1. Introduction
The development of aquaculture is essential to meet growing human food requirements. However, aquaculture can also significantly enrich waters with organic matter and nutrients [
1], thus making them more susceptible to eutrophication and which restricts the broadscale application of aquaculture techniques. In order to overcome these problems, there is a growing global interest in the integrated multitrophic aquaculture (IMTA) mode [
2]. IMTA cocultivates species of different trophic levels and ecological niches with complementary functions to achieve a healthy and sustainable aquaculture system [
3]. The shellfish–algae IMTA mode has proven to be the only economical solution for recycling wasted nutrients in open water [
2,
4]. Additionally, cultured shellfish and algae play an important role in the coastal biocarbon cycle [
5]. Carbon can be removed from the coastal ecosystem through the harvesting of shellfish and algae [
6], thereby increasing the carbon sequestration capacity of marginal shelf seas, accelerating the absorption of atmospheric CO
2 by the ocean, and alleviating the global greenhouse effect. Therefore, shellfish–algae farming is an efficient and economically important mode of environmentally friendly aquaculture that benefits the environment and the coastal carbon cycle [
6,
7]. However, most studies on the role of shellfish–algae culture in the carbon cycle have focused on the inorganic carbon cycle, and few have considered its impact on organic carbon [
7,
8].
At present, the most successful shellfish–algae IMTA modes have been based on the expansion of kelp culture [
9]. However, due to the intolerance of kelp to high temperatures, many shellfish–algae IMTA areas in northern China have large areas where only shellfish are being cultured for up to four months during summer. For example, Sanggou Bay is a key shellfish–algae IMTA farming base in North China that produces about 80,000 tons of kelp annually [
10]. After the summer kelp is harvested, about 1/6 of the kelp farming area will be used to cultivate another macroalgae species, namely the seasonal rotational aquaculture of macroalgae. The annual production of these rotated algae accounts for approximately 1/8 of the kelp yield (data from local government). The physiological activities of shellfish without active algae may stress the natural ecosystems of the aquaculture areas [
11]. Additionally, the stratification of seawater caused by freshwater input and high temperatures make eutrophication more likely in summer [
12,
13]. The combination of multiple environmental stressors such as temperature and eutrophication may also lead to a higher incidence of aquatic diseases in summer [
14,
15,
16]. Japanese scallop (
Patinopecten yessoensis) is a scallop that has become a major aquaculture species in northern China, covering the coastal area from Liaoning Province to Shandong Province. Recently, massive die-offs of Japanese scallop have been observed in Sanggou Bay during summer. This may be attributed to both extreme high temperatures and the absence of macroalgal cultures in summer [
2,
17].
Seaweed (
Gracilaria lemaneiformis) is an algae with a high commercial value and a strong bioremediation effect [
17,
18,
19] which grows fast, has a short growth cycle, and can be harvested in 2–3 months. This species can also adapt well to high temperature environments, so a rotation strategy that includes seaweed in summer can bring significant economic and environmental benefits. The physiological activity of this species in summer may also improve the conditions for the shellfish and reduce the frequency of summer shellfish die-offs. Therefore, in this study the effects of different Japanese scallop/seaweed culture modes on the marine environment and the local ecosystem were investigated in Sanggou Bay during summer. The objectives of this study were to identify the different environmental and ecological impacts of different culture modes, and to explore the benefits and feasibility of rotations that include seaweed in summer. We evaluated the impacts of shellfish–algae IMTA cultures on the marine environment and carbon cycle, identified any biologically mutual benefits, and illustrated the advantages of the IMTA culture mode. These results can be used as a reference for sustainable aquaculture activities in different regions.
4. Discussion
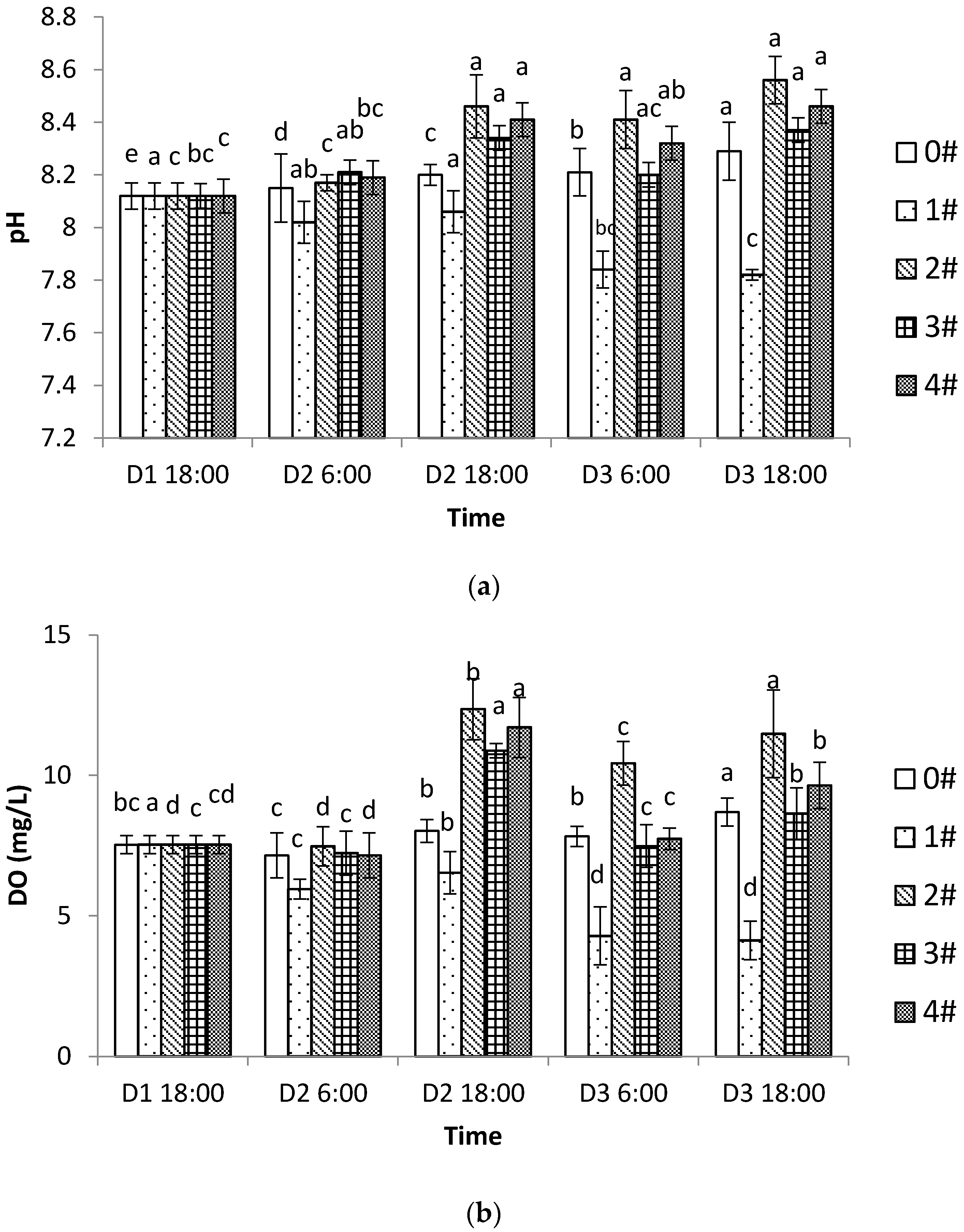
In summer, when much of the kelp is harvested under high temperatures, the main issue of shellfish culture becomes the impact of the substances released and excreted during their growth process on the aquatic environment. In our study, the pH and DO were significantly reduced (
p < 0.05) by the end of the experiment in group 1, where shellfish were cultured alone, and the DO concentration had dropped to below 5 mg/L. The increase in DIN concentration also demonstrated how shellfish cultures can exacerbate the risk of eutrophication. Zhang [
20] has reported that shellfish were unable to form shells at pH 7.3, at DO levels of 4–5 mg/L where the growth of fish was inhibited and some even died, and at DO of 3–4 mg/L where the mass mortality of crustaceans can occur [
21,
22,
23]. Therefore, if the shellfish culture scale is large, mismanagement may easily bring harm to the environment in semienclosed areas or in areas with poor hydrodynamics. Therefore, as during summer the common cocultivated algae, the species of kelp, cannot be grown due to high water temperature, it is necessary to use macroalgal rotation strategies for cocultures to avoid the negative effects of shellfish monocultures.
In the shellfish–algae IMTA areas, after the summer kelp is harvested, the kelp farming area can be used to cultivate seaweed, which can improve environmental parameters and fully absorb the nutrients in the water, thereby alleviating the environmental pressure caused by the shellfish culture [
24,
25,
26,
27,
28]. However, only the appropriate ratio of shellfish to algae can achieve an environmentally friendly aquaculture system [
7]. Generally, it seems that the greater the proportion of algae, the greater the benefit to environmental indicators (e.g., pH and DO) [
2]. However, in low nutrient areas, or areas with poor water exchange, cultivation with too large a proportion of algae may lead to the excessive consumption of nutrients such as DIC, N, and P [
29]. The same conclusion was reached in our study. At the end of the experiment, the DIC of group 2 with only algae was significantly reduced and the DIN was nearing the threshold of 2 μmol/L at which point N becomes a limiting factor for phytoplankton growth [
30,
31]. In contrast, at the end of the experiment in the shellfish–algae IMTA mode group, N:P:Si did not deviate from the Redfield ratio (N:P:Si = 16:1:16) [
32] so that there was no potential limiting nutrient, thus demonstrating that the shellfish–algae mode had little to no negative influence on the ambient nutrient structure.
Chl-
a reflects the biomass of the phytoplankton, which is the main food source for shellfish. The phytoplankton community faces downward controlling effects due to shellfish ingestion and upward controlling effects due to competition with macroalgae for nutrients during growth [
33,
34]. In our study, when the ratio of shellfish to algae was 1:1, the concentration of Chl-
a at end of the experiment was significantly higher than that in other experimental groups. This was because shellfish have phytoplankton size preferences and actively select the size class on which they feed, and Japanese scallop mainly feeds on nanophytoplankton (2–20 μm) [
35,
36]. This meant that the growth of microphytoplankton (>20 μm) and picophytoplankton (0.45–2 μm) were not impeded in this shellfish–algae system. In order to avoid eutrophication and food shortages for shellfish, the Chl-
a concentration should be maintained at a stable level. Furthermore, it appeared that in the actual culture process if the proportion of algae is too low the upward controlling effect of macroalgae on nutrient competition was not strong, which could lead to overpopulating by picophytoplankton and microphytoplankton.
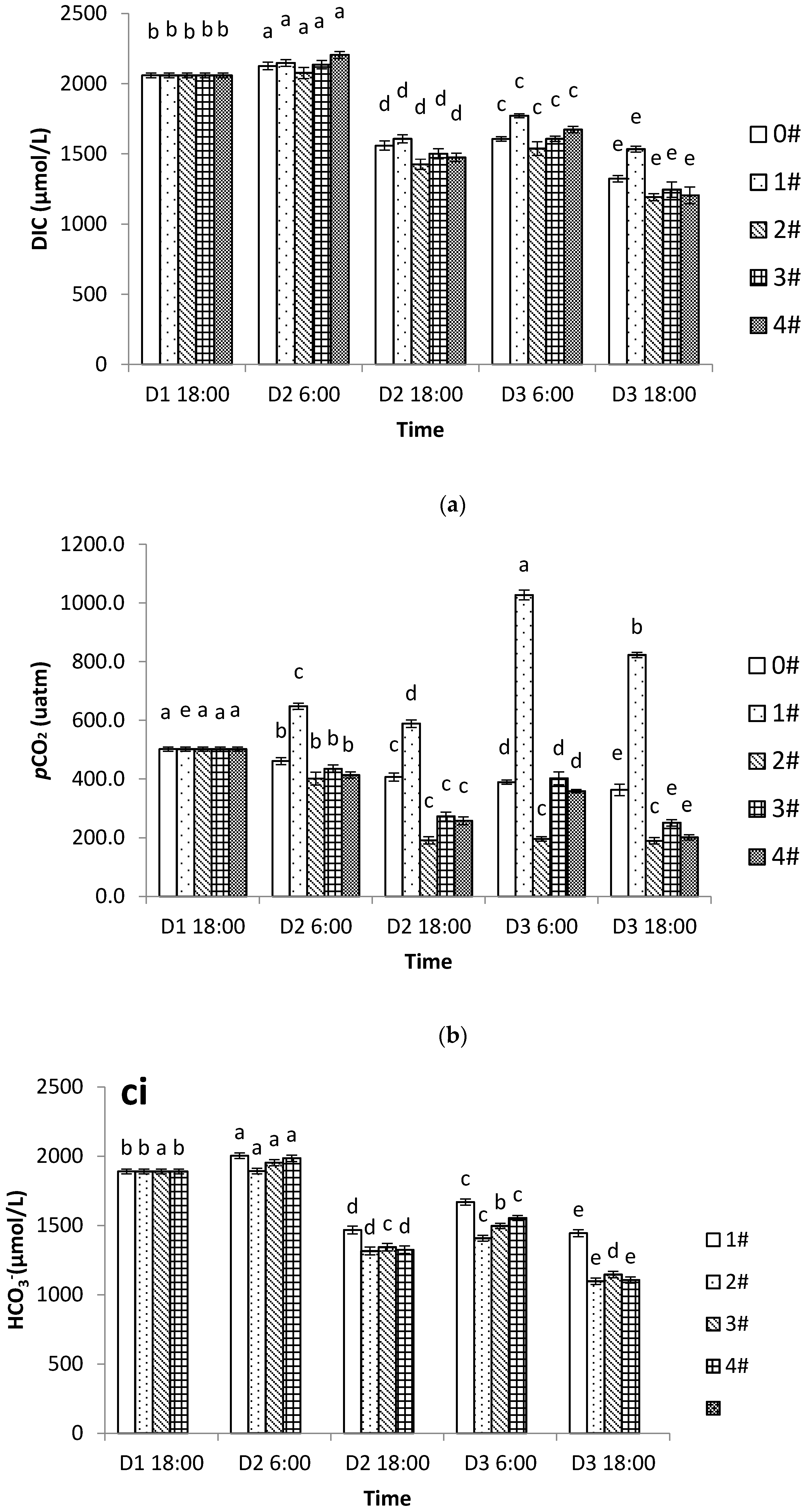
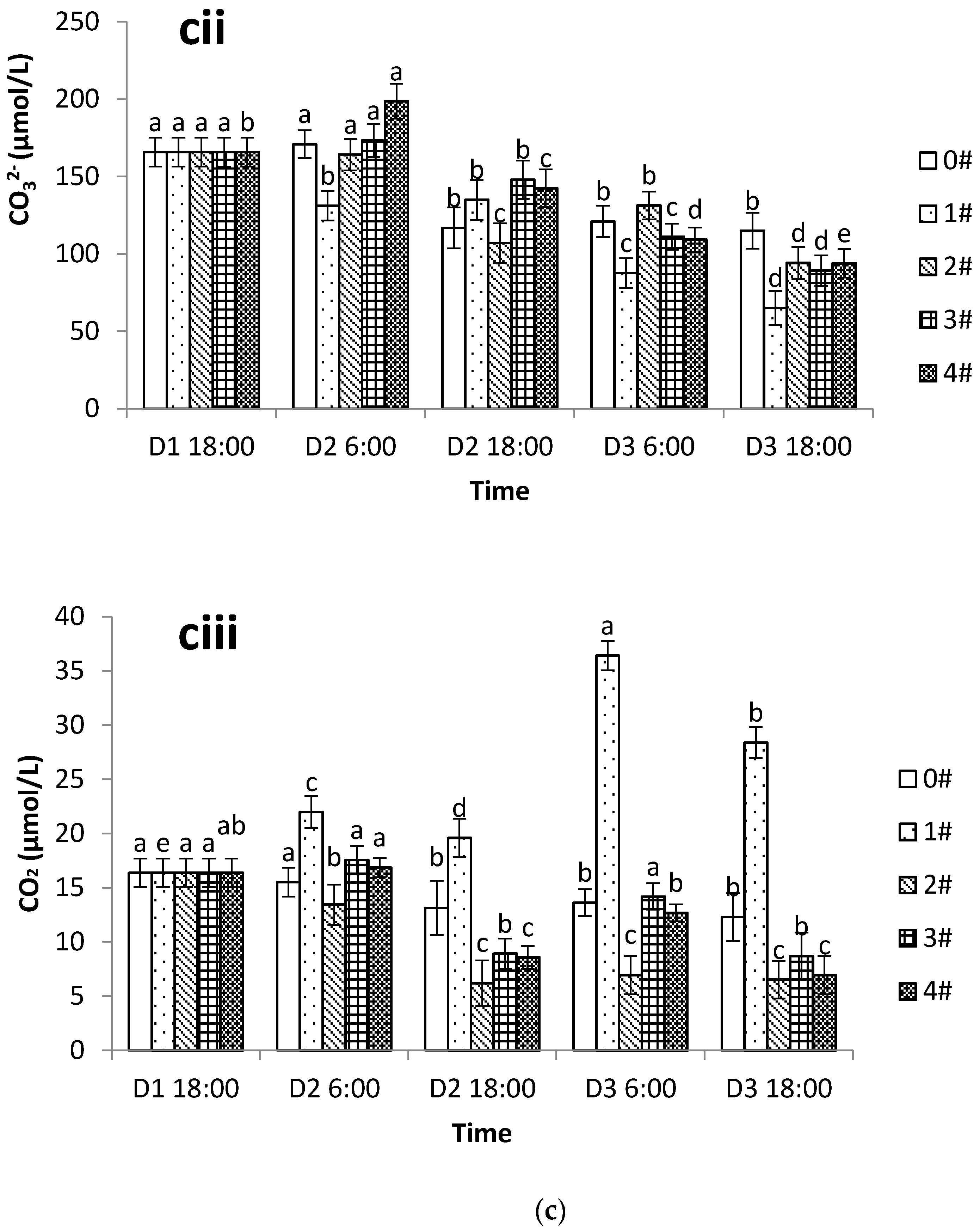
Under high temperature conditions the respiration rate of scallops and CO
2 production increased but the IMTA mode was able to effectively absorb the nutrients released by the shellfish and thereby prevent the accumulation of CO
2 produced by respiration. In the shellfish–algae IMTA mode, 70% of the CO
2 released by shellfish can be absorbed by algae [
8], making the aquaculture system a net carbon sink [
7]. Han [
37] reported that when the
Chlamys farreri to seaweed ratio exceeded 1:0.31, the inorganic carbon system in the water was improved. Likewise, this study found that the DIC, carbonate system components, and the
pCO
2 were all significantly reduced in the shellfish–algae polyculture group. We also found that the greater the proportion of algae, the more significant the impact on the inorganic carbon cycle. It should be noted that cultivation with too large a proportion of algae might lead to the excessive consumption of nutrients, and that the pursuit of improving the inorganic carbon system of the water body needs to be tempered by the awareness of other environmental parameters. Our study verified the importance of the appropriate culture ratios in practical production. At the end of the experiment, when the ratio of shellfish to algae was 1:1.6, the
pCO
2 and the inorganic carbon system components were not significantly different from those in group 2 where the algae were cultured alone. Therefore, it seems possible to assume that the most significant effect on the inorganic carbon cycle was observed in summer when the ratio of Japanese scallop to seaweed was 1:1.6 under experimental conditions. However, in nature there is water exchange in areas occupied by aquaculture, and the real level of water quality parameters may differ significantly from those obtained in a closed experimental system. The optimal ratio of scallop to algae may also differ significantly from that which is experimentally determined in a closed system. Further large-scale field studies are needed to assess the optimal scallop and algae ratio.
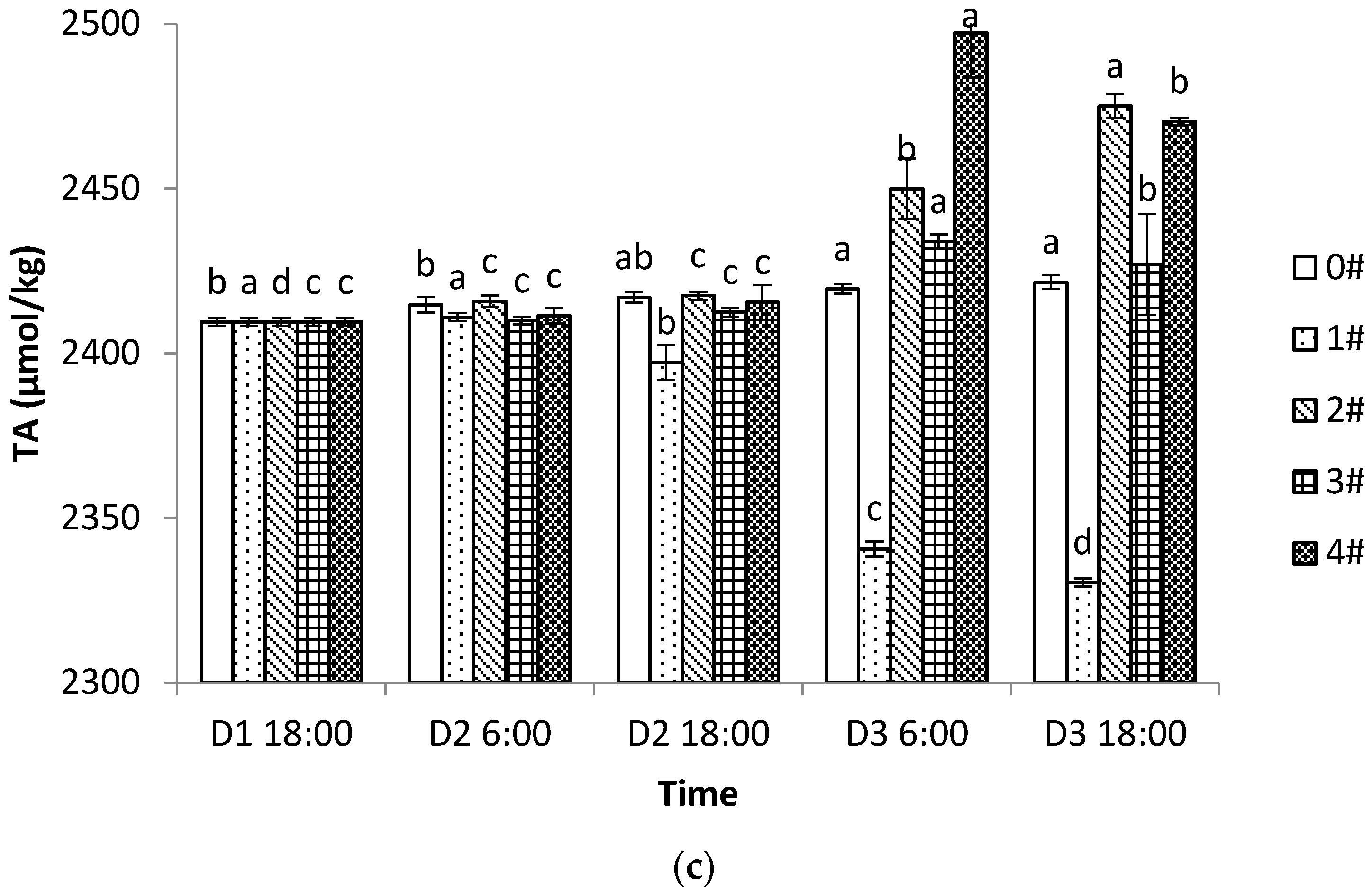
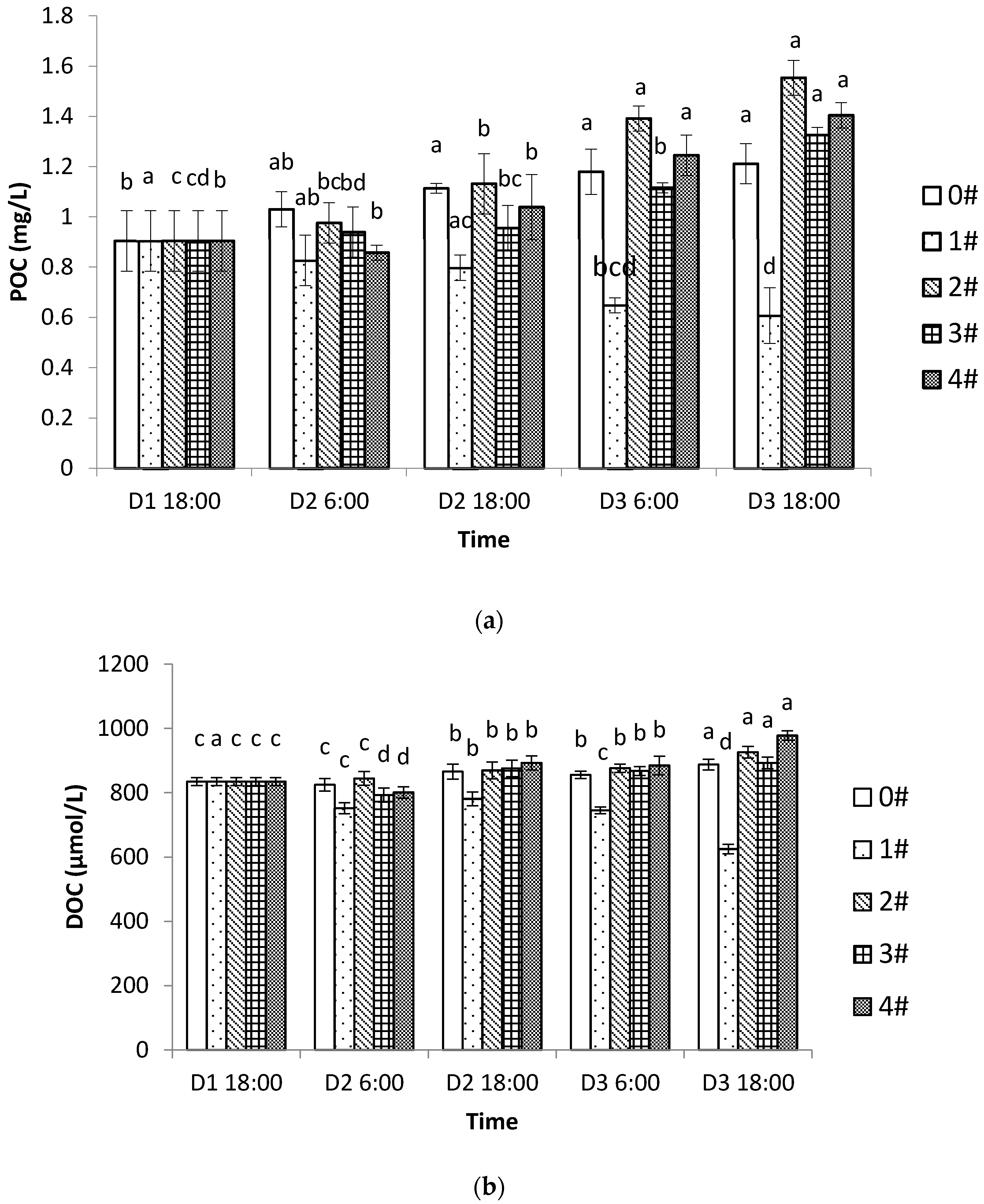
Although shellfish–algae farming influences the inorganic carbon system in the water, the “inorganic carbon pump” only enables the transport of CO
2 from the atmospheric carbon reservoir to the oceanic carbon reservoir. For the long-term storage of CO
2, the CO
2 entering the ocean must be entrained by the “biological carbon pump” [
38]. Therefore, the cycling of organic carbon by the aquaculture process is important and should also be considered. Although CO
2 emissions from shellfish respiration do not affect the TA (i.e., the ability to neutralize hydrogen ions) in seawater [
20], calcification by shellfish decreases the TA, which limits the ability of seawater to absorb CO
2 and further affects the carbonate chemistry equilibrium [
39]. Therefore, if we only look at the direct evidence of inorganic carbon system parameters in the water, shellfish farming should be a source of carbon. However, it has been established that shellfish aquaculture can be a carbon sink, not only via absorbing the DIC and synthesizing the shells, but also through its efficient water filtration and biological activities, which can effectively filter out particulate carbon and transport it to the sediment [
6,
8]. In our study, it was found that the POC and the DOC had decreased significantly by the end of the experiment in group 1, which had only shellfish cultures. Studies on the shellfish carbon balance have found that 30% of the carbon used by shellfish during a growth cycle was retained by the shellfish itself, 30% sunk to the bottom of the sea as waste, and 30–40% was released by the respiration and calcification processes [
40,
41]. Therefore, the controversy around whether shellfish aquaculture is a carbon source or sink should not only be based only on the inorganic carbon cycle but should include more detailed studies that track inorganic and organic carbon as well as the physiological processes of the shellfish themselves. Comprehensive studies such as this one may provide insight into the controversial issue of carbon sequestration in shellfish. Additionally, in the groups containing algae, the DOC content significantly increased. The DOC makes up a portion of organic carbon that is effectively sequestered as it enters food webs via the action of microfood loops or forms inert organic carbon and stays in the sea for a long time [
42,
43]. According to Jiao [
44], microbial carbon sequestration as a carbon sink is often overlooked, and the core mechanism of microbial carbon sequestration is its conversion of DOC into refractory DOC (RDOC) that is stored in seawater for extended periods. Therefore, the organic carbon cycle also plays an important role as a carbon sink in aquaculture systems. The shellfish–algae IMTA mode not only changes the inorganic carbon system, but also effectively changes the organic carbon system, thus increasing the function and role of aquaculture as a carbon sink.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}