
Molecular Tools for Diagnosis and Surveillance of Soil-Transmitted Helminths in Endemic Areas


The Wellcome Trust Research Laboratory, Division of Gastrointestinal Sciences, Christian Medical College, Vellore 632004, Tamil Nadu, India
*
Author to whom correspondence should be addressed.
Parasitologia 2021, 1(3), 105-118; https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia1030012
Submission received: 21 April 2021
/
Revised: 1 July 2021
/
Accepted: 2 July 2021
/
Published: 7 July 2021
Abstract
:Soil-transmitted helminths (STH) including the hookworms Necator americanus and Ancylostoma spp., Ascaris lumbricoides, and Trichuris trichiura affect over 1.5 billion people worldwide and are estimated to have caused 1.9 million disability-adjusted life years (DALYs). With the concerted effort in expanding and improving targeted mass drug administration (MDA) programs over the past decade, along with decreasing prevalence, infections in several endemic areas tend to be of low intensity. Conventional microscopy-based methods recommended for the detection of STH in parasitological surveys have been shown to be less sensitive in these low-intensity settings. As communities progress towards STH elimination through MDA and improved sanitation, there is a pressing need for highly sensitive techniques that detect the true prevalence of STH to evaluate the effectiveness of ongoing programs and interventions. Molecular methods that involve analysis of DNA rather than the morphology of the organism are highly sensitive and specific, allowing for both quantitation and species discrimination. The following review discusses different sample collection strategies, pre-processing steps, DNA extraction platforms, and nucleic acid detection methods available for diagnosis and surveillance of STH. We have contrasted the utility of these molecular tools against conventional microscopy-based methods currently used in most endemic settings. While the detection methods are primarily qPCR based, several newer technologies have also become available along with automation and increased throughput, making these molecular tools increasingly cost-effective and potentially amenable for use in low-resource settings.
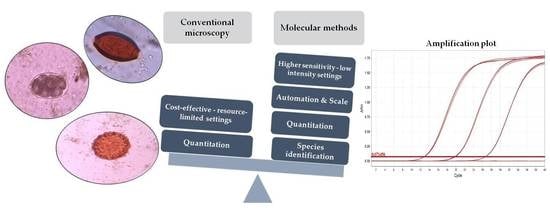
1. Introduction
Soil-transmitted helminths (STH) affect an estimated one billion of the world’s population (772–892 million with Ascaris lumbricoides, 430–508 million with Trichuris trichiura, and 406–480 million with hookworm) [1]. STH infections can lead to adverse health issues such as chronic blood loss leading to anemia, malabsorption of nutrients, loss of appetite, stunting, lethargy, school absenteeism, and lower performance on a range of cognitive tests [1,2,3,4]. In order to reduce STH-related morbidity, the WHO recommends annual or biannual targeted deworming of high-risk populations, including pre-school aged children (PSAC), school-aged children (SAC), and women of reproductive age (WRA) [5]. The frequency of deworming required in a region or country is based on the prevalence and intensity of infection, which is estimated by population-level parasitological surveys (usually carried out in school children) [6]. In these surveys, a known volume of a stool sample is tested for the presence of STH ova, and the worm burden (eggs per gram, EPG) for each species is estimated using microscopy-based methods. The EPG data are then used to classify infections into categories: heavy, moderate, and light intensity, for each species [7].
Several microscopy-based techniques for the detection of STH in both diagnostic laboratories and epidemiological surveys have been developed (Table 1). While direct wet mount preparation and fecal concentration methods (like formol–ether sedimentation and zinc sulfate floatation) remain the primary diagnostic tests for parasites in many laboratories, studies have shown these techniques to be less sensitive than the Kato–Katz method [8,9]. The Kato–Katz method, which also provides the EPG, is currently the most commonly used technique in population-based prevalence surveys but shows better sensitivity for A. lumbricoides and T. trichiura detection than hookworm [10]. The poorer sensitivity of this method for the detection of hookworm is due to the rapid disintegration of the ova [11]. The McMaster technique, originally used in veterinary samples, was shown to have an equivalent sensitivity to Kato–Katz for the detection of hookworm and T. trichiura with a good correlation of egg counts between these two techniques in a multicountry comparison [12]. This technique is relatively easy and less time-consuming. The major drawback with both the Kato–Katz and McMaster methods compared to other microscopy-based methods is their high limits of detection (LODs) of 24 EPG and 50 EPG, respectively. The FLOTAC method, which can be used to enumerate egg counts from a larger volume of fecal material (1 g or more), has a lower LOD of 1–2 EPG. This technique, however, requires a centrifuge, which can be a constraint in resource-limited settings and field surveys [13]. Although this technique has a lower LOD than Kato–Katz, it was shown to underestimate EPG counts [14,15]. The mini-FLOTAC is a simplified form of FLOTAC that does not require costly equipment. A meta-analysis indicated a sensitivity comparable to Kato–Katz for the detection of STH [16]. The FECPAKG2, a recently developed image-based egg counting method, was also less sensitive than single and two-slide Kato–Katz [17]. Lab-on-a-disk is another newly developed egg quantification platform that involves floatation followed by centrifugation and imaging, and showed a good correlation with the mini-FLOTAC when evaluated using samples from pigs and humans in Ethiopia [18]. Point-of-care mobile diagnostics with digital microscopy and deep learning-based image analysis algorithms have also been evaluated in proof-of-concept studies but need to be evaluated more widely in field settings [19,20].
Studies have shown that the rate of egg excretion varies from day to day and hence even the best techniques that rely on microscopy can result in false-negative results [25,26]. Population-based surveys from areas with a predominantly light intensity of infection have shown that standard microscopy-based techniques are suboptimal [27,28,29]. Additionally, the cost of Kato–Katz, while being perceived as cheap, may be higher and more variable when more samples or survey sites are added [30]. Hence, there is a need to develop more sensitive, less labor-intensive, and high-throughput detection methods. Furthermore, these methods should be cost-effective and able to be used in large-scale national surveys to monitor ongoing programs and measure the impact of interventions in endemic settings [31,32,33]. The cost of these newly developed techniques may remain higher than Kato–Katz or other microscopy-based methods, however, the benefits of increased accuracy and sensitivity may outweigh the additional cost [30].
In this review, we discuss the advantages and limitations of the various molecular tools used for STH detection in comparison to the conventional microscopy-based methods used in endemic settings. We have also described in detail the sample collection, storage, and pre-processing steps required for accurate nucleic acid detection and quantitation.
2. Role of Molecular Methods
Molecular methods have significantly improved the sensitivity of detection and quantitation of STH. They are also highly specific as molecular methods can help distinguish between closely related, morphologically similar, human and non-human (zoonotic) species compared to microscopic methods which could misidentify pathogens [27,29]. Some of the different molecular assays used for STH diagnosis are PCR, including real-time PCR and digital PCR, isothermal assays (LAMP), and, more recently, cell-free DNA detection (Table 2). The DNA sequences that have been targeted for the detection of STH include ribosomal DNA [34], repetitive sequences [35], and mitochondrial genes [36]. The rDNA genes and internal transcribed spacers (ITSs) are clusters of tandem repeats. They are targets of choice to detect morphologically similar parasites as they have sufficient heterogeneity and are found to exist as moderate copy number repeats [37]. The mitochondrial genes, especially cytochrome oxidase-I, have also been widely used for the detection of parasites and are more conserved targets than nuclear DNA [38]. More recently, Pilotte et al. developed a PCR that targets non-coding, repetitive DNA sequences using a next-generation sequencing approach that identifies genome-wide eukaryotic repetitive elements [35]. Targeting repetitive DNA sequences that make up several thousand to a million copies per diploid genome can greatly improve sensitivity with the lowest limit of detection being at or above 2 fg/uL, which is less than the quantity of DNA present in a single STH egg [35,39].
3. Sample Storage and DNA Extraction Methods
Sample storage and DNA extraction are critical steps in any molecular assay. Papaiakovou et al. compared standard sample storage at −20 ℃ used in most laboratories with different sample preservation techniques, including a silica bead two-step desiccation process, FTA cards, 5% potassium dichromate, RNAlater, ethanol, Paxgene, and Formalternate, and stored them at two different temperatures (4 ℃ and 32 ℃) for 60 days. This study was carried out using stool samples spiked with N. americanus ova. They found that while all preservation techniques were stable at 4 ℃, only storage using the silica bead two-step desiccation process, 5% potassium dichromate, and FTA card-based preservation had good stability at 32 ℃ with no significant change in the concentration of DNA detected [56]. In another study, storage with 96% ethanol at 4 ℃ was found to detect more concentrations of DNA than potassium dichromate and RNAlater [57]. Replicating these types of studies in real-world endemic settings will help determine alternative approaches for storage and transport of samples. This would be especially useful for surveys in remote or resource-limited settings where setting up a cold chain may not be feasible due to high costs and lack of infrastructure.
Over the years, there have been several advancements in DNA extraction methods for STH ova. Manual extraction followed by commercially available kit-based extractions that vary based on the sample type, to automated extraction platforms have been described. In the case of STH, it is essential to employ steps that homogenize fecal samples and break up the outer layer of the ova to obtain the maximum DNA yield. The commonly used egg disruption steps for fecal homogenates involve heating at high temperatures, multiple freeze–thaw cycles, and bead beating using non-degradable beads like silica, zirconium, etc. Several different lysis protocols have been developed, including: (1) washing with PBS then re-suspending in 2% polyvinylpolypyrrolidone (PVPP) and heating at 100 ℃ for 10 min [34], (2) mixing with lysis buffer, proteinase K, and β-mercaptoethanol, then freezing at 80 ℃ for 30 min followed by heating at 60 ℃ for 2 h [58], (3) multiple freeze–thaw cycles from liquid nitrogen to a water bath at 95 ℃ [42], (4) bead beating with ceramic beads [45], and (5) adding 2% PVPP followed by freezing overnight at −20 ℃ and then bead beating [59]. Ayana et al. compared two commercially available kits with and without a prior bead-beading step and found that bead beating significantly increased DNA yield [57]. Most laboratories now prefer commercially available kit-based DNA extraction methods but these also require pre-processing steps such as bead beating and/or lysis. The different DNA extraction kits evaluated for STH include Fecal Isolate II DNA Extraction Kit (Bioline, London, UK) [60], Powersoil DNA Isolation Kit (MO Bio, Carlsbad, CA, USA) [47], FastDNA Spin Kit for Soil (MP Biomedicals, Irvine, CA, USA) [61], QIAamp DNA Mini Kit (Qiagen Inc., Valencia, CA, USA) [59], DNeasy Tissue Kit (Qiagen, Hilden, Germany) [62], and the DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany) [63]. Automated platforms have also been used for STH, including Automate ExpressTM (Applied BiosystemsTM, Waltham, MA, USA) [64] and the Easy-plex system (AusDiagnostics, Mascot, Australia) [65], but remain challenging to establish in low-resource settings due to the infrastructure required.
4. Conventional PCR
The polymerase chain reaction, PCR, has significantly improved the detection of STH, showing greater sensitivity and detecting DNA at very low concentrations. A study that compared PCR with the formalin–ethyl acetate concentration technique for the detection of A. lumbricoides, N. americanus, and S. stercoralis in stool samples found that PCR was able to detect DNA concentrations as low as 0.001 ng [66]. Conventional PCR has also been used with other downstream techniques such as restriction fragment length polymorphism (RFLP) or Sanger sequencing to identify different species of STH [42]. Alternatively, nested or semi-nested PCRs to detect different species of the same genus have been carried out, for example, a study in a rural community in Thailand used a semi-nested approach to detect two different species of Trichuris—namely T. trichiura and T. vulpis [67]. PCR has also been multiplexed to detect multiple organisms within the same reaction, thereby reducing cost. This was evident in another study in Thailand where they used a triplex PCR approach to detect A. lumbricoides, N. americanus, and T. trichiura in stool samples. Their findings showed a combined STH sensitivity and specificity of 87% and 83%, respectively, with the ability to detect a single copy of DNA [68]. The major drawback with conventional PCR is the need for post-PCR analysis, which increases cost, turn-around time, and has a potential for contamination. With the availability of qPCR, conventional PCR is not commonly used for the detection of STH.
5. Real-Time PCR
Real-time PCR or quantitative PCR (qPCR) has been widely used for the detection of STH by several research groups (Table 2). qPCR assays can detect DNA concentrations that are lower than the amount of DNA found in a single fertilized STH ovum [69] and outperform Kato–Katz sensitivity and specificity even in endemic settings [28]. Two different chemistries, namely, DNA intercalating dyes like SYBR green [70] or probe-based assays like TaqMan, have been developed for STH detection, with the latter allowing for multiplexing.
Some of the real-time PCR-based techniques that have been used for the detection of STH include PCR-Luminex, high-resolution melt curve analysis, and multiplexed-tandem PCR. A PCR-Luminex assay (binding of PCR products to carboxylated Luminex beads with biotinylated primers and amine-modified probes) is a technique that has shown to be a high-throughput alternative to microscopy, with sensitivity and specificity comparable to qPCR [48]. Real-time PCR coupled with high-resolution melt curve analysis is another technique that has been used for the detection of different species of hookworm [49]. Multiplexed-tandem PCR is a newly developed technique that involves a multiplex, conventional PCR followed by a tandem real-time PCR. After this, the amplicons are subjected to a high-resolution melt curve analysis [51]. Detection of multiple helminths species by real-time PCR is, however, more often carried out with either a multiplex or a multiparallel approach.
Multiplex PCR has been shown to be more sensitive than microscopic techniques, including Kato–Katz, for nearly all STH with an LOD of 10 copies [46]. The multiparallel qPCR approach, where different species are tested in different reaction tubes, has a higher sensitivity than multiplex PCR, allowing detection of low-intensity infections (which may remain undetected in multiplex reactions). Another advantage of the multiparallel format is that it provides greater flexibility in selecting species-specific PCRs tailored to the endemicity in a particular region [45]. qPCR has also showed a better sensitivity when used to assess the efficacy of anthelminthic drugs, as it was able to detect infections that were undetected by Kato–Katz due to lower worm burden [71].
qPCR, as the name suggests, has also been employed for the quantification of STH infection. Easton et al. compared the EPG determined by Kato–Katz in rural Kenya with DNA concentration derived from qPCR for A. lumbricoides (ITS primers) and N. americanus (repetitive sequence primers) and found a strong correlation between EPG and average DNA concentration [28]. Mejia et al. also compared Kato–Katz and qPCR (ITS primers) and found a good correlation between egg count and DNA concentration for A. lumbricoides and T. trichiura [45]. A similar strong correlation was found between Kato–Katz EPG and DNA copy number (repetitive sequence primers) for A. lumbricoides and T. trichiura [72], whereas a study by Barda et al. in Tanzania showed poor correlation between EPG and DNA copy number for hookworm and moderate correlation for T. trichiura and A. lumbricoides [73]. Other groups have correlated EPG with raw qPCR Ct values rather than DNA concentration or copy number. A multiplex qPCR targeting N. americanus, A. duodenale, and O. bifurcum carried out in three sites in Africa showed that Ct values for N. americanus (ITS primers) had a good linear correlation with Kato–Katz EPG [34] while a study from Bangladesh showed a moderate correlation between EPG (double-slide Kato–Katz) and Ct values (repetitive sequence primers) [29]. A study by Llewellyn et al. used a multiplex qPCR assay (ITS primers) for the detection of Ascaris spp., Ancylostoma spp., N. americanus, and T. trichiura and converted Ct values to EPG and found a strong linear relationship [47].
Most studies have shown a correlation between DNA concentration or Ct values with EPG. However, when qPCR was used to categorize the intensity of infection into light, moderate, or heavy infections to provide data that are both programmatically and clinically relevant, mixed results for each species were obtained. A recent study found that although the median Ct values of samples which were microscopy negative or of light intensity (as classified by Kato–Katz for A. lumbricoides) were higher than for samples classified as MHI, there was overlap in the range of Ct detected [29]. A study by Knopp et al. found no difference in Ct values for hookworm when Kato–Katz was positive or negative [27]. A recent study was conducted by Levecke et al. using samples from multiple sites to compare categorization of infection intensity between multiple microscopy-based methods and qPCR. They found that that although ~80% of samples were correctly categorized as MHI by qPCR, an additional 10–25% of light-intensity infections (depending on species) were categorized as MHI by qPCR.
Data across these studies are, however, difficult to compare due to the different units of measurement used to represent intensity along with the different methods used [74]. In 2018, an international external quality assurance scheme for STH was initiated as a step towards more uniform reporting. The scheme was for nucleic acid amplification tests and has helped validate the diagnostic performance of these laboratories as well as allowed for comparability [75]. Another suggestion for more uniform reporting has been to express qPCR results more uniformly, in genomic equivalents per mL (GE/mL), to overcome issues such as variable primers and target copy numbers [76].
While the Ct value can be used to determine the infection intensity, multiple factors need to be taken into account while using qPCR as a quantitation method for STH ova. As Papaiakovou et al. mention, it is important to consider that in STH ova, haploid, diploid, or developing larval stages can be present. The copy number of the target sequences also needs to be considered, especially while using multicopy regions, which can improve sensitivity of detection but can lead to over- or under-quantification [77].
Various studies have developed qPCR with very low reaction volumes and improved reagents, significantly reducing the cost of detection. Mejia et al. estimated that a small reaction volume qPCR for the detection of eight parasites in a stool sample costs less than USD 1 [45]. Pooled testing of samples has also been evaluated as a cost-effective method [78,79] but the ability to detect a positive infection decreases when the intensity is low [80].
6. Isothermal Assay (LAMP)
The loop-mediated isothermal assay developed by Notomi et al. [81] works on the principle of auto-strand displacement DNA synthesis. Due to the isothermal conditions, the assay only requires a water bath or thermal block, thereby reducing the cost, and the reaction can be colorimetrically visualized. Two studies that compared LAMP with Kato–Katz for the detection of N. americanus and A. lumbricoides found the sensitivity of LAMP to be over 95% but with variable specificity [54,55]. The Smart Amplification2 (SmartAmp2) assay works in isothermal conditions and uses asymmetrical primers that allow more specific detection of various organisms. This has been applied to STH by Rashwan et al, who found that it allows detection of as little as 1 pg genomic DNA within 50–60 min [53]. The same group also developed a SmartAmp2 assay for the detection of benzimidazole resistance in N. americanus that target codons 167, 198, and 200 in the β-tubulin isotype 1 gene [82].
7. Other Recent Technical Developments
Digital PCR is a recently developed technique used for the detection of various parasitic infections [83]. It allows partitioning of a DNA sample into several individual reactions, with some reactions containing the target molecule, and others being negative. A single molecule is amplified a million-fold or more with TaqMan chemistry. The fraction of negative reactions is used for the absolute quantification of the total target molecule in a sample. It allows for quantitation without the need for a standard curve or endogenous controls. When the LODs of qPCR and digital PCR for the detection of A. lumbricoides egg in reclaimed water were compared, both techniques were able to detect one A. lumbricoides egg in 500 mL of reclaimed water [52]. Droplet digital PCR (ddPCR) is a form of digital PCR that is based on oil–water emulsion technology for the partitioning of the reaction mixture and is currently being evaluated for detection of STH in environmental samples in endemic areas as a potential surveillance tool.
In cell-free DNA assays, DNA released into the bloodstream or appearing in the urine as cells pass through the glomerular barrier or found in other body fluids like sputum, saliva, or stool is detected. Detection of the pathogen in urine and sputum can be less invasive, simple, and cost-effective when conducted at a large scale and has been used for the detection of Leishmania, Plasmodium, Trypanosoma, Schistosoma, and microfilaria. This method can also be used for the detection of STH due to the presence of cell-free DNA from the disintegration or decay of the parasites during different stages of their life cycle [84]. A study conducted in Iran used cell-free DNA from serum to detect Strongyloides infection in immunosuppressed patients [85]. Another study in Argentina detected Strongyloides in urine samples and it was found to have a significant advantage over fecal examination [86]. Thus, this technique can potentially be explored for STH detection.
A new field-adapted method has been developed recently using miniPCR®. Here, genomic DNA is amplified with a portable thermocycler, after which the amplicon is loaded on a quantitative paper-based DNA reader which quantifies DNA based on the distance traveled on a cellulose strip which is then visualized for the naked eye [43]. This fast and relatively simple method can be used in field settings as a point-of-care (POC) test but requires validation and development of an equally portable stool DNA extraction method.
8. Conclusions
Molecular techniques are highly sensitive and specific for the detection of STH and can also detect benzimidazole resistance markers. A current limitation of molecular tools is the difficulty in determining the intensity of infection to provide relevant programmatic data. Limitations also include initial cost, and requirement for specialized equipment (in most methods), infrastructure, and trained personnel, thus making them difficult to be implemented in resource-limited settings. However, when combined with high-throughput automation at scale, molecular methods could potentially replace the more human resource-intensive microscopy-based assays in national-level survey programs. In this scenario, small-volume PCR reactions and pooled testing would also contribute to reduced costs. Issues like stability of samples and requirements for an expensive cold chain can be overcome with preservation steps. Another alternative would be a POC molecular testing strategy, but for STH, assays are not yet at the field-deployable stage due to the extensive pre-processing required. While molecular techniques require building up of expertise and thorough standardization, they have the added advantage of reducing variability due to varying levels of microscopic skills and duration of sample transport, and could also be used for detection of STH in the environment. After repeated rounds of MDA as part of the STH and LF control programs, and with improving sanitation, several endemic communities have transitioned to low-intensity infections. Due to the potential scope for elimination, there is a pressing need for highly sensitive molecular techniques for monitoring and evaluation of control programs and interventions.
Author Contributions
Conceptualization, M.M. and S.S.R.A.; methodology, M.M. and K.R.; software, K.R.; data curation, M.M. and K.R.; writing—original draft preparation, M.M.; writing—review and editing, M.M. and S.S.R.A.; supervision, S.S.R.A.; funding acquisition, S.S.R.A. All authors have read and agreed to the published version of the manuscript.
Funding
S.S.R.A. receives support from an Emerging Global Leader Award (K43) from Fogarty International Center, NIH (1K43TW011415).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank A. D’Souza (University of Sydney) for editing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pullan, R.L.; Smith, J.L.; Jasrasaria, R.; Brooker, S.J. Global numbers of infection and disease burden of soil transmitted helminth infections in 2010. Parasit Vectors 2014, 7, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nokes, C.; Grantham-McGregor, S.M.; Sawyer, A.W.; Cooper, E.S.; Robinson, B.A.; Bundy, D.A. Moderate to heavy infections of Trichuris trichiura affect cognitive function in Jamaican school children. Parasitology 1992, 104, 539–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nokes, C.; Bundy, D.A. Compliance and absenteeism in school children: Implications for helminth control. Trans. R. Soc. Trop. Med. Hyg. 1993, 87, 148–152. [Google Scholar] [CrossRef]
- Savioli, L.; Bundy, D.; Tomkins, A. Intestinal parasitic infections: A soluble public health problem. Trans. R. Soc. Trop. Med. Hyg. 1992, 86, 353–354. [Google Scholar] [CrossRef] [Green Version]
- WHO. 2030 Targets for Soil-Transmitted Helminthiases Control Programmes; World Health Organization: Geneva, Switzerland, 2020; Available online: https://apps.who.int/iris/handle/10665/330611 (accessed on 8 July 2020).
- WHO. Assessing the Epidemiology of Soil-Transmitted Helminths during a Transmission Assessment Survey (TAS); World Health Organization: Geneva, Switzerland, 2015; Available online: http://www.who.int/intestinal_worms/resources/9789241508384/en/ (accessed on 11 July 2020).
- Montresor, A.; WHO. Helminth Control in School Age Children; World Health Organization: Geneva, Switzerland, 2002; Available online: http://www.who.int/intestinal_worms/resources/9789241548267/en/ (accessed on 8 July 2020).
- Mengist, H.M.; Demeke, G.; Zewdie, O.; Belew, A. Diagnostic performance of direct wet mount microscopy in detecting intestinal helminths among pregnant women attending ante-natal care (ANC) in East Wollega, Oromia, Ethiopia. BMC Res. Notes 2018, 11, 276. [Google Scholar] [CrossRef] [Green Version]
- Endris, M.; Tekeste, Z.; Lemma, W.; Kassu, A. Comparison of the Kato-Katz, Wet Mount, and Formol-Ether Concentration Diagnostic Techniques for Intestinal Helminth Infections in Ethiopia. ISRN Parasitol. 2013, 2013, 180439. [Google Scholar] [CrossRef] [Green Version]
- Cools, P.; Vlaminck, J.; Albonico, M.; Ame, S.; Ayana, M.; José Antonio, B.P.; Cringoli, G.; Dana, D.; Keiser, J.; Maurelli, M.P.; et al. Diagnostic performance of a single and duplicate Kato-Katz, Mini-FLOTAC, FECPAKG2 and qPCR for the detection and quantification of soil-transmitted helminths in three endemic countries. PLoS Negl. Trop. Dis. 2019, 13, e0007446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarafder, M.R.; Carabin, H.; Joseph, L.; Balolong, E.; Olveda, R.; McGarvey, S.T. Estimating the sensitivity and specificity of Kato-Katz stool examination technique for detection of hookworms, Ascaris lumbricoides and Trichuris trichiura infections in humans in the absence of a “gold standard”. Int. J. Parasitol. 2010, 40, 399–404. [Google Scholar] [CrossRef] [Green Version]
- Levecke, B.; Behnke, J.M.; Ajjampur, S.S.; Albonico, M.; Ame, S.M.; Charlier, J.; Geiger, S.M.; Hoa, N.T.; Kamwa Ngassam, R.I.; Kotze, A.C.; et al. A comparison of the sensitivity and fecal egg counts of the McMaster egg counting and Kato-Katz thick smear methods for soil-transmitted helminths. PLoS Negl. Trop. Dis. 2011, 5, e1201. [Google Scholar] [CrossRef] [Green Version]
- Cringoli, G.; Rinaldi, L.; Maurelli, M.P.; Utzinger, J. FLOTAC: New multivalent techniques for qualitative and quantitative copromicroscopic diagnosis of parasites in animals and humans. Nat. Protoc. 2010, 5, 503–515. [Google Scholar] [CrossRef]
- Habtamu, K.; Degarege, A.; Ye-Ebiyo, Y.; Erko, B. Comparison of the Kato-Katz and FLOTAC techniques for the diagnosis of soil-transmitted helminth infections. Parasitol. Int. 2011, 60, 398–402. [Google Scholar] [CrossRef]
- Knopp, S.; Speich, B.; Hattendorf, J.; Rinaldi, L.; Mohammed, K.A.; Khamis, I.S.; Mohammed, A.S.; Albonico, M.; Rollinson, D.; Marti, H.; et al. Diagnostic Accuracy of Kato-Katz and FLOTAC for Assessing Anthelmintic Drug Efficacy. PLoS Negl. Trop. Dis. 2011, 5, e1036. [Google Scholar] [CrossRef] [PubMed]
- Nikolay, B.; Brooker, S.J.; Pullan, R.L. Sensitivity of diagnostic tests for human soil-transmitted helminth infections: A meta-analysis in the absence of a true gold standard. Int. J. Parasitol. 2014, 44, 765–774. [Google Scholar] [CrossRef] [Green Version]
- Moser, W.; Bärenbold, O.; Mirams, G.J.; Cools, P.; Vlaminck, J.; Ali, S.M.; Ame, S.M.; Hattendorf, J.; Vounatsou, P.; Levecke, B.; et al. Diagnostic comparison between FECPAKG2 and the Kato-Katz method for analyzing soil-transmitted helminth eggs in stool. PLoS Negl. Trop. Dis. 2018, 12, e0006562. [Google Scholar] [CrossRef] [PubMed]
- Sukas, S.; Van Dorst, B.; Kryj, A.; Lagatie, O.; De Malsche, W.; Stuyver, L.J. Development of a Lab-on-a-Disk Platform with Digital Imaging for Identification and Counting of Parasite Eggs in Human and Animal Stool. Micromachines 2019, 10, 852. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.; Bakhtari, N.; Langdon-Embry, L.; Redwood, E.; Grandjean Lapierre, S.; Rakotomanga, P.; Rafalimanantsoa, A.; De Dios Santos, J.; Vigan-Womas, I.; Knoblauch, A.M.; et al. Kankanet: An artificial neural network-based object detection smartphone application and mobile microscope as a point-of-care diagnostic aid for soil-transmitted helminthiases. PLoS Negl. Trop. Dis. 2019, 13, e0007577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmström, O.; Linder, N.; Ngasala, B.; Mårtensson, A.; Linder, E.; Lundin, M.; Moilanen, H.; Suutala, A.; Diwan, V.; Lundin, J. Point-of-care mobile digital microscopy and deep learning for the detection of soil-transmitted helminths and Schistosoma haematobium. Glob. Health Action 2017, 10 (Suppl. S3), 1337325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Periago, M.V.; Diniz, R.C.; Pinto, S.A.; Yakovleva, A.; Correa-Oliveira, R.; Diemert, D.J.; Bethony, J.M. The Right Tool for the Job: Detection of Soil-Transmitted Helminths in Areas Co-endemic for Other Helminths. PLoS Negl. Trop. Dis. 2015, 9, e0003967. [Google Scholar] [CrossRef]
- Albonico, M.; Rinaldi, L.; Sciascia, S.; Morgoglione, M.E.; Piemonte, M.; Maurelli, M.P.; Musella, V.; Utzinger, J.; Ali, S.M.; Ame, S.M.; et al. Comparison of three copromicroscopic methods to assess albendazole efficacy against soil-transmitted helminth infections in school-aged children on Pemba Island. Trans. R. Soc. Trop. Med. Hyg. 2013, 107, 493–501. [Google Scholar] [CrossRef] [Green Version]
- Barda, B.; Cajal, P.; Villagran, E.; Cimino, R.; Juarez, M.; Krolewiecki, A.; Rinaldi, L.; Cringoli, G.; Burioni, R.; Albonico, M. Mini-FLOTAC, Kato-Katz and McMaster: Three methods, one goal; highlights from north Argentina. Parasit. Vectors 2014, 7, 271. [Google Scholar] [CrossRef] [Green Version]
- Albonico, M.; Ame, S.M.; Vercruysse, J.; Levecke, B. Comparison of the Kato-Katz thick smear and McMaster egg counting techniques for monitoring drug efficacy against soil-transmitted helminths in schoolchildren on Pemba Island, Tanzania. Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 199–201. [Google Scholar] [CrossRef]
- Bärenbold, O.; Raso, G.; Coulibaly, J.T.; N’Goran, E.K.; Utzinger, J.; Vounatsou, P. Estimating sensitivity of the Kato-Katz technique for the diagnosis of Schistosoma mansoni and hookworm in relation to infection intensity. PLoS Negl. Trop. Dis. 2017, 11, e0005953. [Google Scholar] [CrossRef]
- Liu, C.; Lu, L.; Zhang, L.; Bai, Y.; Medina, A.; Rozelle, S.; Smith, D.S.; Zhou, C.; Zang, W. More Poop, More Precision: Improving Epidemiologic Surveillance of Soil-Transmitted Helminths with Multiple Fecal Sampling using the Kato–Katz Technique. Am. J. Trop. Med. Hyg. 2017, 97, 870–875. [Google Scholar] [CrossRef]
- Knopp, S.; Salim, N.; Schindler, T.; Voules, D.A.K.; Rothen, J.; Lweno, O.; Mohammed, A.S.; Singo, R.; Benninghoff, M.; Nsojo, A.A.; et al. Diagnostic Accuracy of Kato–Katz, FLOTAC, Baermann, and PCR Methods for the Detection of Light-Intensity Hookworm and Strongyloides stercoralis Infections in Tanzania. Am. J. Trop. Med. Hyg. 2014, 90, 535–545. [Google Scholar] [CrossRef] [Green Version]
- Easton, A.V.; Oliveira, R.G.; O’Connell, E.M.; Kepha, S.; Mwandawiro, C.S.; Njenga, S.M.; Kihara, J.H.; Mwatele, C.; Odiere, M.R.; Brooker, S.J.; et al. Multi-parallel qPCR provides increased sensitivity and diagnostic breadth for gastrointestinal parasites of humans: Field-based inferences on the impact of mass deworming. Parasit. Vectors 2016, 9, 38. [Google Scholar] [CrossRef] [Green Version]
- Benjamin-Chung, J.; Pilotte, N.; Ercumen, A.; Grant, J.R.; Maasch, J.R.; Gonzalez, A.M.; Ester, A.C.; Arnold, B.F.; Rahman, M.; Haque, R.; et al. Comparison of multi-parallel qPCR and double-slide Kato-Katz for detection of soil-transmitted helminth infection among children in rural Bangladesh. PLoS Negl. Trop. Dis. 2020, 14, e0008087. [Google Scholar] [CrossRef]
- Turner, H.C.; Bettis, A.A.; Dunn, J.C.; Whitton, J.M.; Hollingsworth, T.D.; Fleming, F.M.; Anderson, R.M. Economic Considerations for Moving beyond the Kato-Katz Technique for Diagnosing Intestinal Parasites as We Move Towards Elimination. Trends Parasitol. 2017, 33, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ásbjörnsdóttir, K.H.; Ajjampur, S.S.R.; Anderson, R.M.; Bailey, R.; Gardiner, I.; Halliday, K.E.; Ibikounle, M.; Kalua, K.; Kang, G.; Littlewood, D.T.J.; et al. Assessing the feasibility of interrupting the transmission of soil-transmitted helminths through mass drug administration: The DeWorm3 cluster randomized trial protocol. PLoS Negl. Trop. Dis. 2018, 12, e0006166. [Google Scholar] [CrossRef] [PubMed]
- Brooker, S.J.; Mwandawiro, C.S.; Halliday, K.E.; Njenga, S.M.; Mcharo, C.; Gichuki, P.M.; Wasunna, B.; Kihara, J.H.; Njomo, D.; Alusala, D.; et al. Interrupting transmission of soil-transmitted helminths: A study protocol for cluster randomised trials evaluating alternative treatment strategies and delivery systems in Kenya. BMJ Open 2015, 5, e008950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The DeWorm3 Project Team; Hasegawa, M.; Pilotte, N.; Kikuchi, M.; Means, A.R.; Papaiakovou, M.; Gonzalez, A.M.; Maasch, J.R.; Ikuno, H.; Sunahara, T.; et al. What does soil-transmitted helminth elimination look like? Results from a targeted molecular detection survey in Japan. Parasit. Vectors 2020, 13, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verweij, J.J.; Brienen, E.A.T.; Ziem, J.; Polderman, A.M.; Yelifari, L.; Van Lieshout, L. Simultaneous Detection and Quantification of Ancylostoma duodenale, Necator americanus, and Oesophagostomum bifurcum in Fecal Samples Using Multiplex Real-Time PCR. Am. J. Trop. Med. Hyg. 2007, 77, 685–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilotte, N.; Papaiakovou, M.; Grant, J.R.; Bierwert, L.A.; Llewellyn, S.; McCarthy, J.S.; Williams, S.A. Improved PCR-Based Detection of Soil Transmitted Helminth Infections Using a Next-Generation Sequencing Approach to Assay Design. PLoS Negl. Trop. Dis. 2016, 10, e0004578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, B.; Li, T.; Xiao, S.; Zheng, F.; Hawdon, J.M. Species-Specific Identification of Human Hookworms by PCR of the Mitochondrial Cytochrome Oxidase I Gene. J. Parasitol. 2001, 87, 1227. [Google Scholar] [CrossRef]
- Fairley, T.L.; Kilpatrick, C.W.; Conn, J.E. Intragenomic heterogeneity of internal transcribed spacer rDNA in neotropical malaria vector Anopheles aquasalis (Diptera: Culicidae). J. Med. Entomol. 2005, 42, 795–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.; Singh, M.N.; Bera, A.K.; Bandyopadhyay, S.; Bhattacharya, D. Molecular basis for identification of species/isolates of gastrointestinal nematode parasites. Asian Pac. J. Trop. Med. 2011, 4, 589–593. [Google Scholar] [CrossRef] [Green Version]
- Elder, J.F.; Turner, B.J. Concerted Evolution of Repetitive DNA Sequences in Eukaryotes. Q. Rev. Biol. 1995, 70, 297–320. [Google Scholar] [CrossRef] [PubMed]
- De Gruijter, J.M.; Van Lieshout, L.; Gasser, R.B.; Verweij, J.J.; Brienen, E.A.; Ziem, J.B.; Yelifari, L.; Polderman, A.M. Polymerase chain reaction-based differential diagnosis of Ancylostoma duodenale and Necator americanus infections in humans in northern Ghana. Trop. Med. Int. Health 2005, 10, 574–580. [Google Scholar] [CrossRef]
- Verweij, J.J.; Pit, D.S.S.; Van Lieshout, L.; Baeta, S.M.; Dery, G.D.; Gasser, R.B.; Polderman, A.M. Determining the prevalence of Oesophagostomum bifurcum and Necator americanus infections using specific PCR amplification of DNA from faecal samples. Trop. Med. Int. Health 2001, 6, 726–731. [Google Scholar] [CrossRef] [PubMed]
- George, S.; Geldhof, P.; Albonico, M.; Ame, S.M.; Bethony, J.M.; Engels, D.; Mekonnen, Z.; Montresor, A.; Sopheak, H.; Tchuem-Tchuenté, L.A. Molecular speciation of soil-transmitted helminths egg isolates collected during six drug efficacy trials in endemic countries. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 657–663. [Google Scholar] [CrossRef]
- Wang, A.G.; Dong, T.; Mansour, H.; Matamoros, G.; Sanchez, A.L.; Li, F. Paper-Based DNA Reader for Visualized Quantification of Soil-Transmitted Helminth Infections. ACS Sens. 2018, 3, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Schär, F.; Odermatt, P.; Khieu, V.; Panning, M.; Duong, S.; Muth, S.; Marti, H.; Kramme, S. Evaluation of real-time PCR for Strongyloides stercoralis and hookworm as diagnostic tool in asymptomatic schoolchildren in Cambodia. Acta Trop. 2013, 126, 89–92. [Google Scholar] [CrossRef] [Green Version]
- Mejia, R.; Vicuña, Y.; Broncano, N.; Sandoval, C.; Vaca, M.; Chico, M.; Cooper, P.J.; Nutman, T.B. A Novel, Multi-Parallel, Real-Time Polymerase Chain Reaction Approach for Eight Gastrointestinal Parasites Provides Improved Diagnostic Capabilities to Resource-Limited At-Risk Populations. Am. J. Trop. Med. Hyg. 2013, 88, 1041–1047. [Google Scholar] [CrossRef]
- Basuni, M.; Muhi, J.; Othman, N.; Verweij, J.J.; Ahmad, M.; Miswan, N.; Rahumatullah, A.; Aziz, F.A.; Zainudin, N.S.; Noordin, R. A Pentaplex Real-Time Polymerase Chain Reaction Assay for Detection of Four Species of Soil-Transmitted Helminths. Am. J. Trop. Med. Hyg. 2011, 84, 338–343. [Google Scholar] [CrossRef] [Green Version]
- Llewellyn, S.; Inpankaew, T.; Nery, S.V.; Gray, D.J.; Verweij, J.J.; Clements, A.C.; Gomes, S.J.; Traub, R.; McCarthy, J.S. Application of a Multiplex Quantitative PCR to Assess Prevalence and Intensity Of Intestinal Parasite Infections in a Controlled Clinical Trial. PLoS Negl. Trop. Dis. 2016, 10, e0004380. [Google Scholar] [CrossRef]
- Taniuchi, M.; Verweij, J.J.; Noor, Z.; Sobuz, S.U.; Van Lieshout, L.; Petri, W.A., Jr.; Haque, R.; Houpt, E.R. High Throughput Multiplex PCR and Probe-based Detection with Luminex Beads for Seven Intestinal Parasites. Am. J. Trop. Med. Hyg. 2011, 84, 332–337. [Google Scholar] [CrossRef] [Green Version]
- Ngui, R.; Lim, Y.A.L.; Chua, K.H. Rapid Detection and Identification of Human Hookworm Infections through High Resolution Melting (HRM) Analysis. PLoS ONE 2012, 7, e41996. [Google Scholar] [CrossRef]
- Cunningham, L.J.; Stothard, J.R.; Osei-Atweneboana, M.; Armoo, S.; Verweij, J.J.; Adams, E.R. Developing a real-time PCR assay based on multiplex high-resolution melt-curve analysis: A pilot study in detection and discrimination of soil-transmitted helminth and schistosome species. Parasitology 2018, 145, 1733–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stracke, K.; Clarke, N.; Awburn, C.V.; Vaz Nery, S.; Khieu, V.; Traub, R.J.; Jex, A.R. Development and validation of a multiplexed-tandem qPCR tool for diagnostics of human soil-transmitted helminth infections. PLoS Negl. Trop. Dis. 2019, 13, e0007363. [Google Scholar] [CrossRef] [Green Version]
- Acosta Soto, L.; Santísima-Trinidad, A.B.; Bornay-Llinares, F.J.; Martín González, M.; Pascual Valero, J.A.; Ros Muñoz, M. Quantitative PCR and Digital PCR for Detection of Ascaris lumbricoides Eggs in Reclaimed Water. Biomed. Res. Int. 2017, 2017, 7515409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashwan, N.; Diawara, A.; Scott, M.E.; Prichard, R.K. Isothermal diagnostic assays for the detection of soil-transmitted helminths based on the SmartAmp2 method. Parasit. Vectors 2017, 10, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mugambi, R.M.; Agola, E.L.; Mwangi, I.N.; Kinyua, J.; Shiraho, E.A.; Mkoji, G.M. Development and evaluation of a Loop Mediated Isothermal Amplification (LAMP) technique for the detection of hookworm (Necator americanus) infection in fecal samples. Parasit. Vectors 2015, 8, 574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiraho, E.A.; Eric, A.L.; Mwangi, I.N.; Maina, G.M.; Kinuthia, J.M.; Mutuku, M.W.; Mugambi, R.M.; Mwandi, J.M.; Mkoji, G.M. Development of a Loop Mediated Isothermal Amplification for Diagnosis of Ascaris lumbricoides in Fecal Samples. J. Parasitol. Res. 2016, 2016, 7376207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papaiakovou, M.; Pilotte, N.; Baumer, B.; Grant, J.; Asbjornsdottir, K.; Schaer, F.; Hu, Y.; Aroian, R.; Walson, J.; Williams, S.A. A comparative analysis of preservation techniques for the optimal molecular detection of hookworm DNA in a human fecal specimen. PLoS Negl. Trop. Dis. 2018, 12, e0006130. [Google Scholar] [CrossRef]
- Ayana, M.; Cools, P.; Mekonnen, Z.; Biruksew, A.; Dana, D.; Rashwan, N.; Prichard, R.; Vlaminck, J.; Verweij, J.J.; Levecke, B. Comparison of four DNA extraction and three preservation protocols for the molecular detection and quantification of soil-transmitted helminths in stool. PLoS Negl. Trop. Dis. 2019, 13, e0007778. [Google Scholar] [CrossRef] [Green Version]
- Diawara, A.; Halpenny, C.M.; Churcher, T.S.; Mwandawiro, C.; Kihara, J.; Kaplan, R.M.; Streit, T.G.; Idaghdour, Y.; Scott, M.E.; Basáñez, M.G.; et al. Association between Response to Albendazole Treatment and β-Tubulin Genotype Frequencies in Soil-transmitted Helminths. PLoS Negl. Trop. Dis. 2013, 7, e2247. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, L.J.; Odoom, J.; Pratt, D.; Boatemaa, L.; Asante-Ntim, N.; Attiku, K.; Banahene, B.; Osei-Atweneboana, M.; Verweij, J.J.; Molyneux, D.; et al. Expanding molecular diagnostics of helminthiasis: Piloting use of the GPLN platform for surveillance of soil transmitted helminthiasis and schistosomiasis in Ghana. PLoS Negl. Trop. Dis. 2018, 12, e0006129. [Google Scholar] [CrossRef] [PubMed]
- Hii, S.F.; Senevirathna, D.; Llewellyn, S.; Inpankaew, T.; Odermatt, P.; Khieu, V.; Muth, S.; McCarthy, J.; Traub, R.J. Development and Evaluation of a Multiplex Quantitative Real-Time Polymerase Chain Reaction for Hookworm Species in Human Stool. Am. J. Trop. Med. Hyg. 2018, 99, 1186–1193. [Google Scholar] [CrossRef] [Green Version]
- Velasquez, D.E.; Arvelo, W.; Cama, V.A.; López, B.; Reyes, L.; Roellig, D.M.; Kahn, G.D.; Lindblade, K.A. Molecular Insights for Giardia, Cryptosporidium, and Soil-Transmitted Helminths from a Facility-Based Surveillance System in Guatemala. Am. J. Trop. Med. Hyg. 2011, 85, 1141–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunet, J.; Lemoine, J.P.; Lefebvre, N.; Denis, J.; Pfaff, A.W.; Abou-Bacar, A.; Traub, R.J.; Pesson, B.; Candolfi, E. Bloody Diarrhea Associated with Hookworm Infection in Traveler Returning to France from Myanmar. Emerg. Infect. Dis. 2015, 21, 1878–1879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng-Nguyen, D.; Hii, S.F.; Nguyen, V.-A.T.; Van Nguyen, T.; Van Nguyen, D.; Traub, R.J. Re-evaluation of the species of hookworms infecting dogs in Central Vietnam. Parasit. Vectors 2015, 8, 401. [Google Scholar] [CrossRef] [Green Version]
- Matamoros, G.; Rueda, M.M.; Rodríguez, C.; Gabrie, J.A.; Canales, M.; Fontecha, G.; Sanchez, A. High Endemicity of Soil-Transmitted Helminths in a Population Frequently Exposed to Albendazole but No Evidence of Antiparasitic Resistance. Trop. Med. Infect. Dis. 2019, 4, 73. [Google Scholar] [CrossRef] [Green Version]
- Roeber, F.; Jex, A.R.; Campbell, A.J.; Nielsen, R.; Anderson, G.A.; Stanley, K.K.; Gasser, R.B. Establishment of a robotic, high-throughput platform for the specific diagnosis of gastrointestinal nematode infections in sheep. Int. J. Parasitol. 2012, 42, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Sanprasert, V.; Kerdkaew, R.; Srirungruang, S.; Charuchaibovorn, S.; Phadungsaksawasdi, K.; Nuchprayoon, S. Development of Conventional Multiplex PCR: A Rapid Technique for Simultaneous Detection of Soil-Transmitted Helminths. Pathogens 2019, 8, 152. [Google Scholar] [CrossRef] [Green Version]
- Areekul, P.; Putaporntip, C.; Pattanawong, U.; Sitthicharoenchai, P.; Jongwutiwes, S. Trichuris vulpis and T. trichiura infections among schoolchildren of a rural community in northwestern Thailand: The possible role of dogs in disease transmission. Asian Biomed. 2010, 4, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Phuphisut, O.; Yoonuan, T.; Sanguankiat, S.; Chaisiri, K.; Maipanich, W.; Pubampen, S.; Komalamisra, C.; Adisakwattana, P. Triplex polymerase chain reaction assay for detection of major soil-transmitted helminths, Ascaris lumbricoides, Trichuris trichiura, Necator americanus, in fecal samples. Southeast Asian J. Trop. Med. Public Health 2014, 45, 267–275. [Google Scholar]
- Pilotte, N.; Maasch, J.R.M.A.; Easton, A.V.; Dahlstrom, E.; Nutman, T.B.; Williams, S.A. Targeting a highly repeated germline DNA sequence for improved real-time PCR-based detection of Ascaris infection in human stool. PLoS Negl. Trop. Dis. 2019, 22, e0007593. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.-X.; Pan, C.-S.; Cui, L.-W. Application of a real-time PCR method for detecting and monitoring hookworm Necator americanus infections in Southern China. Asian Pac. J. Trop. Biomed. 2012, 2, 925–929. [Google Scholar] [CrossRef] [Green Version]
- Keller, L.; Patel, C.; Welsche, S.; Schindler, T.; Hürlimann, E.; Keiser, J. Performance of the Kato-Katz method and real time polymerase chain reaction for the diagnosis of soil-transmitted helminthiasis in the framework of a randomised controlled trial: Treatment efficacy and day-to-day variation. Parasit. Vectors 2020, 13, 1–12. [Google Scholar] [CrossRef]
- Dunn, J.C.; Papaiakovou, M.; Han, K.T.; Chooneea, D.; Bettis, A.A.; Wyine, N.Y.; Lwin, A.M.M.; Maung, N.S.; Misra, R.; Littlewood, D.T.J.; et al. The increased sensitivity of qPCR in comparison to Kato-Katz is required for the accurate assessment of the prevalence of soil-transmitted helminth infection in settings that have received multiple rounds of mass drug administration. Parasit Vectors. 2020, 13, 324. [Google Scholar] [CrossRef] [PubMed]
- Barda, B.; Schindler, C.; Wampfler, R.; Ame, S.; Ali, S.M.; Keiser, J. Comparison of real-time PCR and the Kato-Katz method for the diagnosis of soil-transmitted helminthiasis and assessment of cure in a randomized controlled trial. BMC Microbiol. 2020, 20, 298. [Google Scholar] [CrossRef] [PubMed]
- Levecke, B.; Cools, P.; Albonico, M.; Ame, S.; Angebault, C.; Ayana, M.; Behnke, J.M.; Bethony, J.M.; Cringoli, G.; Dana, D.; et al. Identifying thresholds for classifying moderate-to-heavy soil-transmitted helminth intensity infections for FECPAKG2, McMaster, Mini-FLOTAC and qPCR. PLoS Negl. Trop. Dis. 2020, 14, e0008296. [Google Scholar] [CrossRef]
- Cools, P.; Van Lieshout, L.; Koelewijn, R.; Addiss, D.; Ajjampur, S.S.; Ayana, M.; Bradbury, R.S.; Cantera, J.L.; Dana, D.; Fischer, K.; et al. First international external quality assessment scheme of nucleic acid amplification tests for the detection of Schistosoma and soil-transmitted helminths, including Strongyloides: A pilot study. PLoS Negl. Trop. Dis. 2020, 14, e0008231. [Google Scholar] [CrossRef]
- Cools, P.; Vlaminck, J.; Verweij, J.J.; Levecke, B. Quantitative PCR in soil-transmitted helminth epidemiology and control programs: Toward a universal standard. PLoS Negl. Trop. Dis. 2021, 15, e0009134. [Google Scholar] [CrossRef]
- Papaiakovou, M.; Gasser, R.B.; Littlewood, D.T.J. Quantitative PCR-Based Diagnosis of Soil-Transmitted Helminth Infections: Faecal or Fickle? Trends Parasitol. 2019, 35, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Papaiakovou, M.; Wright, J.; Pilotte, N.; Chooneea, D.; Schär, F.; Truscott, J.E.; Dunn, J.C.; Gardiner, I.; Walson, J.L.; Williams, S.A.; et al. Pooling as a strategy for the timely diagnosis of soil-transmitted helminths in stool: Value and reproducibility. Parasit. Vectors. 2019, 12, 443. [Google Scholar] [CrossRef] [Green Version]
- Truscott, J.E.; Dunn, J.C.; Papaiakovou, M.; Schaer, F.; Werkman, M.; Littlewood, D.T.J.; Walson, J.L.; Anderson, R.M. Calculating the prevalence of soil-transmitted helminth infection through pooling of stool samples: Choosing and optimizing the pooling strategy. PLoS Negl. Trop. Dis. 2019, 13, e0007196. [Google Scholar] [CrossRef]
- Leta, G.T.; French, M.; Dorny, P.; Vercruysse, J.; Levecke, B. Comparison of individual and pooled diagnostic examination strategies during the national mapping of soil-transmitted helminths and Schistosoma mansoni in Ethiopia. PLoS Negl. Trop. Dis. 2018, 12, e0006723. [Google Scholar] [CrossRef] [PubMed]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, E63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashwan, N.; Bourguinat, C.; Keller, K.; Gunawardena, N.K.; De Silva, N.; Prichard, R. Isothermal Diagnostic Assays for Monitoring Single Nucleotide Polymorphisms in Necator americanus Associated with Benzimidazole Drug Resistance. PLoS Negl. Trop. Dis. 2016, 10, e0005113. [Google Scholar] [CrossRef]
- Pomari, E.; Piubelli, C.; Perandin, F.; Bisoffi, Z. Digital PCR: A new technology for diagnosis of parasitic infections. Clin. Microbiol. Infect. 2019, 25, 1510–1516. [Google Scholar] [CrossRef] [PubMed]
- Weerakoon, K.G.; McManus, D.P. Cell-Free DNA as a Diagnostic Tool for Human Parasitic Infections. Trends Parasitol. 2016, 32, 378–391. [Google Scholar] [CrossRef] [PubMed]
- Gorgani-Firouzjaee, T.; Kalantari, N.; Javanian, M.; Ghaffari, S. Strongyloides stercoralis: Detection of parasite-derived DNA in serum samples obtained from immunosuppressed patients. Parasitol. Res. 2018, 117, 2927–2932. [Google Scholar] [CrossRef] [PubMed]
- Lodh, N.; Caro, R.; Sofer, S.; Scott, A.; Krolewiecki, A.; Shiff, C. Diagnosis of Strongyloides stercoralis: Detection of parasite-derived DNA in urine. Acta Trop. 2016, 163, 9–13. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
Table 1.
Microscopy-based detection methods for soil-transmitted helminths.
| Method | Principle | Sensitivity | Negative Predictive Value | Reference |
|---|---|---|---|---|
| Wet mount preparation | Preparation of stool sample with saline/iodine on a microscopic slide with a cover glass. | Hookworm: 85.7% A. lumbricoides: 83.3% | Hookworm: 97.5% A. lumbricoides: 98.8% | [8] |
| Hookworm: 37.9% A. lumbricoides: 52% T. trichiura:1 2.5% | [9] | |||
| Formol–ether sedimentation | Stool sample diluted in distilled water is centrifuged with 3% ethyl ether. Of the four layers formed, the lower sediment is mixed with 5% formalin and 50 uL of this sediment screened under a microscope. | Hookworm: 95.8% A. lumbricoides: 94.2% T. trichiura: 86.7% | Hookworm: 98.8% A. lumbricoides: 98.5% T. trichiura: 99.8% | [21] |
| Hookworm: 72.4% A. lumbricoides: 81.4% T. trichiura: 57.8% | [9] | |||
| Kato–Katz | A known amount of sieved stool sample is placed on a glass slide and covered with cellophane soaked in methylene blue solution and screened under a microscope after 30 min and the eggs are counted. | Hookworm: 69% A. lumbricoides: 93.1% T. trichiura: 90.6% | [9] | |
| Hookworm: 72.4% A. lumbricoides: 57.4% Trichuris spp.: 84.4% | [10] | |||
| Hookworm: 62.9% A. lumbricoides: 72.4% T. trichiura: 95% | Hookworm: 58.8% A. lumbricoides: 81.3% T. trichiura: 50% | [22] | ||
| Hookworm: 81% A. lumbricoides: 68% T. trichiura: 88% | [15] | |||
| A. lumbricoides: 84.4% | A. lumbricoides: 96.7% | [23] | ||
| Hookworm: 89.1% A. lumbricoides: 89.8% T. trichiura: 96.1% | [17] | |||
| Hookworm: 19.6% A. lumbricoides: 67.8% T. trichiura: 76.6% | Hookworm: 84.3% A. lumbricoides: 86.7% T. trichiura: 75.3% | [14] | ||
| Hookworm: 93.9% A. lumbricoides: 96.2% T. trichiura: 98.3% | [24] | |||
| Hookworm: 95.1% A. lumbricoides: 97.3% T. trichiura: 86.7% | Hookworm: 98.6% A. lumbricoides: 99.3% T. trichiura: 99.8% | [21] | ||
| McMaster | A known amount of stool is mixed with a saturated salt solution. The top layer of the solution is added to the reading chamber and the eggs are counted. | Hookworm: 67.6% A. lumbricoides: 74.3% T. trichiura: 94.6% | Hookworm: 62% A. lumbricoides: 82.4% T. trichiura: 47.8% | [22] |
| Hookworm: 92.3% A. lumbricoides: 94.9% T. trichiura: 95.2% | [24] | |||
| A. lumbricoides: 48.3% | A. lumbricoides: 91.5% | [23] | ||
| Hookworm: 80.8% A. lumbricoides: 69.5% T. trichiura: 46.7% | Hookworm: 94.6% A. lumbricoides: 92.7% T. trichiura: 99.4% | [21] | ||
| FLOTAC | A weighed amount of stool sample is homogenized and filtered with water or saline. The filtrate is mixed with a floatation solution and is added to the two FLOTAC chambers. The chambers are centrifuged and examined under a microscope. | Hookworm: 80.8% A. lumbricoides: 81.9% T. trichiura: 96.8% | Hookworm: 73.4% A. lumbricoides: 86.9% T. trichiura: 61.1% | [22] |
| Hookworm: 54% A. lumbricoides: 88% T. trichiura: 95% | [15] | |||
| Hookworm: 100% A. lumbricoides: 100% T. trichiura: 100% | Hookworm: 100% A. lumbricoides: 100% T.t richiura: 100% | [14] | ||
| Mini-FLOTAC | A weighed stool sample is homogenized with 5% formalin, filtered, and the filtrate is added to the flotation solution. The suspension is loaded into the chamber. | A. lumbricoides: 61.3% | A. lumbricoides: 93.1% | [23] |
| Hookworm: 70.8% A. lumbricoides: 42.1% T. trichiura: 85.6% | [10] | |||
| FECPAKG2 | FECPAKG2 platform with a cassette, concentrates helminth eggs into one microscopic field of view and this is photographed and is stored on a computer from which the eggs can be counted. | Hookworm: 47.5% A. lumbricoides: 36.8% T. trichiura: 37.5% | [10] | |
| Hookworm: 91.3% A. lumbricoides: 96.9% T. trichiura: 95.3% | [17] |
Table 2.
Molecular detection methods for soil-transmitted helminths.
| Molecular Technique | Target | STH Detected | Sensitivity * | Negative Predictive Value * | Reference | |
|---|---|---|---|---|---|---|
| Conventional PCR | Single-plex | Mitochondrial COI gene | A. duodenale, N. americanus | [36] | ||
| Nested PCR | Semi-nested PCR | ITS-2 | A. duodenale | [40] | ||
| 28S rRNA, ITS-2 | N. americanus | N. americanus: 94.5% | [41] | |||
| Semi-nested PCR+ RFLP | ITS-1, 2 and 5.8S region | A. duodenale, N. americanus, | [42] | |||
| T. trichiura, A. lumbricoides | ||||||
| Quantitative paper-based DNA reader | Single-plex mini-PCR | β-tubulin | T. trichiura | [43] | ||
| Real-time PCR | Single-plex | ITS-2 | A. duodenale, N. americanus | Hookworm: 78.9% | [44] | |
| Repetitive sequence | N. americanus, A. lumbricoides, T. trichiura, A. duodenale | [35] | ||||
| Multiparallel | ITS-1,2 | A. lumbricoides, T. trichiura, A. duodenale, N. americanus | A. lumbricoides: 96.7% T. trichiura: 99.2% | [45] | ||
| Repetitive sequences, ITS-1,2 | A. lumbricoides, T. trichiura, A. duodenale, N. americanus | A. lumbricoides: 98% N. americanus: 98% | [28] | |||
| Multiplex | ITS-2 | A. duodenale, N. americanus | [34] | |||
| ITS-1,2 | A. lumbricoides, Ancylostoma spp., N. americanus | [46] | ||||
| ITS-1,2 | N. americanus, T. trichiura, Ancylostoma spp., Ascaris spp. | [47] | ||||
| PCR-Luminex | ITS-1,2 | N. americanus, A. lumbricoides, A. duodenale | [48] | |||
| Melt curve analysis | ITS-2 | N. americanus, A. duodenale, A. ceylanicum, A. caninum, A. braziliense | Hookworm: 100% | [49] | ||
| 18S, ITS-1,2 | A. lumbricoides, T. trichiura, A. duodenale, N. americanus | [50] | ||||
| Multiplex-tandem PCR–qPCR | β-tubulin | A. lumbricoides, T. trichiura, A. duodenale, N. americanus | [51] | |||
| Digital PCR | ITS-1 | A. lumbricoides | [52] | |||
| Isothermal assay | SmartAmp2 | β-tubulin | N. americanus, A. lumbricoides, T. trichiura | [53] | ||
| LAMP | ITS-2 | N. americanus | N. americanus: 97% | [54] | ||
| LAMP | ITS-1 | A. lumbricoides | A. lumbricoides: 96.3% | A. lumbricoides: 88.9% | [55] | |
* Compared to microscopy.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Manuel, M.; Ramanujam, K.; Ajjampur, S.S.R. Molecular Tools for Diagnosis and Surveillance of Soil-Transmitted Helminths in Endemic Areas. Parasitologia 2021, 1, 105-118. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia1030012
AMA Style
Manuel M, Ramanujam K, Ajjampur SSR. Molecular Tools for Diagnosis and Surveillance of Soil-Transmitted Helminths in Endemic Areas. Parasitologia. 2021; 1(3):105-118. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia1030012
Chicago/Turabian StyleManuel, Malathi, Karthik Ramanujam, and Sitara S. R. Ajjampur. 2021. "Molecular Tools for Diagnosis and Surveillance of Soil-Transmitted Helminths in Endemic Areas" Parasitologia 1, no. 3: 105-118. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia1030012