
San Josecito Cave and Its Paleoecological Contributions for Quaternary Studies in Mexico


1
Laboratorio de Arqueozoología, Subdirección de Laboratorios y Apoyo Académico, I.N.A.H., Moneda 16, Col. Centro, Ciudad de México 06060, Mexico
2
Museum of Texas Tech University, Lubbock, TX 79409-3191, USA
*
Author to whom correspondence should be addressed.
Quaternary 2021, 4(4), 34; https://0-doi-org.brum.beds.ac.uk/10.3390/quat4040034
Submission received: 3 September 2021
/
Revised: 20 October 2021
/
Accepted: 21 October 2021
/
Published: 26 October 2021
(This article belongs to the Special Issue Taphonomy and Palaeoecology of Quaternary Vertebrates: Advances in Fossil and Experimental Studies)
Abstract
:San Josecito Cave (2250 m elevation) is located nearby Aramberri, Nuevo León, northeastern Mexico, with excavations occurring in 1935–1941 and 1990. It is a paleontological cave and the significance of its faunal data rests in the understanding of the Quaternary ecosystems of the Mexican Plateau and the Southern Plains. This significance is underpinned by a consideration of associated stratigraphic and geochronological data. The fauna is composed of mollusks, amphibians, reptiles, birds, and mammals. More than 30 extinct vertebrate species have been identified, constituting one of the most important Quaternary localities in the Americas. Radiocarbon dates and faunal correlations indicate the excavated deposits represent an interval of time between 45,000 and 11,000 14C years BP. The current synthesis demonstrates that the previous view of the assemblage as a single local fauna is erroneous and that, instead, several successive local faunas are present within a stratigraphic framework. This finding underscores the need for detailed studies of single localities in building paleoenvironmental models. As a corollary, results point to the necessity of including all vertebrate classes represented from a locality in building those models. In addition, the field and analytical methodologies demonstrate the importance of very detailed paleontological excavations, with precise spatial and temporal controls, to assess the taphonomic history of a locality, construct a stratigraphic and geochronological framework, and infer the paleoecological conditions during the time span considered based on the number of local faunas represented. The recognition of San Josecito Cave as an important Late Pleistocene vertebrate paleontological locality is enhanced with the consideration of its faunal data for paleoenvironment reconstruction and possible contribution to Quaternary paleoclimatic modeling.
1. Introduction
Paleoenvironmental research data are a resource that should be managed in a way that provides the entire global-change research community with the tools to find, access, and manipulate quickly the data needed for a particular investigation [1]. The Past Global Changes Project [2] has identified several types of data that need to be given high priority for global change studies. The first priority is the original “raw” or primary paleoclimate, palaeoecological, and palaeoceanographic data, including associated chronological information. These raw data are the foundation for paleoenvironmental research, and the ultimate source of paleoenvironmental information.
Documentation of Quaternary environmental change generally follows two complimentary paths [3]. The first examines proxy records that define the state of climatic and biological systems at various times in the past and provides empirical data necessary for testing and refinement of paleoclimatic models. The second focuses on reconstructing the response of surface systems to climatic and environmental change. This synthesis is concerned primarily with the first pathway.
A fundamental goal of Quaternary vertebrate paleontology is to understand the interrelationships between environmental and faunal changes. Such relationships, implied by the cyclic pattern of change in the Quaternary, are complex and incompletely understood [4]. Since integration of data from individual localities is required, it is critical to assure the best understanding for each of those localities.
Sedimentological analysis of paleoenvironments and the taphonomic study of organic remains preserved in these environments are essential components of paleoecology [5]. The disciplines of sedimentology and taphonomy are necessary for the reconstruction of the original ecological relationships among fossil organisms. Sedimentology provides information on the physical settings where organisms lived and were buried and on interactions between biotic and abiotic components of ancient ecosystems [5].
Taphonomy plays diverse roles in paleobiology including ecological, biogeographical, and evolutionary questions, as well as diagnosing the roles of various taphonomic agents, processes, and circumstances in generating fossil records [6]. Taphonomy provides the methodology to delimit and categorize the conditions and processes leading to the preservation of fossil organisms [7]. This approach illuminates biases that affect paleoecological reconstructions, provides additional information of biological processes represented in a fossil assemblage, and allows the identification of similar (isotaphonomic) assemblages among which ecological comparisons can be made.
During the Late Pleistocene, the North American Southern Plains grasslands appears to have extended from Oklahoma and the Southern High Plains to the Texas Gulf Coastal Plains and into México through the Mexican Plateau. This grassland ecosystem appears to have been a parkland setting with trees primarily restricted to the waterways [8,9,10]. A detailed biotic record in the northern part of the Southern Plains grasslands for the last 40,000 years is available [8,11,12,13,14]. In order to gain a regional perspective, a comparable time-transgressive database for the southernmost biotic section is needed. San Josecito Cave (western flank of the Sierra Madre Oriental; southern Nuevo León, Mexico; Figure 1) is one of two known localities with a great potential for a detailed Late Pleistocene record in this southernmost section [15]. Rancho La Amapola, Cedral, San Luis Potosí, is the other locality (an open-air locality rather than a cave). It has a rich grassland mammal fauna consisting of 7 orders, 18 families, and 33 species [16].
Based on numerous previous taxonomic studies of the cave materials, the San Josecito Cave fauna was considered an example of a disharmonious or non-analog fauna [17,18]. The fauna was composed of elements characterizing different climatic regimes and presumably ecological incompatibles, coexisting nowhere in the world at present [18,19]. The fauna had six new fossil species, comprised of the horned lizard (Phrynosoma josecitensis), big-footed turkey (Meleagis crassipes), extinct thicker-billed parrot (Rhynchopsitta phillipsi), extinct vampire bat (Desmodus stocki), dwarf rabbit (Sylvilagus leonensis), and spectacled bear (Tremarctos floridanus). The fauna also included eight extinct birds and 19 extinct mammals [20,21,22]. This disharmonious biota could have been maintained by equable climates with reduced seasonal extremes that have no modern analogs [23]. No specific study addressed taphonomic or stratigraphic analyses of the San Josecito Cave materials, and only short notes were made on the possible causes of bone accumulation [24,25,26].
This synthesis is the first of its kind to bring together the San Josecito Cave data by focusing on three aspects: (1) a consideration of the overall cave fauna that resulted from Stock’s [26] work; (2) results of the 1990 renewed excavation and associated analyses; and (3) the significance that the composite stratum 720 (i.e., combined stratum 720 and Stock’s 57–58 ft level taxa) faunal data has for contributing to the understanding of the Quaternary ecosystems of the Mexican Plateau, Sierra Madre Oriental, and Southern Plains in general. The 1990 excavation and subsequent analyses undertaken were aimed at: (1) establishing a stratigraphic framework and chronology; (2) determining the taphonomic history of the cave fauna; (3) examining the collection and documentation now housed at the Natural History Museum of Los Angeles County (LACM) from Stock’s [26] excavations; and (4) within the established stratigraphic framework, inferring possible paleoenvironmental scenarios in which the cave faunas could have existed. The 1990 stratigraphically-controlled excavation was conducted by the first two authors [27,28,29,30,31]. A working hypothesis was that no drastic climatic change occurred during the deposition of the studied strata in the mid-Wisconsin period (40,000 to 25,000 14C years BP).
2. Locality Setting
The cave is a single drop, multi-entrance fissure that occurs in folded Late Jurassic or Early Cretaceous limestone [26]. All three now-open natural entrances descend vertically 12 to 30 m to the main room or to a sloping tunnel into the main room (Figure 1). None of the entrances provides easy access today and probably did not in the past; a horizontal or walking entrance probably never existed. This situation implies that the cave acted as a natural pit trap for the larger animals. Smaller animals and birds would have been brought in by raptors or from roosting in the cave. Its use as a den (as proposed by [15,25]) would not have been feasible given the lack of easy access or a walking entrance.
San Josecito Cave consists of only one vertical room tapering to the north, 34 m long and 25 m wide, currently with three vertical shafts. Two of the shafts are high (30 m) above the present cave floor and the third consists of a large upper opening (20 m high) that probably served as the primary trap for the fauna (Figure 1). A separate tunnel descends 41° from the east to a 7 m drop to the floor of the main room. This tunnel entrance is an artificial one opened in the late 1930s to facilitate paleontological excavation. A fourth natural entrance, closed in the late 1930s by local inhabitants, drops about 5 m into the tunnel via a narrow shaft.
A vertical shaft was excavated about 30 m down the south headwall. This shaft was excavated by local people looking for treasure prior to paleontological excavations (Stock’s correspondence, 1935, on file at the LACM). Field observations (George Veni, personal communication, 1989) of the shaft indicated the presence of at least 30 m of fossiliferous sediments below the current excavation levels.
On the northern and western walls, thin outgrowths of a calcareous tufa extend the depth of the cave, probably from the original cave floor. The western portion is formed by a boulder-size outgrowth. Enormous amounts of bones are embedded in a hardened matrix on the tufa.
3. Material and Methods
3.1. Stock’s Excavations
Under the direction of Stock [26], field crews representing the California Institute of Technology quarried a section of San Josecito Cave between 1935 and 1941. The cave floor was divided into five north-to-south blocks (Figure 2). Workers shovel-excavated in 5 ft levels in each block, removing all sediments (primarily dumped outside the cave entrance) and bone within the cave from the upper 12 m. This work created a new cave floor (the present one) that corresponds with Stock’s 55 ft level. At that level, the field crew excavated a test pit in block 2 (southern part of the cave) where they had found the highest concentration of bone. The test pit, now known as Stock’s pit, was a 2 × 2 m unit excavated 4.5 m deep. A smaller unit adjacent to (on the east) and contiguous with the larger unit was 1.8 × 1.3 m and excavated 1.8 m deep [28]. The extent of dry screening or screen mesh size was unknown. No wet screening (i.e., sediments water-washed through one or more nested screen or burlap bags) would have been carried out as that technique to recover microvertebrates primarily was a post-WWII development [32].
3.2. The 1990 Excavation and Associated Research
Five steps were undertaken to obtain the basic data for the 1990 work and subsequent study. The first step was the location and reconnaissance of the cave and the establishment of horizontal and vertical controls [30]. Bringing control into the cave proved difficult due to the great distance through rough terrain between the known reference point (INEGI #3146) and the cave entrance. Nevertheless, the datum established at the cave was reliable as it was obtained through two different methods, field survey instrumentation [33] and GPS calculation. Inside the cave, marks on the walls (left by Stock’s [26] crew) were found that helped tie the elevation data to the provenience data salvaged from Stock’s materials and documentation. The north and west walls of Stock’s pit were cleaned and photographed. The deposits showed a great potential for more than 4.5 m of fossiliferous sediments below the current cave floor [28]. Remnants of the upper strata also were located in various parts of the cave.
The second step was the review of most of the materials and available documentation regarding the cave excavations by Cal Tech personnel in 1935–1941 [29,30]. This review proved helpful in reconstructing the history and methodology of those excavations, including the deficiencies with the excavations. A literature search demonstrated the taxonomic importance of the cave fauna [34,35,36]. Several taphonomic features were found on the bones from the collection that formed the initial analysis categories for the remains recovered in the 1990 excavation.
The third step was the 1990 renewed, stratigraphically-controlled excavation to recover high-resolution data that would permit the paleoenvironmental reconstruction of the ecosystems represented by the cave fauna [31]. Four test units were placed. The two main units were adjacent to Stock’s pit along the north face. A 1 × 0.3 m unit was excavated stratigraphically to stratum 570 at a depth of 1.26 m. A 50 × 50 cm unit was excavated to stratum 680 at a depth of approximately 50 cm. The third unit (30 × 30 cm; excavated 23 cm deep) was placed in an alcove (known as the Bone Alcove; in Stock’s block 5) within upper deposits that corresponded to Stock’s 35 ft level. The last unit (20 × 20 cm; excavated 3 cm deep) was placed in another alcove (known as the Bat Alcove; also in Stock’s block 5) within upper deposits ~0.5 m above the present floor. Every step was recorded on field forms and photodocumented. All sediments were collected and several samples were sent to different specialists for study.
Several cave sediment samples were taken for insects, macrobotanical and pollen remains, and mollusks. Samples for insects, macrobotanical remains, and mollusks were water-processed through nested fine-mesh screens. Pollen samples were processed using appropriate, standard lab techniques. The sparse results were not informative [28].
In order to obtain high-quality data for the taphonomic and paleoecologic analyses, a new approach was used in the excavations. A minimal taphonomic unit (taphonomic box) was defined as a 10 × 10 cm square with thickness dependent on stratum thickness. This approach was based on the knowledge that the stratigraphic units to be encountered were no more than 10 cm thick (generally much less). Most previous paleontological studies lacked the very detailed data necessary to analyze the small vertebrates within a short span of time and space in accordance with their biology. The methods for statistical manipulation were: number of individual specimens (NISP); minimum number of elements (MNE); minimum number of individuals (MNI); and MNI by taphonomic box (BOX).
Orientation data were recorded for 88 bones found in-situ. As all but one were small bones, only direction was noted; dip and strike were obtained only for the one large bone. These data did not prove useful and have not been analyzed further.
Sediments were washed through nested screens (1.41, 0.96, and 0.56 mm) and remaining concentrates sorted for biological materials. These materials were separated into the different vertebrate classes (Amphibia, Reptilia, Aves, and Mammalia) and for mammals, different mammal sizes (small, medium, and large). The vertebrate remains were identified preliminarily.
The fourth step was the detailed analysis of the bones from stratum 720, including the paleoecology of the identified taxa [28]. Due to the enormous amount of microvertebrate material recovered from the concentrates, a single stratum was examined in detail. Stratum 720 was chosen because it had good radiometric control and the largest amount of bone material within the excavated strata based on sorted concentrate samples. Species identifications were pursued only for the cranial materials. Postcranial materials were identified as general groups and data used for taphonomic analyses and comparisons.
The fifth step was the paleoenvironmental reconstruction of composite stratum 720. The MNI and NISP values [37,38], habitat weighting analysis [39], and correspondence canonical analysis (CCA) in different taxonomic groups (small mammals, birds, and reptiles) and taxonomic groups were used.
Habitat weighting has been calculated considering the MNI and NISP since becauseboth measurements provide paleoenvironmental insights of finer resolution than taxonomic presence alone [38]. MNI is at best an ordinal-scale measurement unit. NISP is likely to provide an ordinal scale measure of taxonomic abundance at best. MNI underestimates the true abundance of some taxa, overestimates others, and accurately estimates the abundance of still others. NISP is likely to do the same, although the taxa the abundances of which are overestimated may not be the same as those that are overestimated by MNI [37].
The habitat weighting method consisted of establishing the present-day habitat distribution of each taxon. Each species received a score for each habitat type proportional to habitat preference; the sum of scores for all habitat types for each species is 1. The calculated values of habitat weighting (with presence-absence data) were multiplied by the fossil abundance (NISP and MNI) to add abundance into the analysis [39]. The vegetation types were coniferous forest, oak forest, xeric scrub, grassland, and tropical forest. The analysis was conducted in R using CCA [40] and vegan packages [41].
4. Results
4.1. Sediments
Stock’s [26] (p. 3) observations on the cave sediments were limited. He noted: “The sediments consist for the most part of unconsolidated, grayish cave dirt, generally of a fine consistency. These deposits came from outside the cavern and were either washed in or accumulated through aeolian action. Often thickly interspersed in these beds are fragments of limestone, varying greatly in size, that have fallen from the walls and ceiling.”
Cave sediments from different strata defined during the 1990 work were analyzed. Results [42] pointed to a minimal contribution to the cave sediments from external sources (mammal and bird bones) contrary to Stock’s [26] notation. A great diversity of constituents from inside, including abiotic and biotic sources, were found in the sediments. This diversity included: (1) variable-sized fragments of primary limestone; (2) residue from dissolution of the limestone; (3) authigenically formed calcite-cemented tufa-like materials with varying degrees of porosity; and (4) apatite-rich material likely formed in guano.
4.2. Stratigraphy and Geochronology
While excavated materials from Stock’s [26] work were sent back to California with excavation data, those data are lost at some point in the collection’s history. Stock’s collection now is only taxonomically important, as other studies are hampered by the lack of a complete stratigraphic record. The few available excavation data are meaningless regarding paleoenvironmental reconstruction. This unfortunate occurrence is quite contrary to Stock’s [43,44] concern regarding the importance of recording any available stratigraphic data during paleontological excavations.
Despite that concern, Stock’s descriptions of the cave stratigraphy were limited. He recognized four major units (A, B–C, D, E), each many feet thick (Figure 2). Most bone probably came from his lower three units: B-C (“5–6 feet of fine-grained brown sandstone, definitely stratified”); D (“10–15 feet of limestone breccia”); and E (brown sandstone) [28] (pp. 47–48). Most of Stock’s control marks on the cave wall (indicating the 5 ft levels) were preserved and used in correlations between his work and the current study.
During the 1990 excavation, over 28 stratigraphic units were identified, ranging in thickness from 1 to 17 cm (Figure 3). A number have abrupt boundaries indicating a change in sedimentation while others have irregular or wavy boundaries perhaps indicating some sort of deformation before resumption of deposition.
The first and only radiometric controls for the cave sediments and its faunas have been obtained through radiocarbon assays targeting organic sediments. Radiocarbon dating using bone would not be reliable as shown by the results of the amino acid content assays (T.W. Stafford, personal communication, 1993). Six dates now are available for the cave stratigraphy from six strata within the upper and lowermost parts of the described profile [45]. They are critical to unraveling the history of investigations, Stock’s specimen provenience, the stratigraphy, the paleoecology, and the age of the cave record. The deposits date between 28,000 and 45,000 14C years BP (Table 1). A problem in terms of reversed dates is noted for the lower excavated strata. With respect to the lowermost sample, INAH-940 (Table 1), it cross-cuts several strata and, therefore, represents a mixture of organic sediments from different ages. The incongruous age most likely is due to the long exposure of the profile within Stock’s pit and lack of providing a fresh, uncontaminated exposure for sampling. This type of situation is known to yield falsely young ages. The date is discounted. The reversal between samples SMU-2590 and SMU-2532 (Table 1) are a different matter and which date should be discounted has not been resolved. All ages cited in the text are given as radiocarbon years before present (14C BP). Corresponding calendar years are provided in Table 1.
Based on the radiocarbon ages, the removed sediments from Stock’s excavations above the 55 ft level represent a time span between 25,000 and 11,000 14C years BP, including the last glacial maximum (LGM) and the Pleistocene-Holocene boundary. This timespan corroborates that proposed by Harris [11] for the age of the general San Josecito Cave fauna. Holocene sediments may have been present, but their occurrence is not clear from the available documentation.
Bones recovered from Stock’s pit from the 57 to 58 ft level overlap in age with the materials from stratum 720. In general, the renewed excavation and stratigraphic profile overlaps with Stock’s 55 through 71 ft levels (Figure 3). This overlap is important as it permits a correlation of some of Stock’s materials with the 1990 material and creates more representative local faunas for the period 45,000 to 28,000 14C years BP.
4.3. Fauna
Stock (personal correspondence with Francis D. Bode, 1935–1936) points out that below the 65–70 ft level, large bones are absent. The cave deposits thus can be divided into two sections: an upper section with abundant megafaunal remains; and a lower, tested section (Stock’s pit), apparently lacking large bones but having abundant microfauna. The lower section of San Josecito Cave has a great potential for providing a complete record for the time span it represents (i.e., older than 45,000 14C years BP).
Originally, 53 species of mammals have been identified from Stock’s [26] excavations (Table 2) [29,51]. The overall San Josecito Cave fauna is best represented with materials found within Stock’s 55–60 ft level range that is within Stock’s pit [29]. In the lower deposits, the reduction in number of taxa may indicate a faunal change. Stock (personal correspondence with Harold Sebenius, 1940–1941) notes this reduction as the end of the fossil deposits. The lowermost deposits, however, are the least excavated and bone numbers may be low since because those deposits were sampled less intensively. Furthermore, the main focus during Stock’s excavations was on large animals. The crew collected bones of smaller animals only occasionally (as noted in several letters back and forth between Stock and crew members).
4.4. Mammals
Subsequent analyses of the mammalian remains recovered during the 1990 excavation and those housed at LACM [53,54] brought the total taxa number to 62 (Table 2). Overall, 9 species and 4 genera had not been recorded previously from the cave. Also, six species and one genus (Sorex altoensis, Corynorthinus towsendii, Equus conversidens, Marmota flaviventris, Parasciurus alleni, Peromyscus labecula, and Peromyscus levipes) were reported under other names and now are reassigned to different species or genus [53,55,56,57]. Of these taxa, Equus conversidens, Parasciurus alleni, Peromyscus labecula, and Peromyscus levipes were present in the composite stratum 720 deposits. All the extant species occurred in the region (i.e., Sierra Madre Oriental) where the cave is located, and some even inhabited the cave.
No new extinct taxa were added to the cave mammal fauna, but four extralimital taxa (Sorex milleri, S. altoensis (previously reported as S. saussurei), Marmota flaviventris, and Synaptomys cooperi) were confirmed [53,54]. Of these extralimital taxa, only S. altoensis was present in the stratum 720 deposits. Sorex milleri is a minimally extralimital taxon relative to San Josecito Cave. The cave is less than 100 km south of the southernmost range of S. milleri [58]. Marmota flaviventris and Synaptomys cooperi came from deposits above Stock’s 55 ft level (i.e., from upper deposits younger than stratum 720). These two taxa experienced range changes of at least 1000 km north of the cave [58], with M. flaviventris occurring today in the western US into Canada and S. cooperi in the northern to northeastern US into Canada.
Endemic species from other than stratum 720 consisted of Cryptotis mexicanus, S. milleri, and Sylvilagus leonensis. Based on a morphometric analysis, C. mexicana from San Josecito Cave was significantly larger than the extant form while S. milleri from the cave is similar in size to its extant form [54]. The endemic Sylvilagus leonensis (recovered in deposits above stratum 720) was an extinct dwarf rabbit known only from San Josecito Cave (type locality) and La Presita Cave, San Luis Potosí [59].
Eight hundred eighty-four mammalian cranial elements have been identified to species from stratum 720. The 13 species identified include 6 species new for San Josecito Cave (Table 3). Among the mammals are another two species endemic to Mexico. The endemic species are composed of a shrew (Sorex altoensis) and a white-footed mouse (Peromyscus levipes). Based on a morphometric analysis, S. altoensis from San Josecito Cave is similar in size to its extant form [54]. The modern ranges for the shrew and white-footed mouse are sympatric in the Sierra Madre Oriental, although they do not co-occur in the vicinity of San Josecito Cave. They have restricted and differing ranges in Mexico [58,60,61].
The occurrences of Parasciurus alleni and Peromyscus levipes at San Josecito Cave are the first known Quaternary fossil record for this squirrel [62] and mouse [61]. Both have very limited geographic ranges that are centered on the Sierra Madre Oriental. Sorex altoensis is a minimally extralimital taxon relative to San Josecito Cave. The cave is about 50 km north of the northernmost range of S. altoensis [28].
Based on the overlap of the 1990 stratigraphic profile and Stock’s levels (Figure 3), when the mammalian species composition from stratum 720 is combined with the mammalian fauna from Stock’s 57–58 ft level, then 27 species are recorded from this stratigraphic provenience (Table 3). That number represents 43.5% of the total cave mammalian fauna.
4.5. Birds
Miller [24] reported 43 fossil bird taxa from the more than 2100 specimens recovered from Stock’s [26] excavations (Table 4). He segregated the original bird fauna into three groups. Group 1 were those preyed upon by raptors, composed of gallinaceous birds, parrots, doves, woodpeckers, ducks, shore birds, herons, rails, poor wills, and small passerines. Group 2 were those that nested in the cave mouth, represented by Corvus corax, Coragyps atratus, Bubo virginianus, Tyto alba, Falco mexicanus, and Caracara plancus. Group 3 were those that may have roosted outside the cave and senile or injured individuals that had sought refuge in the cave.
Subsequent examination of the 2100 specimens [29,63,64,65,66] increased the taxa count to 54 for the overall bird fauna (Table 4). Steadman et al. [67], based on 90 bird bones from the 1990 excavation, identified 18 species (6 passerines and 12 non-passerines), with nine new species for the overall cave fauna. The new taxa were of temperate rather than tropical affinities. These new data increased the overall cave avifauna from 54 to 63 species (Table 4) and represents in general one of the largest and most diverse Late Pleistocene avifaunas in North America.
That overall avifauna was dominated by scavengers and predators and reflected three distribution categories. One category was that of birds widespread in both temperate and tropical settings. Another category represented birds restricted to temperate settings while the third category had birds restricted to tropical settings. In terms of broad habitat categories, a montane woodland or forest habitat was the most frequent represented in the overall avifauna, followed very closely by a grassland or grassy savannah habitat [67]. The overall San Josecito Cave bird fauna was a time-transgressive (i.e., not contemporaneous) mixture of species that today prefer coniferous or pine-oak forests/woodlands, grasslands/savannahs, and wetlands [24,67].
The 90 bones from the 1990 excavation yielded 39 identifiable elements to genus or species that came from two distinct periods. Of the 39 identifiable elements, 32 came from the unit in the bone alcove that corresponds to Stock’s 35 ft level (Table 4). One of the taxa likely represented an undescribed extinct stork (Ciconia sp. or Mycteria sp.). Six others were extant birds new to the overall cave avifauna. Most notable were the presence of Scolopax minor (American woodcock) and Aphelocoma coerulescens (scrub jay). Scolopax minor was an extralimital, today ranging 500 to 800 km northeast of the cave and southern Nuevo Leon (winter and breeding ranges) [68] and with preferred habitats of brushy swamps or moist woodlands. Aphelocoma coerulescens would be minimally extralimital, where today they occur in Nuevo Leon only in the westernmost mountains of the Sierra Madre Oriental range in a preferred habitat of pinyon-juniper woodlands [68,69].
The remaining seven identifiable bones come from stratum 720. Despite so few bones, they represent six taxa (Table 5), one of which is new to the overall cave avifauna. Gymnorhinus cyanocephalus (pinyon jay) is another extralimital form, breeding today in the mountains of northern Baja California, with accidentals or non-breeding records known for the northern Sierra Madre Occidental [70,71]. This jay also has a preferred habitat of pinyon-juniper woodlands. In addition, small to medium passerines are represented within the unidentified elements (10 bones).
The inclusion of the birds from Stock’s 57–58 ft level brings the avifauna to 10 taxa plus the small to medium passerines (Table 5). Of interest is the referred Pluvialis cf. squatarola (black-bellied plover), a wading shore bird. In the Americas, this globally occurring bird nests in the Arctic and winters as far south as along the coasts of Argentina and Chile. It primarily is a coastal form in Mexico, where they feed in tidal flats, beaches, salt marshes, and meadows. Winter habitats can include inland shallow water and grassland [72,73]. Sightings today in the Sierra Madre Oriental are rare to accidental occurrences [74] (p. 247). With only one specimen and a referred identification, the significance of this bird’s presence in the composite stratum 720 fauna (fortuitous incorporation of a vagrant or actual range change) is speculative.
4.6. Squamate Reptiles
Brattstrom [75] identified two species of reptiles from Stock’s excavation, that of a new extinct species Phrynosoma josecitensis and the extant species P. orbiculare. Phrynosoma josecitensis (Table 6) was known only from San Josecito Cave and is poorly understood due to its rarity. No remains were recovered from the 1990 excavation and the status of the taxon needs to be verified [76].
Mead et al. [76] recognized three taxa from stratum 720 deposits, all three of which occur in the region today around the cave. These were the endemic Mexican Plateau horned lizard (Phrynosoma orbiculare), spiny lizard (Sceloporus sp.) and the endemic Transvolcanic alligator lizard (Barisia cf. B. imbricata) (Table 6). Subsequently, the subspecies of B. imbricata was elevated to species level [77], and other herpetologists have accepted that change [78]. Bryson and Riddle [79] used mtDNA and phylogenic analysis to examine diversification across the genus and timing. Diversification within B. imbricata and B. ciliaris appeared to be around the Pliocene-Pleistocene border and into the Pleistocene. While not addressing the taxonomic status, they concluded that each of the two taxa seemed to be a composite of three different lineages [79] (p. 387). Unrecognized or cryptic species may be within the B. imbricata complex with further investigation required for clarification. Until that time, the taxa as used here followed Smith et al. [77].
Cruz-Silva [80] identified B. imbricata among the stratum 720 remains as well as the endemic B. ciliaris (Sierra alligator lizard) and additional reptile species, seven of which are new to the overall San Josecito Cave fauna (Table 6). Snakes constituted the majority (n = 5; 71.4%) of the new taxa. Barisia ciliaris, Phrynosoma modestum (roundtail horn lizard), Heterodon simus (Southern hognose snake), Tantilla (blackhead snake), and Storeria (brown snake) were the first records for the Pleistocene in Mexico (Table 6).
These new data increased the overall cave squamate reptiles to 11 species (Table 6). Of the 10 identified squamates from stratum 720, all are extant but three do not occur in the region today. In the Late Pleistocene, the squamate diversity represented in stratum 720 is composed of elements from three different modern biogegraphical regions. Heterodon simus ranges throughout the southeastern USA [81]. Barisia imbricata is from the Neovolcanic Transmexican Belt [82]. Phrynosoma modestum inhabits the Mexican Plateau into western Texas and New Mexico [83]. San Josecito Cave contains a relatively large number of taxa for localities in Mexico. The record is exceeded only by those from Santa Cruz Nuevo in Puebla [84], Rancho La Brisca in Sonora [85], and Loltun Cave in Yucatán [86].
Sometime after 28,000 14C years BP, retraction occurs in the distribution of Heterodon simus from the Sierra Madre Oriental to the southeastern USA. Some mammals, such as Synaptomys cooperi (Southern bog lemming) and Microtus pennsylvanicus (meadow vole) follow similar retraction patterns but to the northern and northeastern USA respectively [21,22]. The timing of the retraction for those rodents, however, is much later than 28,000 14C years BP. On the US Southern Plains, the timing for those two rodents is between 10,000 and 8500 14C years BP [8,14].
Barisia imbricata also shows a retraction in its Late Pleistocene distribution from the Sierra Madre Oriental south to the Mexican Transvolcanic Belt at present. Molecular data for Ambystoma, Crotalus, and Phrynosoma indicate a strong phylogenetic relation between the Mexican Transvolcanic Belt and the Sierra Madre Oriental [87,88,89]. The presence of B. imbricata (endemic to the Transvolcanic Belt) and B. ciliaris in the same geographic area has only been recognized at present in the State of Hidalgo [82,90]. This co-occurrence in the Late Pleistocene of San Josecito Cave indicates that sympatry extended as far as 450 km north to the State of Nuevo León.
Although Phrynosoma modestum is found in the State of Nuevo León, it does not inhabit the pine-oak forest around the cave. It inhabits the open shrub areas in arid and semiarid watersheds [83], and Juniperus forest in the State of Nuevo Leon [91,92]. The distributional pattern linking the Sierra Madre Oriental with the Mexican Plateau represented by P. modestum is reinforced by the presence of P. orbiculare that today occurs primarily on the Mexican Plateau. Phrynosoma orbiculare inhabits semiarid shrubland as well as montane woodlands and forests [83]. This pattern also is repeated by the spiny pocket mouse Liomys irroratus [21,22]. This relationship supports Martin’s [93] proposal that a strong connection existed between the Sierra Madre Oriental and Mexican Plateau during the Late Pleistocene. That suggestion is based on biogeographic analysis of amphibians and reptiles, climate, vegetation types, and historical geology in the Gomez Farias Region. This pattern also is inferred for the genus Sceloporus [94,95].
Shifts in the distribution of taxa appear related to environmental changes associated with changes in global weather. Distributional changes found in studies such as phylogeography demonstrate the expansion and contraction of amphibian and reptile ranges during glacial and interglacials [96,97,98,99]. These studies support the biological dynamism during the Quaternary and indicate that the Pleistocene was not a steady state for herptiles as proposed by Holman [100].
The Late Pleistocene (i.e., stratum 720; ~28,000 14C years BP) vegetation surrounding San Josecito Cave based on the reptile record is a mosaic principally of an open forest with savanna, dominated by Pinus and Quercus. A xeric scrub with Juniperus also is present in the cave area. This reconstruction differs from previous environmental inferences for the cave area based on mammals [29,30,31,101], birds [67], and lizards [76]. Those studies indicate the presence of an oak-pine forest similar to the current setting but within a subhumid climate, more marked winter season, and drier conditions than today. The expanded palaeovegetation reconstruction based on vertebrates is in agreement with the pollen data recovered from the Late Pleistocene in the northern part of Mexico. Those data indicate a Juniperus forest into a xeric scrub was widespread in the Late Pleistocene. A Juniperus forest around the cave is 400 km south from its current distribution [102,103,104,105]. This situation underscores that shifts in vegetation distributions also were occurring in the Late Pleistocene.
4.7. Taphonomy
4.7.1. Stratum 720
The taphonomic data collected focused on refitting, minimum number of individuals (MNI), and bone modification for the stratum 720 small mammalian assemblage. Refitting was accomplished only in part as most postcranial material lacked specific identification. Cranial matching was conducted in terms of teeth-mandible, and tooth-tooth for specimens from the same taphonomic box. The findings were not considered valid for individual quantification as little confidence was placed on some of the refits (a more detailed consideration of age and size is required).
Minimum number of individuals is an import taphonomic feature in quantifying the fauna. Large differences have been found with absolute values (Figure 4), but when dealing with relative values for the results from the different counting methods, no significant differences were found (G-test, F = 49.4; p = 0.20). Nonetheless, as the MNI is used most frequently with cave faunas due to low dispersal of bone remains [106], it is considered here as the most reliable for a study involving many disarticulated bone elements from small vertebrates.
Breakage patterns for the small mammal bones were examined using both cranial and postcranial elements [28] (pp. 129–133). A total of 815 postcranial elements were studied, corresponding to long bones. From those, less than 16% (n = 130) were complete bones, 36.9% (n = 301) proximal sections, 24.5% (n = 200) shafts, and 22.6% (n = 184) distal sections. Identified bones included 173 humeri (21.2%), 165 tibiae (20.2%), 133 femora (16.3%), 129 ulnae (15.8%), 125 pelves (15.3%), 49 scapulae (6.0%), and 41 radii (5.0%). The analysis of the overall breakage patterns showed that the proximal regions of the femur, ulna, scapula, and radius, the distal portions of the humerus and tibia, and the pelvic shaft (partial illium and acetabulum) are usually the less-damaged bone parts.
This pattern, however, is modified when looking at taxonomic groups. The breakage patterns differ from one group to another. Only the ones with a large number of specimens are close to the pattern shown by the overall counts, while those groups with fewer numbers are biased. For example, Eulipotyphla has mostly complete individuals; other groups are composed mainly of shafts or section fragments.
Comparisons with other cave faunas had to rely on the element frequencies and breakage pattern for the overall stratum 720 mammalian assemblage. Those patterns were similar to the ones found for some levels from Gran Dolina, Spain [107]. This similarity indicated that owls most likely were the accumulating agents for the stratum 720 small remains. Further studies using SEM should test this hypothesis by looking at the cortical surface damage [108].
Spatial distribution analysis (Figure 5) of the small mammal bones was pursued to learn the taphonomic agents affecting materials prior to burial, as well as to test the usefulness of the excavation methodology approach (taphonomic boxes). A statistically significant correlation (Mantel statistic, z = 5865.7, p = 0.028) was obtained between the distribution of individuals in taphonomic boxes and the spatial position of such boxes. Such correlation not only was derived due to physical causes (gravity), but other abiotic and biotic factors that may have affected the final position of the bones. Biotic factors may have included trampling and bone transportation by other small mammals. Those processes, however, were not recorded clearly on the bones themselves.
The overall taphonomic analysis for stratum 720 indicates that small mammal bone accumulation most likely was due primarily to owls. After deposition of the pellets, bones from the pellets may have been displaced either by gravity or moved by woodrats or other agents to their final position. Breakage seems to correspond to the normal effect while animals are contained in an owl’s gizzard [108].
4.7.2. Stratum 770
Four of the identified strata from the 1990 excavation contained blue and gray colored bone. The diagenetic processes causing the colorations were investigated with bone from stratum 770 [101,109,110]. Results from energy-dispersive X-ray (EDAX) fluorescence and chemical analyses by instrumental neutron activation (INAA) indicated the presence of various trace elements including copper (Cu). Robles et al. [109] (p. 128) concluded that the color was due to impurities within the transition trace metal ions and most likely by copper. The source of the copper, however, was unknown.
Further research involved structural and elemental analyses and spectroscopic investigations. The spectroscopic data were the most informative for understanding the blue and gray colorations. The data revealed a heat-induced modification (apatite crystals of different shapes at least two orders of magnitude larger than those from modern bone) and traces of manganese (Mn) within the typical bone constituents. Different cations of manganese formed by the heat treatment were responsible for the two colorations. For the blue bones, heating took place under oxidizing conditions. Temperature range for color formation was estimated to have been above 500 °C and below 600 °C. The source of the heat, however, was not identified [110].
4.8. Paleoecology
Many studies have explored the relationship between ecological function and species diversity [111], including faunal comparisons and a community structural analysis (methods for describing the evenness in relative abundance among species). Two approaches were used to examine paleoecological influences based on the stratum 720 mammalian assemblage (Table 3). Measures of species number and the distribution of their relative abundances do not require taxonomic similarity as does the taxonomic uniformitarian approach. Dominance-diversity measures, however, do not convey anything about the kinds of ecological roles occupied by species or the interactions between them [112].
4.8.1. Faunal Comparisons
Twenty-one mammal faunas chosen for comparisons with the composite stratum 720 mammals from San Josecito Cave encompassed an approximate range of ages from 21,500 to 38,700 14C years BP and ee values from 3648 through 5540 (these values were available only for nine sites). For most of these localities, specific fauna for a dated stratum was not available. Comparisons were made based on the overall fauna at each locality.
The dendrogram obtained from the cluster analysis (Figure 6) shows that the San Josecito Cave composite stratum 720 mammalian fauna broadly groups with the faunas from southwestern New Mexico (Dry and U-bar caves), Arizona (Rampart Cave), and Nevada (Smith Creek Cave, Nevada). The age for San Josecito Cave stratum 720 (~28,000 14C years BP) is within the age ranges provided for the New Mexican [113] (25,160–33,590 14C years BP) and Arizonan [114,115] (23,540–36,200 14C years BP) caves, and close to the age of the Nevadan cave fauna [116] (28,650 14C years BP). Based on a re-evaluation of the arvicoline rodents, however, the age of the Smith Creek Cave fauna (Nevada) appears much older (from 820,000 to 146,000) than the original age and considerably older than stratum 720 [117].
The Rampart Cave fauna has a very low number of species (n = 7). Two of those species are shared with composite stratum 720 at San Josecito Cave, i.e., the Pleistocene vampire bat and the mountain lion. The sharing of one of the few tropical elements (the vampire bat) may be important in grouping these two caves. If so, the locality grouping is not only based on number of species, but also on the actual species represented.
The New Mexican and Nevadan caves are the localities with the most abundant number of species. The number of shared taxa with San Josecito Cave composite stratum 720, however, is not the same. Dry Cave (New Mexico) and Smith Creek Cave (Nevada) only share five taxa each (mostly carnivores) with composite stratum 720, while U-Bar Cave (New Mexico) shares 12 taxa (bats, carnivores, horses, ungulates, and rodents).
Harris [11] concluded that the U-Bar Cave fauna indicated more equitable conditions than today. Absence of high temperatures together with greater effective moisture during the spring allowed the occurrence of now-distant plants and animals favored by cool summers and moderate year-round moisture. Although this scenario could have occurred around San Josecito Cave, it was not very likely as most of the extant mammal taxa found in stratum 720 occur in the region today but not necessarily in the vicinity of the cave.
4.8.2. Community Structural Analysis
Food habits and habitat preference are the two measures used in the community structural analysis for the composite stratum 720 mammalian assemblage. Food habits provide an understanding of the different environments contributing certain species, and an explanation for bone accumulation in the cave [118]. Affecting the utility of this measure is the lack of knowledge of the detailed food habits of some recent species, and even more so for the fossil ones. Following a methodological approach proposed by Mares and Willig [119], determining habitat preference is based on the general vegetational associations proposed by Barbour and Billings [120]. Data for the different species are easy to obtain for the general associations based on the literature, but a specific association was precluded due to the lack of knowledge of habitat preferences for several extant as well as extinct species.
4.8.3. Food Habits
A total of 1261 specimens represent 27 species. Of the specimens, 976 individuals (77.4%) pertain to the herbivore niche, 201 are carnivores (15.9%), 69 insectivores (5.5%), 12 pollen-eaters (1.0%), 2 omnivores (0.2%), and one blood-eater (0.1%). In terms of food habits by species, percentages are: herbivores (50%), carnivores (26.7%), insectivore (10%), omnivores (6.7%), pollenophage (3.3%), and sanguinivore (3.3%). In comparison, the modern mammalian fauna collected in the area of San Josecito Cave in 1990 is composed of (based on number of taxa in Table 4 of [26] (pp. 57–59): 27 herbivores (45%), 18 insectivores (30%), 9 carnivores (15%), 3 omnivores (5%), 2 pollenophages (3.3%), and one sanguinivore (1.7%). Proportions in the fossil assemblage seem to deviate toward a larger number of carnivores and a smaller number of insectivores than in the modern mammalian fauna.
When comparing the percentage of carnivores present in the composite stratum 720 mammalian assemblage (Table 3) with those provided by White et al. [118] for several North American caves, the San Josecito Cave percentage is greater than the ones for most of those caves. Jaguar, Moonshiner, and Middle Butte in Idaho, however, have higher percentages of carnivores. White et al. [118] proposed two different hypotheses for those high percentages, and those hypotheses are used to evaluate the findings in San Josecito Cave.
One explanation is that San Josecito Cave could have been used as a denning site (as proposed by [15]). A carnivore denning site, however, implies a number of chewed bones, as well as young carnivore remains [118]. Neither is the case for the stratum 720 carnivore assemblage.
The San Josecito Cave carnivore percentage is not as high and disproportionate as those for the Idaho caves [118] that were considered carnivore traps. The great number of large mammals, including carnivores, collected in the overall deposits from San Josecito Cave may suggest that the cave was functioning as a trap throughout the Late Pleistocene. Carnivores may well have gone inside to scavenge the available meat from herbivores that fell down and themselves became entrapped [121]. A similar scenario has been proposed for Rancho La Brea in the Los Angeles basin (southern California) with an assemblage dominated by carnivores and herbivores [122,123]. The available evidence (physiography and taphonomy) indicates the cave functioned as a pit trap in general and the carnivores were present due to entrapment.
4.8.4. Habitat Preferences
The habitat preferences for extant and extinct taxa have been examined. For extinct taxa, past preferences have been inferred from the available literature based on modern analogs and isotope studies. All but one of the species with available data (21) occur in the Rocky Mountain biome. That biome is composed of several kinds of vegetational associations. Although the white-throated wood rat, Neotoma albigula, is not considered within that biome, it is known to occupy the slopes of the hills where the cave is located [124]. Other vegetation regimes where most taxa could occur are: intermountain deserts and woodlands (11 species), deserts (9 species), chaparral (8 species), and grassland (8 species).
Several species are stenotypic (adapted to a narrow range of environmental conditions) and these are the most valuable as paleoenvironmental indicators [119]. Three extant species (Parasciurus alleni, Peromyscus difficilis, P. levipes) presently are known to occur only in the Rocky Mountain forests [125,126]. Three extinct taxa (Desmodus stocki, Navahoceros fricki, and Euceratherium) are thought to occur only in such a vegetational association [15,127]. In addition, two extant species (Sorex altoensis and Microtus mexicanus) occupy only two biomes, that of the Rocky Mountains and Alpine forests [125,128].
While the results of the mammal analysis point to the Rocky Mountain biome as being present around 28,000 14C years BP, the specific vegetational association most likely is not the same as today for the San Josecito Cave area. In order to define more clearly the type of vegetational community that existed, a paleoenvironmental analysis has been undertaken to include all composite stratum 720 vertebrate taxa.
4.8.5. Paleoenvironmental Reconstruction
Several taxa provide additional environmental indications. The extinct Pleistocene vampire bat (Desmodus stocki) is considered to have required a mild climate with winter temperatures warmer than at present [129] and the genus Desmodus is considered to be highly sensitive to environmental temperature [130]. The association of the extinct vampire bat and the Shasta ground sloth (Nothrotheriops shastensis) in the composite stratum 720 at 2250 masl suggests a similar climatic restriction for both species associated with winter temperatures [131]. The presence of the Virginia rail (Rallus limicola) and black-bellied plover (Pluvialis squatarola) indicate a wetlands habitat (freshwater to brackish marshes) and grassland not available in the vicinity of the cave today [72,73,132].
The presence of two Barisia species (Alligator lizards) in the same geographic area is uncommon. Today, B. ciliaris and B. imbricata are sympatric only in Atotonilco el Grande, northern State of Hidalgo [82,90]. Assuming niche conservatism, climatic conditions around 28,000 14C years BP for the San Josecito Cave area would have been similar to that 450 km north. Data from 1951 to 2010 in the Atotonilco el Grande weather station show an average annual temperature of 16.3 °C and an annual precipitation of 787.3 mm (smn.cna.gob.mx). In contrast, San Francisco (the closest weather station to the cave area) has an annual average temperature of 17.3 °C and annual precipitation of 319.9 mm (smn.cna.gob.mx). During the Late Pleistocene, then, the annual average temperature in the cave area may have been 1 °C lower and the annual precipitation as much as 467.4 mm greater than today. Such an increase in annual precipitation would support wetlands in the area as indicated by the Virginia rail and black-bellied plover.
No differences occurred in the results using the habitat weighting method for composite stratum 720 with only the presence of the organism in different habitats or using the presence and the relative abundance (MNI or NISP) of organisms. The small mammals (Table 3) indicated a coniferous forest. The birds (Table 5) indicated a grassland. The squamate reptiles (Table 6) further indicated a coniferous forest (Table 7, Table 8 and Table 9). All vertebrates indicated a combined coniferous and oak forest and grassland at ~28,000 14C years BP for the San Josecito Cave area.
The canonical correlations analysis (CCA) compared the correlation between the presence of the small mammals, birds, squamate reptiles, and all vertebrates in different vegetation types, and the habitat weighting values with relative abundance or only presence. The correlation was regularized and the values of the cross validation (CV-scores) is high in all cases.
The CCA for small mammals infers a coniferous-oak forest (Figure 7), where species such as Sorex altoensis, Parasciururs alleni, Peromyscus difícilis, and Neotoma albigula support high cross correlations values. The CCA for birds indicates that the habitat weighting analysis infer a grassland supported by Sturnella and P. squatarola (Figure 7). The CCA for squamate reptiles infers a coniferous-oak forest (Figure 7), where taxa such as Hypsiglena, Barisia imbricata, B. ciliaris, Phrynosoma orbiculare, and Storeria support high cross correlation values. The CCA for all vertebrates indicates a coniferous-oak forest (Figure 7).
The habitat weighting and CCA analyses, with different groups or all vertebrates, indicate in common the presence of coniferous-oak forest in the vicinity of San Josecito Cave at ~28,000 14C years BP. This result is different than the oak-pine forest present today. The mean annual temperature would have been colder, at least by two degrees in the past [133]. The presence of Barisia imbricata and B. ciliaris in sympatry during this time in San Josecito Cave mirrors their current sympatry at Atotonilco el Grande, Hidalgo [82]. This location has colder and more humid conditions than those of the San Josecito Cave area today.
5. Discussion
Results from the taphonomic studies of the 1990 strata 720 and 770 materials provide insights into the accumulation of the small vertebrates and the cave dynamics. The bone breakage patterns for the stratum 720 vertebrates indicate that owls most likely were the accumulating agents. Three species of owls occur in the composite stratum 720 fauna (Table 5). All three have a varied diet focused on small mammals (e.g., shrews, microtines, mice, bats), birds (particularly small passerines), snakes, and other small vertebrates and insects. This range of prey vertebrates is mirrored in the stratum 720 fauna. The most frequent remains are of Asio otus (long-eared owl) [67]. These owls generally hunt within their home range which can be up to 980 ha (~2422 ac). Home range use changes through the year, with the smallest use area being during the breeding season [134,135,136]. Assuming this behavior was occurring in the Late Pleistocene, home range use by this owl provides a view of the small vertebrate composition within a fairly localized area around San Josecito Cave.
The taphonomic boxes provided an enhanced estimate of abundance and MNI for the stratum 720 fauna. Bones were not distributed randomly across the 720 surfaces. Spatial distribution by taphonomic box indicted a differential distribution of bones across an excavation unit based on the abundance of elements in each box. The MNI by taphonomic box for the stratum 720 mammals (Table 10) increased almost three times from a general count for the overall stratum. The most abundant mammals were a vole (Microtus mexicanus) and mouse (Reithrodontomy megalotis) with the vole more than three times as abundant as the mouse. The accuracy of the individual counts was dependent upon the unit size used [137,138].
Bone discoloration that produced blue and gray bones was due to at least two factors, that of heat and of environmentally available manganese. These physical and chemical processes were occurring within the cave rather than from outside conditions. The heat-induced modifications were caused by an unknown source in the cave (combustible guano deposits may be a likely source) that reached temperatures above 500 °C but below 600 °C. Two different cations of manganese were responsible for the gray and blue colors. Although the cause of the heating events was unknown, these incidents occurred at least four times over a 10,000-year period (from ~35,000 to 25,000 14C years BP).
The squamate reptiles indicate a strong relationship between the Sierra Madre Oriental and Mexican Transvolcanic Belt regions during the Late Pleistocene. Sympatric taxa found in San Josecito Cave at ~28,000 14C years BP now range in one or the other region. The retraction pattern is to the south, whereas other taxa retreated north. The relationship between the Sierra Madre Oriental and the Mexican Transvolcanic Belt regions has not yet been examined for mammals experiencing range changes by the end of the Pleistocene [21,22]. In addition, future climatic studies with the taxa found in the composite stratum 720 local fauna may provide more information on the paleoclimatic conditions in the San Josecito Cave area and Sierra Madre Oriental that allowed co-existence of the taxa and community structure development around 28,000 14C years BP.
The overall fauna of San Josecito Cave now comprises 136 species and prevails as one of the most important Late Pleistocene faunas for North America. The great possibility exists of more unreported taxa within the other strata from the 1990 excavation as well as the non-cataloged materials housed at LACM. Two points stand out. First, paleontological excavations must be undertaken within a field methodology that allows for a very detailed recovery and data recording, with precise spatial (horizontal and vertical) and radiometric controls. Second, general paleoenvironmental conditions can be inferred following the approaches used here. For more detailed results, however, that lead to a better understanding of the Quaternary ecosystems and biotic communities, a greater knowledge of the biology of the recent species and their ecomorphological traits is needed.
The question is raised about the concept of a cave fauna as a single local fauna. Other strata have very different species composition. The term fauna seems better applied to a group of organisms sharing a suite of common species and representing its maximum geographic and temporal limits [139] such as the Late Pleistocene fauna of Mexico. On the other hand, local fauna refers to aggregates of species that have a defined distribution in time and space, based on the record from a restricted geographic area.
The term local fauna could be applied to the overall San Josecito Cave assemblage if all of the materials came from a single stratigraphic level. Stock’s [26] excavations, however, crossed several stratigraphic levels that encompassed many thousands of years. Local faunas should be defined according to stratigraphic provenience and geochronology [8,12,13,14]. Most of Stock’s materials cannot be allocated to a specific local fauna due to their lack of provenience data. The overall vertebrate assemblage reflects many, successive local faunas. Currently, then, a San Josecito Cave local fauna refers only to the composite stratum 720 and Stock’s 57–58 ft level. That local fauna is composed of 47 taxa (Table 11), and dates at ~28,000 14C year BP.
It is on these grounds that it is considered both inappropriate and inadequate to treat the San Josecito Cave fauna as a single entity representing a disharmonious (i.e., non-analog) fauna [17]. The lack of exact provenience of most of Stock’s collection does not allow ascertaining the relationships of the elements from different biomes within one or several temporal communities. The disharmony in the composite stratum 720 local fauna is limited. The level of disharmony within the local fauna is variable, with most non-analog species being minimally extralimital. A bird (Gymnorhinus cyanocephalus) is extralimital by several hundred km. A snake (Heterodon simus) has the most extensive extralimital distribution. Without knowing the provenience of most of the non-analog elements within the overall cave fauna, the trajectory of disharmony is difficult to track. What can be said is that a minimal non-analog fauna existed at ~28,000 14C years BP. Whether the composite stratum 720 is the first non-analog local fauna for the San Josecito Cave record is unknown until the vertebrate remains from the earlier strata are identified and evaluated. An important point, however, is that the non-analog nature of a fauna should be evaluated on all the vertebrate classes represented rather than just one class (typically mammals). Each faunal assemblage needs to be defined not just in terms of faunal composition, but also in terms of the stratigraphic position of the fauna and associated radiometric data. This situation points to the need of stratigraphically-controlled excavations to access and evaluate the actual components of a locality’s overall fauna.
A shortcoming of the community structure analysis is the level of knowledge available for specific relationships between the ecological parameters and the morphological traits. Nevertheless, the approaches taken reveal different paleoenvironmental aspects. First, the Rocky Mountains biome is the general biome for the cave local fauna and regionally at ~28,000 14C years BP similar to that of today. The difference is in the specific vegetational association that appears distinct from today. Second, the vegetational association is a mosaic of coniferous-oak forest and grasslands (different from the pine-oak forest of today). This association in and of itself is a non-analog one. It is not represented in any of the current vegetational associations within the Rocky Mountains biome.
6. Conclusions
The current synthesis examined a paleontological cave deposit and the significance of its faunal data for contributing to the interpretation of Quaternary ecosystems. The hypothesis was that no drastic climatic change occurred during the deposition of the studied strata at the mid-Wisconsin period (40,000 to 25,000 14C years BP). This hypothesis was tested by comparing the independently-derived data sets from the faunal and geochronological results focusing on stratum 720. The focus on one unique stratum decreased the general inferences that could be made regarding the overall late Quaternary ecosystems of northeastern Mexico. This approach, however, assured both the best knowledge of the San Josecito Cave local fauna and the most supported paleoenvironmental and paleoclimatic reconstruction.
Although the hypothesis of no change originally proved valid based only on mammals [28], the broader and more recent analyses have demonstrated the hypothesis to be false. In general, the Rocky Mountains biome is represented in the vicinity of the cave around 28,000 14C years BP as it is today. But, the vegetational association is a non-analog mosaic of coniferous-oak forest and grasslands. The current interpretation points to the importance of detailed paleoecological studies of all vertebrate taxa for each locality as well as within a stratigraphic and geochronological framework.
The continued study of the San Josecito Cave faunal assemblages is a building block toward the understanding of Late Pleistocene environments, climate, and faunal communities on the Mexican Plateau and the Southern Plains in general. The great potential of the sediments for a detailed fossil record for the mid-Wisconsin period exemplified by this synthesis indicates the importance of such research. The San Josecito Cave composite stratum 720 local fauna provides the first inferences of the paleoecology and paleoenvironment at the mid-Wisconsin of northeastern Mexico. Non-analog faunas are beginning to appear by at least ~28,000 14C years BP in the cave record. Equally significant, the local fauna indicates a non-analog vegetational community.
This synthesis of San Josecito Cave and its faunal data has demonstrated that the previous view of the assemblage as a single local fauna is erroneous and instead, several successive local faunas are present within a stratigraphic framework. The overall San Josecito Cave fauna (sensu [17,18]) is a time-transgressive one covering many thousands of years. Treating it as a single fauna masks changes within the faunal community, its paleoecology, and climatic conditions that may have existed.
The study also underscores the need for detailed studies of single localities in building paleoenvironmental models. Furthermore, the recognition of San Josecito Cave as an important Late Pleistocene vertebrate paleontological locality is enhanced with the consideration of its faunal data for paleoenvironment reconstruction and possible contribution to Quaternary paleoclimatic modeling. In addition, the field and analytical methodologies demonstrate the importance of very detailed paleontological excavations, with precise spatial and temporal controls, to assess the taphonomic history of a locality and infer the paleoecological conditions during the time span considered.
Author Contributions
Conceptualization, J.A.-C. and E.J.; methodology, J.A.-C. and E.J.; validation, E.J.; formal analysis, J.A.-C., E.J. and J.A.C.; investigation, J.A.-C. and E.J.; resources, J.A.-C. and E.J.; data curation, J.A.-C.; writing—original draft preparation, J.A.-C. and E.J.; writing—review and editing, J.A.-C., E.J. and J.A.C.; project administration, J.A.-C. and E.J.; funding acquisition, J.A.-C. and E.J. All authors have read and agreed to the published version of the manuscript.
Funding
External funding was received from the National Geographic Society (E.J.), National Speleological Society (E.J. and J.A.-C.), American Society of Mammologists (J.A.-C.), Cave Research Foundation (J.A.-C.), Geological Society of America (J.A.-C.), and the American Museum of Natural History Theodore Roosevelt Memorial Fund (J.A.-C.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are housed at the Archaeozoology Lab, Instituto Nacional de Antropología e Historía, Mexico City, Archaeozoology Lab, National Institute of Anthropology and History, Mexico City, and available on request to Joaquín Arroyo-Cabrales.
Acknowledgments
This manuscript represents part of the ongoing Lubbock Lake Landmark regional research program on the Quaternary paleoecology and taphonomy of the Southern Plains.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nolan, C.; Overpeck, J.T.; Allen, J.R.M.; Anderson, P.M.; Betancourt, J.L.; Binney, H.A.; Brewer, S.; Bush, M.B.; Chase, B.M.; Cheddadi, R.; et al. Past and future global transformation of terrestrial ecosystems under climate change. Science 2018, 361, 920–923. [Google Scholar] [CrossRef] [Green Version]
- Past Global Changes Project. Proposed implementation plans for research activities. IGBP Glob. Chang. Rep. 1992, 19, 1–110. [Google Scholar]
- Blum, M.D.; Toomey, R.S., III; Valastro, S., Jr. Fluvial response to late Quaternary climatic and environmental change, Edwards Plateau, Texas. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994, 108, 1–21. [Google Scholar] [CrossRef]
- Werdelin, L.; von Koenigswald, W. Climate and fauna: A database of the Quaternary mammals of Europe. Eur. Mam. Newsl. 1994, 1, 9–15. [Google Scholar]
- Behrensmeyer, A.K.; Hook, R.W. Paleoenvironmental contexts and taphonomic modes. In Terrestrial Ecosystems through Time. Evolutionary Paleoecology of Terrestrial Plants and Animals; Behrensmeyer, A.K., Damuth, J.D., DiMichele, W.A., Potts, R., Sues, H.-D., Wing, S.L., Eds.; The University of Chicago Press: Chicago, IL, USA, 1992; pp. 15–136. [Google Scholar]
- Behrensmeyer, A.K.; Kidwell, S.M.; Gastaldo, R.A. Taphonomy and paleobiology. Paleobiology 2000, 26, 103–147. [Google Scholar] [CrossRef]
- Martin, R.E. Taphonomy: A Process Approach; Cambridge University Press: Cambridge, MA, USA, 1999; p. 508. [Google Scholar]
- Johnson, E. Lubbock Lake: Late Quaternary Studies on the Southern High Plains; Texas A&M University Press: College Station, TX, USA, 1987; pp. xii + 179. [Google Scholar]
- Pérez-Crespo, V.A.; Arroyo-Cabrales, J.; Benammi, M.; Johnson, E.; Polaco, O.J.; Santos-Moreno, A.; Morales-Puente, P.; Cienfuegos-Alvarado, E. Geographic Variation of Diet and Habitat of the Mexican Populations of Columbian Mammoth (Mammuthus columbi). Quat. Int. 2012, 276–277, 8–16. [Google Scholar] [CrossRef]
- Pérez-Crespo, V.A.; Prado, J.L.; Alberdi, M.T.; Arroyo-Cabrales, J.; Johnson, E. Diet and Habitat for Six American Pleistocene Proboscidean Species Using Carbon and Oxygen Stable Isotopes. Ameghiniana 2016, 53, 39–51. [Google Scholar] [CrossRef]
- Harris, A.H. Wisconsin age environments in the northern Chihuahuan Desert: Evidence from the higher vertebrates. In Transactions of the Symposium on the Biological Resources of the Chihuahuan Desert Region; National Park Service Transactions and Proceedings Series; U.S. Department of the Interior: Chihuahua, CH, USA, 1977; Volume 3, pp. 23–52. [Google Scholar]
- Johnson, E. Late Pleistocene and early Holocene vertebrates and paleoenvironments on the southern High Plains (USA). Geogr. Phys. Quat. 1986, 40, 249–261. [Google Scholar]
- Johnson, J.D.; Wilson, L.D.; Mata-Silva, V.; García-Padilla, E.; DeSantis, D.L. The Endemic Herpetofauna of Mexico: Organisms of Global Significance in Severe Peril. Mesoam. Herpetol. 2017, 4, 544–620. [Google Scholar]
- Moretti, J.A. The Vertebrate Fauna of Macy Locality 100: Exploring Late Pleistocene Community Composition in Non-Analog North America. Master’s Thesis, Texas Tech University, Lubbock, TX, USA, 2018. [Google Scholar]
- Kurten, B.; Anderson, E. Pleistocene Mammals of North America; Columbia University Press: New York, NY, USA, 1980; pp. xvii + 442. [Google Scholar]
- Álvarez, T.; Ocaña, M.A.; Arroyo-Cabrales, J. Restos de mamífero. In Rancho “La Amapola”, Cedral. Un Sitio Arqueológico-Paleontológico Pleistocénico-Holocénico con Restos de Actividad Humana; México, D.F., Mirambell, L.E., Eds.; Colección Interdisciplinaria-Serie Memorias; INAH: México City, Mexico, 2012; pp. 147–194. [Google Scholar]
- Graham, R.W. Response of mammalian communities to environmental changes during the late Quaternary. In Community Ecology; Diamond, J., Case, T.J., Eds.; Harper & Row: New York, NY, USA, 1986; pp. 300–313. [Google Scholar]
- Graham, R.W.; Lundelius, E.L., Jr. Coevolutionary disequilibrium and Pleistocene extinctions. In Quaternary Extinctions; Martin, P.S., Klein, R.G., Eds.; University of Arizona Press: Tucson, AZ, USA, 1984; pp. 223–249. [Google Scholar]
- Semken, H.A., Jr. Micromammal distribution and migration during the Holocene I. In 3rd Annual Meeting Abstracts; Knox, J.C., Mickelson, D.M., Eds.; American Quaternary Association: Madison, WI, USA, 1974; p. 25. [Google Scholar]
- Arroyo-Cabrales, J.; Johnson, E. Catálogo de los ejemplares tipo procedentes de la Cueva de San Josecito, Nuevo León, México. Rev. Mex. Cienc. Geol. 2003, 20, 79–93. [Google Scholar]
- Ceballos, G.; Arroyo-Cabrales, J.; Ponce, E. Effects of Pleistocene environmental changes on the distribution and community structure of the mammalian fauna of Mexico. Quat. Res. 2010, 73, 464–473. [Google Scholar] [CrossRef]
- Ferrusquía-Villafranca, I.; Arroyo-Cabrales, J.; Martínez-Hernández, E.; Gama-Castro, J.; Ruíz-González, J.; Polaco, O.J.; Johnson, E. Pleistocene mammals of Mexico: A critical review of regional chronofaunas, climate change response and biogeographic provinciality. Quat. Int. 2010, 217, 53–104. [Google Scholar] [CrossRef]
- Graham, R.W.; Mead, J.I. Environmental fluctuations and evolution of mammalian faunas during the last deglaciation in North America. In The Geology of North America: North America and Adjacent Oceans during the Last Deglaciation; Geological Society of America: Boulder, CO, USA, 1987; Volume K-3, pp. 371–402. [Google Scholar]
- Miller, L. The Pleistocene Birds of San Josecito Cavern, Mexico. Univ. Calif. Publ. Zool. 1943, 47, 143–167. [Google Scholar]
- Naples, V.L. Morphological changes in the facial region and a model of dental growth and wear pattern in Nothrotheriops shastensis. J. Vertebr. Paleontol. 1990, 10, 372–389. [Google Scholar] [CrossRef]
- Stock, C. The Cave of San Josecito, Mexico. Calif. Inst. Technol. Balch Grad. Sch. Geol. Sci. Contrib. 1943, 361, 1–5. [Google Scholar]
- Arroyo-Cabrales, J. New Studies on San Josecito Cave, Nuevo Leon, Mexico. Cave Res. Found. Rep. 1991, 1990, 43–45. [Google Scholar]
- Arroyo-Cabrales, J. Taphonomy and Paleoecology of San Josecito Cave, Nuevo León, México. Ph.D. Thesis, Department of Biological Sciences, Texas Tech University, Lubbock, TX, USA, 1994. [Google Scholar]
- Arroyo-Cabrales, J.; Johnson, E. Arroyo-Cabrales, J.; Johnson, E. A Reappraisal of Fossil Vertebrates From San Josecito Cave, Nuevo Leon, Mexico. In Ancient Peoples and Landscapes; Johnson, E., Ed.; Museum of Texas Tech University: Lubbock, TX, USA, 1995; pp. 217–231. [Google Scholar]
- Arroyo-Cabrales, J.; Johnson, E.; Ralph, R.W. An initial reappraisal of San Josecito Cave and Its Late Pleistocene fauna. Curr. Res. Pleistocene 1989, 6, 63–65. [Google Scholar]
- Arroyo-Cabrales, J.; Johnson, E.; Ralph, R.W. New excavations at San Josecito Cave, Nuevo León, México. Curr. Res. Pleistocene 1993, 10, 91–94. [Google Scholar]
- Hibbard, C.W. Techniques of Collecting Microvertebrate Fossils. Univ. Mich. Contrib. Mus. Paleontol. 1949, 8, 7–19. [Google Scholar]
- Ralph, R.W.; Arroyo-Cabrales, J.; Johnson, E. San Josecito Cave. Recent Explorations into the Past. NSS News 1994, 52, 230–235. [Google Scholar]
- Jones, J.K., Jr. Pleistocene bats from San Josecito Cave, Nuevo León, México. Univ. Kans. Mus. Nat. Hist. 1958, 9, 389–396. [Google Scholar]
- Russell, R.J. Pleistocene pocket gophers from San Josecito Cave, Nuevo León, México. Kans. Mus. Nat. Hist. 1960, 9, 539–548. [Google Scholar]
- Webb, S.D. A cranium of Navahoceros and its phylogenetic place among New World Cervidae. Ann. Zool. Fenn. 1992, 28, 401–410. [Google Scholar]
- Lyman, R.L. Quantitative Paleozoology; Cambridge University Press: New York, NY, USA, 2008; p. 348. [Google Scholar]
- Faith, J.T.; Lyman, R.L. Paleozoology and Paleoenvironments: Fundamentals, Assumptions, Techniques; Cambridge University Press: New York, NY, USA, 2019; p. 398. [Google Scholar]
- Blain, H.A.; Bailon, S.; Cuenca-Bescós, G. The Early–Middle Pleistocene palaeoenvironmental change based on the squamate reptile and amphibian proxies at the Gran Dolina site, Atapuerca, Spain. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 261, 177–192. [Google Scholar] [CrossRef]
- González, I.; Déjean, S.; Martin, P.; Baccini, A. CCA: An R package to extend canonical correlation analysis. J. Stat. Softw. 2008, 23, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package Vegan: Community Ecology Package. 2016. Available online: https://CRAN.R-project.org/package=vegan (accessed on 10 August 2020).
- Rolong, N.A.; Arroyo-Cabrales, J.; Allen, B.L.; Johnson, E.; Flores-Díaz, A. Sediments Properties from San Josecito Cave, Nuevo León, México. In Proceedings of the Transactions of the 15th World Congress of Soil Science, Commission V: Symposia, Acapulco, Mexico, 10–16 July 1994; Etchevers, B., Ed.; Sociedad Mexicana de la Ciencia del Suelo, International Society of Soil Science: Acapulco, México, 1994; Volume 6a, pp. 129–141. [Google Scholar]
- Stock, C. The succession of mammalian forms within the period in which human remains are known to occur in America. Am. Nat. 1936, 70, 324–331. [Google Scholar] [CrossRef]
- Simpson, G.G. Chester Stock 1892–1950. Natl. Acad. Biogr. Mem. 1952, 27, 334–362. [Google Scholar]
- Arroyo-Cabrales, J.; Johnson, E.; Hass, H.; de los Ríos-Paredes, M.; Ralph, R.W.; Hartwell, W.T. First Radiocarbon Dates for San Josecito Cave, Nuevo León, México. Quat. Res. 1995, 43, 255–258. [Google Scholar] [CrossRef]
- Bronk-Ramsey, C. Radiocarbon calibration and analysis of stratigraphy: The OxCal program. Radiocarbon 1995, 37, 425–430. [Google Scholar] [CrossRef] [Green Version]
- Bronk-Ramsey, C. Bayesian analysis of radiocarbon dates. Radiocarbon 2009, 51, 337–360. [Google Scholar] [CrossRef] [Green Version]
- Bronk-Ramsey, C.; Dee, M.; Lee, S.; Nakagawa, T.; Staff, R. Developments in the calibration and modelling of radiocarbon dates. Radiocarbon 2010, 52, 953–961. [Google Scholar] [CrossRef] [Green Version]
- Bronk-Ramsey, C. Methods for Summarizing Radiocarbon Datasets. Radiocarbon 2017, 59, 1809–1833. [Google Scholar] [CrossRef] [Green Version]
- Reimer, P.; Austin, W.; Bard, E.; Bayliss, A.; Blackwell, P.; Bronk-Ramsey, C.; Butzin, M.; Cheng, H.; Edwards, R.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Jakway, G.E. Pleistocene Lagomorpha and Rodentia from the San Josecito Cave, Nuevo Leon, Mexico. Trans. Kans. Acad. Sci. 1958, 61, 313–327. [Google Scholar] [CrossRef]
- Mammal Diversity Database. Version 1.6. 2021. Available online: http://0-doi-org.brum.beds.ac.uk/10.5281/zenodo.4139818 (accessed on 12 May 2021). [CrossRef]
- Arroyo-Cabrales, J.; Johnson, E. Mammalian additions to the faunal assemblages from San Josecito Cave, Nuevo León, México. In Avances en el Estudio de los Mamíferos de México II; Lorenzo, C., Espinoza, E., Ortega, J., Eds.; Asociación Mexicana de Mastozoología Asociación Civi: San Cristóbal de las Casas, CS, Mexico, 2008; pp. 65–87. [Google Scholar]
- Esteva, M.; Arroyo-Cabrales, J.; Flores Martínez, A.; Johnson, E.; Polaco, O.J. Fossil Shrews from San Josecito Cave, Nuevo Leon, Mexico. In Advances in the Biology of Shrews II; Merritt, J.F., Churchfield, S., Hutterer, R., Sheftel, B.L., Eds.; Special Publication of the International Society of Shrew Biologists: New York, NY, USA, 2005; Volume 1, pp. 31–43. [Google Scholar]
- Woodman, N. American Recent Eulipotyphla: Nesophontids, Solenodons, Moles, and Shrews in the New World. Smithson. Contr. Zool. 2018, 650, 1–107. [Google Scholar] [CrossRef]
- Greenbaum, I.F.; Honeycutt, R.L.; Chirhart, S.E. Taxonomy and Phylogenetics of the Peromyscus maniculatus Species Group. Spec. Publ. Mus. Tex. Tech. Univ. 2019, 71, 559–575. [Google Scholar]
- de Abreu, E.F., Jr.; Pavan, S.E.; Tsuchiya, M.T.N.; Wilson, D.E.; Percequillo, A.R.; Maldonado, J.E. Museomics of Tree Squirrels: A Dense Taxon Sampling of Mitogenomes Reveals Hidden Diversity, Phenotypic Convergence, and the Need of a Taxonomic Overhaul. BMC Evol. Biol. 2020, 20, 77. [Google Scholar] [CrossRef]
- Hall, E.R. The Mammals of North America; John Wiley & Sons: New York, NY, USA, 1981; p. 1300. [Google Scholar]
- Polaco, J.O.; Butrón, M.L. Mamíferos pleistocénicos de la cueva La Presita, San Luis Potosí. México. In Homenaje al Profesor Ticul Álvarez; Colección Científica; Arroyo-Cabrales, J., Polaco, J.O., Eds.; Instituto Nacional de Antropología e Historia: Mexico City, Mexico, 1997; pp. 279–296. [Google Scholar]
- Chávez-Tovar, J.C. Peromyscus levipes Merriam, 1898. In Los Mamíferos Silvestres de México; Ceballos, G., Oliva, G., Eds.; Comisíon Nacional para el Conocimiento y Uso de la Biodiversidad, Fondo de Cultura Económica: México City, Mexico, 2005; pp. 745–746. [Google Scholar]
- Álvarez-Castañeda, S.T.; González-Ruiz, N. Peromyscus levipes (Rodentia: Cricetidae). Mamm. Species 2009, 824, 1–6. [Google Scholar] [CrossRef]
- Best, T.L. Sciurus alleni. Mamm. Species 1995, 501, 1–4. [Google Scholar]
- Howard, H. Quaternary avian remains from Dark Canyon Cave, New Mexico. Condor 1971, 73, 237–240. [Google Scholar] [CrossRef]
- Olson, S.L. The Pleistocene rails of North America. Condor 1974, 76, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Olson, S.L. An evaluation of the fossil curlew Palnumenius victima L. Miller (Aves: Scolopacidae). In Contributions in Quaternary Vertebrate Paleontology: A Volume in Memorial to John E. Guilday; Genoways, H.H., Dawson, M.R., Eds.; Special Publication; Carnegie Museum of Natural History: Pittsburgh, PA, USA, 1984; pp. 53–55. [Google Scholar]
- Rea, A.M. The Indeterminate Parrot of Nuevo León. In The Era of Allan R. Phillips: A Festschrift; Dickerman, R.W., Ed.; Horizon Communications: Albuquerque, NM, USA, 1991; pp. 167–176. [Google Scholar]
- Steadman, D.W.; Arroyo-Cabrales, J.; Johnson, E.; Guzmán, A.F. New information on the Late Pleistocene birds from San Josecito Cave, Nuevo León, México. Condor 1994, 96, 577–589. [Google Scholar] [CrossRef]
- American Ornithologists’ Union (AOU). Check-List of North American Birds, 6th ed.; Allen Press: Lawrence, KS, USA, 1983; p. 877. [Google Scholar]
- Phillips, A.R. The Known Birds of North and Middle America. Part I. Hirundinidae to Mimidae, Certhiidae; UNKNO: Denver, CO, USA, 1986; p. 320. [Google Scholar]
- Moore, R.T. Records of two North American corvids in Mexico. Condor 1951, 53, 101. [Google Scholar]
- Selander, R.K. Additional record of the Pinon Jay in Mexico. Condor 1956, 58, 239. [Google Scholar]
- Hayman, P.; Marchant, J.; Prater, T. Shorebirds. An Identification Guide to the Wader of the World; Croom Helm: London, UK, 1986; p. 412. [Google Scholar]
- Johnsgard, P.A. The Plovers, Sandpipers and Snipes of the World; University of Nebraska Press: Lincoln, NE, USA, 1981; p. 519. [Google Scholar]
- Van Perlo, B. Birds of Mexico and Central America; Princeton University Press: Princeton, NJ, USA, 2006; p. 336. [Google Scholar]
- Brattstrom, B.H. Pleistocene lizards from San Josecito Cavern, Mexico, with description of new species. Copeia 1955, 1955, 133–134. [Google Scholar] [CrossRef]
- Mead, J.I.; Arroyo-Cabrales, J.; Johnson, E. Pleistocene lizards (Reptilia: Squamata) from San Josecito cave, Nuevo Leon, Mexico. Copeia 1999, 1999, 163–173. [Google Scholar] [CrossRef]
- Smith, H.M.; Burg, T.M.; Chiszar, D. Evolutionary speciation in the alligator lizard of the genus Barisia. Bull. Md. Herpetol. Soc. 2002, 38, 23–26. [Google Scholar]
- Cruz-Sáenz, D.; Muñoz-Nolasco, F.J.; Mata-Silva, V.; Johnson, J.D.; García-Padilla, E.; Wilson, L.D. The Herpetofauna of Jalisco, Mexico: Composition, Distribution, and Conservation Status. Mesoam. Herpetol. 2017, 4, 23–118. [Google Scholar]
- Bryson, R.W., Jr.; Riddle, B.R. Tracing the Origins of Widespread Highland Species: A Case of Neogene Diversification Across the Mexican Sierras in an Endemic Lizard. Biol. J. Linn. Soc. 2012, 105, 382–394. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Silva, J.A. Reconstrucción paleoambiental del Pleistoceno final usando modelos de nicho ecológico inferidos con vertebrados: El caso de la Cueva de San Josecito, Nuevo León. Master’s Thesis, Instituto de Biología, Universidad Nacional Autónoma de México, México City, Mexico, 2012; p. 132. [Google Scholar]
- Conant, R. Collins, J.T. A Field Guide to Reptiles and Amphibians of Eastern/Central North America, 3rd ed.; Houghton Mifflin: Boston, MA, USA; New York, NY, USA, 1991; 450p. [Google Scholar]
- Zaldivar-Riveron, A.; Nieto-Montes de Oca, A.; Laclette, J.P. Phylogeny and evolution of dorsal pattern in the Mexican endemic lizard genus Barisia (Anguidae: Gerrhonotinae). J. Zool. Syst. Evol. Res. 2005, 43, 243–257. [Google Scholar] [CrossRef]
- Sherbrooke, W.C. Introduction to Horned Lizards of North America; California Natural History Guides; University of California Press: Berkeley, CA, USA, 2003; p. 191. [Google Scholar]
- Tovar-Liceaga, R.E.; Montellano-Ballesteros, M. Pleistocene microvertebrates from Santa Cruz Nuevo, Puebla, Mexico. Curr. Res. Pleistocene 2006, 23, 72–74. [Google Scholar]
- Van Devender, T.R.; Rea, A.M.; Smith, M.L. The Sangamon interglacial vertebrate fauna from Rancho La Brisca, Sonora, Mexico. Trans. San Diego Soc. Nat. Hist. 1985, 21, 23–55. [Google Scholar] [CrossRef]
- Cruz, J.A.; Arroyo-Cabrales, J.; Reynoso, V.H. Reconstructing the paleoenvironment of Loltún Cave, Yucatán, México, with Pleistocene amphibians and reptiles and their paleobiogeographic implications. Rev. Mex. Cienc. Geol. 2016, 33, 342–354. [Google Scholar]
- Shaffer, H.B.; McKnight, M.L. The polytypic species revisited: Genetic differentiation and molecular phylogenetics of the tiger salamander Ambystoma tigrinum (Amphibia: Caudata) complex. Evolution 1996, 50, 417–433. [Google Scholar] [CrossRef]
- Bryson, R.W., Jr.; Murphy, R.W.; Graham, M.R.; Lathrop, A.; Lozano, D. Ephemeral Pleistocene woodlands connect the dots for highland rattlesnakes of the Crotalus intermedius group. J. Biogeogr. 2011, 38, 2299–2310. [Google Scholar] [CrossRef]
- Bryson, R.W., Jr.; García-Vasquez, U.O.; Riddle, B.R. Diversification in the mexican horned lizard Phrynosoma orbiculare across a dynamic landscape. Mol. Phylogenet. Evol. 2012, 62, 87–96. [Google Scholar] [CrossRef]
- Guillette, L.J., Jr.; Smith, H.M. A review of the Mexican lizard Barisia imbricata, and the description of a new subspecies. Trans. Kans. Acad. Sci. 1982, 85, 13–33. [Google Scholar] [CrossRef]
- Canseco-Márquez, L.; Mendoza-Quijano, F.; Gutiérrez-Mayen, M.G. Análisis de la distribución de la herpetofauna. In Biodiversidad de la Sierra Madre Oriental; Luna, I., Morrone, J.J., Espinosa, D., Eds.; Las Prensas de Ciencias: México City, Mexico, 2004; pp. 417–437. [Google Scholar]
- Lazcano, D.; Sánchez-Almazán, A.; García-de la Peña, C.; Castañeda, G.; Contreras-Balderas, A.J. Notes on Mexican herpetofauna 9: Herpetofauna of a fragmented Juniperus forest in the state Natural Protected Area of San Juan y Puentes, Arramberri, Nuevo León, México. Bull. Chic. Herpetol. Soc. 2007, 42, 21–26. [Google Scholar]
- Martin, P.S. A biogeography of reptiles and amphibians in the Gomez Farias region, Tamaulipas, Mexico. Misc. Publ. Mus. Zool. Univ. Mich. 1958, 101, 1–102. [Google Scholar]
- Gadsden, H.; Rodríguez-Romero, F.J.; Méndez-de la Cruz, F.R.; Gil-Martínez, R. Ciclo reproductor de Sceloporus poinsettii Baird y Girard 1852 (Squamata: Phrynosomatidae) en el centro del desierto chihuahuense, México. Acta Zool. Mex. 2005, 21, 93–107. [Google Scholar]
- Méndez-de la Cruz, F.R.; Villagrán-Santa Cruz, M.; Cuellar, O. Geographic variation of spermatogenesis in the Mexican viviparous lizard Sceloporus mucronatus. Biogeographica 1994, 70, 59–67. [Google Scholar]
- Douglas, M.E.; Douglas, M.R.; Schuett, G.W.; Porras, L.W. Evolution of rattlesnakes (Viperidae; Crotalus) in the warm deserts of western North America shaped by Neogene vicariance and Quaternary climate change. Mol. Ecol. 2006, 15, 3353–3374. [Google Scholar] [CrossRef]
- Hutchison, D.W.; Malcomber, S.T.; Pletscher, L.S. A multidisciplinary investigation of the applicability of the Pleistocene herpetofaunal stability model to collared lizards (Crotaphytus collaris). Herpetol. Monogr. 1999, 13, 81–141. [Google Scholar] [CrossRef]
- McGuire, J.A.; Linkem, C.W.; Koo, M.S.; Hutchison, D.W.; Lappin, A.K.; Orange, D.I.; Lemos-Espinal, J.; Riddle, B.R.; Jaeger, J.R. Mitochondrial introgression and incomplete lineage sorting through space and time: Phylogenetics of crotaphytid lizards. Evolution 2007, 61, 2879–2897. [Google Scholar] [CrossRef] [PubMed]
- Waltari, E.; Hijmans, R.J.; Townsend-Peterson, A.; Nyári, A.S.; Perkins, S.L.; Guralnick, R.P. Locating Pleistocene refugia: Comparing phylogeography and ecological niche model predictions. PLoS ONE 2007, 2, e563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holman, J.A. Pleistocene Amphibians and Reptiles in North America; Oxford University Press: New York, NY, USA, 1995; p. 256. [Google Scholar]
- Arroyo-Cabrales, J.; Johnson, E. Preliminary analysis of bone from San Josecito Cave, Nuevo Leon, Mexico. In Proceedings of the 1993 Bone Modification Conference; Occasional Papers; Hannus, L.A., Rossum, L., Winham, R.P., Eds.; Archeology Lab Augustana College: Hot Springs, SD, USA, 1997; pp. 50–64. [Google Scholar]
- Caballero, M.; Lozano-García, S.; Vázquez-Selem, L.; Ortega, B. Evidencias de cambio climático y ambiental en registros glaciales y en cuencas lacustres del centro de México durante el último máximo glacial. Boletín Soc. Geol. Mex. 2010, 62, 359–377. [Google Scholar] [CrossRef]
- Metcalfe, S.E.; O’Hara, S.L.; Caballero, M.; Davies, S.J. Records of Late Pleistocene-Holocene climatic change in Mexico: A review. Quat. Sci. Rev. 2000, 19, 699–721. [Google Scholar] [CrossRef]
- Metcalfe, S.E. Late Quaternary environments of the northern deserts and Central Transvolcanic Belt of Mexico. Ann. Mol. Bot. Gard. 2006, 93, 258–273. [Google Scholar] [CrossRef]
- Van Devender, T.R.; Mead, J.I. Early Holocene and Late Pleistocene Amphibians and Reptiles in Sonoran Desert Packrat Middens. Copeia 1978, 1978, 464–475. [Google Scholar] [CrossRef]
- Badgley, C. Counting individuals in mammalian fossil assemblages from fluvial environments. Palaios 1986, 1, 328–338. [Google Scholar] [CrossRef]
- Fernández-Jalvo, Y.; Andrews, P. Small mammal taphonomy of Gran Dolina, Atapuerca (Burgos), Spain. J. Archaeol. Sci. 1992, 19, 407–428. [Google Scholar] [CrossRef]
- Andrews, P. Owls, Caves, and Fossils, 1st ed.; The University of Chicago Press and British Museum (Natural History): Chicago, IL, USA, 1990; p. 231. [Google Scholar]
- Robles, J.; Arroyo-Cabrales, J.; Johnson, E.; Allen, B.L.; Izquierdo, G. Blue Bone Analyses as a Contribution to the Study of Bone Taphonomy in San Josecito Cave, Nuevo León, México. J. Cave Karst Stud. 2002, 64, 147–152. [Google Scholar]
- Chadefaux, C.; Vignaud, C.; Chalmin, E.; Robles-Camacho, J.; Arroyo-Cabrales, J.; Johnson, E.; Reiche, I. Colour Origin and Heat Evidence of Paleontological Bones: Case Study of Blue and Gray Bones from San Josecito Cave, México. Am. Mineral. 2009, 94, 27–33. [Google Scholar] [CrossRef]
- Balvanera, P.; Kremen, C.; Martínez-Ramos, M. Applying community structure analysis to ecosystem function: Examples from pollination and carbon storage. Ecol. Appl. 2005, 15, 360–375. [Google Scholar] [CrossRef]
- Wing, S.L.; Sues, H.-D.; Potts, R.; DiMichele, W.A.; Behrensmeyer, A.K. Evolutionary Paleoecology. In Terrestrial Ecosystems through Time. Evolutionary Paleoecology of Terrestrial Plants and Animals; Behrensmeyer, A.K., Damuth, J.D., DiMichele, W.A., Potts, R., Sues, H.-D., Wing, S.L., Eds.; The University of Chicago Press: Chicago, IL, USA, 1992; pp. 1–13. [Google Scholar]
- Harris, A.H. Reconstruction of mid-Wisconsin environments in southern New Mexico. Natl. Geogr. Res. 1987, 3, 142–151. [Google Scholar]
- Meltzer, D.J.; Mead, J.I. Dating Late Pleistocene extinctions: Theoretical issues, analytical bias, and substantive results. In Environments and Extinctions: Man in Late Glacial North America; Peopling of the Americas Series; Mead, J.I., Meltzer, D.J., Eds.; Center for the Study of Early Man, University of Maine: Orono, MI, USA, 1985; pp. 145–173. [Google Scholar]
- Wilson, R.W. Preliminary study of the fauna of Rampart Cave, Arizona. Publ. Carnegie Inst. Wash. 1942, 530, 169–185. [Google Scholar]
- Miller, S.J. The archaeological fauna of four sites in Smith Creek Canyon. In The Archaeology of Smith Creek Canyon, Eastern Nevada; Anthropological Papers; Tuohy, D.R., Randall, D.L., Eds.; Nevada State Museum: Carson City, Nevada, 1979; Volume 17, pp. 271–329. [Google Scholar]
- Jass, C. Caves, Arvicoline Rodents, and Chronologic Resolution. Palaeontol. Electron. 2011, 14, 1–21. [Google Scholar]
- White, J.A.; McDonald, H.G.; Anderson, E.; Soiset, J.M. Lava blisters as carnivore 25s. In Contributions in Quaternary Vertebrate Paleontology: A Volume in Memorial of John E. Guilday; Genoways, H.H., Dawson, M.R., Eds.; Carnegie Museum of Natural History: Carnegie, PA, USA, 1984; Volume 8, pp. 241–256. [Google Scholar]
- Mares, M.A.; Willig, M.R. Inferring biome associations of recent mammals from samples of temperate and tropical faunas: Paleoecological considerations. Hist. Biol. 1994, 8, 31–48. [Google Scholar] [CrossRef]
- Barbour, M.G.; Billings, W.D. North American Terrestrial Vegetation; Cambridge University Press: New York, NY, USA, 1988; p. 434. [Google Scholar]
- Sutcliffe, A.J. On the Track of the Ice Age Mammals; British Museum (Natural History): London, UK, 1985; p. 224. [Google Scholar]
- Carbone, C.; Maddox, T.; Funston, P.J.; Mills, M.G.L.; Grether, G.F.; Van Valkenburgh, B. Parallels between Playbacks and Pleistocene Tar Seeps Suggest Sociality in an Extinct Sabretooth cat, Smilodon. Biol. Lett. 2009, 5, 81–85. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, L.R.G.; Crites, J.M.; Feranec, R.S.; Fox-Dobbs, K.; Farrell, A.B.; Harris, J.M.; Takeuchi, G.T.; Cerling, T.E. Causes and Consequences of Pleistocene Megafaunal Extinctions as Revealed from Rancho La Brea Mammals. Curr. Biol. 2019, 29, 2488–2495. [Google Scholar] [CrossRef] [PubMed]
- Zepeda, G.J. Los Mamíferos Recientes de la Región de la Cueva de San Josecito, Nuevo León, México. Unpublished Bachelor’s Thesis, Escuela Nacional de Ciencias Biológicas, IPN, México City, Mexico, 2003. [Google Scholar]
- Baker, R.H. Mammals of Coahuila, Mexico. Univ. Kans. Publ. Kans. Mus. Nat. Hist. 1956, 9, 125–335. [Google Scholar]
- Schmidly, D.J. The Mammals of Trans-Pecos Texas; Texas A&M University: College Station, TX, USA, 1977; p. 225. [Google Scholar]
- Ray, C.E.; Linares, O.J.; Morgan, G.S. Paleontology. In Natural History of Vampire Bats; Greenhall, A.M., Schmidt, U., Eds.; CRC Press, Inc.: Boca Raton, FL, USA, 1988; pp. 19–29. [Google Scholar]
- Koestner, E.J. An annotated list of mammals collected in Nuevo Leon, Mexico, in 1938. Great Basin Nat. 1941, 2, 9–15. [Google Scholar]
- Morgan, G.S. Neotropical Chiroptera from the Pliocene and Pleistocene of Florida. Bull. Am. Mus. Nat. Hist. 1991, 206, 176–213. [Google Scholar]
- McNab, B.K. Energetics and the distribution of vampires. J. Mammal. 1973, 54, 131–144. [Google Scholar] [CrossRef]
- Grayson, D.K. Minimum Numbers and Sample Size in Vertebrate Faunal Analysis. Am. Antiq. 1978, 43, 53–65. [Google Scholar] [CrossRef]
- Sibley, D.A. The Sibley Guide to Birds; Alfred A. Knopf: New York, NY, USA, 2000; p. 624. [Google Scholar]
- Rzedowski, J. Vegetación de México; CONABIO: México City, Mexico, 2006; p. 505. [Google Scholar]
- Craig, E.H.; Craig, T.H.; Powers, L.R. Patterns and Home-Range Use of Nesting Long-Eared Owls. Wilson J. Ornithol. 1988, 100, 204–213. [Google Scholar]
- Henrioux, F. Home Range and Habitat Use by the Long-Eared Owl in Northwestern Switzerland. J. Raptor Res. 2000, 34, 93–101. [Google Scholar]
- Lövy, M.; Riegert, J. Home Range and Land Use of Urban Long-Eared Owls. Condor 2013, 115, 551–557. [Google Scholar] [CrossRef]
- Lyman, R.L. Quantitative Units and Terminology in Zooarchaeology. Am. Antiq. 1994, 59, 36–71. [Google Scholar] [CrossRef]
- McDonald, H.G.; Jefferson, G.T. Distribution and habitat of Nothrotheriops (Xenarthra, Nothrotheridae) in the Pleistocene of North America. In Geology and Vertebrate Paleontology of Western and Southern North America, Contributions in Honor of David P. Whistler; Wang, X., Barnes, L.G., Eds.; Natural History Museum of Los Angeles County Science Series: Los Angeles, CA, USA, 2008; No. 41; pp. 313–331. [Google Scholar]
- Tedford, R.H. Principles and Practices of Mammalian Geochronology in North America. In Proceedings of the North American Paleontological Convention: Field Museum of Natural History, Chicago, IL, USA, 5–7 September 1969; Yochelson, E.L., Ed.; pp. 666–703. [Google Scholar]
Figure 1.
Location of San Josecito Cave (black point) in the Sierra Madre Oriental (SMO), Mexico, with plan and profile (dashed lines) facing west of San Josecito Cave. Modified from [28] and adapted with permission from George Veni, National Speleological Society.
Figure 1.
Location of San Josecito Cave (black point) in the Sierra Madre Oriental (SMO), Mexico, with plan and profile (dashed lines) facing west of San Josecito Cave. Modified from [28] and adapted with permission from George Veni, National Speleological Society.

Figure 2.
(A) Original cave profile indicating the spatial position of bones considered noteworthy (letter of Bode to Stock-Stock, personal correspondence, 19 January 1936). (B) Stock´s cave plan and profile showing excavations blocks.
Figure 2.
(A) Original cave profile indicating the spatial position of bones considered noteworthy (letter of Bode to Stock-Stock, personal correspondence, 19 January 1936). (B) Stock´s cave plan and profile showing excavations blocks.

Figure 3.
Stock’s block 2 and excavated levels by foot increments superimposed over the 1990 stratigraphic profile of the north wall of Stock’s pit [28].
Figure 3.
Stock’s block 2 and excavated levels by foot increments superimposed over the 1990 stratigraphic profile of the north wall of Stock’s pit [28].

Figure 4.
The plotting of the absolute values obtained for the different quantification methods used in the taphonomic study. Plots represent the utilized method, as follow: 1—NISP; 2—MNE; 3—MNI; 4—BOX. Species are represented by the following abbreviations: (a) Sorex altoensis; (b) Chiroptera; (c) Leptonycteris nivalis; (d) Desmodus stocki; (e) Myotis californicus; (f) Myotis thysanodes; (g) Myotis sp.; (h) Sylvilagus floridanus; (i) Sylvilagus sp.; (j) Parasciurus alleni; (k) Neotoma leucodon; (l) Neotoma mexicana; (m) Neotoma sp.; (n) Peromyscus difficilis; (o) Peromyscus levipes; (p) Peromyscus labecula; (q) Peromyscus sp.; (r) Reithrodontomys megalotis; (s) Microtus mexicanus.
Figure 4.
The plotting of the absolute values obtained for the different quantification methods used in the taphonomic study. Plots represent the utilized method, as follow: 1—NISP; 2—MNE; 3—MNI; 4—BOX. Species are represented by the following abbreviations: (a) Sorex altoensis; (b) Chiroptera; (c) Leptonycteris nivalis; (d) Desmodus stocki; (e) Myotis californicus; (f) Myotis thysanodes; (g) Myotis sp.; (h) Sylvilagus floridanus; (i) Sylvilagus sp.; (j) Parasciurus alleni; (k) Neotoma leucodon; (l) Neotoma mexicana; (m) Neotoma sp.; (n) Peromyscus difficilis; (o) Peromyscus levipes; (p) Peromyscus labecula; (q) Peromyscus sp.; (r) Reithrodontomys megalotis; (s) Microtus mexicanus.

Figure 5.
The distribution of the overall number of individuals (based on MNI) by taphonomic box for stratum 720 in excavation unit 534N197E, San Josecito Cave, Nuevo León, Mexico.
Figure 5.
The distribution of the overall number of individuals (based on MNI) by taphonomic box for stratum 720 in excavation unit 534N197E, San Josecito Cave, Nuevo León, Mexico.

Figure 6.
Dendrogram obtained from the UPGMA analysis of Pre-LGM faunas from different localities. Site number is located before the level value; those numbers correspond to the ones utilized Arroyo-Cabrales ([28], Table 28). (1) Dry Cave, New Mexico (25,610–33,590 14C years BP), (2) U-Bar Cave, New Mexico (26,150–35,890 14C years BP), (3) San Josecito Cave, Nuevo Leon, Mexico (27,970 14C years BP), (4) Hueco Mountains. New Mexico (>33,000 14C years BP), (5) Vulture Cave, Arizona (29,000–30,000 14C years BP), (6) Ambystoma Faunule, Jones Local Fauna, Meade Co., Kansas (26,700–29,000 14C years BP), (7) Rampart Cave, Arizona (23,540–36,200 14C years BP), (8) Hot Springs, South Dakota (26,075 14C years BP), (9) El Cedral, San Luis Potosi, Mexico (31,800 14C years BP), (10) Lewisville, Denton Co., Texas (>37,000 14C years BP), (11) Channel Islands, Santa Barbara Co., California (29,650 14C years BP), (12) Easley Ranch Local Fauna, Foard Co., Texas (>35,000 14C years BP), (13) Quitaque Local Fauna, Motley Co., Texas (>35,000 14C years BP), (14) Carrol Creek Local Fauna, Donley Co., Texas (32,400 14C years BP), (15) Canadian Local Fauna, Hemphill Co., Texas (34,900 14C years BP), (16) Rainbow Beach Local Fauna, Power Co., Idaho (21,500–31,300 14C years BP), (17) Smith Creek Cave, Nevada (28,650 14C years BP), (18) Fort Qu’appelle, Saskatchewan (>30,000 14C years BP), (19) Medicine Hat Area, Alberta (37,900–38,700 14C years BP), (20) Gold Run Creek, Yukon Territory (22,200–32,350 14C years BP), (21) Selby-Dutton Fauna (29,000 14C years BP), (22) Tesequite Canyon, Cimarron Co., Oklahoma (31,360 14C years BP).
Figure 6.
Dendrogram obtained from the UPGMA analysis of Pre-LGM faunas from different localities. Site number is located before the level value; those numbers correspond to the ones utilized Arroyo-Cabrales ([28], Table 28). (1) Dry Cave, New Mexico (25,610–33,590 14C years BP), (2) U-Bar Cave, New Mexico (26,150–35,890 14C years BP), (3) San Josecito Cave, Nuevo Leon, Mexico (27,970 14C years BP), (4) Hueco Mountains. New Mexico (>33,000 14C years BP), (5) Vulture Cave, Arizona (29,000–30,000 14C years BP), (6) Ambystoma Faunule, Jones Local Fauna, Meade Co., Kansas (26,700–29,000 14C years BP), (7) Rampart Cave, Arizona (23,540–36,200 14C years BP), (8) Hot Springs, South Dakota (26,075 14C years BP), (9) El Cedral, San Luis Potosi, Mexico (31,800 14C years BP), (10) Lewisville, Denton Co., Texas (>37,000 14C years BP), (11) Channel Islands, Santa Barbara Co., California (29,650 14C years BP), (12) Easley Ranch Local Fauna, Foard Co., Texas (>35,000 14C years BP), (13) Quitaque Local Fauna, Motley Co., Texas (>35,000 14C years BP), (14) Carrol Creek Local Fauna, Donley Co., Texas (32,400 14C years BP), (15) Canadian Local Fauna, Hemphill Co., Texas (34,900 14C years BP), (16) Rainbow Beach Local Fauna, Power Co., Idaho (21,500–31,300 14C years BP), (17) Smith Creek Cave, Nevada (28,650 14C years BP), (18) Fort Qu’appelle, Saskatchewan (>30,000 14C years BP), (19) Medicine Hat Area, Alberta (37,900–38,700 14C years BP), (20) Gold Run Creek, Yukon Territory (22,200–32,350 14C years BP), (21) Selby-Dutton Fauna (29,000 14C years BP), (22) Tesequite Canyon, Cimarron Co., Oklahoma (31,360 14C years BP).

Figure 7.
Correspondence canonical analysis (CCA) in different taxonomic groups from composite stratum 720, using the habitat weighting with presence species (HW) and habitat weighting with relative abundance employing number of individual specimens (NISP) and number of individuals (MNI). (A) mammals, (B) reptiles, (C) birds, and (D) all vertebrates.
Figure 7.
Correspondence canonical analysis (CCA) in different taxonomic groups from composite stratum 720, using the habitat weighting with presence species (HW) and habitat weighting with relative abundance employing number of individual specimens (NISP) and number of individuals (MNI). (A) mammals, (B) reptiles, (C) birds, and (D) all vertebrates.



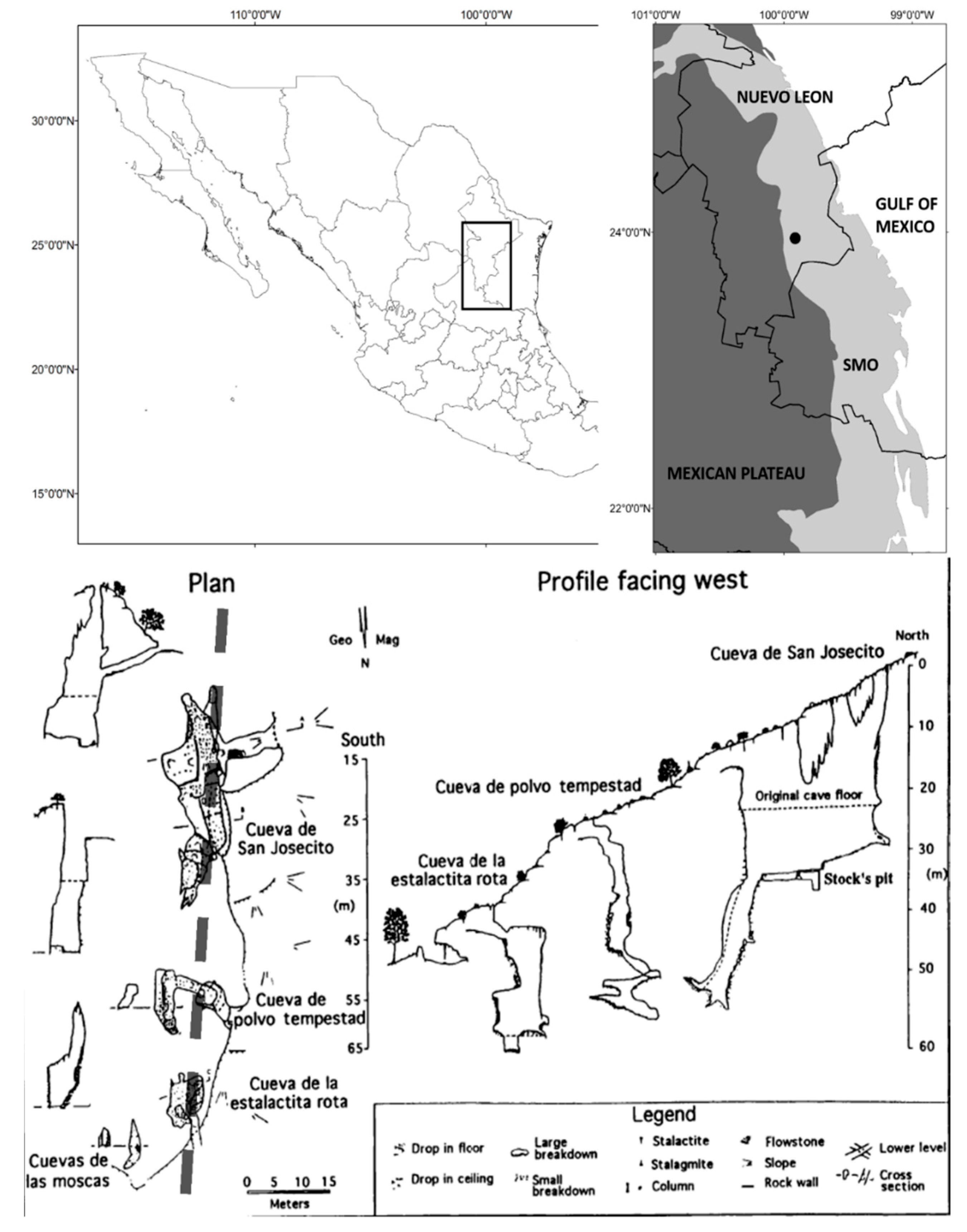{kind=link}
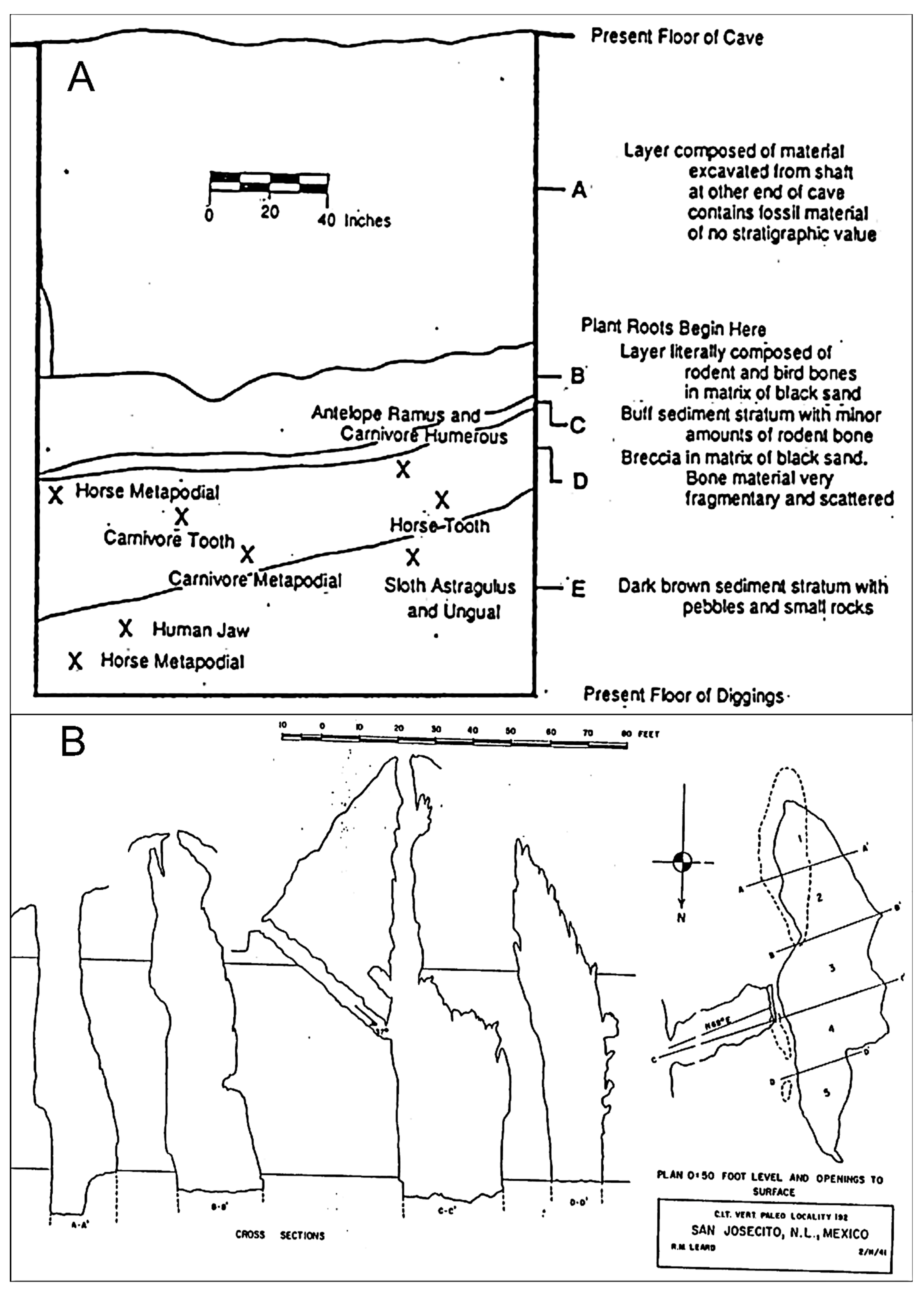{kind=link}
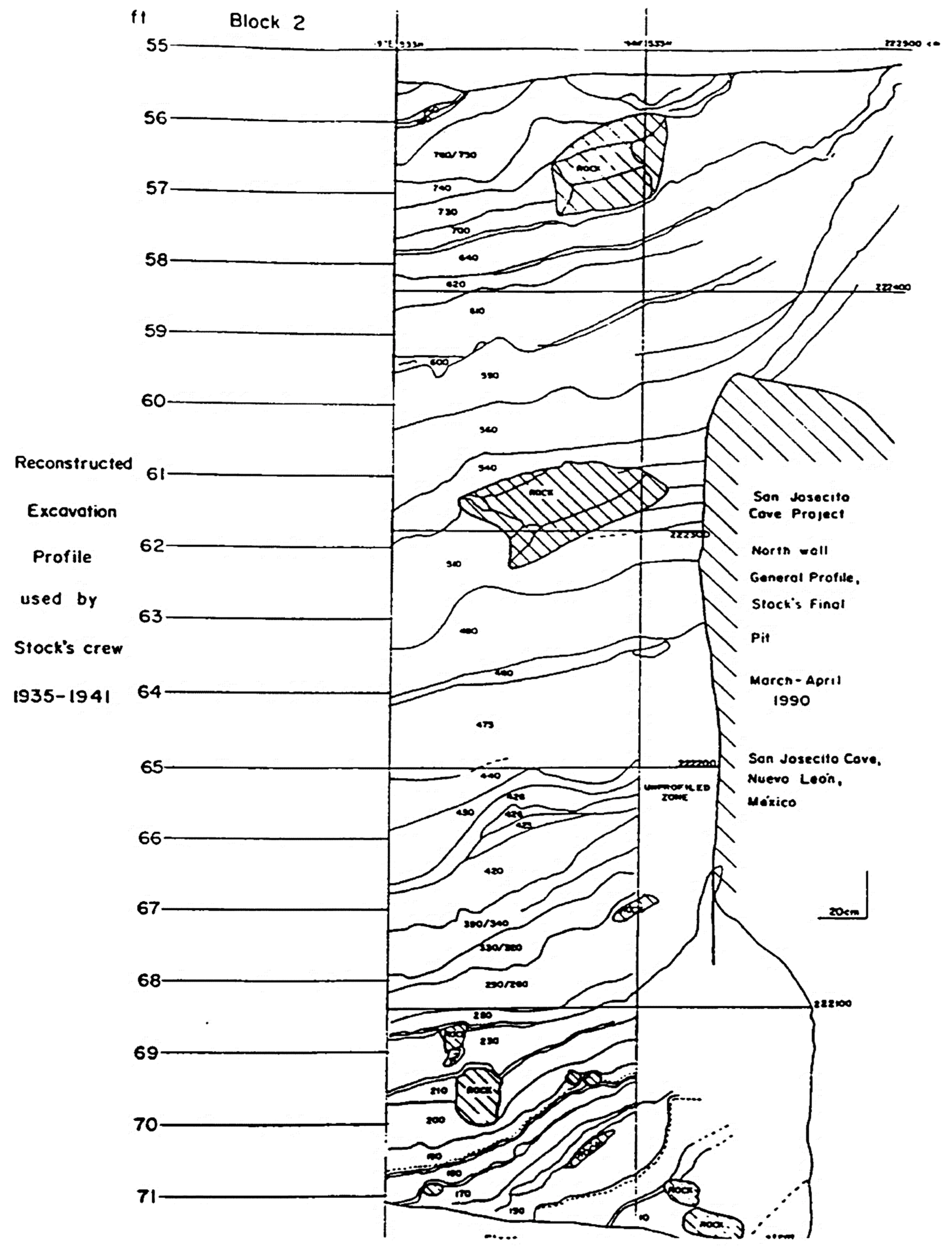{kind=link}
{kind=link}
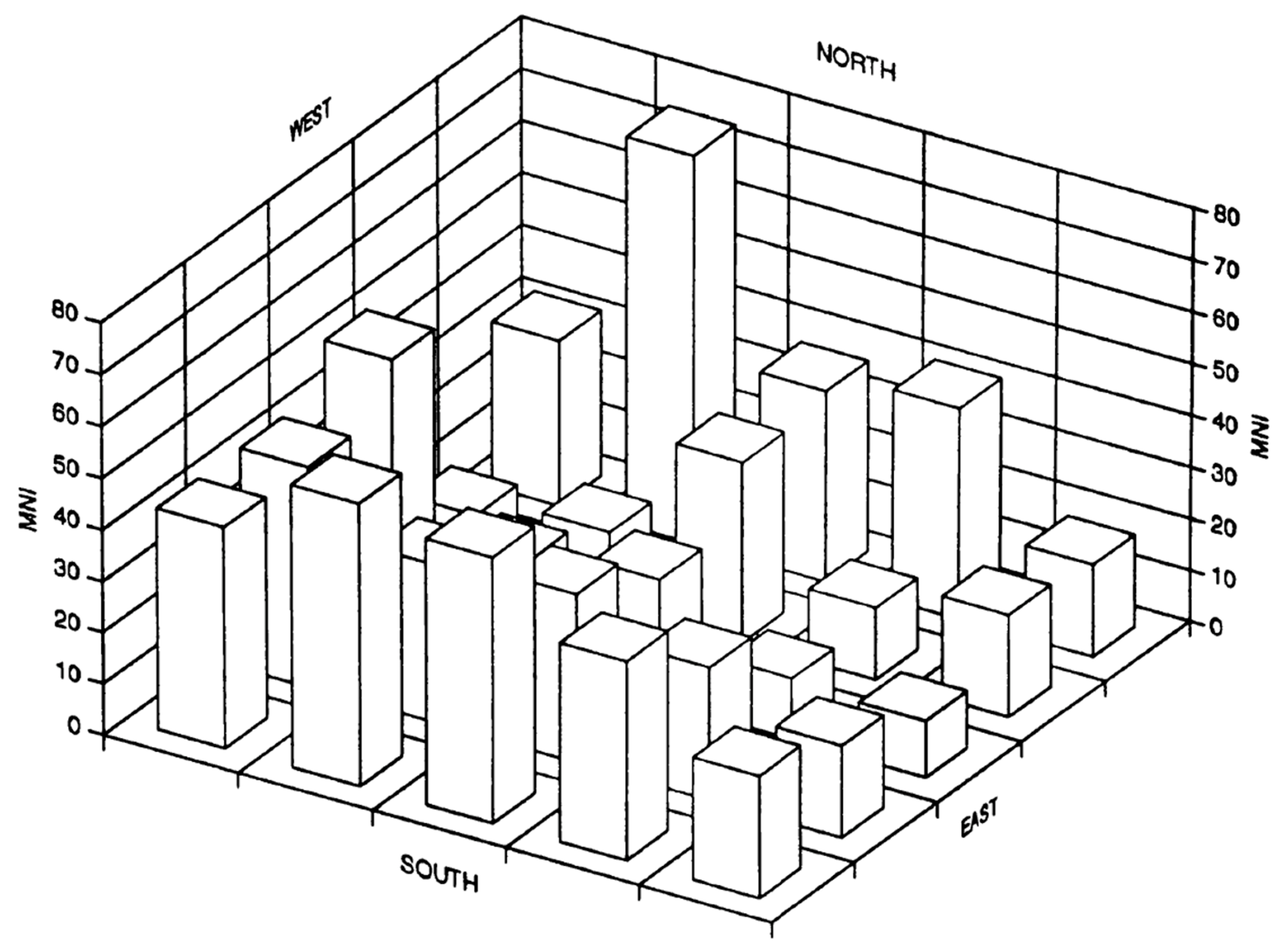{kind=link}
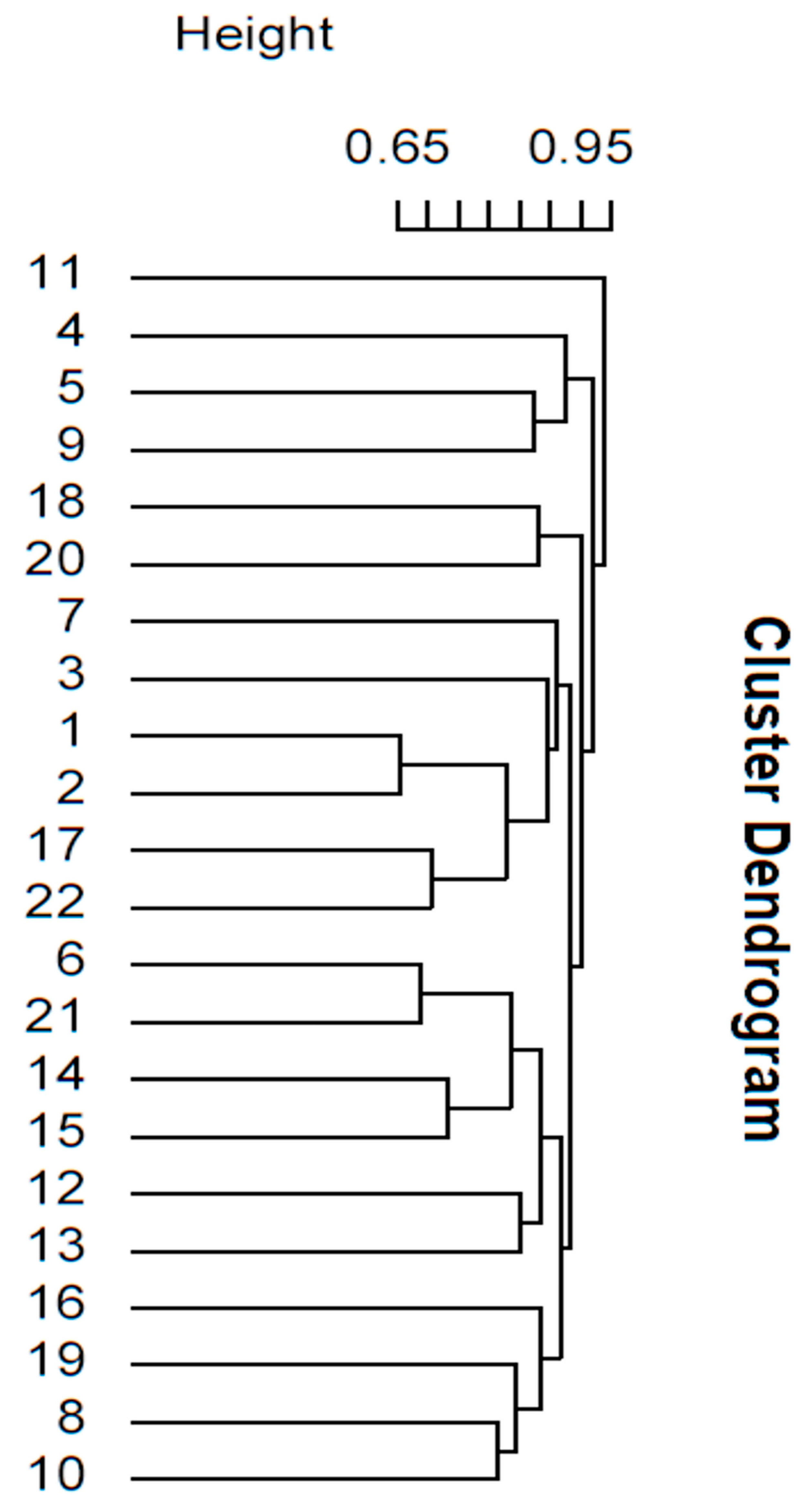{kind=link}
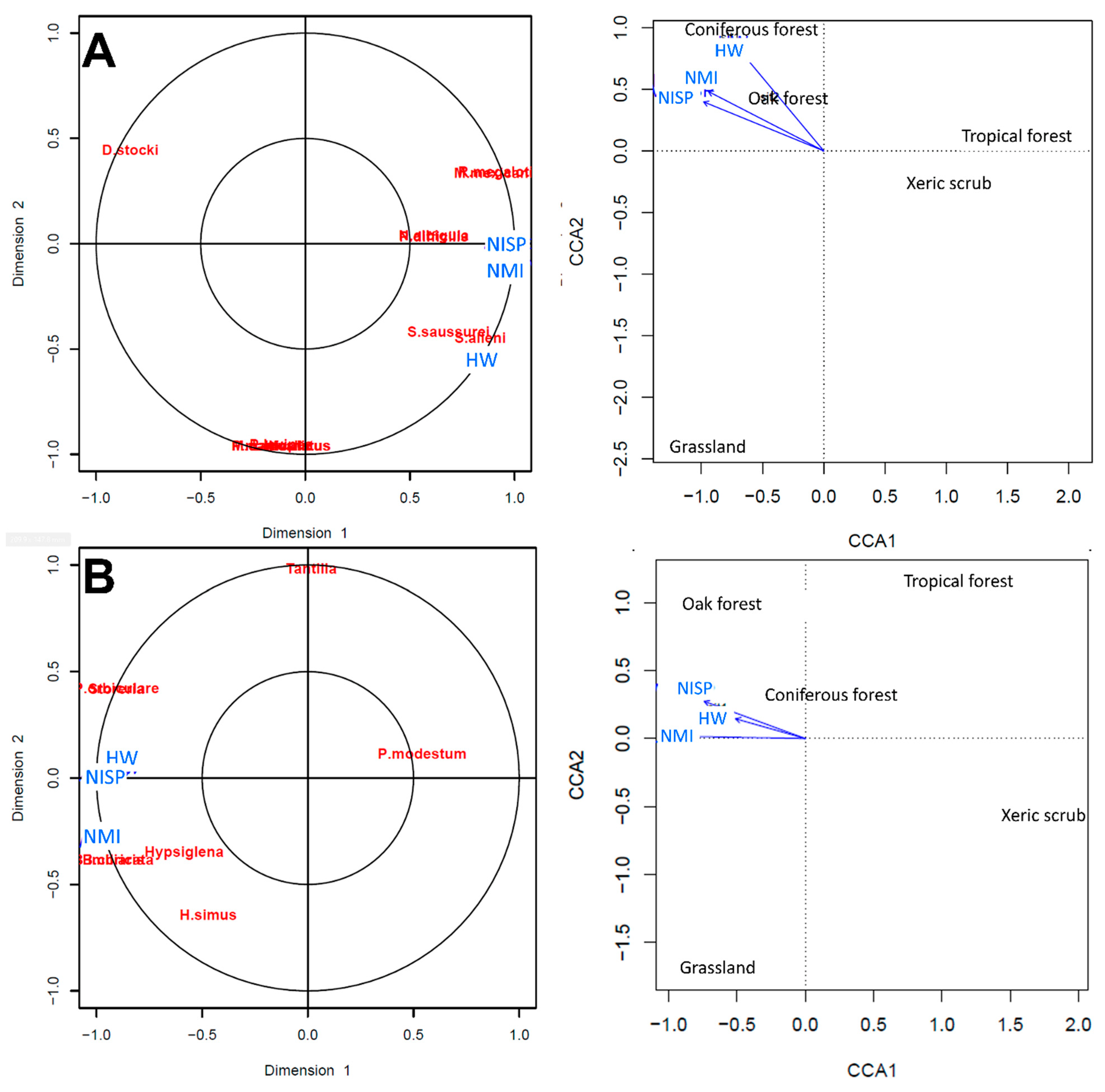{kind=link}
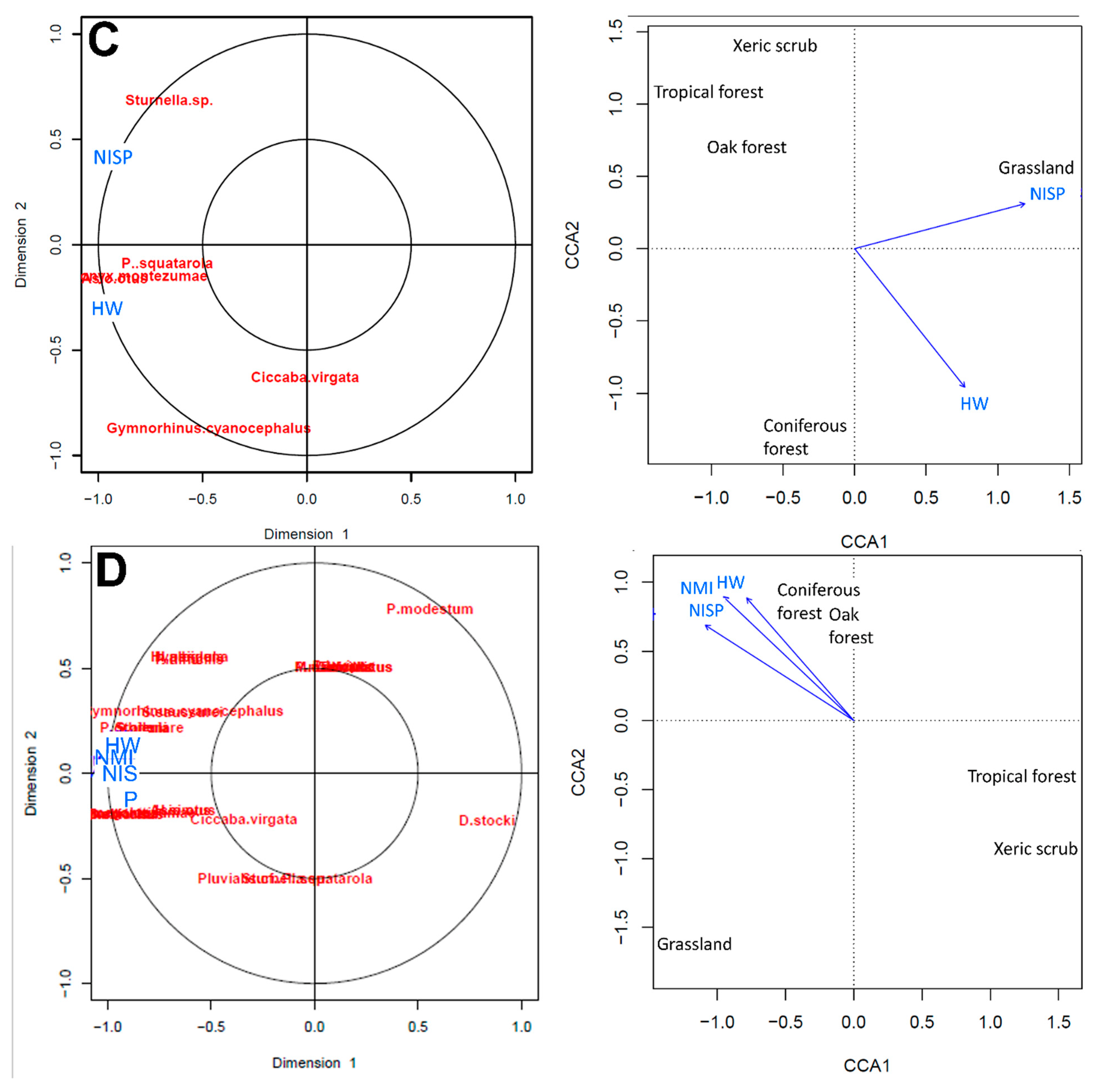{kind=link}
Table 1.
Radiocarbon ages from San Josecito Cave based on organic sediments [45].
Table 1.
Radiocarbon ages from San Josecito Cave based on organic sediments [45].
| Lab Number 1 | Elevation (cm) | Stratum | Uncalibrated Radiocarbon Age | ± | δ13C | Corrected Radiocarbon Age 2 | Calendar Years 3 | ± |
|---|---|---|---|---|---|---|---|---|
| INAH-943A | 222,434–222,425 | 720 | 28,005 | 1035 | 34,892 | 32,942 | 4077 | |
| SMU-2518 | 222,425–222,422 | 710 | 31,290 | 1270 | −16.9 | 39,390 | 37,440 | 4967 |
| SMU-2589 | 222,415–222,413 | 670 | 34,830 | 470 | −15.8 | 39,592 | 37,642 | 1536 |
| SMU-2590 | 222,406–222,402 | 630 | 44,520 | 2590 | −15.5 | 48,507 | 46,557 | 3751 |
| SMU-2532 | 222,371–222,363 | 595 | 37,975 | 700 | −14.4 | 42,684 | 40,734 | 1590 |
| INAH-940 | 222,203–222,162 | 420–340 | 33,700 | 1365 | 43,974 | 42,024 | 6367 |
Table 2.
Mammal species identified from paleontological excavations in San Josecito Cave, Nuevo León, Mexico from the original excavations (1935–1941) and the 1990 excavation (MNI are provided for bones from stratum 720). Nomenclature follows that proposed by [52]. An asterisk denotes extinct taxa (*).
Table 2.
Mammal species identified from paleontological excavations in San Josecito Cave, Nuevo León, Mexico from the original excavations (1935–1941) and the 1990 excavation (MNI are provided for bones from stratum 720). Nomenclature follows that proposed by [52]. An asterisk denotes extinct taxa (*).
| Taxa | Original Excavations | 1990 Excavation |
|---|---|---|
| Didelphidae | ||
| Didelphis virginiana | X | |
| Megatheriidae | ||
| Megalonyx jeffersonii * | X | |
| Nothrotheriops shastensis * | X | X |
| Soricidae | ||
| Cryptotis mexicana | X | X |
| Sorex cinereus or S. milleri | X | X |
| Sorex altoensis | 9 | |
| Mormoopidae | ||
| Mormoops megalophylla | X | |
| Phyllostomidae | ||
| Choeronycteris mexicana | X | |
| Desmodus stocki * | X | X |
| Leptonycteris nivalis | X | 4 |
| Vespertilionidae | ||
| Corynorhinus townsendii | X | |
| Eptesicus fuscus | X | |
| Lasiurus cinereus | X | |
| Myotis californicus | 1 | |
| Myotis thysanodes | 2 | |
| Canidae | ||
| Aenocyon dirus * | X | |
| Canis latrans | X | |
| Canis lupus | X | |
| Cuon alpinus * | X | |
| Urocyon cinereoargenteus | X | |
| Felidae | ||
| Herpailurus yagouaroundi | X | |
| Lynx rufus | X | |
| Panthera atrox * | X | |
| Panthera onca | X | |
| Puma concolor | X | |
| Smilodon fatalis * | X | |
| Mephitidae | ||
| Conepatus mesoleucus | X | |
| Mephitis mephitis | X | |
| Spilogale angustifrons/gracilis | X | |
| Mustelidae | ||
| Mustela frenata | X | |
| Taxidea taxus | X | |
| Procyonidae | ||
| Bassariscus astutus | X | |
| Ursidae | ||
| Tremarctos floridanus * | X | |
| Ursus americanus | X | |
| Equidae | ||
| Equus conversidens * | X | 1 |
| Tapiridae | ||
| Tapirus haysii * | 1 | |
| Tayassuidae | ||
| Platygonus * | X | |
| Camelidae | ||
| Camelops * | X | |
| Cervidae | ||
| Navahoceros fricki * | X | |
| Odocoileus | X | |
| Antilocapridae | ||
| Stockoceros conklingi * | X | |
| Bovidae | ||
| Euceratherium * | X | |
| Oreamnos harringtoni * | X | |
| Sciuridae | ||
| Marmota flaviventris | X | |
| Parasciurus alleni | X | X |
| Xerospermophilus spilosoma | X | |
| Geomyidae | ||
| Cratogeomys castanops | X | |
| Orthogeomys onerous * | X | |
| Thomomys umbrinus | X | X |
| Heteromyidae | ||
| Heteromys irroratus | X | X |
| Cricetidae | ||
| Microtus mexicanus | X | 44 |
| Neotoma albigula | X | 4 |
| Neotoma mexicana | X | 9 |
| Peromyscus difficilis | 8 | |
| Peromyscus labecula | 9 | |
| Peromyscus levipes | X | 16 |
| Reithrodontomy megalotis | X | 16 |
| Sigmodon hispidus | X | X |
| Synaptomys cooperi | X | X |
| Erethizontidae | ||
| Erethizon dorsatum | X | X |
| Leporidae | ||
| Sylvilagus floridanus | X | 2 |
| Sylvilagus leonensis * | X | X |
Table 3.
Mammal species assigned to the composite stratum 720 and Stock’s 57–58 ft level (~28,000 14C years BP). Nomenclature follows that proposed by [52]. An asterisk denotes extinct taxa (*).
Table 3.
Mammal species assigned to the composite stratum 720 and Stock’s 57–58 ft level (~28,000 14C years BP). Nomenclature follows that proposed by [52]. An asterisk denotes extinct taxa (*).
| Taxa | Stock’s 57–58 ft Level | Stratum 720 (MNI) |
|---|---|---|
| Megatheriidae | ||
| Nothrotheriops shastensis * | X | |
| Soricidae | ||
| Sorex altoensis | 9 | |
| Phyllostomidae | ||
| Desmodus stocki * | X | |
| Leptonycteris nivalis | X | 4 |
| Vespertilionidae | ||
| Myotis californicus | 1 | |
| Myotis thysanodes | 2 | |
| Canidae | ||
| Aenocyon dirus * | X | |
| Canis latrans | X | |
| Felidae | ||
| Panthera atrox * | X | |
| Panthera onca | X | |
| Puma concolor | X | |
| Smilodon fatalis * | X | |
| Ursidae | ||
| Tremarctos floridanus * | X | |
| Ursus americanus | X | |
| Equidae | ||
| Equus conversidens * | X | 1 |
| Cervidae | ||
| Navahoceros fricki * | X | |
| Antilocapridae | ||
| Stockoceros conklingi * | X | |
| Bovidae | ||
| Euceratherium * | X | |
| Sciuridae | ||
| Parasciurus alleni | X | |
| Cricetidae | ||
| Microtus mexicanus | X | 44 |
| Neotoma albigula | X | 4 |
| Neotoma mexicana | X | 9 |
| Peromyscus difficilis | 8 | |
| Peromyscus labecula | 9 | |
| Peromyscus levipes | X | 16 |
| Reithrodontomy megalotis | X | 16 |
| Leporidae | ||
| Sylvilagus floridanus | X | 2 |
Table 4.
Bird species identified from paleontological excavations in San Josecito Cave, Nuevo León, Mexico, from the original excavations (1935–1941) and the 1990 excavation. An asterisk denotes extinct taxa (*).
Table 4.
Bird species identified from paleontological excavations in San Josecito Cave, Nuevo León, Mexico, from the original excavations (1935–1941) and the 1990 excavation. An asterisk denotes extinct taxa (*).
| Taxa | Original Excavations | 1990 Excavation (NISP) |
|---|---|---|
| Podicipedidade | ||
| Podilymbus podiceps | X | |
| Ardeidae | ||
| Nycticorax nycticorax | X | |
| Anatidae | ||
| Aix cf. A. sponsa | X | |
| Anas sp. | X | |
| cf. Histrionicus sp. | X | |
| Oxyura cf. O. jamaicensis | X | |
| Ciconiidae | ||
| Ciconia sp. or Mycteria sp. | 1 | |
| Teratornithidae | ||
| Teratornis merriami * | X | |
| Vulturidae | ||
| Gymnogyps californianus | X | |
| Coragyps atratus | X | |
| Accipitridae | ||
| Elanus leucurus | X | |
| Parabuteo unicinctus | X | |
| Buteo cf. B. regalis | X | |
| Buteo jamaicensis | X | 1 |
| Buteo nitidus | X | |
| Aquila chrysaetos | X | |
| Spizaetus grinnelli * | X | |
| Buteogallus daggetti * | X | |
| Neogyps errans * | X | |
| Neophrontops americanus | X | |
| Circus cyaneus | X | |
| Falconidae | ||
| Caracara plancus | X | |
| Falco mexicanus | X | |
| Falco sparverius | X | 20 |
| Falco sp. * | X | |
| Phasianidae | ||
| Cyrtonyx montezumae | X | 85 |
| Dendrortyx sp. | X | |
| Meleagris crassipes * | X | |
| Rallidae | ||
| Rallus elegans/longirostris | X | |
| Rallus limicola | 2 | |
| Fulica americana | X | |
| Charadriidae | ||
| Pluvialis cf. P. squatarola | X | 1 |
| Scolopacidae | ||
| Scolopax minor | 2 | |
| Numenius cf. N. americanus | X | |
| Burhinidae | ||
| Burhinus cf. B. bistriatus | X | |
| Columbidae | ||
| Patagioenas fasciata | X | 10 |
| Zenaida macroura | X | |
| Psittacidae | ||
| Rhynchopsitta terrisi | X | |
| Rhynchopsitta pachyrhyncha | X | |
| Rhynchopsitta phillipsi * | X | |
| Cuculidae | ||
| Geococcyx californianus conklingi * | X | 41 |
| Tytonidae | ||
| Tyto alba | X | |
| Strigidae | ||
| Asio flammeus | X | 1 |
| Asio otus | X | 107 |
| Megascops asio/kennicotti | X | |
| Megascops trichopsis | X | |
| Bubo virginianus | X | |
| Bubo sp. | X | |
| Glaucidium gnoma | X | |
| Micrathene cf. M. whitneyi | X | |
| Strix occidentalis | X | |
| Aegolius acadicus | X | |
| Psiloscops flammeolus | 12 | |
| Ciccaba virgata | X | |
| Caprimulgidae | ||
| Phalaenoptilus nuttallii | X | 2 |
| Picidae | ||
| Colaptes auratus | X | |
| Hirundinidae | ||
| Petrochelidon pyrrhonota | 2 | |
| Corvidae | ||
| Gymnorhinus cyanocephalus | 1 | |
| Aphelocoma coerulescens | 1 | |
| Corvus corax | X | 1051 |
| Muscicapidae | ||
| Turdus migratorius | 2 | |
| Icteridae | ||
| Sturnella sp. | 3 | |
| Small-medium passerines | X | X |
Table 5.
Bird species assigned to the composite stratum 720 and Stock’s 57–58 ft level (~28,000 14C years BP).
Table 5.
Bird species assigned to the composite stratum 720 and Stock’s 57–58 ft level (~28,000 14C years BP).
| Taxa | Original Excavations | Stratum 720 (NISP) |
|---|---|---|
| Accipitridae | ||
| Aquila chrysaetos | X | 75 |
| Phasianidae | ||
| Cyrtonyx montezumae | 85 | |
| Rallidae | ||
| Rallus limicola | 2 | |
| Charadriidae | ||
| Pluvialis cf. P. squatarola | X | 1 |
| Strigidae | ||
| Megascops asio/kennicottii | 7 | |
| Asio otus | X | 107 |
| Ciccaba virgata | X | 20 |
| Corvidae | ||
| Gymnorhinus cyanocephalus | 1 | |
| Corvus corax | X | 1051 |
| Icteridae | ||
| Sturnella sp. | 3 | |
| Small-medium passerines | X | X |
Table 6.
Squamate reptiles identified from paleontological excavations in San Josecito Cave, Nuevo León, Mexico, from the original excavations (1935–1941) and the 1990 excavation (all MNI from stratum 720 [~28,000 14C years BP]). An asterisk denotes extinct taxa (*).
Table 6.
Squamate reptiles identified from paleontological excavations in San Josecito Cave, Nuevo León, Mexico, from the original excavations (1935–1941) and the 1990 excavation (all MNI from stratum 720 [~28,000 14C years BP]). An asterisk denotes extinct taxa (*).
| Taxa | Original Excavations | Stratum 720 (1990 Excavation) |
|---|---|---|
| Anguidae | ||
| Barisia ciliaris | 23 | |
| Barisia imbricata | 8 | |
| Phrynosomatidae | ||
| Phrynosoma modestum | 1 | |
| Phrynosoma josecitensis * | X | |
| Phrynosoma orbiculare | X | 2 |
| Sceloporus spp. | 4 | |
| Colubridae | ||
| Tantilla sp. | 1 | |
| Heterodon cf. H. simus | 1 | |
| Hypsiglena | 1 | |
| Storeria sp. | 1 | |
| Viperidae | ||
| Crotalus spp. | 1 |
Table 7.
Habitat weighting values for composite stratum 720 (~28,000 14C years BP) from San Josecito Cave using small mammals and habitat preferences.
Table 7.
Habitat weighting values for composite stratum 720 (~28,000 14C years BP) from San Josecito Cave using small mammals and habitat preferences.
| Taxa | Coniferous Forest | Oak Forest | Xeric Scrub | Grassland | Tropical Forest | |
| Habitat weighting using presence data | ||||||
| Sorex altoensis | 1 | 0 | 0 | 0 | 0 | |
| Desmondus stocki | 0 | 0 | 0.33 | 0.33 | 0.33 | |
| Leptonycteris nivalis | 0.25 | 0.25 | 0.25 | 0 | 0.25 | |
| Myotis californicus | 0.25 | 0.25 | 0.25 | 0 | 0.25 | |
| Myotis thysanodes | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | |
| Parasciurus alleni | 0.5 | 0.5 | 0 | 0 | 0 | |
| Microtus mexicanus | 0.33 | 0.33 | 0 | 0.33 | 0 | |
| Neotoma albigula | 0.25 | 0.25 | 0.25 | 0.25 | 0 | |
| Neotoma mexicana | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | |
| Peromyscus difficilis | 0.25 | 0.25 | 0.25 | 0.25 | 0 | |
| Peromyscus levipes | 0.25 | 0.25 | 0.25 | 0 | 0.25 | |
| Peromyscus labecula | 0.25 | 0.25 | 0.25 | 0 | 0.25 | |
| Reithrodontomys megalotis | 0.33 | 0.33 | 0 | 0.33 | 0 | |
| Sylvilagus floridanus | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | |
| TOTAL | 4.26 | 3.26 | 2.43 | 2.09 | 1.93 | |
| Habitat weighting using presence and MNI data | ||||||
| Taxa | MNI | Coniferous Forest | Oak Forest | Xeric Scrub | Grassland | Tropical Forest |
| Sorex altoensis | 9 | 9 | 0 | 0 | 0 | 0 |
| Desmondus stocki | 1 | 0 | 0 | 0.33 | 0.33 | 0.33 |
| Leptonycteris nivalis | 4 | 1 | 1 | 1 | 0 | 1 |
| Myotis californicus | 1 | 0.25 | 0.25 | 0.25 | 0 | 0.25 |
| Myotis thysanodes | 2 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 |
| Parasciurus alleni | 2 | 1 | 1 | 0 | 0 | 0 |
| Microtus mexicanus | 44 | 14.52 | 14.52 | 0 | 14.52 | 0 |
| Neotoma albigula | 4 | 1 | 1 | 1 | 1 | 0 |
| Neotoma mexicana | 9 | 1.8 | 1.8 | 1.8 | 1.8 | 1.8 |
| Peromyscus difficilis | 8 | 2 | 2 | 2 | 2 | 0 |
| Peromyscus levipes | 16 | 4 | 4 | 4 | 0 | 4 |
| Peromyscus labecula | 9 | 2.25 | 2.25 | 2.25 | 0 | 2.25 |
| Reithrodontomys megalotis | 16 | 5.28 | 5.28 | 0 | 5.28 | 0 |
| Sylvilagus floridanus | 2 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 |
| TOTAL | 127 | 42.9 | 33.9 | 13.43 | 25.73 | 10.43 |
| Habitat weighting using presence and NISP data | ||||||
| Taxa | NISP | Coniferous Forest | Oak Forest | Xeric Scrub | Grassland | Tropical Forest |
| Sorex altoensis | 60 | 60 | 0 | 0 | 0 | 0 |
| Desmondus stocki | 1 | 0 | 0 | 0.33 | 0.33 | 0.33 |
| Leptonycteris nivalis | 12 | 3 | 3 | 3 | 0 | 3 |
| Myotis californicus | 2 | 0.5 | 0.5 | 0.5 | 0 | 0.5 |
| Myotis thysanodes | 7 | 1.4 | 1.4 | 1.4 | 1.4 | 1.4 |
| Parasciurus alleni | 3 | 1.5 | 1.5 | 0 | 0 | 0 |
| Microtus mexicanus | 342 | 112.86 | 112.86 | 0 | 112.86 | 0 |
| Neotoma albigula | 16 | 4 | 4 | 4 | 4 | 0 |
| Neotoma mexicana | 44 | 8.8 | 8.8 | 8.8 | 8.8 | 8.8 |
| Peromyscus difficilis | 46 | 11.5 | 11.5 | 11.5 | 11.5 | 0 |
| Peromyscus levipes | 83 | 20.75 | 20.75 | 20.75 | 0 | 20.75 |
| Peromyscus labecula | 53 | 13.25 | 13.25 | 13.25 | 0 | 13.25 |
| Reithrodontomys megalotis | 110 | 36.3 | 36.3 | 0 | 36.3 | 0 |
| Sylvilagus floridanus | 3 | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 |
| TOTAL | 782 | 274.46 | 214.46 | 64.13 | 175.79 | 48.63 |
Table 8.
Habitat weighting values for composite stratum 720 (~28,000 14C years BP) from San Josecito Cave using birds and habitat preferences.
Table 8.
Habitat weighting values for composite stratum 720 (~28,000 14C years BP) from San Josecito Cave using birds and habitat preferences.
| Taxa | Coniferous Forest | Oak Forest | Xeric Scrub | Grassland | Tropical Forest | |
| Habitat weighting using presence data | ||||||
| Aquila chrysaetos | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | |
| Asio otus | 0.5 | 0 | 0 | 0.5 | 0 | |
| Ciccaba virgata | 0.33 | 0.33 | 0 | 0 | 0.33 | |
| Corvus corax | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | |
| Cyrtonyx montezumae | 0.33 | 0.33 | 0 | 0.33 | 0 | |
| Gymnorhinus cyanocephalus | 1 | 0 | 0 | 0 | 0 | |
| Megascops asio/kennicottii | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | |
| Pluvialis cf. P. squatarola | 0 | 0 | 0 | 1 | 0 | |
| Rallus limicola | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | |
| Sturnella sp. | 0 | 0 | 0 | 1 | 0 | |
| TOTAL | 2.96 | 1.46 | 0.8 | 3.63 | 1.13 | |
| Habitat weighting using presence and NISP data | ||||||
| Taxa | NISP | Coniferous Forest | Oak Forest | Xeric Scrub | Grassland | Tropical Forest |
| Aquila chrysaetos | 75 | 15 | 15 | 15 | 15 | 15 |
| Asio otus | 107 | 0.5 | 0 | 0 | 0.5 | 0 |
| Ciccaba virgata | 20 | 6.6 | 6.6 | 0 | 0 | 6.6 |
| Corvus corax | 1051 | 15 | 15 | 15 | 15 | 15 |
| Cyrtonyx montezumae | 85 | 0.33 | 0.33 | 0 | 0.33 | 0 |
| Gymnorhinus cyanocephalus | 1 | 1 | 0 | 0 | 0 | 0 |
| Megascops asio/kennicottii | 7 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Pluvialis cf. P. squatarola | 1 | 0 | 0 | 0 | 1 | 0 |
| Rallus limicola | 2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Sturnella sp. | 3 | 0 | 0 | 0 | 20 | 0 |
| TOTAL | 1352 | 38.83 | 37.33 | 30.4 | 52.23 | 37.0 |
Table 9.
Habitat weighting values for composite stratum 720 (~28,000 14C years BP) from San Josecito Cave using squamate reptiles and habitat preferences.
Table 9.
Habitat weighting values for composite stratum 720 (~28,000 14C years BP) from San Josecito Cave using squamate reptiles and habitat preferences.
| Taxa | Coniferous Forest | Oak Forest | Xeric Scrub | Grassland | Tropical Forest | |
| Habitat weighting using presence data | ||||||
| Barisia imbricata | 0.33 | 0.33 | 0 | 0.33 | 0 | |
| Barisia ciliaris | 0.33 | 0.33 | 0 | 0.33 | 0 | |
| Phrynosoma modestum | 0 | 0 | 1 | 0 | 0 | |
| Phrynosoma orbiculare | 0.5 | 0.5 | 0 | 0 | 0 | |
| Sceloporus spp. | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | |
| Tantilla sp. | 0.25 | 0.25 | 0.25 | 0 | 0.25 | |
| Heterodon cf. H. simus | 0.5 | 0 | 0 | 0.5 | 0 | |
| Hypsiglena sp. | 0.25 | 0.25 | 0.25 | 0.25 | 0 | |
| Storeria sp. | 0.5 | 0.5 | 0 | 0 | 0 | |
| Crotalus sp. | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | |
| TOTAL | 3.06 | 2.56 | 1.9 | 1.81 | 0.65 | |
| Habitat weighting using presence and MNI data | ||||||
| Taxa | MNI | Coniferous Forest | Oak Forest | Xeric Scrub | Grassland | Tropical Forest |
| Barisia imbricata | 8 | 2.64 | 2.64 | 0 | 2.64 | 0 |
| Barisia ciliaris | 23 | 7.59 | 7.59 | 0 | 7.59 | 0 |
| Phrynosoma modestum | 1 | 0 | 0 | 1 | 0 | 0 |
| Phrynosoma orbiculare | 2 | 1 | 1 | 0 | 0 | 0 |
| Sceloporus spp. | 4 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 |
| Tantilla sp. | 1 | 0.25 | 0.25 | 0.25 | 0 | 0.25 |
| Heterodon cf. H. simus | 1 | 0.5 | 0 | 0 | 0.5 | 0 |
| Hypsiglena sp. | 1 | 0.25 | 0.25 | 0.25 | 0.25 | 0 |
| Storeria sp. | 1 | 0.5 | 0.5 | 0 | 0 | 0 |
| Crotalus sp. | 1 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| TOTAL | 43 | 13.73 | 13.23 | 2.5 | 11.98 | 1.25 |
| Habitat weighting using presence and NISP data | ||||||
| Taxa | NISP | Coniferous Forest | Oak Forest | Xeric Scrub | Grassland | Tropical Forest |
| Barisia imbricata | 32 | 10.56 | 10.56 | 0 | 10.56 | 0 |
| Barisia ciliaris | 75 | 24.75 | 24.75 | 0 | 24.75 | 0 |
| Phrynosoma modestum | 1 | 0 | 0 | 1 | 0 | 0 |
| Phrynosoma orbiculare | 6 | 3 | 3 | 0 | 0 | 0 |
| Sceloporus spp. | 14 | 2.8 | 2.8 | 2.8 | 2.8 | 2.8 |
| Tantilla sp. | 3 | 0.75 | 0.75 | 0.75 | 0 | 0.75 |
| Heterodon cf. H. simus | 1 | 0.5 | 0 | 0 | 0.5 | 0 |
| Hypsiglena sp. | 6 | 1.5 | 1.5 | 1.5 | 1.5 | 0 |
| Storeria sp. | 61 | 30.5 | 30.5 | 0 | 0 | 0 |
| Crotalus sp. | 41 | 8.2 | 8.2 | 8.2 | 8.2 | 8.2 |
| TOTAL | 240 | 82.56 | 82.06 | 14.25 | 48.31 | 11.75 |
Table 10.
Mammal species assigned to stratum 720 (~28,000 14C years BP) and minimum number of individuals (MNI) based on taphonomic box occurrences. Nomenclature follows that proposed by [52]. An asterisk denotes extinct taxa (*).
Table 10.
Mammal species assigned to stratum 720 (~28,000 14C years BP) and minimum number of individuals (MNI) based on taphonomic box occurrences. Nomenclature follows that proposed by [52]. An asterisk denotes extinct taxa (*).
| Taxa | MNI |
|---|---|
| Sorex altoensis | 60 |
| Leptonycteris nivalis | 12 |
| Myotis californicus | 2 |
| Myotis thysanodes | 7 |
| Equus conversidens * | 1 |
| Microtus mexicanus | 342 |
| Neotoma albigula | 16 |
| Neotoma mexicana | 44 |
| Peromyscus difficilis | 46 |
| Peromyscus labecula | 53 |
| Peromyscus levipes | 83 |
| Reithrodontomy megalotis | 110 |
| Sylvilagus floridanus | 3 |
| Total MNI | 779 |
Table 11.
The San Josecito Cave composite 720 local fauna, dated at ~28,000 14C years BP (MNI are provided for bones from stratum 720). Extinct taxa are denoted by an asterisk (*).
Table 11.
The San Josecito Cave composite 720 local fauna, dated at ~28,000 14C years BP (MNI are provided for bones from stratum 720). Extinct taxa are denoted by an asterisk (*).
| Taxa | Stock’s 57–58 ft Level | Stratum 720 |
|---|---|---|
| Nothrotheriops shastensis * | X | |
| Sorex altoensis | 9 | |
| Desmodus stocki * | X | |
| Leptonycteris nivalis | X | 4 |
| Myotis californicus | 1 | |
| Myotis thysanodes | 2 | |
| Aenocyon dirus * | X | |
| Canis latrans | X | |
| Panthera atrox * | X | |
| Panthera onca | X | |
| Puma concolor | X | |
| Smilodon fatalis * | X | |
| Tremarctos floridanus * | X | |
| Ursus americanus | X | |
| Equus conversidens * | X | 1 |
| Navahoceros fricki * | X | |
| Stockoceros conklingi * | X | |
| Euceratherium * | X | |
| Parasciurus alleni | X | |
| Microtus mexicanus | X | 44 |
| Neotoma albigula | X | 4 |
| Neotoma mexicana | X | 9 |
| Peromyscus difficilis | 8 | |
| Peromyscus labecula | 9 | |
| Peromyscus levipes | X | 16 |
| Reithrodontomy megalotis | X | 16 |
| Sylvilagus floridanus | X | 2 |
| Aquila chrysaetos | X | |
| Cyrtonyx montezumae | 85 | |
| Rallus limicola | 2 | |
| Pluvialis cf. squatarola. | X | |
| Megascops asio/kennicottii | 7 | |
| Asio otus | X | 107 |
| Ciccaba virgata | X | |
| Gymnorhinus cyanocephalus | 1 | |
| Corvus corax | X | |
| Sturnella sp. | 3 | |
| Barisia ciliaris | 23 | |
| Barisia imbricata | 8 | |
| Phrynosoma modestum | 1 | |
| Phrynosoma orbiculare | X | 2 |
| Sceloporus sp. | 4 | |
| Tantilla sp. | 1 | |
| Heterodon cf. simus | 1 | |
| Hypsiglena sp. | 1 | |
| Storeria sp. | 1 | |
| Crotalus spp. | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Arroyo-Cabrales, J.; Johnson, E.; Cruz, J.A. San Josecito Cave and Its Paleoecological Contributions for Quaternary Studies in Mexico. Quaternary 2021, 4, 34. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4040034
AMA Style
Arroyo-Cabrales J, Johnson E, Cruz JA. San Josecito Cave and Its Paleoecological Contributions for Quaternary Studies in Mexico. Quaternary. 2021; 4(4):34. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4040034
Chicago/Turabian StyleArroyo-Cabrales, Joaquin, Eileen Johnson, and J. Alberto Cruz. 2021. "San Josecito Cave and Its Paleoecological Contributions for Quaternary Studies in Mexico" Quaternary 4, no. 4: 34. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4040034