
Multi-Taxa Neo-Taphonomic Analysis of Bone Remains from Barn Owl Pellets and Cross-Validation of Observations: A Case Study from Dominica (Lesser Antilles)

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
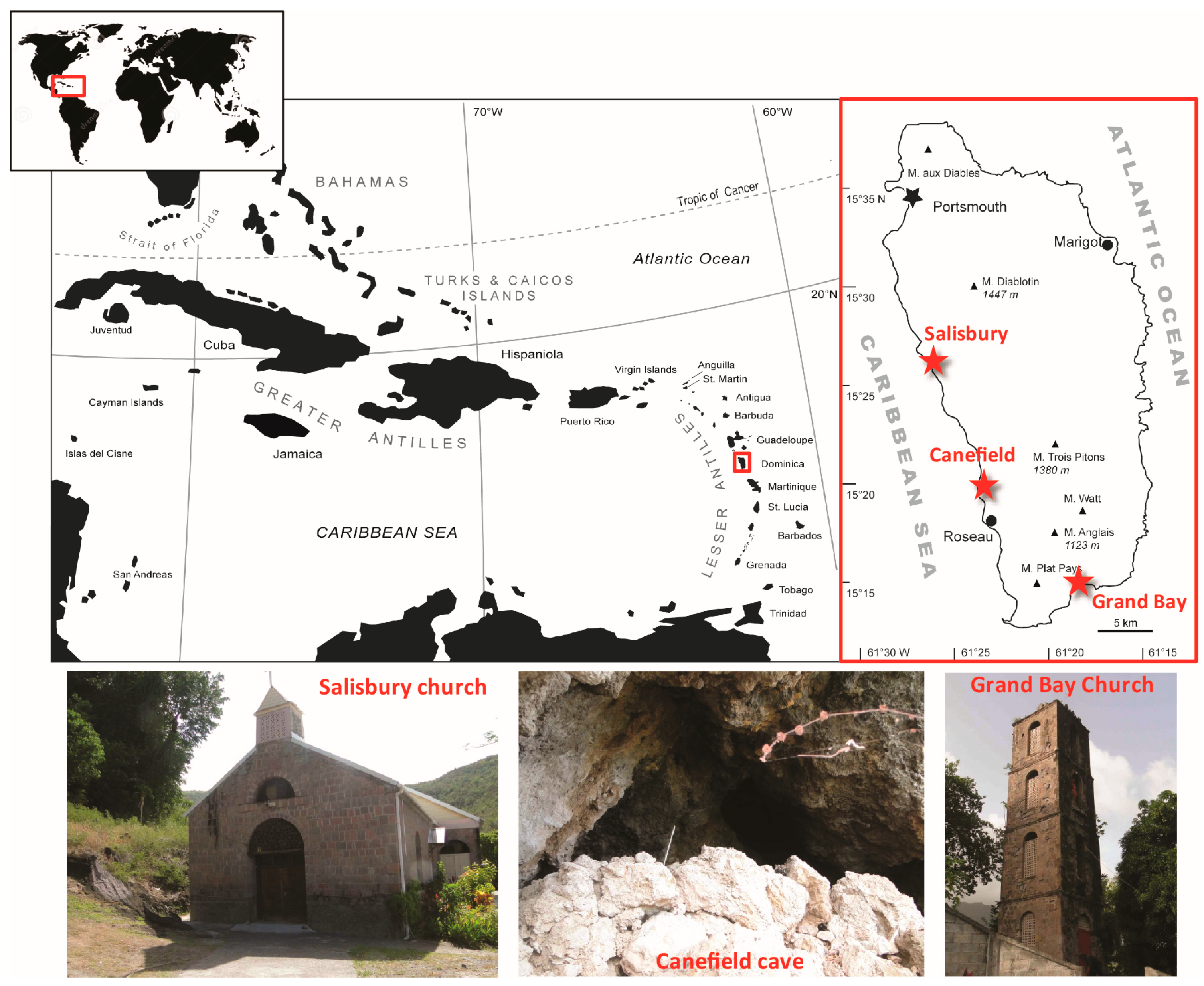
2.1. Owl Pellets Sampling
2.2. Prey Identification
2.3. Taphonomic Analysis
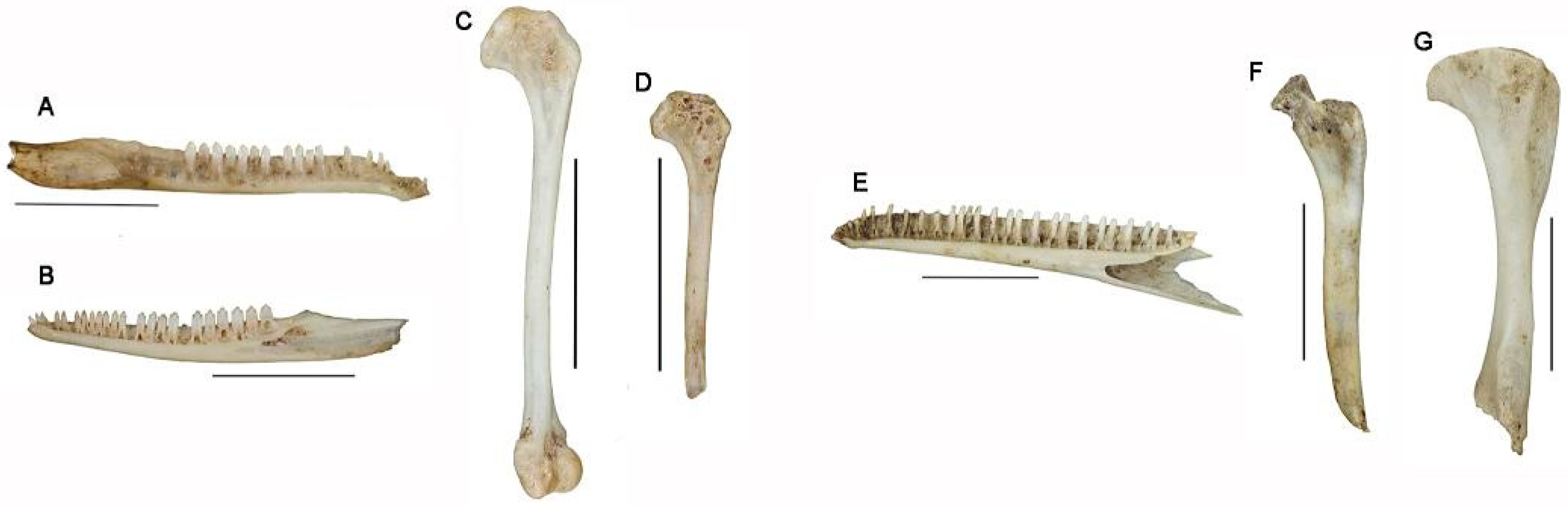
2.3.1. Anatomical Representation
2.3.2. Fragmentation
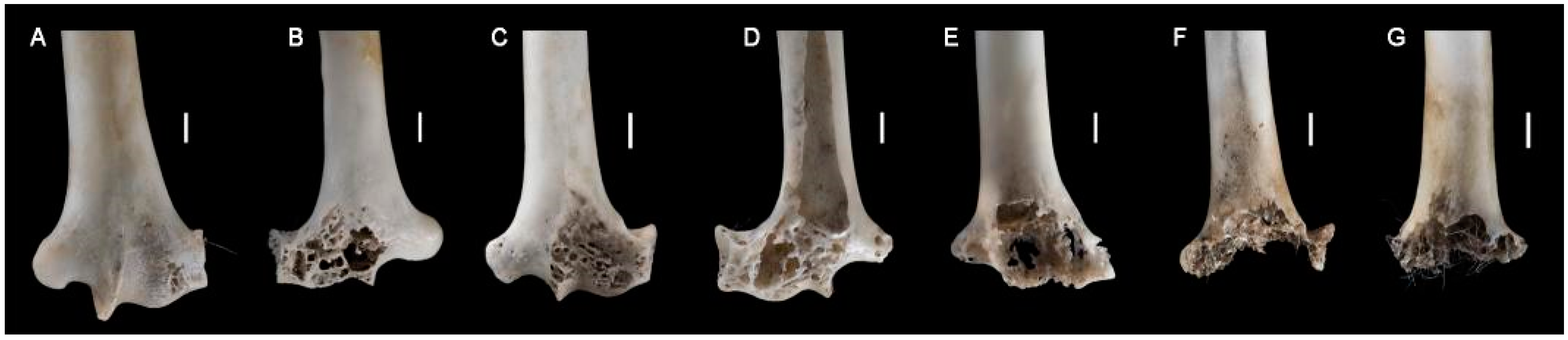
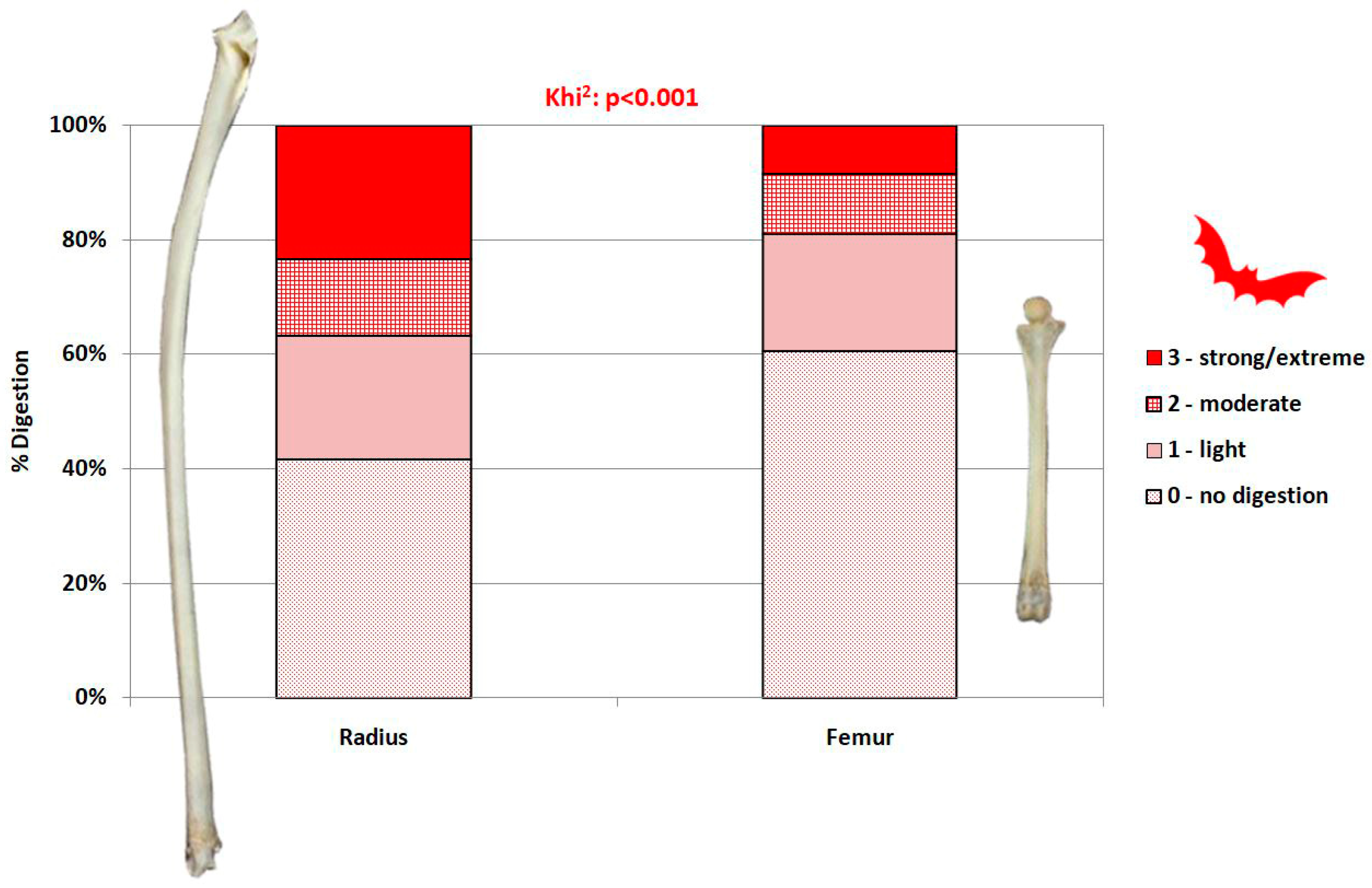
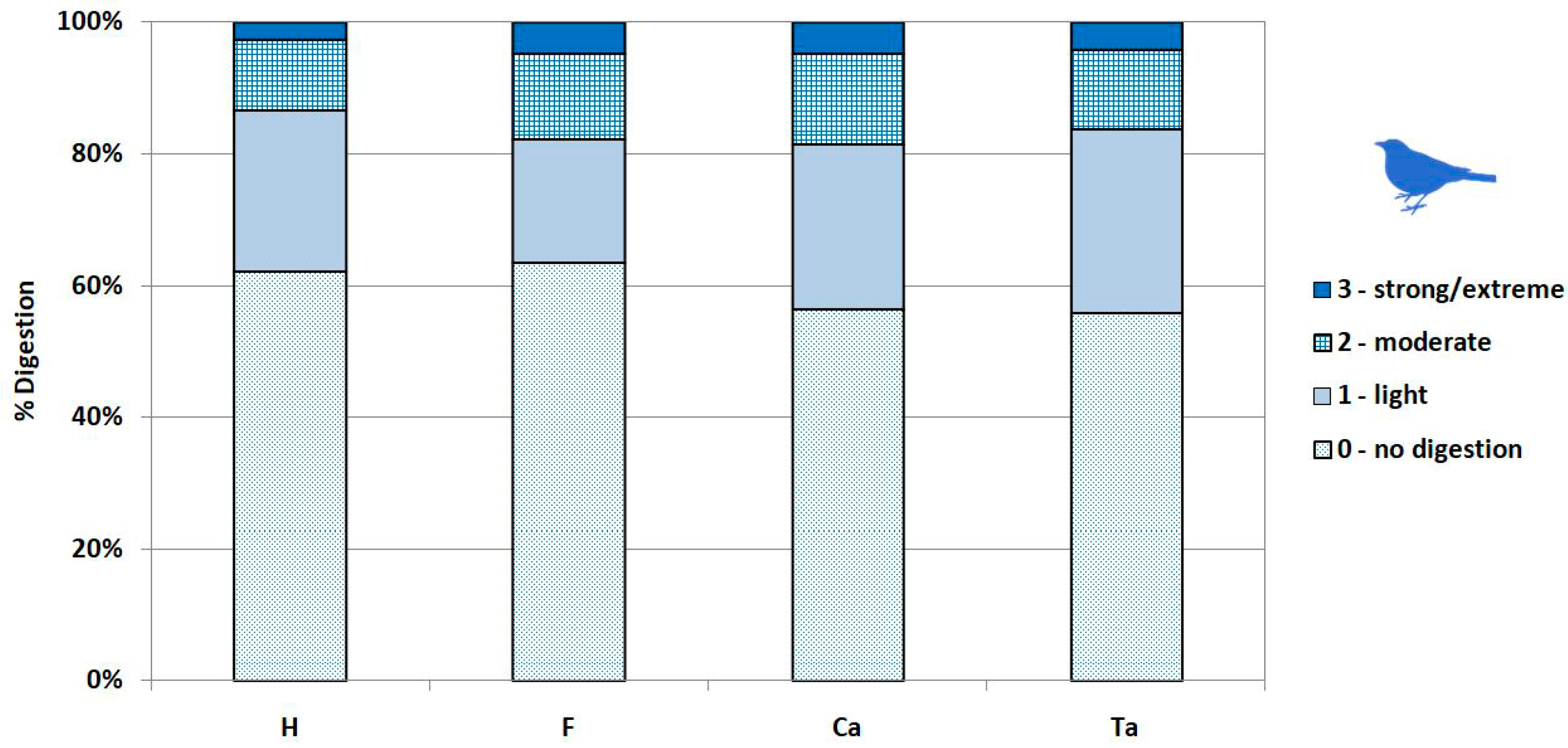
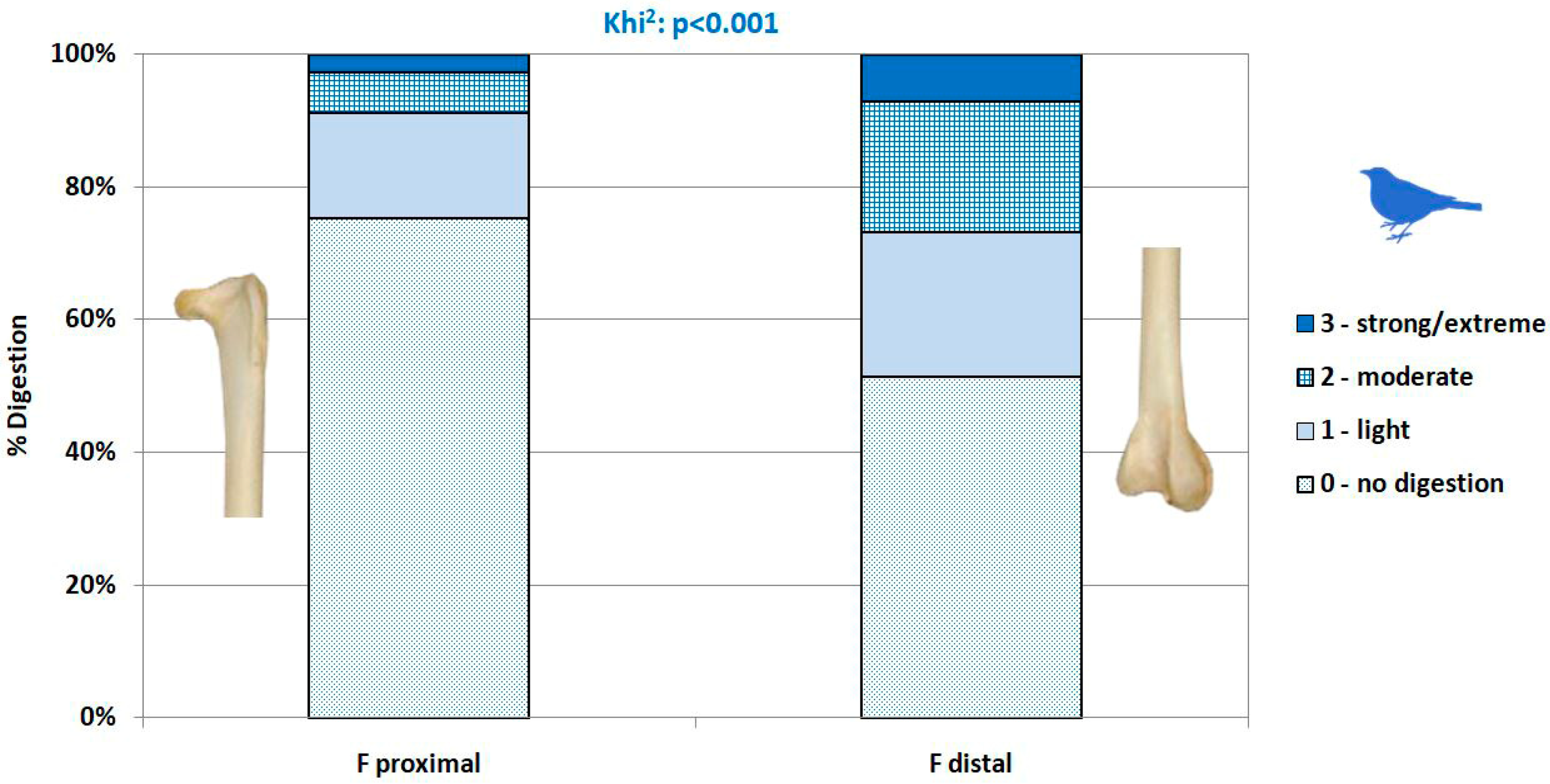
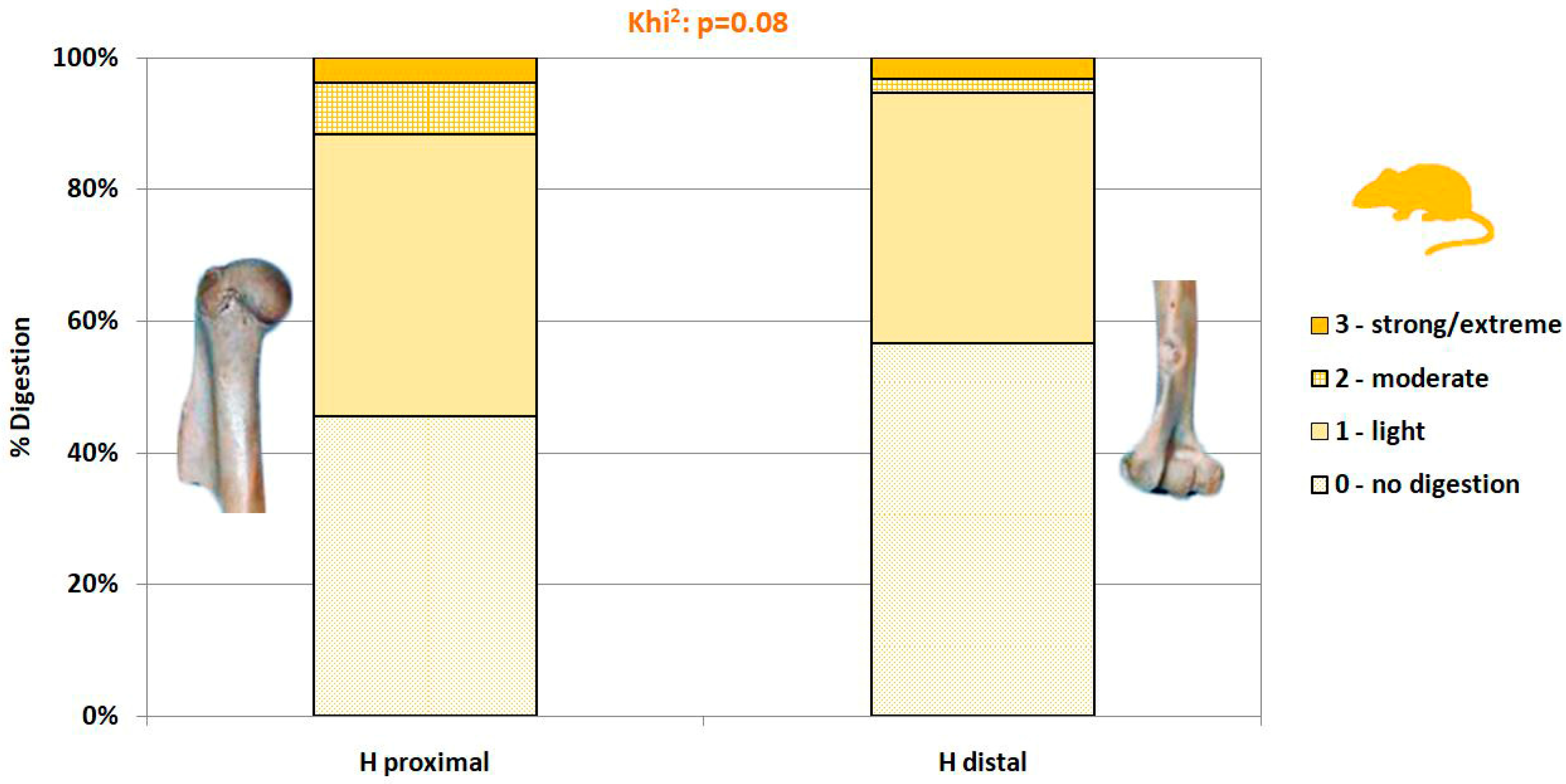
2.3.3. Surface Modifications
- Not digested.
- Slight digestion. On long bones: polished/rounded articular surfaces, cortical bone slightly dissolved and substantia spongiosa slightly visible. On mandibles/dentaries: weak alteration of the anterior part of the bone, but the shape of the bone remains weakly or not modified.
- Moderate digestion. On long bones: substantia spongiosa much more exposed with holes in the cortical bone, but not necessarily on the whole articular surface; the global form of the extremity is preserved. On mandibles/dentaries: strong alteration and partial destruction of the anterior part and alteration of the surface of the corpus of the mandible, the shape of the bone and teeth (mammals, lizards) are modified.
- Strong to extreme digestion. On long bones: cortical bone totally dissolved and substantia spongiosa totally exposed or even dissolved; the extremity can be totally destroyed. On mandibles/dentaries: more than one third of the bone is destroyed, teeth (mammals, lizards) are altered.
2.3.4. Size/Weight Classes of Preys
- Size category 0: below 10 g (the gecko Sphaerodactylus sp.; birds such as small Passeriformes (Parulidae) and hummingbirds (Trochilidae)).
- Size category 1: 10 to 100 g (small lizards of Anolis genus; birds such as Columbina passerina, Salatator albicollis, Mimus gilvus; Mus musculus, young Rattus spp., all identified bat species).
- Size category 2: 100 to 300 g (lizards such as large Anolis spp., medium size Pholidoscelis spp., Leiocephalus sp. and newborn Iguana sp.; birds such as Margarops fuscatus and largers; adult Rattus rattus, young Rattus norvegicus).
2.4. Cross-Validation of Observations
3. Results
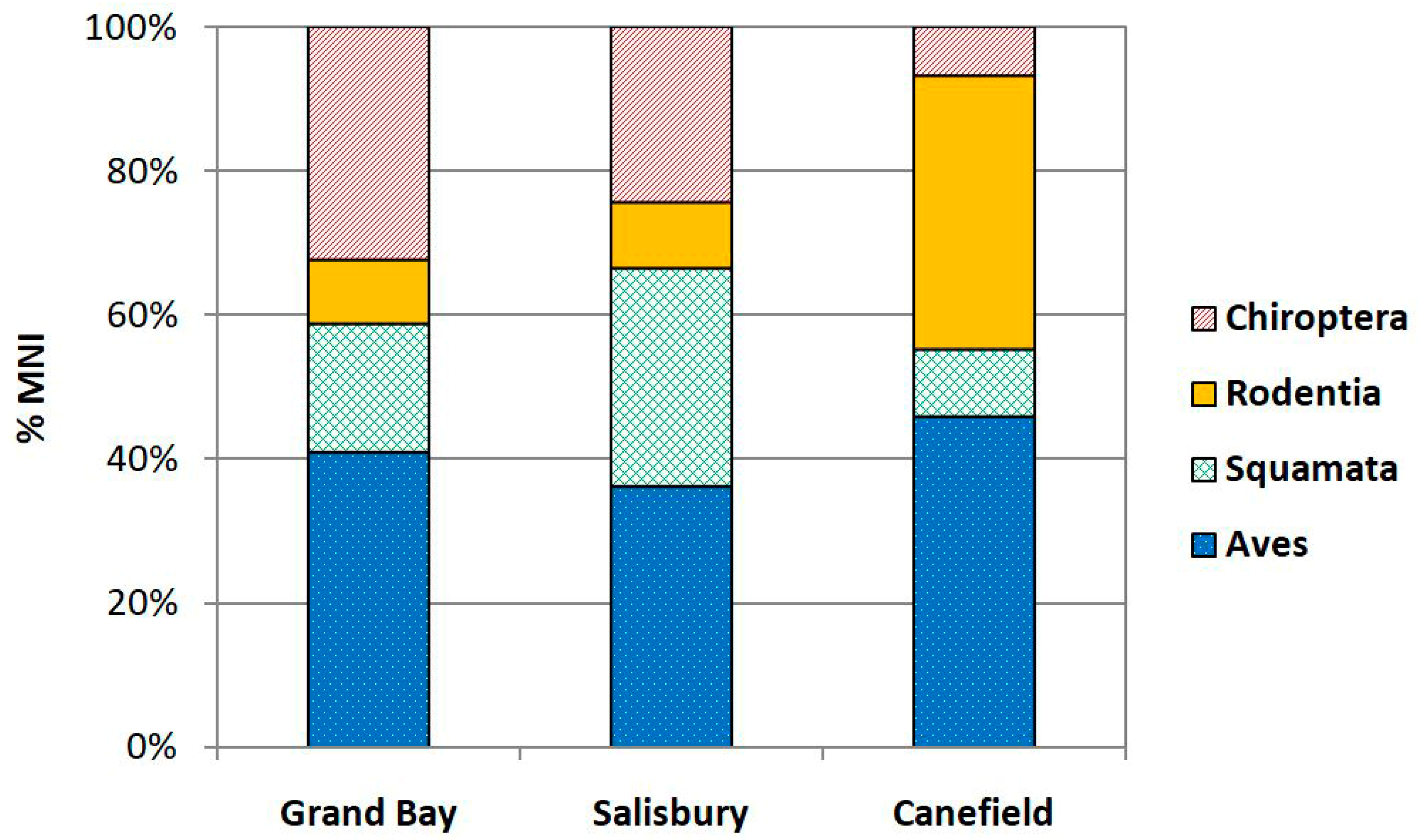
3.1. Faunal Spectrum
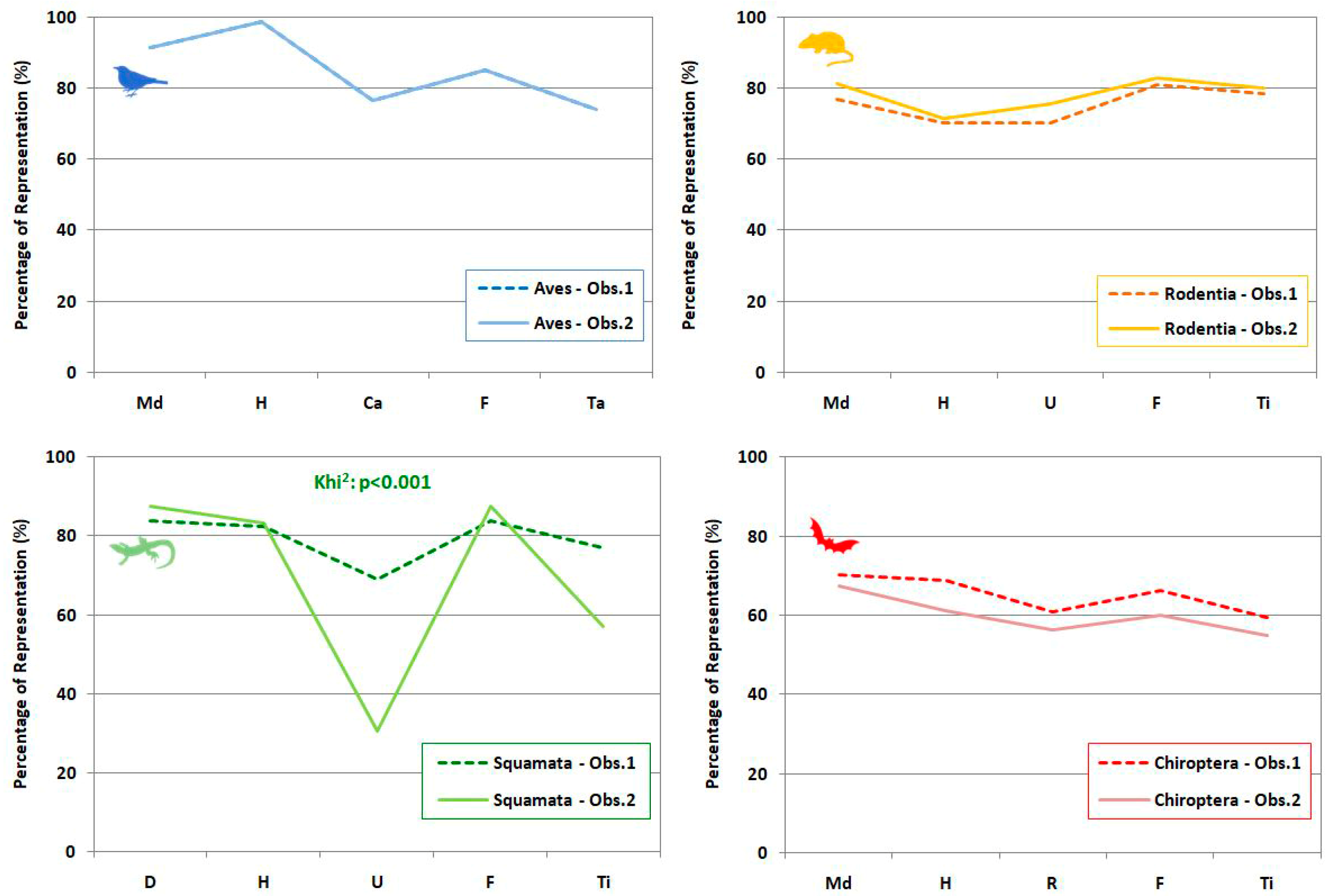
3.2. Anatomical Representation
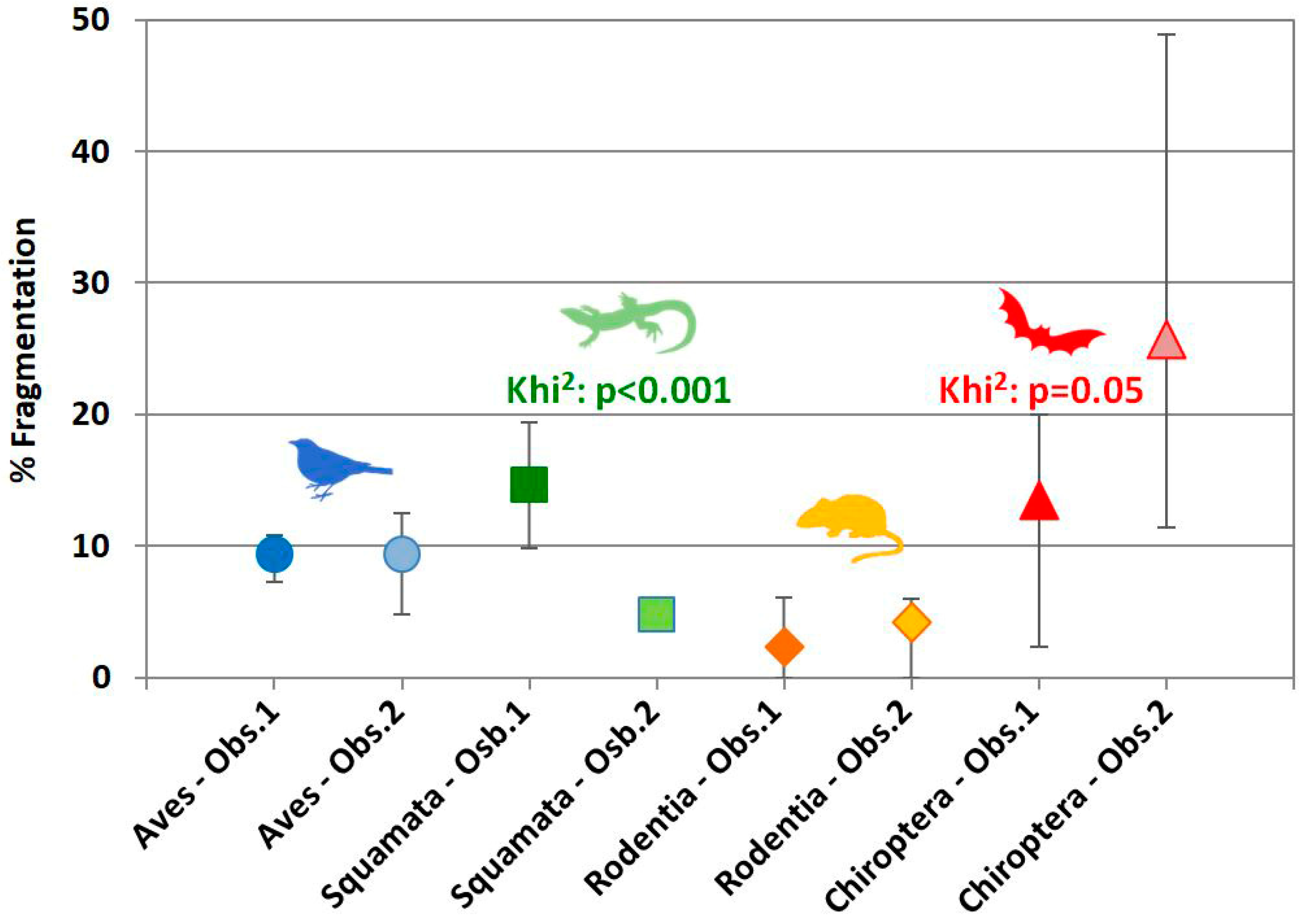
3.3. Fragmentation
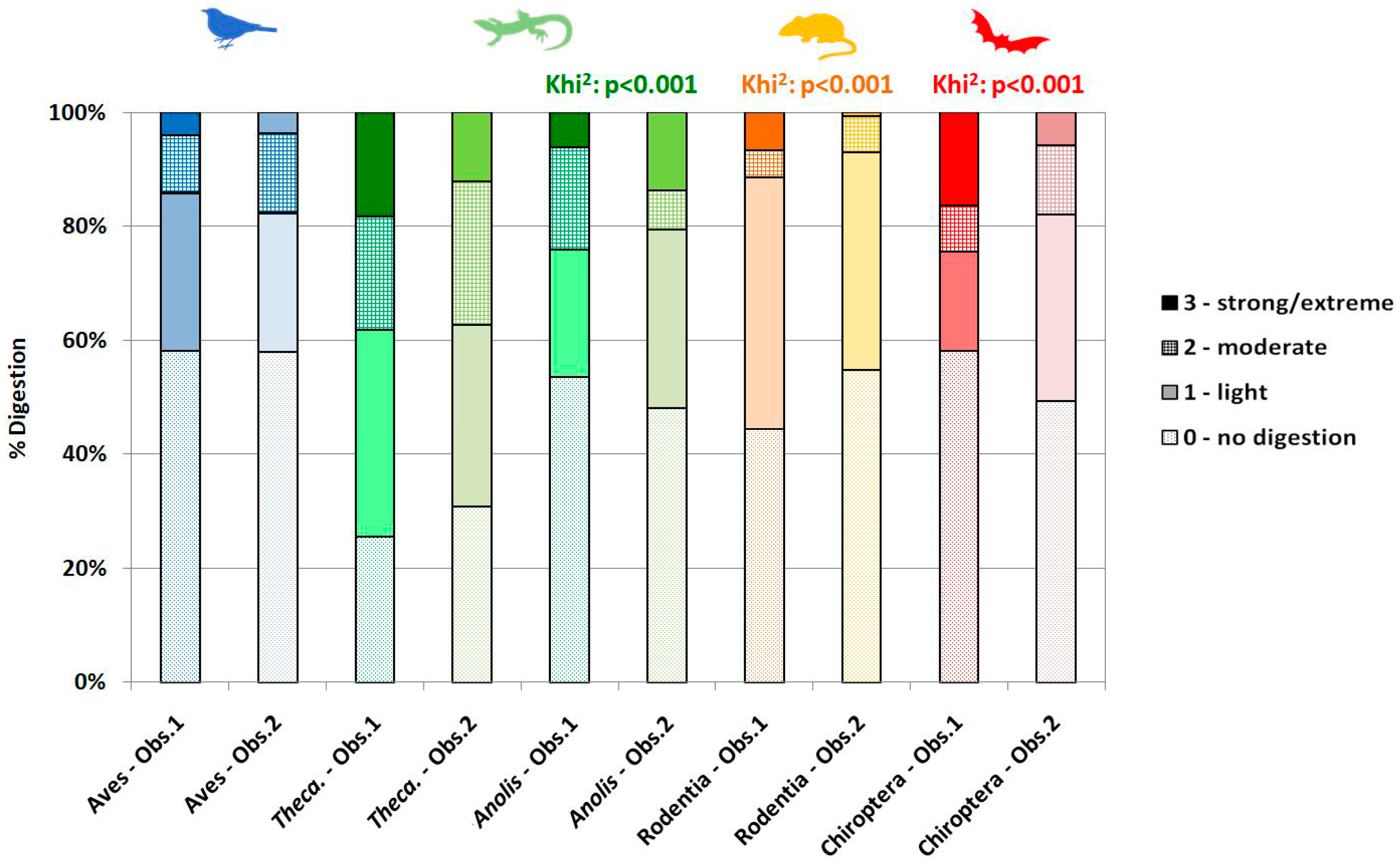
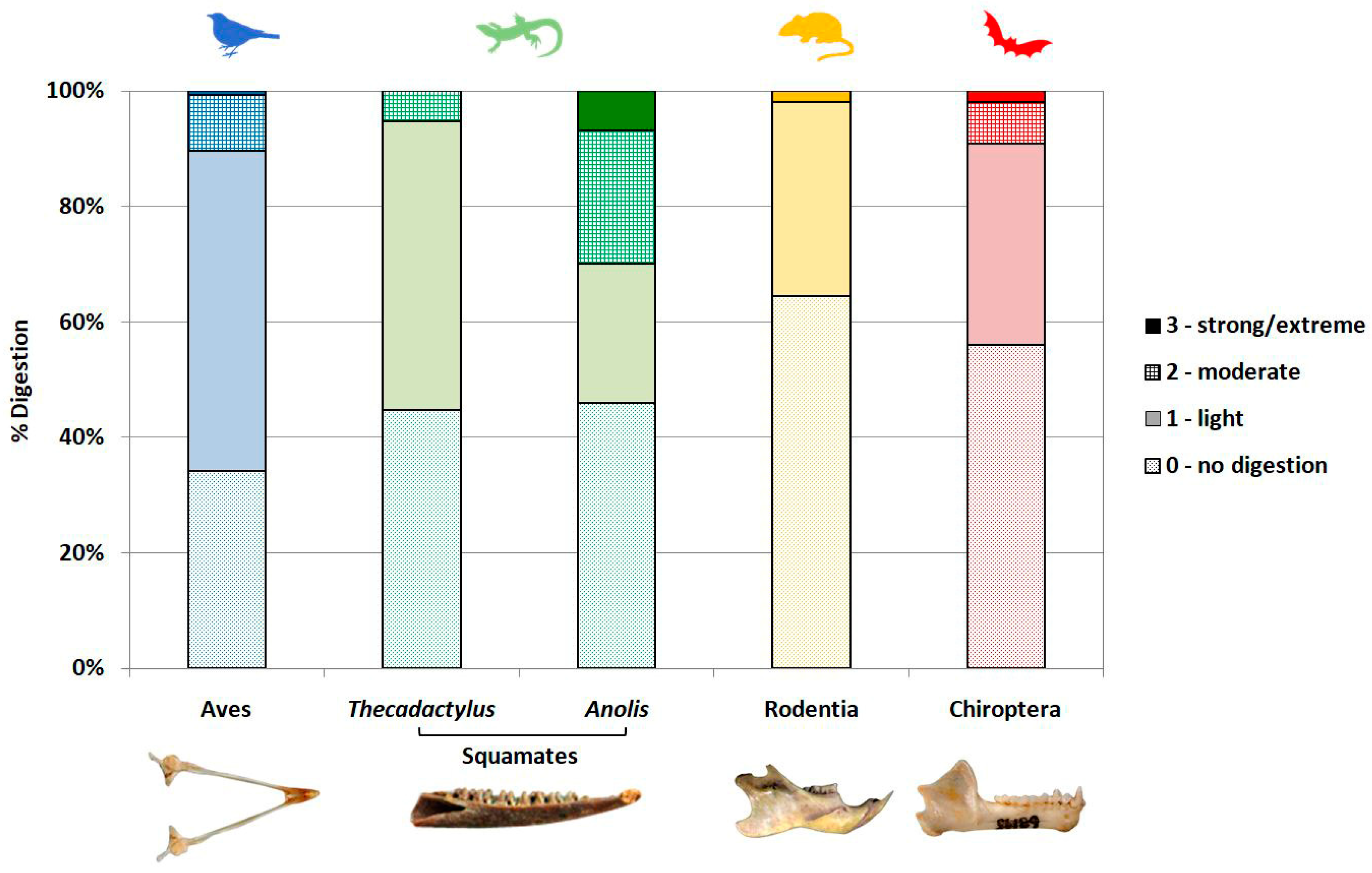
3.4. Modifications of Bone Surface
4. Discussion
4.1. Diet of Tyto Insularis in Dominica
4.2. Taphonomic Impact of Tyto Insularis on Small Vertebrate Bone Assemblage
4.2.1. Remarks on the Size/Weight Classes of Preys
4.2.2. Anatomical Representation
4.2.3. Fragmentation
4.2.4. Digestion
4.3. Degree of Inter-Observer Differences and Potential Outcomes
4.4. Towards an “Inter-Taxa Calibration”
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ridgway, R. Birds of North and Middle America. Bull. U.S. Nat. Mus. 1914, 50, 594–825. [Google Scholar]
- Wetmore, A.; Swales, B.H. The Birds of Haiti and the Dominican Republic. Bull. U.S. Nat. Mus. 1931, 155, 1–483. [Google Scholar] [CrossRef]
- Weick, F. Owls (Strigiformes): Annotated and Illustrated Checklist; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Suarez, W.; Olson, S.L. Systematics and distribution of the living and fossil small barn owls of the West Indies (Aves: Strigiformes: Tytonidae). Zootaxa 2020, 4830, 544–564. [Google Scholar] [CrossRef] [PubMed]
- Avibase. The World Bird Database; Lepage, D., Ed.; Available online: https://avibase.bsc-eoc.org/avibase.jsp (accessed on 15 July 2021).
- Johnston, D.W. Food of the Barn Owl on Grand Cayman, BWI. Q. J. Fla. Acad. Sci. 1972, 35, 171–172. Available online: https://0-www-jstor-org.brum.beds.ac.uk/stable/i24313839 (accessed on 20 July 2021).
- Buden, D.W. Prey remains of Barn Owls in the southern Bahama Islands. Wils. Bull. 1974, 86, 336–343. [Google Scholar]
- McFarlane, D.A.; Garrett, K.L. The prey of common Barn Owls (Tyto alba) in dry limestone scrub forest of southern Jamaica. Caribb. J. Sci. 1989, 25, 21–23. [Google Scholar]
- Arredondo, C.; Chirino, V.N. Consideraciones sobre la alimentación de Tyto alba furcata (aves: Strigiformes) con implicaciones ecológicas en Cuba. J. Caribb. Ornithol. 2002, 15, 16–24. [Google Scholar]
- Hernández-Munoz, A.; Mancina, C.A. La dieta de la lechuza (Tyto alba) (Aves: Strigiformes) en hábitats naturales y antropogénicos de la región central de Cuba. Rev. Mex. Biodiv. 2011, 82, 217–226. [Google Scholar] [CrossRef]
- López-Ricardo, Y. Alimentacion de la Lechuza (Tyto Alba Furcata) en Cuba Central: Presas Introducias y Autoctonas. Tesis de diploma, Universidad de La Habana, La Habana, Cuba, 2012. [Google Scholar]
- Stoetzel, E.; Fraysse, A.; Bochaton, C.; Gala, M.; Grouard, S.; Lenoble, A.; Denys, C. Diet of the Lesser Antillean barn owl Tyto insularis (Aves: Strigiformes) in Dominica, Lesser Antilles. Caribb. J. Sci. 2016, 49, 91–100. [Google Scholar] [CrossRef]
- Stoetzel, E.; Royer, A.; Cochard, D.; Lenoble, A. Late Quaternary changes in bat palaeobiodiversity and palaeobiogeography under climatic and anthropogenic pressure: New insights from Marie-Galante, Lesser Antilles. Quat. Sci. Rev. 2016, 143, 150–174. [Google Scholar] [CrossRef]
- Andrews, P. Owls, Caves and Fossils; Natural History Museum Publications: London, UK, 1990. [Google Scholar]
- Denys, C.; Stoetzel, E.; Andrews, P.; Bailon, S.; Rihane, A.; Huchet, J.B.; Fernandez-Jalvo, Y.; Laroulandie, V. Taphonomy of small predators accumulations: About palaeoecological implications. Hist. Biol. 2018, 30, 868–881. [Google Scholar] [CrossRef]
- Fernández Jalvo, Y.; Andrews, P.; Denys, C.; Sese, C.; Stoetzel, E.; Marin-Monfort, D.; Pesquero, D. Taphonomy for taxonomists: Implications of predation in small mammal studies. Quat. Sci. Rev. 2016, 139, 138–157. [Google Scholar] [CrossRef]
- Lorvelec, O.; Pascal, M.; Delloupe, X.; Chapuis, J.L. Les mammifères terrestres non volants des Antilles françaises et l’introduction récente d’un écureuil. Rev. Écol. 2007, 62, 295–314. Available online: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&ved=2ahUKEwim0qDX7tvzAhXZ6OAKHWhZBJcQFnoECAIQAw&url=https%3A%2F%2Fecureuils.mnhn.fr%2Fsites%2Fdefault%2Ffiles%2Fdocuments%2Florvelec-et-al-rev-ecol-terr-vie.pdf&usg=AOvVaw20M02pZpFMS8O_t8aGb4gX (accessed on 20 July 2021).
- Giovas, C.M. The beasts at large—Perennial questions and new paradigms for caribbean translocation research. Part II: Mammalian introductions in cultural context. Environ. Archaeol. 2018, 24, 294–305. [Google Scholar] [CrossRef]
- Kemp, M.E.; Mychajliw, A.M.; Wadman, J.; Goldberg, A. 7000 years of turnover: Historical contingency and human niche construction shape the Caribbean’s Anthropocene biota. Proc. R. Soc. B 2020, 287, 20200447. [Google Scholar] [CrossRef]
- Rabinow, S.; Giovas, C. A systematic review of agouti (Dasyproctidae: Dasyprocta) records from the pre-1492 Lesser Antilles: New perspectives on an introduced commensal. Int. J. Osteoarchaeol. 2021, 31, 758–769. [Google Scholar] [CrossRef]
- Mallye, J.B.; Cochard, D.; Laroulandie, V. Accumulation osseuse en périphérie de terriers de petits carnivores: Les stigmates de prédation et de fréquentation. Ann. Paléontol. 2008, 3, 187–208. [Google Scholar] [CrossRef]
- Royer, A.; Montuire, S.; Gilg, O.; Laroulandie, V. A taphonomic investigation of small vertebrate accumulations produced by the snowy owl (Bubo scandiacus) and its implications for fossil studies. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 514, 189–205. [Google Scholar] [CrossRef]
- Lebreton, L.; Bailon, S.; Guilllaud, E.; Testu, A.; Perrenoud, C. Multi-taxa referential of a modern Eurasian Eagle-Owl (Bubo bubo) aerie. J. Archaeol. Sci. Rep. 2020, 32, 102417. [Google Scholar] [CrossRef]
- Erome, G.; Aulagnier, S. Contribution à l’identification des proies des Rapaces. Bièvre 1982, 4, 129–135. [Google Scholar]
- Radulet, N. Comparative anatomy of the mandible in the mammal systematics (Mammalia: Insectvora, Chiroptera, Rodentia) from Romania (II). Trav. Mus. Natl. Hist. Nat. Grigore Antipa 2006, XLIX, 365–374. [Google Scholar]
- Radulet, N. Morphology of the inner side of the mandible in micromammals (Mammalia: Insectivora, Chiroptera, Rodentia) of Romania. Trav. Mus. Natl. Hist. Nat. Grigore Antipa 2007, L, 371–393. [Google Scholar]
- Fraysse, A. Contribution à L’identification Morphologique et Moléculaire des Chiroptères des Petites Antilles. Master’s Thesis, Muséum National d’Histoire Naturelle, Paris, France, 13 September 2013. [Google Scholar]
- Ibéné, B.; Leblanc, F.; Houllemare, C.; Tarlier, C.; Pentier, C. Contribution à L’étude des Chiroptères de la Guadeloupe; Diren Guadeloupe: Basse-Terre, France, 2007. [Google Scholar]
- Arendt, W.J.; Faaborg, J.; Wallace, G.E.; Garrido, O.H. Biometrics of birds throughout the Greater Caribbean Basin. Proc. West. Found. Vert. Zool. 2004, 8, 1–33. [Google Scholar]
- Bochaton, C.; Bailon, S.; Herrel, A.; Grouard, S.; Ineich, I.; Tresset, A.; Cornette, R. Human impacts reduce morphological diversity in an insular species of lizard. Proc. R. Soc. B 2017, 284, 20170921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bochaton, C.; Daza, J.D.; Lenoble, A. Identifying gecko species from Lesser Antillean paleontological assemblages: Intraspecific osteological variation within and interspecific osteological differences between Thecadactylus rapicauda (Houttuyn, 1782) (Phyllodactylidae) and Hemidactylus mabouia (Moreau de Jonnès, 1818) (Gekkonidae). J. Herpetol. 2018, 52, 313–320. [Google Scholar] [CrossRef]
- Lloveras, L.; Moreno-Garcia, M.; Nadal, J.; Thomas, R. Blind test evaluation of accuracy in the identification and quantification of digestion corrosion damage on leporid bones. Quat. Int. 2014, 330, 150–155. [Google Scholar] [CrossRef]
- Goedert, J.; Cochard, D.; Lenoble, A.; Lorvelec, O.; Pisanu, B.; Royer, A. Seasonal demography of different black rat (Rattus rattus) populations under contrasting natural habitats in Guadeloupe (Lesser Antilles, Caribbean). Mamm. Res. 2020, 65, 793–804. [Google Scholar] [CrossRef]
- Flikweert, M.; Prins, T.G.; de Freitas, J.A.; Nijman, V. Spatial variation in the diet of the Barn Owl Tyto alba in the Caribbean. Ardea 2007, 95, 75–82. [Google Scholar] [CrossRef]
- Wiley, J.W. Food habits of the endemic ashy-faced owl (Tyto glaucops) and recently arrived barn owl (T. alba) in Hispaniola. J. Raptor Res. 2010, 44, 87–100. [Google Scholar] [CrossRef]
- Lopez, J.M. Actualistic taphonomy of barn owl pellet-derived small mammal bone accumulations in arid environments of South America. J. Quat. Sci. 2020, 35, 1057–1069. [Google Scholar] [CrossRef]
- Linchamps, P.; Stoetzel, E.; Hanon, R.; Denys, C. Neotaphonomic study of two Tyto alba assemblages from Botswana: Paleoecological implications. J. Archaeol. Sci. Rep. 2021, 38, 103085. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anatomical Representation | Ratios | ||||
|---|---|---|---|---|---|
| Cranial Elements | Post-Cranial Elements | Cranial /Post-Cranial * | Anterior /Posterior | Stylopodia /Zeugopodia | |
| Aves | Mandible (Md) | Humerus (H), Carpometacarpus (Ca), Femur (F), Tarsometatarsus (Ta) | Md/(H + Ca + F + Ta) | (H + Ca)/(F + Ta) | (H + F)/(Ca + Ta) |
| Squamata | Dentary (D) | Humerus (H), Ulna (U), Femur (F), Tibia (Ti) | D/(H + U+F + Ti) | (H + U)/(F + Ti) | (H + F)/(U + Ti) |
| Rodentia | Mandible (Md) | Humerus (H), Ulna (U), Femur (F), Tibia (Ti) | Md/(H + U+F + Ti) | (H + U)/(F + Ti) | (H + F)/(U + Ti) |
| Chiroptera | Mandible (Md) | Humerus (H), Radius (R), Femur (F), Tibia (Ti) | Md/(H + R+F + Ti) | (H + R)/(F + Ti) | (H + F)/(R + Ti) |
| Prey Taxa | Grand Bay | Salisbury | Canefield | TOTAL | ||||
|---|---|---|---|---|---|---|---|---|
| NISP | MNI | NISP | MNI | NISP | MNI | NISP | MNI | |
| Aves | 186.0 | 23.0 | 206.0 | 24.0 | 295.0 | 34.0 | 687.0 | 81.0 |
| Squamata | 75.5 | 9.5 | 159.0 | 20.0 | 36.5 | 7.3 | 271.0 | 36.0 |
| Rodentia | 27.0 | 5.0 | 39.0 | 5.5 | 207.5 | 27.5 | 273.5 | 38.0 |
| Chiroptera | 102.0 | 18.0 | 117.0 | 15.5 | 21.0 | 5.0 | 240.0 | 38.5 |
| TOTAL | 390.5 | 55.5 | 521.0 | 65.0 | 560.0 | 73.8 | 1471.5 | 193.5 |
| Size Classes | Grand Bay | Salisbury | Canefield | TOTAL | ||||
|---|---|---|---|---|---|---|---|---|
| NISP | NMI | NISP | NMI | NISP | NMI | NISP | NMI | |
| 0 (<10 g) | 107.0 | 13.0 | 71.5 | 9.0 | 119.0 | 14.0 | 297.5 | 36.0 |
| 1 (10–100 g) | 275.5 | 40.5 | 447.5 | 55.0 | 431.0 | 57.0 | 1154.0 | 152.5 |
| 2 (100–300 g) | 8.0 | 2.0 | 2.0 | 0.5 | 10.0 | 3.0 | 20.0 | 5.5 |
| TOTAL | 390.5 | 55.5 | 521.0 | 64.5 | 560.0 | 74.0 | 1471.5 | 194.0 |
| Cranial/Post-Cranial | Anterior/Posterior | Stylopodia/Zeugopodia | |
|---|---|---|---|
| Aves | 1.09 | 1.10 | 1.22 |
| Squamata | 1.20 | 0.87 | 1.44 |
| Rodentia | 1.04 | 0.89 | 1.00 |
| Chiroptera | 1.13 | 1.03 | 1.11 |
| <30% Digested | 30–50% Digested | 50–80% Digested | >80% Digested | |
|---|---|---|---|---|
| Aves | Hum, CMC, Fem, TMT | Md | ||
| Squamata (Thecadactylusrapicauda) | Md, Hum, Ulna, Tib | Fem | ||
| Squamata (Anolis sp.) | Ulna | Tibia | Md, Hum, Fem | |
| Rodentia | Md | Hum | Ulna, Fem, Tib | |
| Chiroptera | Md, Fem, Tib | Hum, Radius |
| Majority Not Digested | Majority Light Digestion | Majority Moderate Digestion | Majority Strong Digestion | |
|---|---|---|---|---|
| Aves | Hum, CMC, Fem, TMT | Md | ||
| Squamata (Thecadactylusrapicauda) | Md, ulna, Tib | Hum | Fem | |
| Squamata (Anolis sp.) | Md, Hum, Ulna, Fem, Tib | |||
| Rodentia | Md, Hum, Tib | Ulna, Fem | ||
| Chiroptera | Md, Hum, Ulna, Fem, Tib | Radius |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoetzel, E.; Bochaton, C.; Bailon, S.; Cochard, D.; Gala, M.; Laroulandie, V. Multi-Taxa Neo-Taphonomic Analysis of Bone Remains from Barn Owl Pellets and Cross-Validation of Observations: A Case Study from Dominica (Lesser Antilles). Quaternary 2021, 4, 38. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4040038
Stoetzel E, Bochaton C, Bailon S, Cochard D, Gala M, Laroulandie V. Multi-Taxa Neo-Taphonomic Analysis of Bone Remains from Barn Owl Pellets and Cross-Validation of Observations: A Case Study from Dominica (Lesser Antilles). Quaternary. 2021; 4(4):38. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4040038
Chicago/Turabian StyleStoetzel, Emmanuelle, Corentin Bochaton, Salvador Bailon, David Cochard, Monica Gala, and Véronique Laroulandie. 2021. "Multi-Taxa Neo-Taphonomic Analysis of Bone Remains from Barn Owl Pellets and Cross-Validation of Observations: A Case Study from Dominica (Lesser Antilles)" Quaternary 4, no. 4: 38. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4040038