
Human Activities, Biostratigraphy and Past Environment Revealed by Small-Mammal Associations at the Chalcolithic Levels of El Portalón de Cueva Mayor (Atapuerca, Spain)

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Site
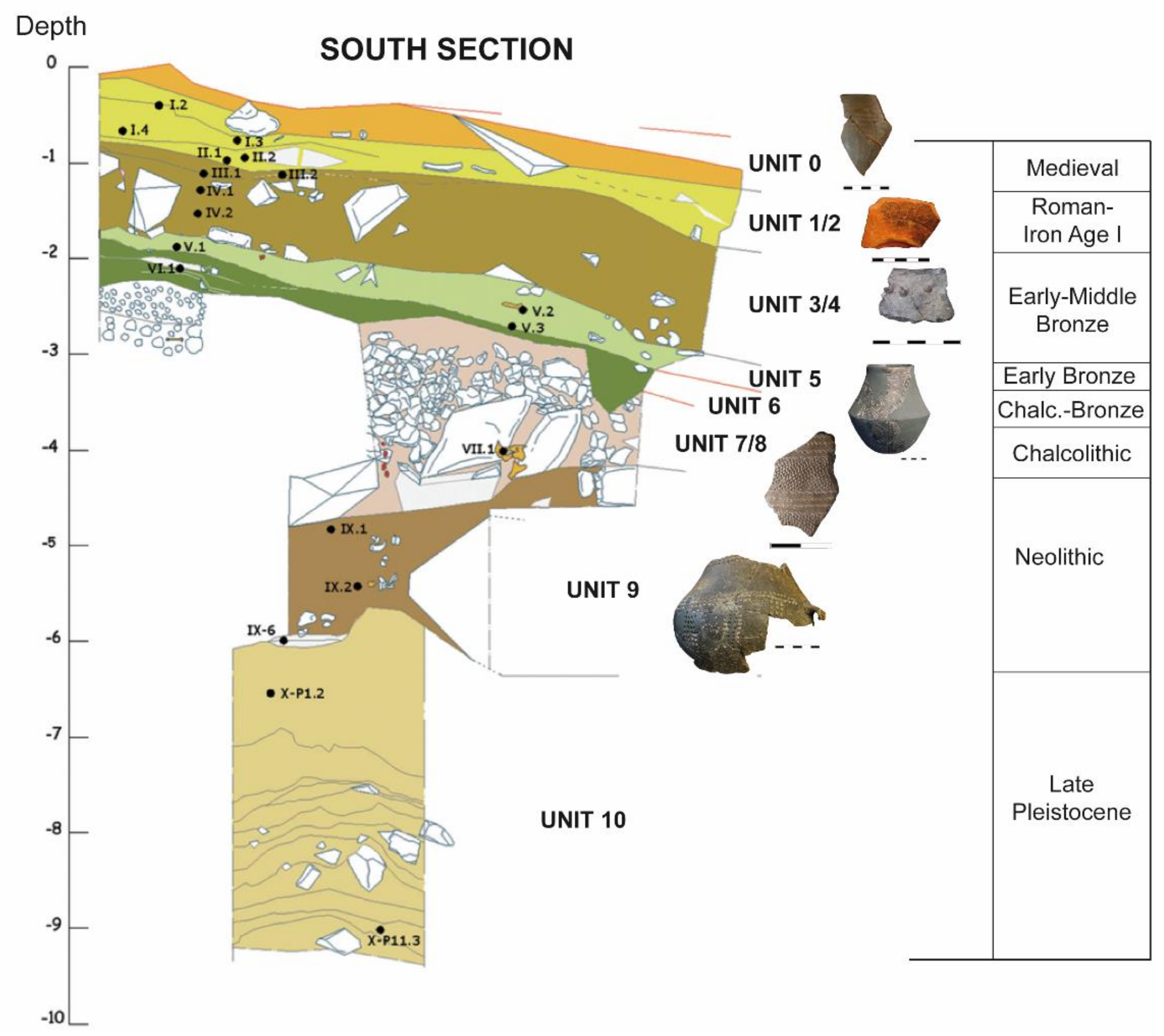
2.2. Archaeo-Stratigraphic Units and Chronology
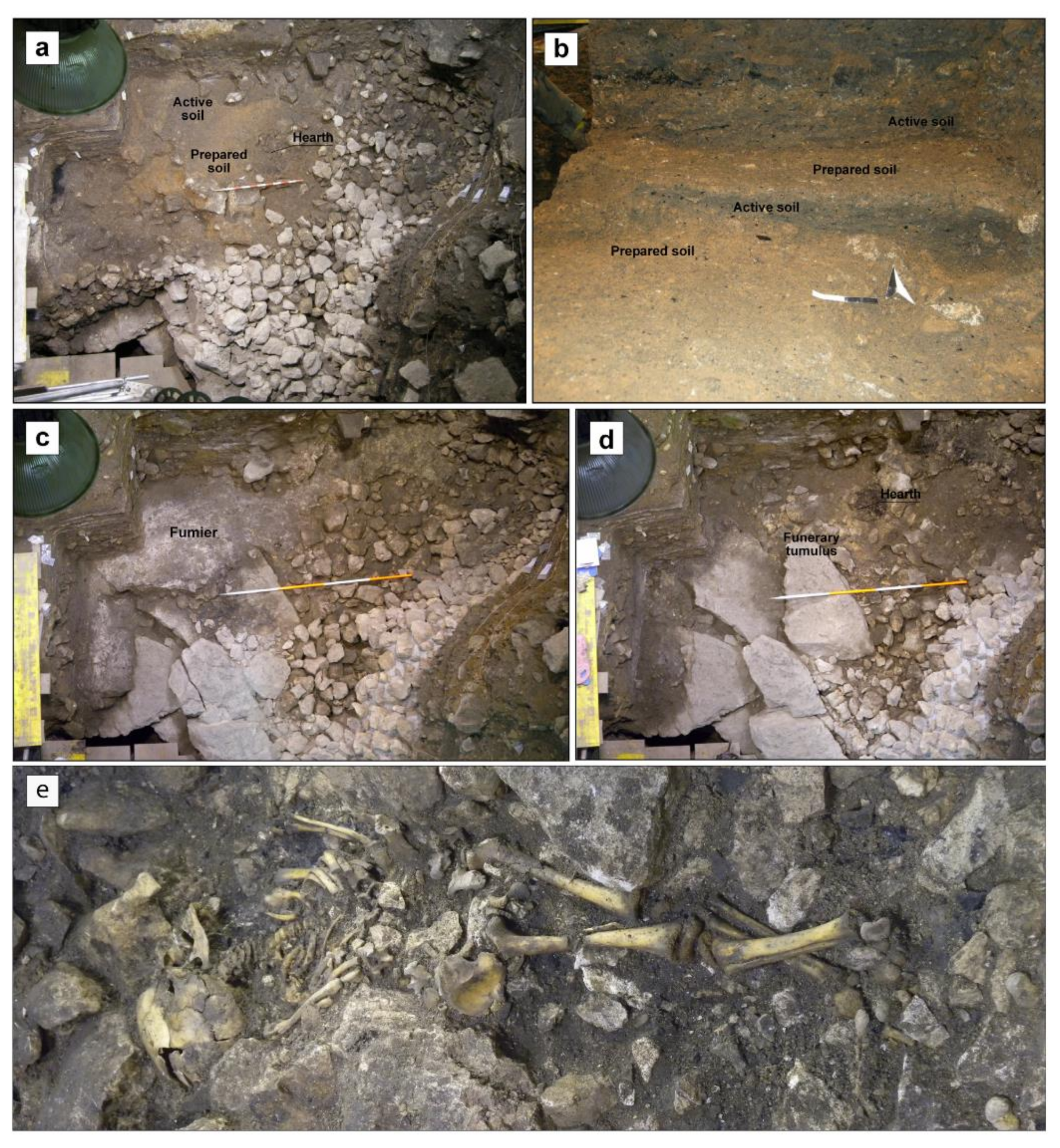
2.2.1. Early Chalcolithic (Pre-Bell-Beaker) Funerary Context (UE79)
2.2.2. Late Chalcolithic (Late Pre-Bell-Beaker and Bell-Beaker) Habitat and Stabling Context
2.2.3. Chronology
2.3. Samples
2.4. Taxonomic Identification and Quantification
2.5. Statistical Analysis
2.6. Paleo-Environmental Estimations
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mellet, J.S. Scatological origin of microvertebrate fossil accumulations. Science 1974, 185, 349–350. [Google Scholar] [CrossRef] [PubMed]
- Andrews, P.O. Caves and Fossils: Predation, Preservation and Accumulation of Small Mammal Bones in Caves, with an Analysis of the Pleistocene Cave Faunas from Westbury-Sub-Mendip, Somerset, UK; University of Chicago Press: Chicago, IL, USA, 1990. [Google Scholar]
- Fernández-Jalvo, Y.; Andrews, P.; Denys, C.; Sesé, C.; Stoetzel, E.; Marin-Monfort, D.; Pesquero, D. Taphonomy for taxonomists: Implications of predation in small mammal studies. Quat. Sci. Rev. 2016, 139, 138–157. [Google Scholar] [CrossRef]
- Cuenca-Bescós, G.; Rofes, J.; López-García, J.M.; Blain, H.-A.; De Marfá, R.J.; Galindo-Pellicena, M.A.; Bennásar-Serra, M.L.; Melero-Rubio, M.; Arsuaga, J.L.; Bermúdez de Castro, J.M.; et al. Biochronology of Spanish Quaternary small vertebrate faunas. Quat. Int. 2010, 212, 109–119. [Google Scholar] [CrossRef]
- Cuenca-Bescós, G.; Blain, H.-A.; Rofes, J.; López-García, J.M.; Lozano-Fernández, I.; Galán, J.; Núñez-Lahuerta, C. Updated Atapuerca biostratigraphy: Small-mammal distribution and its implications for the biochronology of the Quaternary in Spain. C. R. Palevol 2016, 15, 621–634. [Google Scholar] [CrossRef]
- Cuenca-Bescós, G.; Straus, L.G.; González-Morales, M.R.; García-Pimienta, J.C. The reconstruction of past environments through small mammals: From the Mousterian to the Bronze Age in El Mirón Cave (Cantabria, Spain). JAS 2009, 36, 947–955. [Google Scholar] [CrossRef]
- López-García, J.M.; Berto, C.; Colamussi, V.; Dalla Valle, C.; Lo Vetro, D.; Luzi, E.; Malavasi, G.; Martini, F.; Sala, B. Palaeoenvironmental and palaeoclimatic reconstruction of the Latest Pleistocene-Holocene sequence from Grotta del Romito (Calabria, southern Italy) using the small-mammal assemblages. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 409, 169–179. [Google Scholar] [CrossRef]
- Rofes, J.; Murelaga, X.; Martínez-García, B.; Bailon, S.; López-Quintana, J.C.; Guenaga-Lizasuc, A.; Ortega, L.A.; Zuluaga, M.C.; Alonso-Olazabal, A.; Castaños, J.; et al. The long paleoenvironmental sequence of Santimamiñe (Bizkaia, Spain): 20,000 years of small mammal record from the latest Late Pleistocene to the middle Holocene. Quat. Int. 2014, 339–340, 62–75. [Google Scholar] [CrossRef]
- Rofes, J.; Garcia-Ibaibarriaga, N.; Aguirre, M.; Martínez-García, B.; Ortega, L.; Zuluaga, M.C.; Bailon, S.; Alonso-Olazabal, A.; Jone Castaños, J.; Murelaga, X. Combining Small-Vertebrate, Marine and Stable-Isotope Data to Reconstruct Past Environments. Sci. Rep. 2015, 5, 14219. [Google Scholar] [CrossRef]
- Galán, J.; Núñez-Lahuerta, C.; López-García, J.; Cuenca-Bescós, G. Did humans disturb bats? Exploring the hominin-chiropter interactions in the Sierra de Atapuerca sites (early to Middle Pleistocene, Spain). Quat. Sci. Rev. 2019, 226, 106018. [Google Scholar] [CrossRef]
- Cucchi, T.; Barnett, R.; Martínková, N.; Renaud, S.; Renvoisé, E.; Evin, A.; Sheridan, A.; Mainland, I.; Wickham-Jones, C.; Tougard, C.; et al. The changing pace of insular life: 5000 Years of microevolution in the Orkney vole (Microtus Arvalis Orcadensis). Evolution 2014, 68, 2804–2820. [Google Scholar] [CrossRef] [Green Version]
- Rofes, J.; Cucchi, T.; Hanot, P.; Herman, J.; Stephan, P.; Cersoy, S.; Ivan Horácek, I.; Kerr, E.; Allberry, K.; Valenzuela, S.; et al. Postglacial recolonization and Holocene diversification of Crocidura suaveolens (Mammalia, Soricidae) on the north-western fringe of the European continent. Quat. Sci. Rev. 2018, 190, 1–10. [Google Scholar] [CrossRef]
- Wood, W.R.; Lee Johnson, D. 9-A Survey of Disturbance Processes in Archaeological Site Formation. In Advances in Archaeological Method and Theory; Schiffer, M.B., Ed.; Academic Press: San Diego, CA, USA, 1978; pp. 315–381. [Google Scholar]
- Texier, J. A propos des processus de formation des sites préhistoriques/About prehistoric site formation processes. Paléo, Revue d’Archéologie Préhistorique 2000, 12, 379–386. [Google Scholar] [CrossRef]
- Dalland, M.; Carter, S. The evaluation of a prehistoric mound damaged by rabbit burrowing at Maryton Law, Angus. Tayside Fife Archaeol. J. 1998, 4, 20–30. [Google Scholar]
- Mallye, J.-B. Badger (Meles meles) remains within caves as an analytical tool to test the integrity of stratified sites: The contribution of Unikoté Cave (Pyrénées-Atlantiques, France). J. Taphon. 2011, 9, 15–36. [Google Scholar]
- Pelletier, M.; Brugal, J.-P.; Cochard, D.; Lenoble, A.; Mallye, J.-B.; Royer, A. Identifying fossil rabbit warrens: Insights from a taphonomical analysis of a modern warren. JAS Rep. 2016, 10, 331–344. [Google Scholar] [CrossRef]
- Rofes, J.; Cersoy, S.; Zazzo, A.; Royer, A.; Nicod, P.-Y.; Laroulandie, V.; Langlais, M.; Pailler, Y.; Leandri, C.; Leandri, F.; et al. Detecting stratigraphical issues using direct radiocarbon dating from small-mammal remains. J. Quat. Sci. 2020, 35, 505–513. [Google Scholar] [CrossRef]
- Ordiales, A.; Rofes, J.; Iriarte, E.; Mardones, V.; Cuenca-Bescós, G.; Carretero, J.M.; Arsuaga, J.L.; García-Ibaibarriaga, N. Small mammals as tools to characterize archaeological contexts in the Chalcolithic of Portalón de Cueva Mayor (Atapuerca, Burgos, Spain). In Proceedings of the 1st Young Natural History Scientists’ Meeting, Paris, France, 12–14 February 2014; p. 12. [Google Scholar]
- Carbonell, E.; Bermúdez de Castro, J.M.; Arsuaga, J.L.; Díez, J.C.; Rosas, A.; Cuenca-Bescós, G.; Sala, R.; Mosquera, M.; Rodríguez, X.P. Lower Pleistocene hominids and artifacts from Atapuerca TD6 (Spain). Science 1995, 269, 826–830. [Google Scholar] [CrossRef] [Green Version]
- Carbonell, E.; Bermúdez de Castro, J.M.; Parés, J.M.; Pérez-González, A.; Cuenca-Bescós, G.; Olle, A.; Mosquera, M.; Huguet, R.; van der Made, J.; Rosas, A.; et al. The first hominin of Europe. Nature 2008, 452, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Arsuaga, J.L.; Bermúdez de Castro, J.M.; Carbonell, E. (Eds.) The Sima de los Huesos hominid Site. J. Hum. Evol. 1997, 33, 105–421. [Google Scholar] [CrossRef]
- Bermúdez de Castro, J.M.; Carbonell, E.; Arsuaga, J.L. (Eds.) Gran Dolina Site: TD6 Aurora Stratum (Burgos, Spain). J. Hum. Evol. 1999, 37, 309–700. [Google Scholar]
- Clark, G.; Straus, L.G.; Burton, S.S.; Jackson-Clark, V. The North Burgos Archaelogical Survey: An inventory of cultural remains. In The North Burgos Archaelogical Survey. Bronze and Iron Age archaeology on the Meseta del Norte (Province of Burgos, North-Central Spain) 19, 18 156; Clark, G., Ed.; Arizona State University, Department of Antropology: Tempe, AZ, USA, 1979. [Google Scholar]
- Apellaniz, J.M.; Domingo, D. Estudios sobre Atapuerca (Burgos): II. Los materiales de superficie del Santuario de la Galería del Sílex. Cuad. Arqueol. Univ. Deusto 1987, 10, 1–342. [Google Scholar]
- Carretero, J.M.; Ortega, A.I.; Juez, L.; Pérez-González, A.; Arsuaga, J.L.; Pérez, R.; Ortega, M.C. A Late Pleistocene-Early Holocene archaeological sequence of Portalón de Cueva Mayor (Sierra de Atapuerca, Burgos, Spain). Munibe 2008, 59, 67–80. [Google Scholar]
- Pérez-Romero, A.; Carretero, J.M.; Juez, L.; Ortega, A.I.; Ortega, M.C.; Arsuaga, J.L. Una dobla almohade del Siglo XIII en el yacimiento de El portalón de Cueva Mayor, Sierra de Atapuerca (Burgos). Nvmisma 2010, 254, 85–106. [Google Scholar]
- Pérez-Romero, A.; Carretero, J.M.; Alday, A.; Arsuaga, J.L. La Cerámica Protohistórica e Histórica en el yacimiento de El Portalón de Cueva Mayor, Sierra de Atapuerca, Burgos. Bol. Soc. Esp. Ceram. Vidr. 2013, 52, 183–193. [Google Scholar] [CrossRef]
- Pérez-Romero, A.; Iriarte, E.; Galindo-Pellicena, M.A.; García-González, R.; Rodríguez, L.; Castilla, M.; Francés-Negro, M.; Santos, E.; Valdiosera, C.; Arsuaga, J.L.; et al. An unusual Pre-bell beaker copper age cave burial context from El Portalón de Cueva Mayor site (Sierra de Atapuerca, Burgos). Quat. Int. 2017, 433, 142–155. [Google Scholar] [CrossRef]
- Vergés, J.M.; Allué, E.; Angeluccí, D.E.; Cebriá, A.; Fontanals, M.D.; Mányanos, A.; Montero, S.; Moral, S.; Vaquero, M.; Zaragoza, J. La sierra de Atapuerca durante el Holoceno: Datos preliminares sobre las ocupaciones de la Edad del Bronce en la cueva de Mirador (Ibeas de Juarros, Burgos). Trab. Prehist. 2002, 59, 107–126. [Google Scholar] [CrossRef] [Green Version]
- Vergés, J.M.; Allué, E.; Angeluccí, D.E.; Burjachs, F.; Carrancho, A.; Cebriá, A.; Expósito, I.; Fontanals, M.; Moral, S.; Rodríguez, A.; et al. Los niveles neolíticos de la cueva del Mirador (Sierra de Atapuerca Burgos): Nuevos datos sobre la implantación y el desarrollo de la economía agropecuaria en la submeseta norte. In Proceedings of the IV Peninsular Neolithic Congres, MARQ, Alicante, Spain; Hernández, M.S., Soler, J.A., López, J.A., Eds.; Museo Arqueológico de Alicante, Diputación de Alicante: Alicante, Spain, 2008; pp. 418–427. [Google Scholar]
- López-García, J.M.; Blain, H.-A.; Cuenca-Bescós, G.; Ruiz-Zapata, M.B.; Dorado-Valiño, M.; Gil-García, M.J.; Valdeolmillos, A.; Ortega, A.I.; Carretero, J.M.; Arsuaga, J.L.; et al. Palaeoenvironmental and palaeoclimatic reconstruction of the Latest Pleistocene of El Portalón Site, Sierra de Atapuerca, northwestern Spain. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 292, 453–464. [Google Scholar] [CrossRef]
- Castilla, M.; Carretero, J.M.; Gracia, A.; Arsuaga, J.L. Evidence of Rickets and/or Scurry in a complete Chalcolithic Child Skeleton from the El Portalón Site (Sierra de Atapuerca, Spain). JASs Rep. 2014, 92, 1–16. [Google Scholar]
- Günther, T.; Valdiosera, C.; Malmström, H.; Ureña, I.; Rodriguez-Varela, R.; Osk Sverrisdóttir, O.; Daskalaki, E.A.; Skoglund, P.; Naidoo, T.; Svensson, E.M.; et al. Ancient genomes link early farmers from Atapuerca in Spain to modern-day Basques. Proc. Natl. Acad. Sci. USA 2015. [Google Scholar] [CrossRef] [Green Version]
- Reimer, P.J.; Austin, W.E.N.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Ramsey, C.B.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Arsuaga, J.L.; Gómez-Olivencia, A.; Sala, N.; Martínez-Pillado, V.; Pablos, A.; Bonmatí, A.; Pantoja, A.; Lira, J.; Alcázar de Velasco, A.; Ortega, A.I.; et al. Evidence of paleoecological changes and Mousterian occupations at the Galería de las Estatuas site, Sierra de Atapuerca, northern Iberian plateau, Spain. Quat. Res. 2017, 88, 345–367. [Google Scholar] [CrossRef] [Green Version]
- Bañuls-Cardona, S.; López-García, J.M.; Morales-Hidalgo, J.I.; Cuenca-Bescós, G.; Vergés, J.M. Late Glacial to Late Holocene palaeoclimatic and palaeoenvironmental reconstruction of El Mirador cave (Sierra de Atapuerca, Burgos, Spain) using the small-mammal assemblages. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 471, 71–81. [Google Scholar] [CrossRef]
- Cuenca-Bescós, G.; Rossell, J.; Morcillo-Amo, A.; Galindo-Pellicena, M.A.; Santos, E.; Moya-Costa, R. Beavers (Castoridae, Rodentia, Mammalia) from the Quaternary sites of the Sierra de Atapuerca, in Burgos, Spain. Quat. Int. 2017, 433, 263–277. [Google Scholar] [CrossRef]
- Benito-Calvo, A.; Ortega, A.I.; Pérez-González, A.; Campaña, I.; Bermúdez de Castro, J.M.; Carbonell, E. Palaeogeographical reconstruction of the Sierra de Atapuerca Pleistocene sites (Burgos, Spain). Quat. Int. 2017, 433, 379–392. [Google Scholar] [CrossRef]
- Wilson, D.E.; Reeder, D.-A.M. Mammal Species of the World. In A Taxonomic and Geographic Reference; The Johns Hopkins University Press: Baltimore, MD, USA, 2005. [Google Scholar]
- IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 23 February 2021).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. Available online: http://folk.uio.no/ohammer/past/ (accessed on 1 October 2018).
- Cuenca-Bescós, G.; Straus, L.G.; García-Pimienta, J.C.; González Morales, M.R.; López-García, J.M. Late Quaternary small mammal turnover in the Cantabrian Region: The extinction of Pliomys lenki (Rodentia, Mammalia). Quat. Int. 2010, 212, 129–136. [Google Scholar] [CrossRef]
- Galindo-Pellicena, M.A. Estudio de la macrofauna de los niveles holocenos del yacimiento de El Portalón (sierra de Atapuerca, Burgos). Ph.D. Thesis, Department of Paleontology, Complutense University of Madrid, Madrid, Spain, 2014. [Google Scholar]
- Galindo-Pellicena, M.A.; Arsuaga, J.L.; Carretero, J.M. Pig management in the Chalcolithic and Bronze Age periods at the El Portalón site: Integration in the Iberian context. Quat. Int. 2019, 515, 138–149. [Google Scholar] [CrossRef]
- Galindo-Pellicena, M.A.; Arsuaga, J.L.; Pérez-Romero, A.; Iriarte, E.; DeGaspar, I.; Carretero, J.M. Metrical analysis of bovine bone remains from the Neolithic to the Bronze Age at the El Portalón site (Atapuerca, Burgos) in the Iberian context. Quat. Int. 2020, 566–567, 211–213. [Google Scholar] [CrossRef]
- Bañuls-Cardona, S. Human Impact on Small-Mammals from Late Glacial to Late Holocene of Western Mediterranean Region: New Environmental and Climate Approach. Ph.D. Thesis, Università degli Studi di Ferrara, Ferrara, Italy, 2017. [Google Scholar]
- Cuenca-Bescós, G.; García, N. Biostratigraphic succession of the Early and Middle Pleistocene mammal faunas of the Atapuerca cave sites (Burgos, Spain). Cour. Forschunginst. Senckenberg 2007, 259, 99–110. [Google Scholar]
- Rofes, J.; Zuluaga, M.C.; Murelaga, X.; Fernández-Eraso, J.; Bailon, S.; Iriarte, M.J.; Ortega, L.A.; Alonso-Olazabal, A. Palaeoenvironmental reconstruction of the early Neolithic to middle Bronze Age Peña Larga rock shelter (Álava, Spain) from the small mammal record. Quat. Res. 2013, 79, 158–167. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Chalcolithic Cultural Context | Laboratory Code | Con. Age (yr BP) | Age (yr cal BP) | Reference |
|---|---|---|---|---|---|
| Human tibia | Bell-Beaker (UE4) | Beta-269494 | 3900 ± 40 | 4423–4158 | [26] |
| Triticum sp. seed | Pre-Bell Beaker/Stabling (UE209) | Beta-347579 | 4230 ± 30 | 4859–4646 | [26] |
| Triticum sp. seed | Pre-Bell Beaker/Stabling (UE212a) | Beta-347580 | 4280 ± 30 | 4957–4732 | [29] |
| Human adult proximal foot phalange | Pre-Bell Beaker/Funerary (UE79) | Beta-368290 | 4280 ± 30 | 4957–4732 | [34] |
| Human tooth | Pre-Bell Beaker/Funerary (UE80) | Beta-337300 | 4300 ± 30 | 4960–4830 | [34] |
| Bos sp. | Pre-Bell Beaker/Funerary (UE80) | Beta-337299 | 4380 ± 30 | 5042–4860 | [29] |
| Human adult thoracic vertebra | Pre-Bell Beaker/Funerary (UE79) | Beta-368289 | 4400 ± 30 | 5214–4862 | [34] |
| Ovicaprine tooth | Pre-Bell Beaker/Funerary (UE79) | Beta-197389 | 4440 ± 50 | 5285–4874 | [29] |
| Human fragment of a right tibial shaft | Pre-Bell Beaker/Funerary (UE79) | Beta-368285 | 4460 ± 40 | 5294–4886 | [34] |
| Taxa | Fo | Sh | Gr | WI | RA |
|---|---|---|---|---|---|
| Crocidura suaveolens | |||||
| Sorex gr. araneus-coronatus | |||||
| Sorex minutus | |||||
| Neomys fodiens | |||||
| Talpa sp. | |||||
| Miniopterus schreibersii | |||||
| Myotis gr. myotis-blythii | |||||
| Arvicola sapidus | |||||
| Microtus (T.) duodecimcostatus | |||||
| Microtus (T.) lusitanicus | |||||
| Microtus agrestis | |||||
| Microtus arvalis | |||||
| Apodemus gr. sylvaticus-flavicollis | |||||
| Eliomys quercinus | |||||
| Sciurus vulgaris | |||||
| Oryctolagus cuniculus |
| Chrono-Cultural Period | Early Chalcolithic | Late Chalcolithic | ||||||
|---|---|---|---|---|---|---|---|---|
| Context | Funerary | Habitat/Stabling | ||||||
| Floor | Prepared | Activity | Fumier | |||||
| Taxa | NISP | MNI | NISP | MNI | NISP | MNI | NISP | MNI |
| Crocidura suaveolens | 281 | 83 | 31 | 6 | 46 | 11 | 67 | 25 |
| Sorex gr. araneus-coronatus | 326 | 79 | 19 | 3 | 67 | 16 | 104 | 38 |
| Sorex minutus | 4 | 3 | 0 | 0 | 0 | 0 | 2 | 2 |
| Neomys fodiens | 3 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Talpa sp. | 10 | 4 | 0 | 0 | 0 | 0 | 7 | 3 |
| Miniopterus schreibersii | 9 | 5 | 1 | 1 | 4 | 3 | 0 | 0 |
| Myotis gr. myotis-blythii | 27 | 6 | 2 | 1 | 13 | 5 | 5 | 2 |
| Arvicola sapidus | 7 | 4 | 1 | 1 | 0 | 0 | 0 | 1 |
| Pliomys lenki | 0 | 0 | 2 | 1 | 0 | 0 | 0 | 0 |
| Chionomys nivalis | 0 | 0 | 2 | 1 | 0 | 0 | 0 | 0 |
| Microtus (T.) duodecimcostatus | 15 | 10 | 2 | 2 | 2 | 2 | 7 | 6 |
| Microtus (T.) lusitanicus | 49 | 33 | 2 | 1 | 7 | 5 | 14 | 9 |
| Microtus agrestis | 58 | 36 | 9 | 6 | 16 | 11 | 40 | 25 |
| Microtus arvalis | 95 | 54 | 12 | 10 | 14 | 9 | 37 | 24 |
| Apodemus gr. sylvaticus-flavicollis | 555 | 121 | 37 | 9 | 94 | 17 | 143 | 33 |
| Eliomys quercinus | 35 | 10 | 3 | 1 | 7 | 3 | 12 | 7 |
| Sciurus vulgaris | 2 | 2 | 1 | 1 | 1 | 1 | 0 | 0 |
| Oryctolagus cuniculus | 8 | 3 | 6 | 2 | 2 | 2 | 0 | 0 |
| Totals | 1484 | 454 | 130 | 46 | 273 | 85 | 438 | 175 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rofes, J.; Ordiales, A.; Iriarte, E.; Cuenca-Bescós, G.; Galindo-Pellicena, M.Á.; Pérez-Romero, A.; Carretero, J.M.; Arsuaga, J.L. Human Activities, Biostratigraphy and Past Environment Revealed by Small-Mammal Associations at the Chalcolithic Levels of El Portalón de Cueva Mayor (Atapuerca, Spain). Quaternary 2021, 4, 16. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4020016
Rofes J, Ordiales A, Iriarte E, Cuenca-Bescós G, Galindo-Pellicena MÁ, Pérez-Romero A, Carretero JM, Arsuaga JL. Human Activities, Biostratigraphy and Past Environment Revealed by Small-Mammal Associations at the Chalcolithic Levels of El Portalón de Cueva Mayor (Atapuerca, Spain). Quaternary. 2021; 4(2):16. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4020016
Chicago/Turabian StyleRofes, Juan, Amaia Ordiales, Eneko Iriarte, Gloria Cuenca-Bescós, María Ángeles Galindo-Pellicena, Amalia Pérez-Romero, José Miguel Carretero, and Juan Luis Arsuaga. 2021. "Human Activities, Biostratigraphy and Past Environment Revealed by Small-Mammal Associations at the Chalcolithic Levels of El Portalón de Cueva Mayor (Atapuerca, Spain)" Quaternary 4, no. 2: 16. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4020016