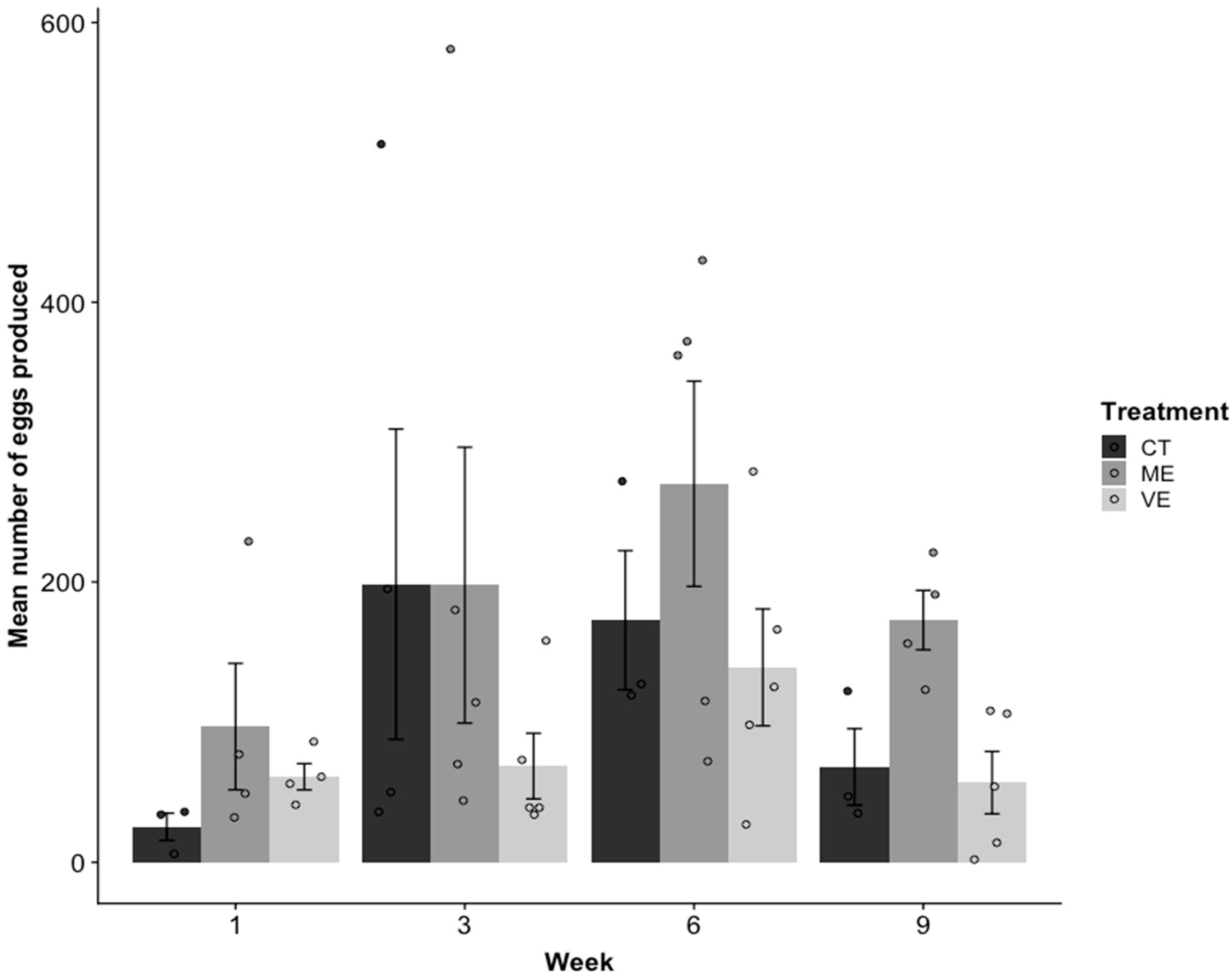
The mean number of eggs produced by females was significantly affected by the type of tank enhancement (R2 = 0.32, p = 0.004; Figure 3). Mild enhancement (ME) saw females producing 54% more eggs than control tanks (CT) and 81% more than very enhanced (VE) (CT: p = 0.05; VE: p = 0.001). The mean number of eggs produced in CT and VE tanks was not significantly different from each other (p = 0.32). Week also significantly affected the mean number of eggs produced (p = 0.003; Figure 3). Fewer eggs were produced in Week 1 than in Weeks 3 and 6 (Week 3: estimate = 0.83, p = 0.01; Week 6: estimate = 1.11, p < 0.001). However, the mean number of eggs was not significantly different between Weeks 1 and 9 (p = 0.24).
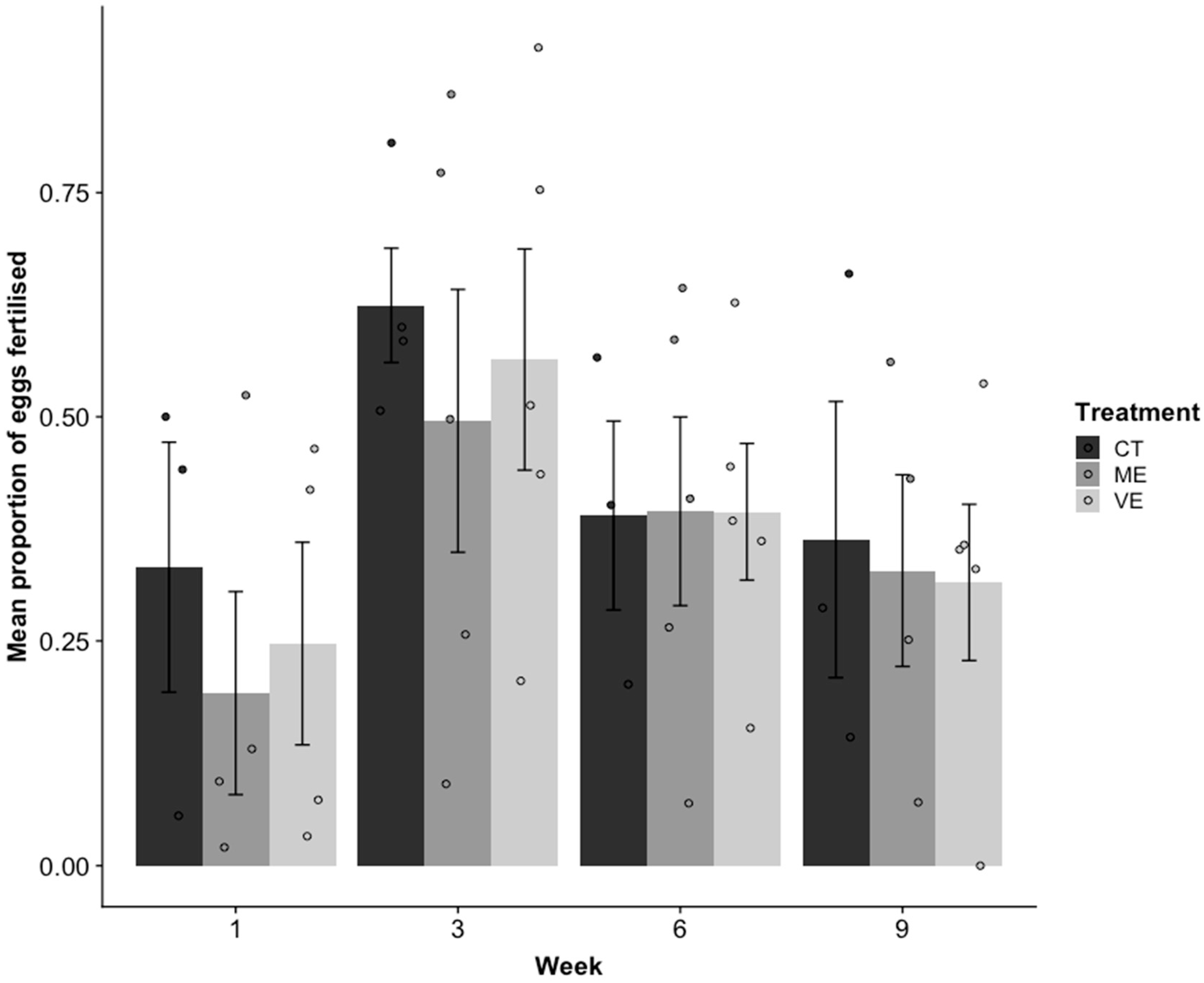
The proportion of fertilised eggs differed significantly between weeks (R2 = 0.22, p = 0.01; Figure 4). Fertilisation was significantly lower in Week 1 than Week 3 (p = 0.001), but was not significantly different from Week 6 (p = 0.09) or Week 9 (p = 0.32). Week 3 had a higher proportion of eggs fertilised than all other weeks (p < 0.001). There was no significant difference in the proportion of eggs fertilised between treatments (p = 0.57).
2.5. Growth
At the start of the experiment, the mean mass of fish differed significantly between the different enhancement tanks (Mean ± SE: CT 0.487 ± 0.008; ME 0.514 ± 0.010; VE 0.462 ± 0.008; ANOVA: F2,12 = 8.92, p < 0.01) because the fish randomly placed into the ME tanks were significantly heavier than those placed into the VE tanks (Tukey’s test: p < 0.01). This pattern remained consistent at the end of the experiment in Week 9. Again, enhancement treatment had a significant effect on mean mass (Mean ± SE: CT 0.484 ± 0.017; ME 0.505 ± 0.011; VE 0.445 ± 0.009; ANOVA: F2,12 = 5.90, p < 0.05), with ME fish being significantly heavier than VE fish (Tukey’s test: p < 0.05), and there was no significant difference between fish in CT tanks and either ME tanks (Tukey’s test: p = 0.49) or VE tanks (Tukey’s test: p = 0.11). There was no significant change in mass between the start and the end for fish in any treatment (CT: Paired t-test: t = 0.21, df = 4, p = 0.84; ME: Paired t-test: t = 1.25, df = 4, p = 0.28; VE: Paired t-test: t = 1.35, df = 4, p = 0.25).
At the start of the experiment, there was no significant difference in the mean length of the fish allocated to each enhancement type (Mean ± SE: CT 30.459 ± 0.160; ME 30.480 ± 0.178; VE 30.030 ± 0.233; ANOVA: F = 1.74, df = 2, p = 0.22). However, at the end of the experiment, in Week 9, there was a significant effect of enhancement treatment on mean length (Mean ± SE: CT 30.526 ± 0.341; ME 30.138 ± 0.255; VE 29.436 ± 0.105; ANOVA: F = 4.76, df = 2, p < 0.05) and fish housed in VE tanks were significantly shorter than fish in CT tanks (Tukey’s test: p < 0.05).
3. Discussion
Overall, our results show that the tank environment can significantly affect the behaviour, and potentially, the growth of zebrafish, at least in the strain studied. Aggression levels, both in terms of the number of aggressive attacks made and the time spent close to their mirror image, were significantly increased in fish from VE tanks compared to fish from the less (ME) or non-enhanced (CT) treatments. This increase in aggression towards their mirror image was seen from Week 3 of the experiment onwards, and time spent close to the mirror was found to correlate with the number of aggressive attacks throughout the experiment. We did not measure aggression in the tanks themselves, so it is possible that fish behaved differently when tested in front of a mirror in isolation.
In contrast, previous studies have found a decrease in aggression with structural complexity both in a laboratory setting (crayfish, Orconectes propinquus [27], zebrafish [20,21]) and in semi-natural environments (white-spotted char, Salvelinus leucomaenis [22]). Both Basquill and Grant [20] and Carfagnini et al. [21] found that zebrafish aggression rate was significantly reduced when measured in structurally complex versus bare environments, though the densities that Basquill and Grant [20] used were very low, so the fish may have been visually obscured by structures in the tank. Furthermore, both Carfagnini et al. [21] and Basquill and Grant [20] used juvenile, rather than adult zebrafish, which could suggest that life stage is important. Hamilton and Dill [28] found no difference in the aggression levels of adult zebrafish in a vegetated and a non-vegetated habitat. Our experiments ran for nine weeks and it was only after three weeks that aggression was seen to increase, suggesting that there were long-term effects on aggression when structure was present. Furthermore, as mentioned already, we measured aggression under controlled conditions, whereas in these other studies it was measured in the experimental tanks.
A possible explanation for the increased aggression in the VE fish in our study is that the fish become territorial over the objects within the tank and so their aggression level increased when measured with the mirror test. The zebrafish is a shoaling species, known to develop dominance hierarchies [29,30,31]. Previous studies have shown that zebrafish are highly territorial over spawning sites [32] and will defend food sources [28,33]. The structure within the VE tanks could have represented potential spawning sites, though there was no difference in the number of eggs collected in this treatment, which might be expected if fish were spawning in alternative sites in some of the treatments. Alternatively, VE fish may have become territorial over the upturned plant pots and artificial plants because food often became trapped under or around them, and so the fish could have been more aggressive when there was more structure because they were defending this resource. McCarthy et al. [34] found that in rainbow trout (Oncorhynchus mykiss), territories were very easy to defend when the density was low, but almost impossible to defend at high densities, and so aggressiveness over potential territories could vary with density, as found in the damselfish, Stegastes partitus [35,36], with aggression highest at intermediate densities like those used here. In our study, a random sample of fish was chosen from each tank for the mirror test and an equal sex ratio of fish was maintained in each tank, making it highly likely that the increased aggression observed was in both sexes, unless one sex was easier to catch than the other and so was always tested, but we have no evidence for this.
Increased plasma cortisol levels, known to be associated with stress in fish (e.g., salmonids [37]), have been demonstrated in fish displaying aggressive interactions during the development of territories and dominance hierarchies (cichlid, Haplochromis burtoni [38], rainbow trout [39]). Wilkes et al. [40] found no difference in cortisol levels or aggression between juvenile zebrafish kept in structured tanks and those in bare tanks. Pairs of zebrafish were found to show increased cortisol levels initially when housed with vegetation, but this declined with time, so by day 10 fish had lower levels than those kept individually or in pairs without vegetation [41]. Cortisol was not measured in our experiments, but the increased aggression within VE tanks is likely to have a negative impact on fish physiology and welfare, in which case enhancement of housing as implemented here may be detrimental, rather than beneficial, as usually assumed [20,21,22]. However, some degree of aggression may not be completely negative, according to the concept of allostasis [42]. The effect of enrichment on aggression may be age- and species-specific, and dependent on the territoriality of the species in question.
Fish housed in the VE tanks were significantly shorter than those in the CT tanks by the end of the experiment, and a significant change in length of these fish was detected, though no comparable change in mass was detected. This suggests there may have been an effect of treatment on growth rate, but the results are not conclusive. Previous studies have found that structure in a tank can have a positive effect on growth. For example, Finstad et al. [24] found that in Atlantic salmon, mass loss (common over winter [43]) was lowest when structural complexity was highest, possibly because there was less need for anti-predator behaviour and less competition for shelters. However, consistent with our study, others have found that fish in bare tanks have higher growth rates than those with structure (Chinook salmon [11], zebrafish [12]), though some have found no effect of structure on growth (zebrafish [44]), but an effect on morphology (three-spine stickleback, Gasterosteus aculeatus [45]). Kihslinger et al. [11] found that after one year of rearing, hatchery Chinook salmon were significantly heavier than their wild counterparts and similar results have been seen in the laboratory. Spence et al. [12] found that zebrafish reared in structurally complex environments were significantly shorter than those from bare tanks. A suggested explanation for reduced growth of fish subjected to structural complexity is that the objects within the tank trap food, which is consequently less accessible to the fish, as was observed in our study. Alternatively, an increase in aggression and a decrease in growth could be due to a trade-off in energy balance.
We did not find an increase in fertilisation success with increasing enhancement. While ME tanks produced more eggs than either of the other two housing treatments, the proportion of fertilised eggs did not differ. Carfagnini et al. [21] housed zebrafish in structurally complex and bare tanks and found that fecundity was not affected by housing complexity. However, they did find that when levels of aggression were highest, fecundity decreased [21], possibly due to elevated metabolic costs associated with high aggression levels.
We found no significant difference in the boldness of fish from the different treatments, a result in common with other studies on guppies (Poecilia reticulata) and three-spine sticklebacks [46,47]. There has been considerable debate about the measurement of boldness and whether it could simply be a reflection of the animal’s general activity level [48] but this has been found not to be the case [49]. In this study, fish were found to cross significantly more lines in the open-field test than they did in their home tanks but there was no difference between treatments. In contrast, von Krogh et al. [44] found that zebrafish from bare tanks had higher locomotor activity than those from structurally complex tanks.
The current study found no significant effect of tank background (ME tanks) on the behaviour of zebrafish, and so it appears that structural complexity within the tank and not tank background alone affected the fish. Previous work has found that fish housed in tanks with light-coloured backgrounds are more aggressive (Atlantic charr, Salvelinus alpines [50]), more exploratory (zebrafish [51]), have higher growth rates (common carp [14], thinlip mullet [19]) and have a higher oxygen consumption (scaled carp, Cyprinus carpio [13]). However, no significant effect of tank background was found in pot-bellied seahorses (Hippocampus abdominalis) [15] and Atlantic cod, Gadus morhua [17], suggesting that the effect of tank colour or background may be species-specific. It could also be dependent on the variability of the light conditions in which species live naturally, as the more variable they are, the less likely it is that differences in tank colour will have an effect.
In conclusion, this study has found that the enhancement of fish tanks, by adding structure, can significantly affect the aggressive behaviour of adult zebrafish, at least in this strain and when tested in isolation, and, potentially, their growth. Laboratories need to consider species-specific behaviour before enrichment is implemented.
4. Methods
4.1. Fish, Treatments and Growth
All fish used in the experiments were adult Nacre zebrafish. Stock fish were housed in 10-litre tanks (30 × 15 × 24 cm) in a recirculating system. Tanks were kept at 26 ± 1 °C under a 12:12 h light:dark photoperiod with 40 min dusk to dawn simulations. Fish were fed twice a day with ground-up flake food and day-old brine shrimp (Artemia salina).
Fish were housed in tanks of one of three treatment groups: (1) very enhanced (VE), (2) mildly enhanced (ME) or (3) control (CT) (Figure 5). Very enhanced tanks contained one artificial plant and one upturned plant pot (7 cm diameter). Additionally, plastic boards were backed with an aquarium backing paper (a blue seascape design) and placed against the rear walls of these tanks. Mildly enhanced tanks contained only the seascape boards against the rear walls, and CT tanks were left completely bare. These definitions were used purely to describe the experimental treatments and were not used as value judgements. Ten size-matched fish (five males, five females) were randomly placed in each treatment tank and there were five replicate tanks of each treatment group.
Fish were given a nine-day acclimatisation period, after which behavioural measurements were taken. The behavioural tests were done on a subset of three randomly-chosen fish per tank, and fecundity and fertilisation success were measured per tank. These measurements were taken at regular intervals throughout the course of the experiment at weeks 1, 3, 6, and 9. Standard length (tip of snout to caudal peduncle) and mass of each fish was measured at the start and end of the experiment in weeks one and nine. Previous work has shown that boldness and aggression are not related to size in either sex in this strain of zebrafish [49]. Lighting during behavioural trials was provided by two 18 W daylight fluorescent tubes placed approximately 40 cm above the test tank. Test tanks were filled with water from the recirculating system kept at the same condition as water within the housing tank system. Fish were tested before feeding in order to standardise this.
4.2. Behavioural Tests
4.2.1. Aggression
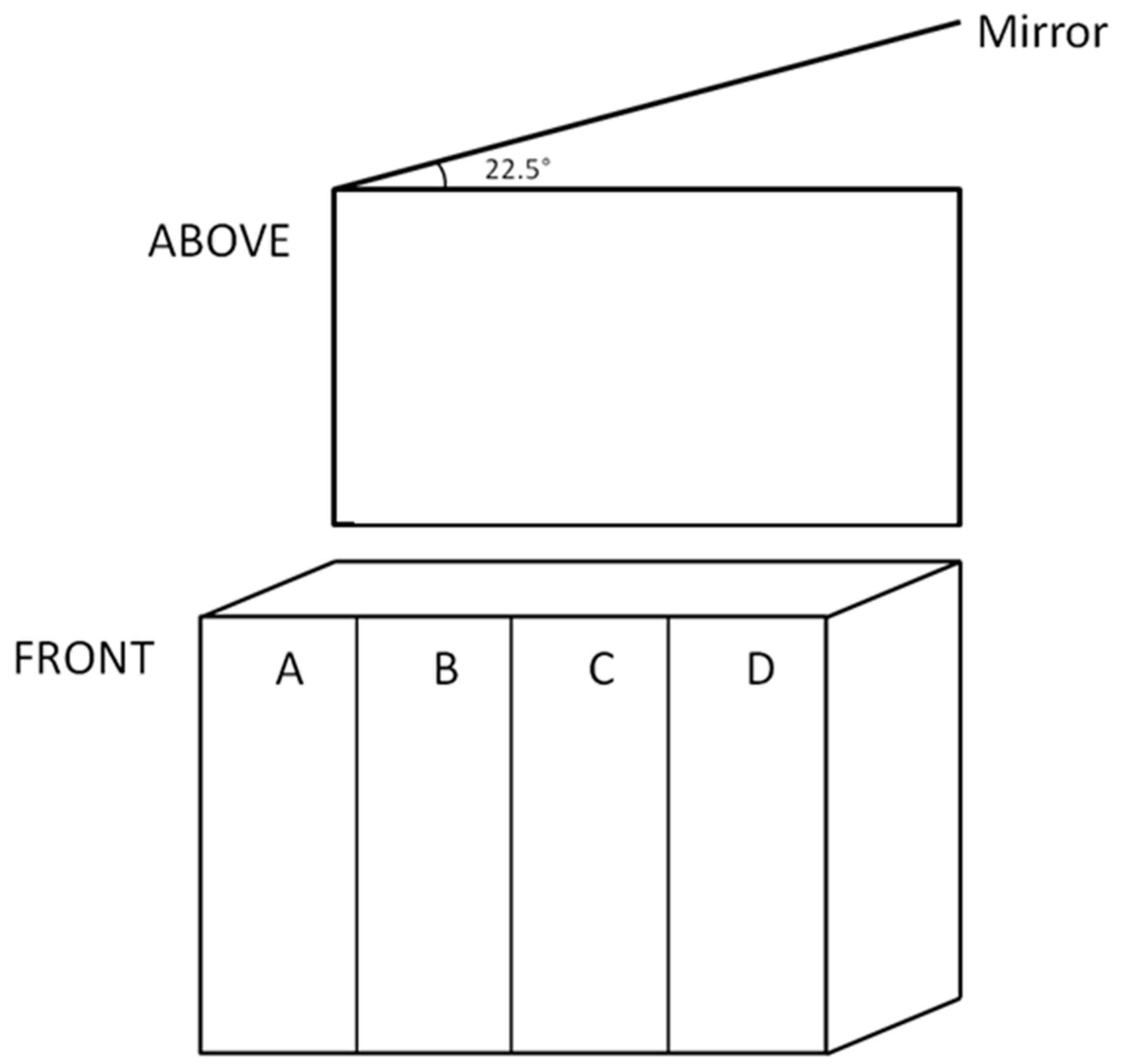
Aggression was tested using the mirror test [52,53]. A test tank (28 × 17.5 × 17 cm) was filled with 4.8 L of water and a mirror (45 × 38 cm) was placed at the back of the tank at an angle of 22.5° (Figure 6). A single fish was added to the tank and allowed to acclimatise for 60 s with the mirror covered. The cover was then removed and the number of aggressive attacks that a fish performed towards its mirror image was recorded over a 180 s period. The area of the test tank in which a fish spent most of its time was also quantified immediately after the aggressive attacks were recorded. The front of the test tank was marked into four equal vertical sections (Figure 6), and the section in which the fish was located was recorded every 5 s for 180 s. A fish that spent much of its time in section A could be regarded as aggressive because it was closer to its image, whereas a fish in section D may be avoiding aggression.
4.2.2. Boldness
To test for boldness, the open-field test [54,55,56] was used. A single fish was added to the test tank (48 × 23 × 26.5 cm), which had a 4 × 6 grid on the bottom and was filled with 3.25 L of water; the fish was left to acclimatise for 60 s. Immediately after this period, the number of lines the fish crossed, taken as a measure of boldness [49], and the total freezing time (time spent completely still) within a 180 s period, were recorded. The sides of the test tank were covered with opaque paper to minimise disturbance to the fish caused by the close proximity of the observer.
4.2.3. Activity Level
Activity level was measured in home tanks by placing a 2 × 5 cm grid onto the front of the tank and recording the movements of a single fish over a period of 180 s. The number of lines the fish crossed in this period was counted. Three randomly chosen fish per tank were tested.
4.2.4. Reproduction
A spawning dish (12.5 cm in diameter) containing two layers of glass marbles was placed into each tank at 16:00 on the first day of each measurement period. When fish spawned over the marbles, the eggs dropped into the spaces, stopping fish from eating them and aiding in their collection [57,58]. Moreover, zebrafish are likely to choose to spawn in these dishes as they have been shown to prefer to spawn in a substratum that provides protection to the eggs [59]. At 10:00 the following day, all dishes were taken out of the tanks, marbles were carefully removed, and all eggs within each dish were counted and transferred to a Petri dish containing aquarium water. Egg production can be variable in zebrafish [32], and so the first introduction of a spawning dish did not always lead to egg production. Therefore, if the fish in a tank did not produce any eggs, a spawning dish was placed in the tank at 16:00 every day for one week until eggs were produced. After the eggs had been in the Petri dishes for 24 h, the number that were fertilised was counted. Fertilised eggs remain transparent after this time period, but dead and unfertilised eggs become white and opaque [60,61].
4.3. Data Analysis
All data analyses were conducted using the R statistical package, version 2.14.1 [62]. For all datasets, means were calculated for each tank and used in the analysis. All behavioural data were analysed with linear mixed-effects models using the nlme package (model outputs shown in Supplementary Material, Table S1). The mean attack number was log-transformed before being modelled with enhancement treatment and week as interaction terms; tank was included as a random effect in the model. The mean number of entries into a section was modelled with section, enhancement treatment, and week as a three-way interaction, with tank as a random effect. For both the mean number of attacks and number of entries, model fit was tested using the Akaike information criterion (AIC), and in both cases, no terms were dropped for either model.
Boldness was measured in two separate models: first, using the number of lines crossed within the tank as the response variable, and secondly with freeze time as the response variable. Fish activity was also modelled with the number of lines crossed in the tank as a response variable. In all the boldness and activity models, treatment and week were fixed effects, with an interaction between the two proving to be non-significant, and therefore dropped from all models, as determined by the model fit through AIC.
Reproduction was first examined with the mean number of eggs as the response variable using a negative binomial model in the “ggfortify” package, to account for over-dispersion in the data. Enhancement treatment and week were used as fixed effects and no significant interaction was found between the two. Secondly, the mean proportion of eggs fertilised was investigated using generalized linear models in the “stats” package, version 3.5.2, after being log-transformed to give normally distributed data. An interaction between enhancement treatment and week proved non-significant in explaining variation through AIC assessment of single-term deletions of fixed effects, and was therefore dropped from the model. Treatment also proved non-significant as a fixed effect and was also removed from the final model. Outputs for the reproduction models are shown in Supplementary Materials, Table S2. Paired t-tests were used to compare the before and after weight and length data.
This work had appropriate ethical approval from the University of Sheffield.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2410-3888/4/1/22/s1, Table S1: Outputs from behavioural generalised linear mixed-effect models of 1) aggression, 2) boldness, and 3) activity, Table S2: Model outputs of 1) egg number (mean), and 2) proportion of eggs fertilised.
Author Contributions
P.J.W. and M.A.W. conceived of and designed the experiments, M.A.W. conducted the work, L.A.W. and M.A.W. analysed the data, M.A.W. and P.J.W. wrote the paper.
Funding
This research received no external funding.
Acknowledgments
We thank Tolulope Ariyomo and Terry Burke for advice.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Williams, T.D.; Readman, G.D.; Owen, S.F. Key issues concerning environmental enrichment for laboratory-held fish species. Lab. Anim. 2009, 43, 107–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.P.; Taylor, V.P. Environmental enrichment resources for laboratory animals: 1965–1995: Birds, cats, dogs, farm animals, ferrets, rabbits and rodents. AWIC Resour. Ser. 1995, 2, 145–212. [Google Scholar]
- Newberry, R. Environmental enrichment: Increasing the biological relevance of captive environments. Appl. Anim. Behav. Sci. 1995, 44, 229–243. [Google Scholar] [CrossRef]
- Braithwaite, V.A.; Salvanes, A.G.V. Environmental variability in the early rearing environment generates behaviourally flexible cod: Implications for rehabilitating wild populations. Proc. R. Soc. Lond. B 2005, 272, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Johnsson, J.I.; Brockmark, S.; Näslund, J. Environmental effects on behavioural development consequences for fitness of captive-reared fishes in the wild. J. Fish Biol. 2014, 85, 1946–1971. [Google Scholar] [CrossRef] [PubMed]
- Rosengren, M.; Kvingedal, E.; Näslund, J.; Johnsson, J.I.; Sundell, K. Born to be wild: Effects of rearing density and environmental enrichment on stress, welfare, and smolt migration in hatchery-reared Atlantic salmon. Can. J. Fish. Aquat. Sci. 2017, 74, 396–405. [Google Scholar] [CrossRef]
- Ellis, T.; North, B.; Scott, A.P.; Bromage, N.R.; Porter, M.; Gadd, D. The relationships between stocking density and welfare in farmed rainbow trout. J. Fish Biol. 2002, 61, 493–531. [Google Scholar] [CrossRef]
- Ashley, P.J. Fish welfare: Current issues in aquaculture. Appl. Anim. Behav. Sci. 2007, 104, 199–235. [Google Scholar] [CrossRef]
- Näslund, J.; Johnsson, J.I. Environmental enrichment for fish in captive environments: Effects of physical structures and substrates. Fish Fish. 2016, 17, 1–30. [Google Scholar] [CrossRef]
- Kemp, P.S.; Armstrong, J.D.; Gilvear, D.J. Behavioural responses of juvenile Atlantic salmon (Salmo salar) to presence of boulders. River Res. Appl. 2005, 21, 1053–1060. [Google Scholar] [CrossRef]
- Kihslinger, R.L.; Lema, S.C.; Nevitt, G.A. Environmental rearing conditions produce forebrain differences in wild Chinook salmon Oncorhynchus tshawytscha. Comp. Biochem. Physiol. A 2006, 145, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Spence, R.; Magurran, A.E.; Smith, C. Spatial cognition in zebrafish: The role of strain and rearing environment. Anim. Cogn. 2011, 14, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Papoutsoglou, S.E.; Mylonakis, G.; Miliou, H.; Karakatsouli, N.P.; Chadio, S. Effects of background colour on growth performances and physiological responses of scaled carp (Cyprinus carpio L.) reared in a closed circulated system. Aquac. Eng. 2000, 22, 309–318. [Google Scholar] [CrossRef]
- Papoutsoglou, S.E.; Karakatsouli, N.; Louizos, E.; Chadio, S.; Kalogiannis, D.; Dalla, C.; Polissidis, A.; Papadopoulou-Daifoti, Z. Effect of Mozart’s music (Romanze-Andante of “Eine Kleine Nacht Musik”, sol major, K525) stimulus on common carp (Cyprinus carpio L.) physiology under different light conditions. Aquac. Eng. 2007, 36, 61–72. [Google Scholar] [CrossRef]
- Martinez-Cardenas, L.; Purser, G.J. Effect of tank colour on Artemia ingestion, growth and survival in cultured early juvenile pot-bellied seahorses (Hippocampus abdominalis). Aquaculture 2007, 264, 92–100. [Google Scholar] [CrossRef]
- Mesquita, F.O.; Godinho, H.P.; Azevedo, P.G.; Young, R.J. A preliminary study into the effectiveness of stroboscopic light as an aversive stimulus for fish. Appl. Anim. Behav. Sci. 2008, 111, 402–407. [Google Scholar] [CrossRef]
- Monk, J.; Puvanendran, V.; Brown, J.A. Does different tank bottom colour affect the growth, survival and foraging behaviour of Atlantic cod (Gadus morhua) larvae? Aquaculture 2008, 277, 197–202. [Google Scholar] [CrossRef]
- Cobcroft, J.M.; Battaglene, S.C. Jaw malformation in striped trumpeter Latris lineate larvae linked to walling behaviour and tank colour. Aquaculture 2009, 289, 274–282. [Google Scholar] [CrossRef]
- El-Sayed, A.F.M.; El-Ghobashy, A.E. Effects of tank colour and feed colour on growth and feed utilization of thinlip mullet (Liza ramada) larvae. Aquac. Res. 2011, 42, 1163–1169. [Google Scholar] [CrossRef]
- Basquill, S.P.; Grant, J.W.A. An increase in habitat complexity reduces aggression and monopolization of food by zebra fish (Danio rerio). Can. J. Zool. 1998, 76, 770–772. [Google Scholar] [CrossRef]
- Carfagnini, A.G.; Rodd, F.H.; Jeffers, K.B.; Bruce, A.E.E. The effects of habitat complexity on aggression and fecundity in zebrafish (Danio rerio). Environ. Biol. Fishes 2009, 86, 403–409. [Google Scholar] [CrossRef]
- Hasegawa, K.; Maekawa, K. Potential of habitat complexity for mitigating interference competition between native and non-native salmonid species. Can. J. Zool. 2008, 86, 386–393. [Google Scholar] [CrossRef]
- Roberts, L.J.; Taylor, J.; Garcia de Leaniz, C. Environmental enrichment reduces maladaptive risk-taking behaviour in salmon reared for conservation. Biol. Conserv. 2011, 144, 1972–1979. [Google Scholar] [CrossRef]
- Finstad, A.G.; Einum, S.; Forseth, T.; Ugedal, O. Shelter availability affects behaviour, size-dependent and mean growth of juvenile Atlantic salmon. Freshwat. Biol. 2007, 52, 1710–1718. [Google Scholar] [CrossRef]
- Reed, B.; Jennings, M. Guidance on the housing and care of zebrafish, Danio rerio; RSPCA: Horsham, UK, 2010. [Google Scholar]
- Kistler, C.; Hegglin, D.; Würbel, H.; König, B. Preference for structured environment on zebrafish (Danio rerio) and checker barbs (Puntius oligolepis). Appl. Anim. Behav. Sci. 2011, 135, 318–327. [Google Scholar] [CrossRef]
- Corkum, L.D.; Cronin, D.J. Habitat complexity reduces aggression and enhances consumption in crayfish. J. Ethol. 2004, 22, 23–27. [Google Scholar] [CrossRef]
- Hamilton, I.M.; Dill, L.M. Monopolization of food by zebrafish (Danio rerio) increases in risky habitats. Can. J. Zool. 2002, 80, 2164–2169. [Google Scholar] [CrossRef]
- Larson, E.T.; O’Malley, D.M.; Melloni, R.H. Aggression and vasotocin are associated with dominant-subordinate relationships in zebrafish. Behav. Brain Res. 2006, 167, 94–102. [Google Scholar] [CrossRef]
- Spence, R.; Gerlach, G.; Lawrence, C.; Smith, C. The behaviour and ecology of the zebrafish, Danio rerio. Biol. Rev. 2008, 83, 13–34. [Google Scholar] [CrossRef]
- Watt, P.J.; Skinner, A.; Hale, M.; Nakagawa, S.; Burke, T. Small subordinate male advantage in the zebrafish. Ethology 2011, 117, 1003–1008. [Google Scholar] [CrossRef]
- Spence, R.; Smith, C. Male territoriality mediates density and sex ratio effects on oviposition in the zebrafish, Danio rerio. Anim. Behav. 2005, 69, 1317–1323. [Google Scholar] [CrossRef]
- Grant, J.W.A.; Kramer, D.L. Temporal clumping of food arrival reduces its monopolization and defense by zebrafish, Brachydanio rerio. Anim. Behav. 1992, 44, 101–110. [Google Scholar] [CrossRef]
- McCarthy, I.D.; Carter, C.G.; Houlihan, D.F. The effect of feeding hierarchy on individual variability in daily feeding of rainbow trout, Oncorhynchus mykiss. J. Fish Biol. 1992, 41, 257–263. [Google Scholar] [CrossRef]
- Johnson, D.W. Combined effects of condition and density on post-settlement survival and growth of a marine fish. Oecologia 2008, 155, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Samhouri, J.F. Food supply influences offspring provisioning but not density-dependent fecundity in a marine fish. Ecology 2009, 90, 3478–3488. [Google Scholar] [CrossRef]
- Pickering, A.D.; Pottinger, T.G. Stress responses and disease resistance in salmonid fish: Effects of chronic elevation of plasma cortisol. Fish Physiol. Biochem. 1989, 7, 253–258. [Google Scholar] [CrossRef]
- Fox, H.E.; White, S.A.; Kao, M.H.F.; Fernald, R.D. Stress and dominance in a social fish. J. Neurosci. 1997, 17, 6463–6469. [Google Scholar] [CrossRef]
- Pottinger, T.G.; Pickering, A.D. The influence of social interaction on the acclimation of rainbow trout, Oncorhynchus mykiss, to chronic stress. J. Fish Biol. 1992, 41, 435–447. [Google Scholar] [CrossRef]
- Wilkes, L.; Owen, S.F.; Readman, G.D.; Sloman, K.A.; Wilson, R.W. Does structural enrichment for toxicology studies improve zebrafish welfare? Appl. Anim. Behav. Sci. 2012, 139, 143–150. [Google Scholar] [CrossRef]
- Keck, V.A.; Edgerton, D.S.; Hajizadeh, S.; Swift, L.L.; Dupont, W.D.; Lawrence, C.; Boyd, K.L. Effects of complexity on pair-housed zebrafish. J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 378–383. [Google Scholar]
- Korte, S.M.; Olivier, B.; Koolhaas, J.M. A new animal welfare concept based on allostasis. Phys. Behav. 2007, 92, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Næsje, T.F.; Thorstad, E.B.; Forseth, T.; Aursand, M.; Saksgård, R.; Finstad, A.G. Lipid class content as an indicator of critical periods for survival in juvenile Atlantic salmon (Salmo salar). Ecol. Freshw. Fish 2006, 15, 572–577. [Google Scholar] [CrossRef]
- Von Krogh, K.; Sørensen, C.; Nilsson, G.E.; Øverli, Ø. Forebrain cell proliferation, behavior, and physiology of zebrafish, Danio rerio, kept in enriched or barren environments. Physiol. Behav. 2010, 101, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Garduño-Paz, M.V.; Couderc, S.; Adams, C.E. Habitat complexity modulates phenotype expression through developmental plasticity in the threespine stickleback. Biol. J. Linn. Soc. 2010, 100, 407–413. [Google Scholar] [CrossRef] [Green Version]
- Brydges, N.M.; Braithwaite, V.A. Does environmental enrichment affect the behaviour of fish commonly used in laboratory work? Appl. Anim. Behav. Sci. 2009, 118, 137–143. [Google Scholar] [CrossRef]
- Burns, J.G.; Saravanan, A.; Rodd, F.H. Rearing environment affects the brain size of guppies: Lab-reared guppies have smaller brains than wild-caught guppies. Ethology 2009, 115, 122–133. [Google Scholar] [CrossRef]
- Réale, D.; Reader, S.M.; Sol, D.; McDougall, P.T.; Dingemanse, N.J. Intergrating animal temperament within ecology and evolution. Biol. Rev. 2007, 82, 291–318. [Google Scholar] [CrossRef]
- Ariyomo, T.O.; Watt, P.J. The effect of variation in boldness and aggressiveness on the reproductive success of zebrafish. Anim. Behav. 2012, 83, 41–46. [Google Scholar] [CrossRef]
- Höglund, E.; Balm, P.H.M.; Winberg, S. Behavioural and neuroendocrine effects of environmental background colour and social interaction in Arctic charr (Salvelinus alpines). J. Exp. Biol. 2002, 205, 2535–2543. [Google Scholar]
- Rosemberg, D.B.; Rico, E.P.; Mussulini, B.H.M.; Paiato, A.L.; Calcagnotto, M.E.; Bonan, C.D.; Dias, R.E.; Blaser, R.E.; Souza, D.O.; de Oliveira, D.L. Differences in spatio-temporal behavior of zebrafish in the open tank paradigm after a short-period confinement into dark and bright environments. PLoS ONE 2011, 6, e19317. [Google Scholar] [CrossRef]
- Gerlai, R.; Lahav, M.; Guo, S.; Rosenthal, A. Drinks like a fish: Zebra fish (Danio rerio) as a behaviour genetic model to study alcohol effects. Pharmacol. Biochem. Behav. 2000, 67, 773–782. [Google Scholar] [CrossRef]
- Moretz, J.A.; Martins, E.P.; Robison, B.D. The effects of early and adult social environment on zebrafish (Danio rerio) behavior. Environ. Biol. Fish. 2007, 80, 91–101. [Google Scholar] [CrossRef]
- Walsh, R.N.; Cummins, R.A. The open-field test: A critical review. Psychol. Bull. 1976, 83, 482–504. [Google Scholar] [CrossRef]
- Moretz, J.A.; Martins, E.P.; Robison, B.D. Behavioral syndromes and the evolution of correlated behaviour in zebrafish. Behav. Ecol. 2007, 18, 556–562. [Google Scholar] [CrossRef]
- Burns, J.G. The validity of three tests of temperament in guppies, Poecilia reticulata. J. Comp. Psychol. 2008, 122, 344–356. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio rerio), 4th ed.; Chapman and Hall: London, UK, 1995. [Google Scholar]
- Brand, M.; Granato, M.; Nüsslein-Volhard, C. Keeping and raising zebrafish. In Zebrafish: A Practical Approach; Nüsslein-Volhard, C., Dahm, R., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 7–37. [Google Scholar]
- Spence, R.; Ashton, R.; Smith, C. Oviposition decisions are mediated by spawning site quality in wild and domesticated zebrafish, Danio rerio. Behaviour 2007, 144, 953–966. [Google Scholar] [CrossRef]
- Mertens, J. Year-round controlled mass reproduction of the zebrafish Brachydanio rerio (Hamilton-Buchanan). Aquaculture 1973, 2, 245–249. [Google Scholar] [CrossRef]
- Gellert, G.; Heinrichsdorff, J. Effect of age on the susceptibility of zebrafish eggs to industrial waste water. Water Res. 2001, 35, 3754–3757. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011. [Google Scholar]
Figure 1.
Mean ± SE number of aggressive attacks made towards a mirror image in the mirror test in control (CT), mildly enhanced (ME), and very enhanced (VE) treatments in weeks 1, 3, 6, and 9. There were five replicates of each treatment in each week (each replicate is shown as a point).
Figure 1.
Mean ± SE number of aggressive attacks made towards a mirror image in the mirror test in control (CT), mildly enhanced (ME), and very enhanced (VE) treatments in weeks 1, 3, 6, and 9. There were five replicates of each treatment in each week (each replicate is shown as a point).

Figure 2.
Mean ± SE number of times fish entered sections A, B, C, and D in the mirror test in control (CT), mildly enhanced (ME), and very enhanced (VE) tank treatments in weeks 1, 3, 6, and 9. Section A was closest to the mirror, section D was furthest from the mirror, and sections B and C were in the middle of the tank. There were five replicates per treatment.
Figure 2.
Mean ± SE number of times fish entered sections A, B, C, and D in the mirror test in control (CT), mildly enhanced (ME), and very enhanced (VE) tank treatments in weeks 1, 3, 6, and 9. Section A was closest to the mirror, section D was furthest from the mirror, and sections B and C were in the middle of the tank. There were five replicates per treatment.

Figure 3.
Mean ± SE number of eggs produced from the control (CT), mildly enhanced (ME), and very enhanced (VE) tanks over weeks 1, 3, 6, and 9. Sample sizes varied between 3 and 5 tanks, depending on the number of tanks that produced eggs. There was a significant difference in mean number of eggs between treatments in week 9 only, where more eggs were produced in ME tanks.
Figure 3.
Mean ± SE number of eggs produced from the control (CT), mildly enhanced (ME), and very enhanced (VE) tanks over weeks 1, 3, 6, and 9. Sample sizes varied between 3 and 5 tanks, depending on the number of tanks that produced eggs. There was a significant difference in mean number of eggs between treatments in week 9 only, where more eggs were produced in ME tanks.

Figure 4.
Mean ± SE proportion of eggs fertilised from the control (CT), mildly enhanced (ME), and very enhanced (VE) tanks in weeks 1, 3, 6, and 9. Sample sizes vary between 3 and 5 tanks depending on the number of tanks that produced eggs.
Figure 4.
Mean ± SE proportion of eggs fertilised from the control (CT), mildly enhanced (ME), and very enhanced (VE) tanks in weeks 1, 3, 6, and 9. Sample sizes vary between 3 and 5 tanks depending on the number of tanks that produced eggs.

Figure 5.
The three housing treatments: (a) very enhanced (VE), (b) mildly enhanced (ME), and (c) control (CT).
Figure 5.
The three housing treatments: (a) very enhanced (VE), (b) mildly enhanced (ME), and (c) control (CT).

Figure 6.
Schematic of the front and top view of the test tank used for the mirror test.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Woodward, M.A.; Winder, L.A.; Watt, P.J. Enrichment Increases Aggression in Zebrafish. Fishes 2019, 4, 22. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4010022
AMA Style
Woodward MA, Winder LA, Watt PJ. Enrichment Increases Aggression in Zebrafish. Fishes. 2019; 4(1):22. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4010022
Chicago/Turabian StyleWoodward, Melanie A., Lucy A. Winder, and Penelope J. Watt. 2019. "Enrichment Increases Aggression in Zebrafish" Fishes 4, no. 1: 22. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4010022