Drought Increases Vulnerability of Pinus ponderosa Saplings to Fire-Induced Mortality

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pinus Ponderosa Saplings and Study Treatments
2.2. Physiological Measurements
2.3. Bud Development and Sapling Mortality
2.4. Statistical Analysis
3. Results
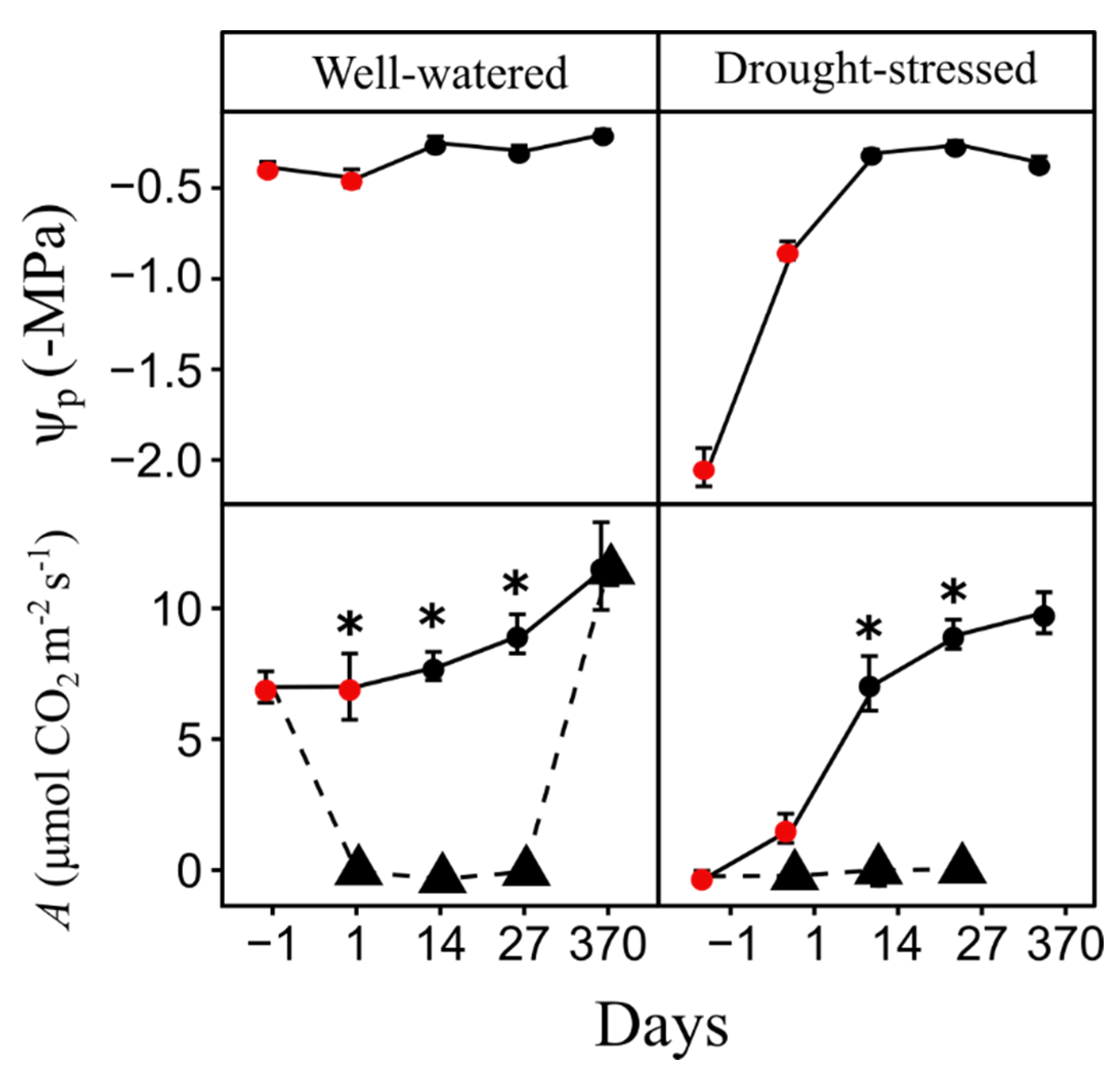
3.1. Sapling Physiology
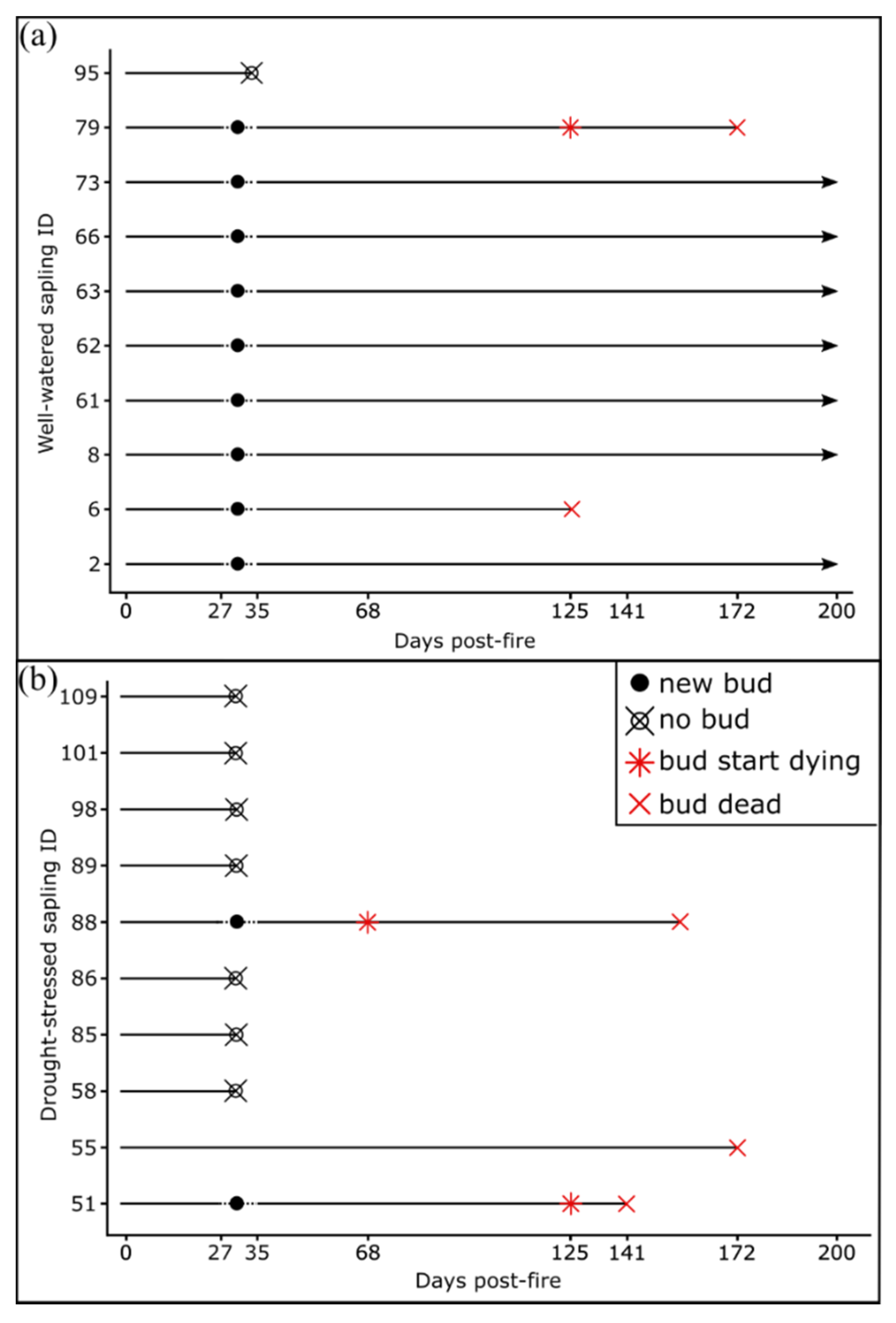
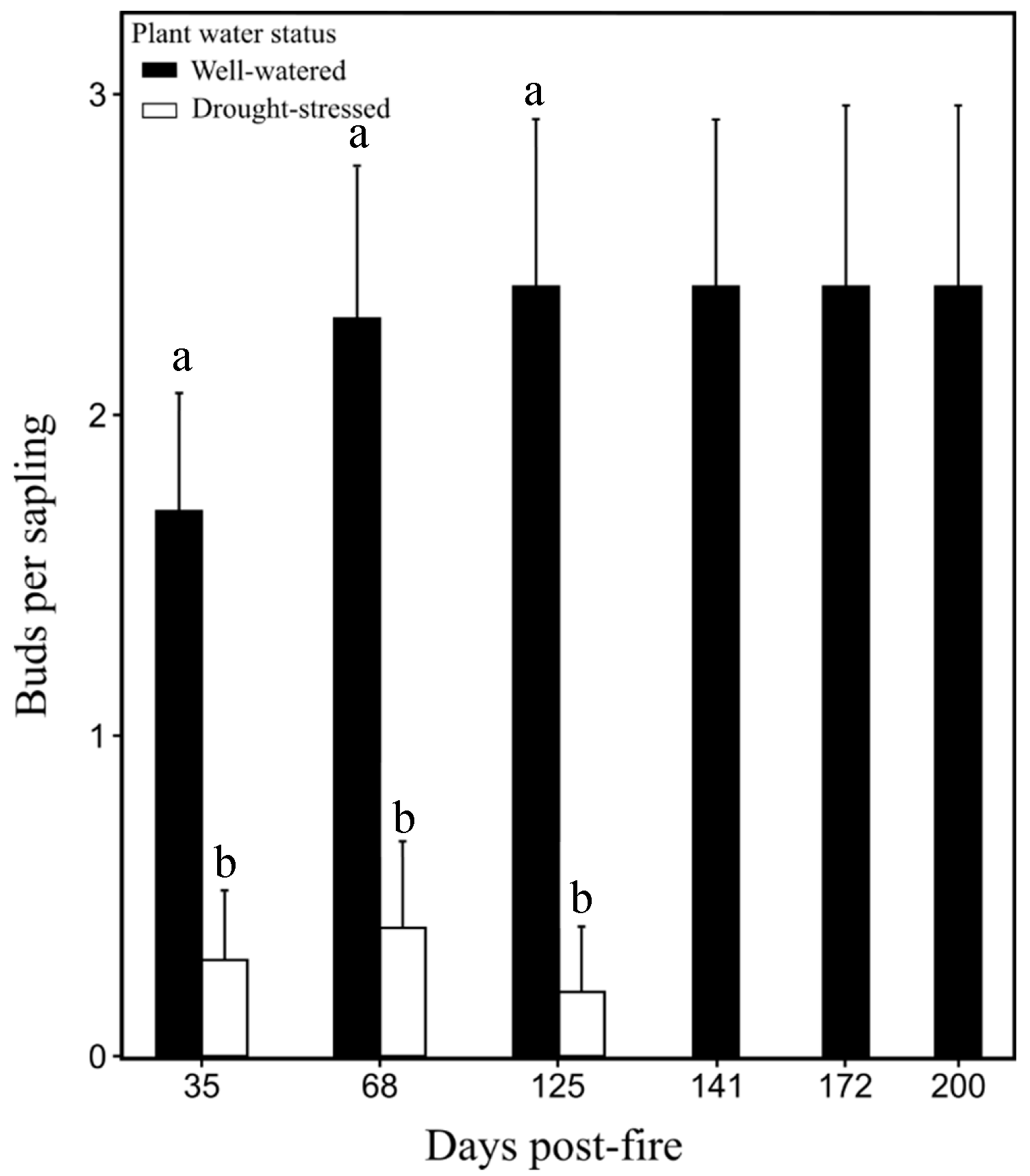
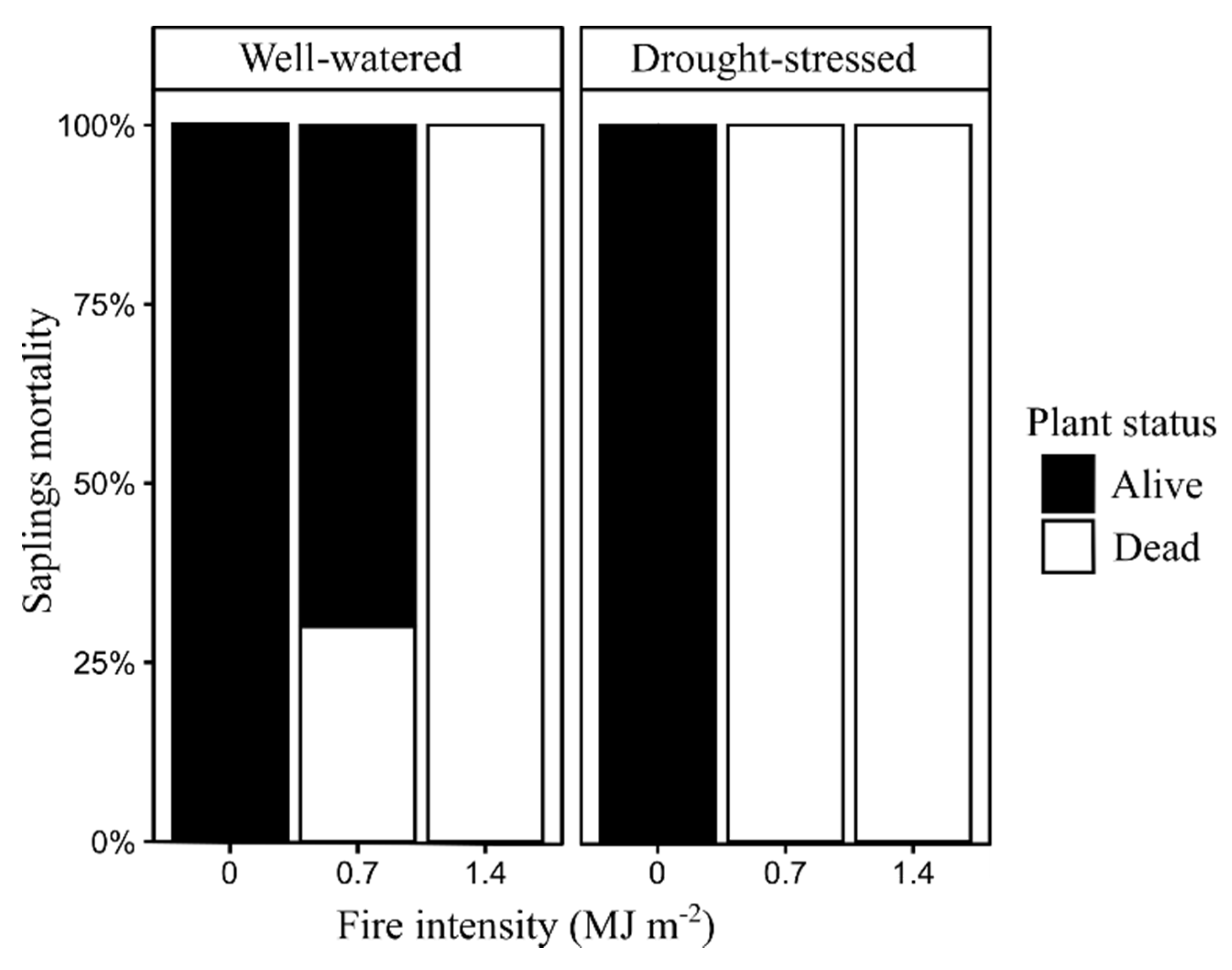
3.2. Bud Development and Sapling Mortality
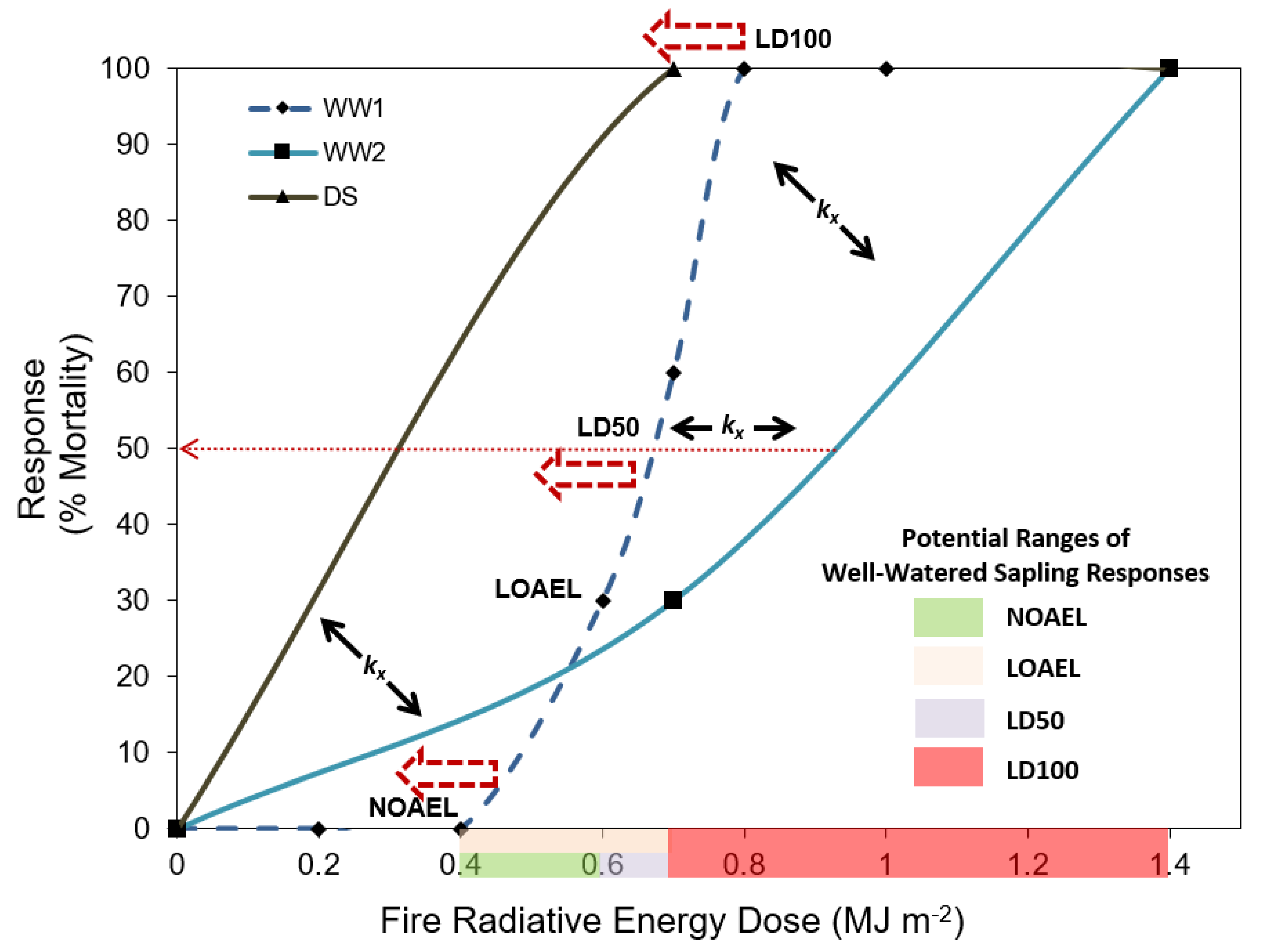
4. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J.; et al. Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, T.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2009, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Adams, H.D.; Luce, C.H.; Breshears, D.D.; Weiler, M.; Hale, C.H.; Allen, C.D.; Smith, A.M.S.; Huxman, T.E. Ecohydrological consequences of drought- and infestation-triggered tree die-off. Ecohydrology 2012, 5, 145–159. [Google Scholar] [CrossRef]
- Barbero, R.; Abatzoglou, J.T.; Larkin, N.K.; Kolden, C.A.; Stocks, B. Climate change presents increased potential for very large fires in the contiguous United States. Int. J. Wildland Fire 2015, 24, 892–899. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Williams, A.P. Impact of anthropogenic climate change on wildfire across western US forests. Proc. Natl. Acad. Sci. USA 2016, 113, 11770–11775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, D.M.J.S.; Williamson, G.; Kolden, C.A.; Abatzoglou, J.T.; Cochrane, M.A.; Smith, A.M.S. Human exposure and sensitivity to globally extreme wildfire events. Nat. Ecol. Evol. 2017, 1, 58. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L. Temperate forest health in an era of emerging mega disturbance. Science 2015, 349, 823–826. [Google Scholar] [CrossRef]
- Davis, K.T.; Dobrowski, S.Z.; Higuera, P.E.; Holden, Z.A.; Veblen, T.T.; Rother, M.T.; Parks, S.A.; Sala, A.; Maneta, M. Wildfires and climate change push low-elevation forests across a critical climate threshold for tree regeneration. Proc. Natl. Acad. Sci. USA 2019, 116, 6193–6198. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, E.J.; Sturtevant, B.R. Modeling forest mortality caused by drought stress: Implications for climate change. Ecosystems 2013, 16, 60–74. [Google Scholar] [CrossRef]
- Reinhardt, E.D.; Keane, R.E.; Brown, J.K. First Order Fire Effects Model: FOFEM 4.0 User’s Guide INT-GTR-344 US Department of Agriculture; Forest Service, Intermountain Research Station: Ogden, UT, USA, 1997. [Google Scholar]
- Rebain, S.A. The Fire and Fuels Extension to the Forest Vegetation Simulator: Updated Model Documentation; Internal Rep. U.S. Department of Agriculture 2015, Forest Service, Forest Management Service Center: Fort Collins, CO, USA, 2015; p. 403.
- Lutes, D. FOFEM 6.5 First Order Fire Effects Model User Guide, Fire and Aviation Management, Rocky Mountain Research Station Fire Modelling Institute; United States Department of Agriculture: Fort Collins, CO, USA, 2018; p. 86.
- Woolley, T.; Shaw, D.C.; Ganio, L.M.; Fitzgerald, S. A review of logistic regression models used to predict post-fire tree mortality of western North American conifers. Int. J. Wildland Fire 2012, 21, 1–35. [Google Scholar] [CrossRef]
- Smith, A.M.S.; Sparks, A.M.; Kolden, C.A.; Abatzoglou, J.T.; Talhelm, A.F.; Johnson, D.M.; Boschetti, L.; Lutz, J.A.; Apostol, K.G.; Yedinak, K.M.; et al. Towards a new paradigm in fire severity research using dose-response experiments. Int. J. Wildland Fire 2016, 25, 158–166. [Google Scholar] [CrossRef]
- Hood, S.M.; Varner, J.M.; van Mantgem, P.; Cansler, C.A. Fire and tree death: Understanding and improving modeling of fire-induced tree mortality. Environ. Res. Lett. 2018, 13, 113004. [Google Scholar] [CrossRef]
- Pfeiffer, M.; Spessa, A.; Kaplan, J.O. A model for global biomass burning in preindustrial time: LPJ-LMfire (v1. 0). Geosci. Model Dev. 2013, 6, 643–685. [Google Scholar] [CrossRef] [Green Version]
- Thonicke, K.; Spessa, A.; Prentice, I.C.; Harrison, S.P.; Dong, L.; Carmona-Moreno, C. The influence of vegetation, fire spread and fire behaviour on biomass burning and trace gas emissions: Results from a process-based model. Biogeosciences 2010, 7, 1991–2011. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, D.M.; Fisher, R.A.; Koven, C.D.; Oleson, K.W.; Swenson, S.C.; Bonan, G.; Collier, N.; Ghimire, B.; van Kampenhout, L.; Kennedy, D.; et al. The Community Land Model Version 5: Description of New Features, Benchmarking, and Impact of Forcing Uncertainty. J. Adv. Modeling Earth Syst. 2019, 11, 4245–4287. [Google Scholar] [CrossRef] [Green Version]
- Adam, J.C.; Stephens, J.C.; Chung, S.H.; Brady, M.P.; Evans, R.D.; Kruger, C.E.; Lamb, B.K.; Liu, M.; Stökle, C.O.; Vaughan, J.K.; et al. BioEarth: Envisioning and developing a new regional earth system model to inform natural and agricultural resource management. Clim. Chang. 2015, 129, 555–571. [Google Scholar] [CrossRef]
- Zierl, B.; Bugmann, H.; Tague, C.L. Water and carbon fluxes of European ecosystems: An evaluation of the ecohydrological model RHESSys. Hydrol. Process. 2007, 21, 3328–3339. [Google Scholar] [CrossRef]
- Bart, R.R.; Kennedy, M.C.; Tague, C.L.; McKenzie, D. Integrating fire effects on vegetation carbon cycling within an ecohydrologic model. Ecol. Model. 2000, 416, 108880. [Google Scholar] [CrossRef]
- Jolly, W.M.; Johnson, D.M. Pyro-Ecophysiology: Shifting the Paradigm of Live Wildland Fuel Research. Fire 2018, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.J.; Hiers, J.K.; Varner, J.M.; Hoffman, C.M.; Dickinson, M.B.; Michaletz, S.T.; Loudermilk, E.L.; Butler, B.W. Advances in Mechanistic Approaches to Quantifying Biophysical Fire Effects. Curr. For. Rep. 2018, 4, 161–177. [Google Scholar] [CrossRef]
- Smith, A.M.S.; Talhelm, A.F.; Johnson, D.M.; Sparks, A.M.; Yedinak, K.M.; Apostol, K.G.; Tinkham, W.T.; Kolden, C.A.; Abatzoglou, J.T.; Lutz, J.A.; et al. Effects of fire radiative energy density intensity on Pinus contorta and Larix occidentalis seedling physiology and mortality. Int. J. Wildland Fire 2017, 26, 82–94. [Google Scholar] [CrossRef] [Green Version]
- Sparks, A.M.; Kolden, C.A.; Talhelm, A.F.; Smith, A.M.S.; Apostol, K.G.; Johnson, D.M.; Boschetti, L. Spectral indices accurately quantify changes in seedling physiology following fire: Towards mechanistic assessments of post-fire carbon cycling. Remote Sens. 2016, 8, 572. [Google Scholar] [CrossRef] [Green Version]
- Sparks, A.M.; Smith, A.M.S.; Talhelm, A.F.; Kolden, C.A.; Yedinak, K.M.; Johnson, D.M. Impacts of fire radiative flux on mature Pinus ponderosa growth and vulnerability to secondary mortality agents. Int. J. Wildland Fire 2017, 26, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Sparks, A.M.; Talhelm, A.F.; Partelli-Feltrin, R.; Smith, A.M.S.; Johnson, D.M.; Kolden, C.A.; Boschetti, L. An experimental assessment of the impact of drought and fire on western larch mortality and recovery. Int. J. Wildland Fire 2018, 27, 490–497. [Google Scholar] [CrossRef] [Green Version]
- Sparks, A.M.; Kolden, C.A.; Smith, A.M.S.; Boschetti, L.; Johnson, D.M.; Cochrane, M.A. Fire intensity impacts on post-fire response of temperate coniferous forest net primary productivity. Biogeosciences 2018, 15, 1173–1183. [Google Scholar] [CrossRef] [Green Version]
- Steady, W.D.; Partelli-Feltrin, R.; Johnson, D.M.; Sparks, A.M.; Kolden, C.A.; Talhelm, A.F.; Lutz, J.A.; Boschelli, L.; Hudak, A.T.; Nelson, A.S.; et al. The survival of Pinus ponderosa saplings subjected to increasing levels of fire behavior and impacts on post-fire growth. Fire 2019, 2, 23. [Google Scholar] [CrossRef] [Green Version]
- Partelli-Feltrin, R.; Smith, A.M.S.; Adams, H.D.; Kolden, C.A.; Johnson, D.M. Short- and long-term effects of fire on stem hydraulics in Pinus ponderosa saplings. Plant. Cell Environ. 2020. [Google Scholar] [CrossRef]
- Bär, A.; Michaeltz, S.T.; Mayr, S. Fire effects on tree physiology. New Phytol. 2019, 223, 1728–1741. [Google Scholar] [CrossRef] [Green Version]
- Cochard, H.; Coll, L.; Roux, X.L.; Améglio, T. Unraveling the effects of plant hydraulics on stomatal closure during water stress in walnut. Plant. Physiol. 2002, 128, 282–290. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.G.; Beerling, D.J.; Breshears, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Grieu, P.; Guehl, J.M.; Aussenac, G. The effects of soil and atmospheric drought on photosynthesis and stomatal control of gas exchange in three coniferous species. Physiol. Plant. 1988, 73, 97–104. [Google Scholar] [CrossRef]
- Rouhi, V.; Samson, R.; Lemeur, R.; Damme, P.V. Photosynthetic gas exchange characteristics in three different almond species during drought stress and subsequent recovery. Environ. Exp. Bot. 2007, 59, 117–129. [Google Scholar] [CrossRef]
- Adams, H.D.; Zeppel, M.J.B.; Anderegg, W.R.L.; Hartmann, H.; Landhaüsser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Hartmann, H.; Adams, H.D.; Zhang, H.; Jin, C.; Zhao, C.; Guan, D.; Wang, A.; Yuan, F.; Wu, J. The sweet side of global change-dynamic responses of non-structural carbohydrates to drought, elevated CO2 and nitrogen fertilization in tree species. Tree Physiol. 2018, 38, 1–18. [Google Scholar] [CrossRef]
- Michaletz, S.T.; Johnson, E.A. How forest fires kill trees: A review of the fundamental biophysical processes. Scand. J. For. Res. 2007, 22, 500–515. [Google Scholar] [CrossRef]
- Kavanagh, K.L.; Dickinson, M.B.; Bova, A.S. A way forward for fire-caused tree mortality prediction: Modeling a physiological consequence of fire. Fire Ecol. 2010, 6, 80–94. [Google Scholar] [CrossRef]
- Michaletz, S.T.; Johnson, E.A.; Tyree, M.T. Moving beyond the cambium necrosis hypothesis of post-fire tree mortality: Cavitation and deformation of xylem in forest fires. New Phytol. 2012, 194, 254–263. [Google Scholar] [CrossRef]
- West, A.G.; Nel, J.A.; Bond, W.J.; Midgley, J.J. Experimental evidence for heat plume-induced cavitation and xylem deformation as a mechanism of rapid post-fire tree mortality. New Phytol. 2016, 211, 828–838. [Google Scholar] [CrossRef] [Green Version]
- Bär, A.; Nardini, A.; Mayr, S. Post-fire effects in xylem hydraulics of Picea abies, Pinus sylvestris and Fagus sylvatica. New Phytol. 2018, 217, 1484–1493. [Google Scholar] [CrossRef] [Green Version]
- Ryan, K.C.; Frandsen, W.H. Basal injury from smoldering fires in mature Pinus ponderosa Laws. Int. J. Wildland Fire 1991, 2, 107–118. [Google Scholar] [CrossRef]
- Dickinson, M.B.; Johnsonm, E.A. Temperature-dependent rate models of vascular cambium cell mortality. Can. J. For. Res. 2004, 34, 546–559. [Google Scholar] [CrossRef]
- Kleinman, J.S.; Goode, J.D.; Fries, A.C.; Hart, J.L. Ecological consequences of compound disturbances in forest ecosystems: A systematic review. Ecosphere 2019, 10, e02962. [Google Scholar] [CrossRef] [Green Version]
- van Mantgem, P.J.; Nesmith, J.C.B.; Keifer, M.; Knapp, E.E.; Flint, A.; Flint, L. Climatic stress increases forest fire severity across the western United States. Ecol. Lett. 2013, 16, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Van Nieuwstadt, M.G.L.; Sheil, D. Drought, fire and tree survival in a Borneo rain forest, East Kalimantan, Indonesia. J. Ecol. 2005, 93, 191–201. [Google Scholar] [CrossRef]
- Brando, P.M.; Balch, J.K.; Nepstad, D.C.; Morton, D.C.; Putz, F.E.; Coe, M.T.; Silverio, D.; Macedo, M.N.; Davidson, E.A.; Nobrega, C.C.; et al. Abrupt increases in Amazonian tree mortality due to drought–fire interactions. Proc. Natl. Acad. Sci. USA 2014, 111, 6347–6352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Iorio, A.; Montagnoli, A.; Scippa, G.S.; Chiatante, D. Fine root growth of Quercus pubescens seedlings after drought stress and fire disturbance. Environ. Exp. Bot. 2011, 74, 272–279. [Google Scholar] [CrossRef]
- Chiatante, D.; Tognetti, R.; Scippa, G.S.; Congiu, T.; Baesso, B.; Terzaghi, M.; Montagnoli, A. Interspecific variation in functional traits of oak seedlings (Quercus ilex, Quercus trojana, Quercus virgiliana) grown under artificial drought and fire conditions. J. Plant. Res. 2015, 128, 595–611. [Google Scholar] [CrossRef]
- van Mantgem, P.J.; Caprio, A.C.; Stephenson, N.L.; Das, A.J. Does prescribed fire promote resistance to drought in low-elevation forests of the Sierra Nevada, California, USA? Fire Ecol. 2016, 12, 13–25. [Google Scholar] [CrossRef]
- Smith, A.M.S.; Kolden, C.A.; Bowman, D.M.J.S. Biomimicry can help humans to sustainably coexist with fire. Nat. Ecol. Evol. 2018, 2, 1827–1829. [Google Scholar] [CrossRef]
- Brodersen, C.R.; Germino, M.J.; Johnson, D.M.; Reinhardt, K.; Smith, W.K.; Resler, L.M.; Bader, M.Y.; Sala, A.; Kueppers, L.M.; Broll, G.; et al. Seedling survival at timberline is critical to conifer mountain forest elevation and extent. Front. For. Glob. Chang. 2019, 2, 9. [Google Scholar] [CrossRef]
- Baker, K.V.; Tai, X.; Miller, M.L.; Johnson, D.M. Six co-occurring conifer species in northern Idaho exhibit a continuum of hydraulic strategies during an extreme drought year. AoB Plants 2019, 11, plz056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wooster, M.J.; Roberts, G.; Smith, A.M.S.; Johnson, J.; Freeborn, P.; Amici, S.; Hudak, A.T. Thermal Remote Sensing of Active Vegetation Fires and Biomass Burning Events, chapter 18. In Thermal Infrared Remote Sensing; Kuenzer, C., Dech, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 978-94-007-6638-9. [Google Scholar]
- Smith, A.M.S.; Tinkham, W.T.; Roy, D.P.; Boschetti, L.; Kumar, S.; Sparks, A.M.; Kremens, R.L.; Falkowski, M.J. Quantification of fuel moisture effects on biomass consumed derived from fire radiative energy retrievals. Geophys. Res. Lett. 2013, 40, 6298–6302. [Google Scholar] [CrossRef]
- Kremens, R.; Smith, A.M.S.; Dickinson, M. Fire Metrology: Current and future directions in physics-based measurements. Fire Ecol. 2010, 6, 13–35. [Google Scholar] [CrossRef]
- Tinkham, W.T.; Smith, A.M.S.; Higuera, P.E.; Hatten, J.A.; Brewer, N.B.; Doerr, S.H. Replacing time with space: Using laboratory fires to explore the effects of repeated burning on black carbon degradation. Int. J. Wildland Fire 2016, 25, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Brewer, N.W.; Smith, A.M.S.; Hatten, J.A.; Higuera, P.E.; Hudak, A.T.; Ottmar, R.D.; Tinkham, W.T. Fuel Moisture Influences on Fire-altered Carbon in Masticated Fuels: An Experimental Study. J. Geophys. Res. 2013, 118, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mäechler, M.; Bolker, B.; Walker, S.; Christensen, R.H.B. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R. Emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.4.3.01. 2019. Available online: https://www.r-project.org/ (accessed on 1 August 2020).
- Agee, J.K.; Wright, C.S.; Williamson, N.; Huff, M.H. Foliar moisture content of Pacific Northwest vegetation and its relation to wildland fire behavior. For. Ecol. Manag. 2002, 167, 57–66. [Google Scholar] [CrossRef]
- McDowell, N.G.; Michaletz, S.T.; Bennett, K.E.; Solander, K.C.; Xu, C.; Maxwell, R.M.; Allen, C.D.; Middleton, R.S. Predicting chronic climate- driven disturbances and their mitigation. Trends Ecol. Evol. 2018, 33, 15–27. [Google Scholar] [CrossRef]
- Flannigan, M.; Stocks, B.J.; Wotton, B.M. Climate change and forest fires. Sci. Total Environ. 2000, 262, 221–229. [Google Scholar] [CrossRef]
- Flannigan, M.; Cantin, A.S.; De Groot, W.J.; Wotton, M.; Newbery, A.; Gowman, L.M. Global wildland fire season severity in the 21st century. For. Ecol. Manag. 2013, 294, 54–61. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Days | Units | Comments |
|---|---|---|---|
| Leaf water potential (Ψp) | −1, 1, 14, 27, 370 | - MPa | Ψp was measured only in unburned saplings. |
| Photosynthesis (A) | −1, 1, 14, 27, 370 | μmol CO2 m−2 s−1 | After saplings were exposed to the fire, A was measured in unburned and burned saplings exposed to 0.7 MJ m−2. |
| Bud development | 27, 35, 68, 125, 141, 172, 200 | Number of buds per sapling | |
| Sapling mortality | 200 | Live/Dead |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Partelli-Feltrin, R.; Johnson, D.M.; Sparks, A.M.; Adams, H.D.; Kolden, C.A.; Nelson, A.S.; Smith, A.M.S. Drought Increases Vulnerability of Pinus ponderosa Saplings to Fire-Induced Mortality. Fire 2020, 3, 56. https://0-doi-org.brum.beds.ac.uk/10.3390/fire3040056
Partelli-Feltrin R, Johnson DM, Sparks AM, Adams HD, Kolden CA, Nelson AS, Smith AMS. Drought Increases Vulnerability of Pinus ponderosa Saplings to Fire-Induced Mortality. Fire. 2020; 3(4):56. https://0-doi-org.brum.beds.ac.uk/10.3390/fire3040056
Chicago/Turabian StylePartelli-Feltrin, Raquel, Daniel M. Johnson, Aaron M. Sparks, Henry D. Adams, Crystal A. Kolden, Andrew S. Nelson, and Alistair M. S. Smith. 2020. "Drought Increases Vulnerability of Pinus ponderosa Saplings to Fire-Induced Mortality" Fire 3, no. 4: 56. https://0-doi-org.brum.beds.ac.uk/10.3390/fire3040056