
Discovery of Antigens and Cellular Mechanisms in the Protozoan Parasite Sarcocystis aucheniae Using Immunoproteomics
 , , and
, , and
Abstract
:1. Introduction
2. Results
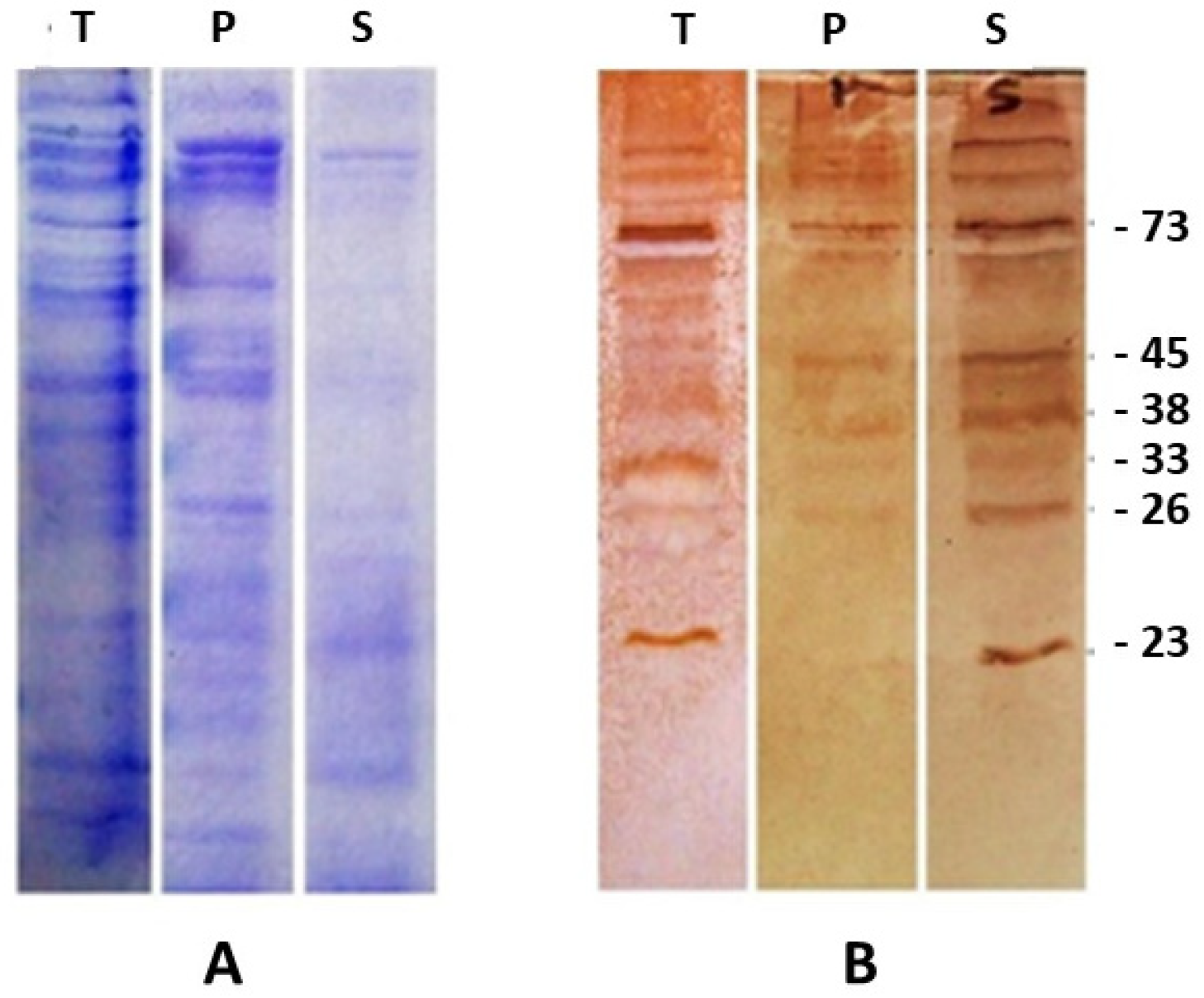
2.1. Llama Anti-S. aucheniae IgG Antibodies Recognize Six Immunoreactive Protein Bands from Sarcocysts
2.2. Sarcocystis aucheniae Immunoreactive Bands Contain Soluble B-Cell Epitopes Not Conserved in Other Coccidia
2.3. Conserved Domains Reveal Metabolic Pathways and Cellular Mechanisms Active in Sarcocyst Parasitic Stages
3. Discussion
4. Materials and Methods
4.1. Serum Samples
4.2. Electrophoretic Separation of S. aucheniae Cyst Proteins
4.3. Determination of Immunoreactivity of Llama Sera against S. aucheniae Cyst Proteins via Western Blot
4.4. Determination of Immunoglobulin Isotype Reacting with S. aucheniae Proteins
4.5. Discrimination between Soluble and Membrane-Bound Proteins by Triton X-114 Partition
4.6. Mass Spectrometry Analysis of Immunoreactive Bands
4.7. Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P.; Calero-Bernal, R.; Rosenthal, B.M.; Speer, C.A.; Fayer, R. Sarcocystosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016; ISBN 9781498710152. [Google Scholar]
- Wassermann, M.; Raisch, L.; Lyons, J.A.; Natusch, D.J.D.; Richter, S.; Wirth, M.; Preeprem, P.; Khoprasert, Y.; Ginting, S.; Mackenstedt, U.; et al. Examination of Sarcocystis spp. of Giant Snakes from Australia and Southeast Asia Confirms Presence of a Known Pathogen—Sarcocystis Nesbitti. PLoS ONE 2017, 12, e0187984. [Google Scholar] [CrossRef] [PubMed]
- Decker Franco, C.; Schnittger, L.; Florin-Christensen, M. Sarcocystis. In Parasitic Protozoa of Farm Animals and Pets; Springer International Publishing: Cham, Switzerland, 2018; pp. 103–124. ISBN 9783319701325. [Google Scholar]
- Gabor, M.; Gabor, L.J.; Srivastava, M.; Booth, M.; Reece, R. Chronic Myositis in an Australian Alpaca (Llama pacos) Associated with Sarcocystis spp. J. Vet. Diagn. Investig. 2010, 22, 966–969. [Google Scholar] [CrossRef] [PubMed]
- La Perle, K.M.D.; Silveria, F.; Anderson, D.E.; Blomme, E.A.G. Dalmeny Disease in an Alpaca (Lama pacos): Sarcocystosis, Eosinophilic Myositis and Abortion. J. Comp. Pathol. 1999, 121, 287–293. [Google Scholar] [CrossRef] [PubMed]
- MacKay, R.J.; Howe, D.K. Equine Protozoal Myeloencephalitis. Vet. Clin. Equine Pract. 2022, 38, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Sarcocystis neurona, Neospora spp. and Toxoplasma gondii Infections in Horses and Equine Protozoal Myeloencephalitis (EPM): Five Decades of Personal Experience, Perspectives and Update. Parasitology 2022, 149, 717–728. [Google Scholar] [CrossRef]
- Blazejewski, T.; Nursimulu, N.; Pszenny, V.; Dangoudoubiyam, S.; Namasivayam, S.; Chiasson, M.A.; Chessman, K.; Tonkin, M.; Swapna, L.S.; Hung, S.S.; et al. Systems-Based Analysis of the Sarcocystis neurona Genome Identifies Pathways That Contribute to a Heteroxenous Life Cycle. mBio 2015, 6, 1–16. [Google Scholar] [CrossRef]
- Rosenthal, B.M. Zoonotic Sarcocystis. Res. Vet. Sci. 2021, 136, 151–157. [Google Scholar] [CrossRef]
- Wheeler Jane, C. Evolution and Present Situation of the South American Camelidae. Biol. J. Linn. Soc. 1995, 54, 271–295. [Google Scholar] [CrossRef]
- Vilá, B.; Arzamendia, Y. South American Camelids: Their Values and Contributions to People. Sustain. Sci. 2022, 17, 707–724. [Google Scholar] [CrossRef]
- Saeed, M.A.; Rashid, M.H.; Vaughan, J.; Jabbar, A. Sarcocystosis in South American Camelids: The State of Play Revisited. Parasit. Vectors 2018, 11, 146. [Google Scholar] [CrossRef]
- Fraser, M.D.; Vallin, H.E.; Roberts, B.P. Animal Board Invited Review: Grassland-Based Livestock Farming and Biodiversity. Animal 2022, 16, 100671. [Google Scholar] [CrossRef] [PubMed]
- Leguía, G. The Epidemiology and Economic Impact of Llama Parasites. Parasitol. Today 1991, 7, 54–56. [Google Scholar] [CrossRef] [PubMed]
- Vilca, M. Producción, Tecnología e Higiene de La Carne. In Avances y Perspectivas del Conocimiento de los Camélidos Sudamericanos; FAO: Santiago de Chile, Chile, 1991; pp. 42–49. [Google Scholar]
- Popova, T.; Tejeda, L.; Peñarrieta, J.M.; Smith, M.A.; Bush, R.D.; Hopkins, D.L. Meat of South American Camelids—Sensory Quality and Nutritional Composition. Meat Sci. 2021, 171, 108285. [Google Scholar] [CrossRef] [PubMed]
- Carletti, T.; Martin, M.; Romero, S.; Morrison, D.A.; Marcoppido, G.; Florin-Christensen, M.; Schnittger, L. Molecular Identification of Sarcocystis aucheniae as the Macrocyst-Forming Parasite of Llamas. Vet. Parasitol. 2013, 198, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Moré, G.; Regensburger, C.; Gos, M.L.; Pardini, L.; Verma, S.K.; Ctibor, J.; Serrano-Martínez, M.E.; Dubey, J.P.; Venturini, M.C. Sarcocystis masoni, n. Sp. (Apicomplexa: Sarcocystidae), and Redescription of Sarcocystis Aucheniae from Llama (Lama glama), Guanaco (Lama guanicoe) and Alpaca (Vicugna pacos). Parasitology 2016, 143, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Irikura, D.; Saito, M.; Sugita-Konishi, Y.; Ohnishi, T.; Sugiyama, K.; Watanabe, M.; Yamazaki, A.; Izumiyama, S.; Sato, H.; Kimura, Y.; et al. Characterization of Sarcocystis fayeri’s Actin-Depolymerizing Factor as a Toxin That Causes Diarrhea. Genes Cells 2017, 22, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Kamata, Y.; Saito, M.; Irikura, D.; Yahata, Y.; Ohnishi, T.; Bessho, T.; Inui, T.; Watanabe, M.; Sugita-Konishi, Y. A Toxin Isolated from Sarcocystis fayeri in Raw Horsemeat May Be Responsible for Food Poisoning. J. Food Prot. 2014, 77, 814–819. [Google Scholar] [CrossRef]
- Decker Franco, C.; Wieser, S.N.; Soria, M.; de Alba, P.; Florin-Christensen, M.; Schnittger, L. In Silico Identification of Immunotherapeutic and Diagnostic Targets in the Glycosylphosphatidylinositol Metabolism of the Coccidian Sarcocystis aucheniae. Transbound Emerg. Dis. 2020, 67, 165–174. [Google Scholar] [CrossRef]
- Martin, M.; Decker Franco, C.; Romero, S.; Carletti, T.; Schnittger, L.; Florin-Christensen, M. Molecular Detection of Sarcocystis aucheniae in the Blood of Llamas from Argentina. Rev. Argent Microbiol. 2016, 48, 200–205. [Google Scholar] [CrossRef]
- Decker Franco, C.; Romero, S.; Ferrari, A.; Schnittger, L.; Florin-Christensen, M. Detection of Sarcocystis Aucheniae in Blood of Llama Using a Duplex Semi-Nested PCR Assay and Its Association with Cyst Infestation. Heliyon 2018, 4, e00928. [Google Scholar] [CrossRef]
- Moré, G.; Pardini, L.; Basso, W.; Marín, R.; Bacigalupe, D.; Auad, G.; Venturini, L.; Venturini, M.C. Seroprevalence of Neospora caninum, Toxoplasma gondii and Sarcocystis sp. in Llamas (Lama glama) from Jujuy, Argentina. Vet. Parasitol. 2008, 155, 158–160. [Google Scholar] [CrossRef] [PubMed]
- Romero, S.; Carletti, T.; Decker Franco, C.; Moré, G.; Schnittger, L.; Florin-Christensen, M. Seropositivity to Sarcocystis Infection of Llamas Correlates with Breeding Practices. Vet. Parasitol. Reg. Stud. Rep. 2017, 10, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. A Review of Coccidiosis in South American Camelids. Parasitol. Res. 2018, 117, 1999–2013. [Google Scholar] [CrossRef] [PubMed]
- Cañal, V.; Beltrame, M.O. Gastrointestinal Parasite Diversity of South American Camelids (Artiodactyla: Camelidae): First Review throughout the Native Range of Distribution. Int. J. Parasitol. Parasites Wildl. 2022, 19, 222–242. [Google Scholar] [CrossRef] [PubMed]
- Can, H.; Aksoy Gökmen, A.; Döşkaya, M.; Erkunt Alak, S.; Değirmenci Döşkaya, A.; Karakavuk, M.; Köseoğlu, A.E.; Karakavuk, T.; Gül, C.; Güvendi, M.; et al. Development of a New Serotyping ELISA for Toxoplasma gondii Type II, Type III and Africa 1 Lineages Using in Silico Peptide Discovery Methods, Well Categorized Feline and Human Outbreak Serum Samples. BMC Infect Dis. 2022, 22, 110. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.J.; Korcsmaros, T.; Carding, S.R. Mechanisms and Pathways of Toxoplasma gondii Transepithelial Migration. Tissue Barriers 2017, 5, e1273865. [Google Scholar] [CrossRef] [PubMed]
- Dangoudoubiyam, S.; Oliveira, J.B.; Víquezà, C.; Gó Mezgarcíà, A.; González, O.; Romeroà, J.J.; Kwok, O.C.H.; Dubey, J.P.; Howe, D.K. Detection of Antibodies against Sarcocystis neurona, Neospora spp., and Toxoplasma gondii in Horses from Costa Rica. J. Parasitol. 2011, 97, 522–524. [Google Scholar] [CrossRef]
- Dominguez, M.; Echaide, I.; de Echaide, S.T.; Wilkowsky, S.; Zabal, O.; Mosqueda, J.J.; Schnittger, L.; Florin-Christensen, M. Validation and Field Evaluation of a Competitive Enzyme-Linked Immunosorbent Assay for Diagnosis of Babesia bovis Infections in Argentina. Clin. Vaccine Immunol. 2012, 19, 924–928. [Google Scholar] [CrossRef]
- Dubois, D.J.; Soldati-Favre, D. Biogenesis and Secretion of Micronemes in Toxoplasma gondii. Cell Microbiol. 2019, 21, e13018. [Google Scholar] [CrossRef]
- Döşkaya, M.; Liang, L.; Jain, A.; Can, H.; Gülçe Iz, S.; Felgner, P.L.; Deǧirmenci Döşkaya, A.; Davies, D.H.; Gürüz, A.Y. Discovery of New Toxoplasma gondii Antigenic Proteins Using a High Throughput Protein Microarray Approach Screening Sera of Murine Model Infected Orally with Oocysts and Tissue Cysts. Parasit. Vectors 2018, 11, 393. [Google Scholar] [CrossRef]
- Juárez-Estrada, M.A.; Tellez-Isaias, G.; Graham, D.M.; Laverty, L.; Gayosso-Vázquez, A.; Alonso-Morales, R.A. Identification of Eimeria tenella Sporozoite Immunodominant Mimotopes by Random Phage-Display Peptide Libraries—A Proof of Concept Study. Front. Vet. Sci. 2023, 10, 1223436. [Google Scholar] [CrossRef] [PubMed]
- Frénal, K.; Dubremetz, J.F.; Lebrun, M.; Soldati-Favre, D. Gliding Motility Powers Invasion and Egress in Apicomplexa. Nat. Rev. Microbiol. 2017, 15, 645–660. [Google Scholar] [CrossRef]
- Baum, J.; Papenfuss, A.T.; Baum, B.; Speed, T.P.; Cowman, A.F. Regulation of Apicomplexan Actin-Based Motility. Nat. Rev. Microbiol. 2006, 4, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Heintzelman, M.B. Gliding Motility in Apicomplexan Parasites. Semin Cell Dev. Biol. 2015, 46, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.R.; Barrios-Arpi, M.; Rodríguez, J.; Balcázarnakamatsu, S.; Zarria, J.; Namiyama, G.; Taniwaki, N.; Gonzales-Viera, O. Ultrastructural Description of Sarcocystis sp. in Cardiac Muscle of Naturally Infected Alpacas (Vicugna pacos). Iran. J. Parasitol. 2019, 14, 174–179. [Google Scholar]
- Blackman, M.J.; Bannister, L.H. Apical Organelles of Apicomplexa: Biology and Isolation by Subcellular Fractionation. Mol. Biochem. Parasitol. 2001, 117, 11–25. [Google Scholar] [CrossRef]
- Mercier, C.; Adjogble, K.D.Z.; Däubener, W.; Delauw, M.F.C. Dense Granules: Are They Key Organelles to Help Understand the Parasitophorous Vacuole of All Apicomplexa Parasites? Int. J. Parasitol. 2005, 35, 829–849. [Google Scholar] [CrossRef]
- Sajid, M.; Blackman, M.J.; Doyle, P.; He, C.; Land, K.M.; Lobo, C.; Mackey, Z.; Ndao, M.; Reed, S.L.; Shiels, B.; et al. Proteases of Parasitic Protozoa—Current Status and Validation. In Antiparasitic and Antibacterial Drug Discovery: From Molecular Targets to Drug Candidates; Wiley-VCH Verlag GmbH & Co.: Weinheim, Germany, 2009; pp. 175–209. [Google Scholar] [CrossRef]
- Florin-Christensen, M.; Sojka, D.; Ganzinelli, S.; Šnebergerová, P.; Suarez, C.E.; Schnittger, L. Degrade to Survive: The Intricate World of Piroplasmid Proteases. Trends Parasitol. 2023, 39, 532–546. [Google Scholar] [CrossRef]
- Hajagos, B.E.; Turetzky, J.M.; Peng, E.D.; Cheng, S.J.; Ryan, C.M.; Souda, P.; Whitelegge, J.P.; Lebrun, M.; Dubremetz, J.F.; Bradley, P.J. Molecular Dissection of Novel Trafficking and Processing of the Toxoplasma gondii Rhoptry Metalloprotease Toxolysin-1. Traffic 2012, 13, 292–304. [Google Scholar] [CrossRef]
- Xie, S.C.; Dick, L.R.; Gould, A.; Brand, S.; Tilley, L. The Proteasome as a Target for Protozoan Parasites. Expert Opin. Ther. Targets 2019, 23, 903–914. [Google Scholar] [CrossRef]
- Soto, A.S.; Fenoy, I.M.; Sanchez, V.R.; March, F.; Perrone Sibilia, M.D.; Aldirico, M.D.L.A.; Picchio, M.S.; Arcon, N.; Acosta, P.L.; Polack, F.P.; et al. Toxoplasma gondii Serine-Protease Inhibitor-1: A New Adjuvant Candidate for Asthma Therapy. PLoS ONE 2017, 12, e0187002. [Google Scholar] [CrossRef]
- Fleige, T.; Pfaff, N.; Gross, U.; Bohne, W. Localisation of Gluconeogenesis and Tricarboxylic Acid (TCA)-Cycle Enzymes and First Functional Analysis of the TCA Cycle in Toxoplasma gondii. Int. J. Parasitol. 2008, 38, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Ghaffar, F.; Bashtar, A.R.; Al-Quraishy, S.; Al Nasr, I.; Mehlhorn, H. Sarcocystis Infecting Reptiles in Saudi Arabia: 11-Light and Electron Microscopic Study on Sarcocysts of Sarcocystis turcicii sp. nov. Infecting the Gecko Hemidactylus turcicus linnaeus. Parasitol. Res. 2009, 104, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Qian, W.; Li, X.; Wang, T.; Ding, K.; Huang, T. Morphological and Molecular Characterization of Sarcocystis miescheriana from Pigs in the Central Region of China. Parasitol. Res. 2013, 112, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Ashwinder, K.; Kho, M.T.; Chee, P.M.; Lim, W.Z.; Yap, I.K.S.; Choi, S.B.; Yam, W.K. Targeting Heat Shock Proteins 60 and 70 of Toxoplasma gondii as a Potential Drug Target: In Silico Approach. Interdiscip. Sci. 2016, 8, 374–387. [Google Scholar] [CrossRef] [PubMed]
- Anas, M.; Kumari, V.; Gupta, N.; Dube, A.; Kumar, N. Protein Quality Control Machinery in Intracellular Protozoan Parasites: Hopes and Challenges for Therapeutic Targeting. Cell Stress Chaperones 2019, 24, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Welti, R.; Mui, E.; Sparks, A.; Wernimont, S.; Isaac, G.; Kirisits, M.; Roth, M.; Roberts, C.W.; Botté, C.; Maréchal, E.; et al. Lipidomic Analysis of Toxoplasma gondii Reveals Unusual Polar Lipids. Biochemistry 2007, 46, 13882–13890. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Liang, X.; Brouwers, J.F.; Miron, R.C.; Shen, B.; Gupta, N. Synthesis vs. Salvage of Ester- and Ether-Linked Phosphatidylethanolamine in the Intracellular Protozoan Pathogen Toxoplasma gondii. Commun. Biol. 2023, 6, 306. [Google Scholar] [CrossRef]
- Pszenny, V.; Ehrenman, K.; Romano, J.D.; Kennard, A.; Schultz, A.; Roos, D.S.; Grigg, M.E.; Carruthers, V.B.; Coppens, I. A Lipolytic Lecithin:Cholesterol Acyltransferase Secreted by Toxoplasma Facilitates Parasite Replication and Egress. J. Biol. Chem. 2016, 291, 3725–3746. [Google Scholar] [CrossRef]
- Li, L.; Brunk, B.P.; Kissinger, J.C.; Pape, D.; Tang, K.; Cole, R.H.; Martin, J.; Wylie, T.; Dante, M.; Fogarty, S.J.; et al. Gene Discovery in the Apicomplexa as Revealed by EST Sequencing and Assembly of a Comparative Gene Database. Genome Res. 2003, 13, 443–454. [Google Scholar] [CrossRef]
- Hunt, A.G.; Howe, D.K.; Brown, A.; Yeargan, M. Transcriptional Dynamics in the Protozoan Parasite Sarcocystis neurona and Mammalian Host Cells after Treatment with a Specific Inhibitor of Apicomplexan MRNA Polyadenylation. PLoS ONE 2021, 16, e0259109. [Google Scholar] [CrossRef]
- Friedrich, A.; Ledesma, M.; Landone, I.; Ferrari, A.; Leoni, J. Production of a Monoclonal Antibody against Serum Immunoglobulin M of South American Camelids and Assessment of Its Suitability in Two Immunoassays. J. Vet. Diagn. Investig. 2014, 26, 646–650. [Google Scholar] [CrossRef]
- Wang, L.; Coppel, R.L. Triton X-114 Phase Partitioning Triton X-114 Phase Partitioning for Antigen Characterization. In Malaria Methods and Protocols: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2002; pp. 581–585. [Google Scholar]
- Yao, B.; Zheng, D.; Liang, S.; Zhang, C. SVMTriP: A Method to Predict B-Cell Linear Antigenic Epitopes. Methods Mol. Biol. 2020, 2131, 299–307. [Google Scholar] [CrossRef]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving Sequence-Based B-Cell Epitope Prediction Using Conformational Epitopes. Nucleic Acids Res. 2017, 45, W24–W29. [Google Scholar] [CrossRef]
- Madden, T. The BLAST Sequence Analysis Tool; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2013. [Google Scholar]
- McGinnis, S.; Madden, T.L. BLAST: At the Core of a Powerful and Diverse Set of Sequence Analysis Tools. Nucleic Acids Res. 2004, 32, W20–W25. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. Clustal Omega. Curr. Protoc. Bioinform. 2014, 48, 3.13.1–3.13.16. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating Signal Peptides from Transmembrane Regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L.L. Predicting Transmembrane Protein Topology with a Hidden Markov Model: Application to Complete Genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef]
- Eisenhaber, B.; Bork, P.; Eisenhaber, F. Prediction of Potential GPI-Modification Sites in Proprotein Sequences. J. Mol. Biol. 1999, 292, 741–758. [Google Scholar] [CrossRef]
- Fankhauser, N.; Mäser, P. Identification of GPI Anchor Attachment Signals by a Kohonen Self-Organizing Map. Bioinformatics 2005, 21, 1846–1852. [Google Scholar] [CrossRef]
- Pierleoni, A.; Martelli, P.; Casadio, R. PredGPI: A GPI-Anchor Predictor. BMC Bioinform. 2008, 9, 392. [Google Scholar] [CrossRef] [PubMed]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro Protein Families and Domains Database: 20 Years On. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The Protein Families Database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Band Size | Protein ID | Antigenicity Index | B-Cell Epitope | Algorithm | Score | Water Solubility | Cross-Reactivity Propensity |
|---|---|---|---|---|---|---|---|
| 23 kDa | OR538339 | 0.64 | SATLVLYSSMVSHLKRDT | S | 1.000 | Poor | no |
| IELALDLTDKGSEDIERV | S | 0.942 | Good | no | |||
| RDTEILANSIRDFAVRYA | S | 0.917 | Good | yes | |||
| DINGYEV | B | Good | no | ||||
| NVQSTMQ | B | Poor | no | ||||
| 26 kDa | OR538349 | 0.9 | EKRTLSSCTLATIEIDSL | S | 1.000 | Good | yes |
| EARGGTN | B | Good | yes | ||||
| 33 kDa | OR538360 | IRLGLALNYSVFFYEILN | S | 1.000 | Poor | yes | |
| 0.93 | TLIMQLLRDNLTLWTSDL | S | 0.942 | Poor | yes | ||
| DEEEKAAE | B | Good | yes | ||||
| 38 kDa | OR538361 | 0.87 | RCVLKIGEHTPSSLAIME | S | 1.000 | Good | no |
| VRSLSPVADPRSSTATPRQP | B | Good | yes | ||||
| 45 kDa | OR538336 | 0.93 | RRCLNGNLVENRMCLEEE | S | 1.000 | Good | no |
| GEGVGGGVGGGAA | B | Poor | yes | ||||
| PPAAEGGAP | B | Good | yes | ||||
| GDEAGAGE | B | Good | yes | ||||
| 73 kDa | OR538343 | 0.93 | VQKDTELHTYDFSRLKWN | S | 1.000 | Good | no |
| Process | Protein Name | Protein ID | Comment |
|---|---|---|---|
| Motility, host cell invasion, and cyst formation | Surface protein (SRS domain) | OR538334 | Host cell recognition and attachment |
| OR538335 | |||
| MIC 12 | OR538336 | Glideosome, host cell invasion | |
| GAP-45 | OR538337 | ||
| Dense granule protein GRA9 | OR538338 | Cyst formation | |
| Host protein degradation for nutrition and egress from host cells | Secreted metalloprotease | OR538339 | M16; in T. gondii: toxolysin 1 |
| Trypsin | OR538340 | S1 serine protease | |
| Leucine aminopeptidase | OR538341 | M17 metalloprotease | |
| Alanyl aminopeptidase | OR538342 | M1 metalloprotease | |
| Serine peptidase | OR538343 | S9 prolyl oligopeptidase | |
| Protein synthesis, regulation, and processing | Elongation factor 2 | OR538344 | Protein synthesis |
| Ubiquitin | OR538345 | Tagging of proteins for processing in the proteasome | |
| Threonine protease | OR538346 | T1; protein degradation in the proteasome | |
| Heat-shock protein 60 | OR538347 | Chaperone: protein folding | |
| Heat-shock protein 70 | OR538348 OR538349 | ||
| Heat-shock protein 90 | OR538350 | ||
| Serpin | OR538351 | Serine protease inhibitor | |
| Energy storage | Fructose 1,6 bisphosphatase | OR538352 | Gluconeogenesis |
| Lipid metabolism | Phosphatidylserine decarboxylase | OR538353 | Phosphatidylethanolamine synthesis |
| Phosphatidylcholine-sterol O-acyltransferase | OR538354 | Cholesteryl ester synthesis, phospholipase A2 activity? | |
| Cell division and assorted metabolic pathways | Tudor domain-containing protein | OR538355 | RNA metabolism, DNA repair |
| Chromosome-segregation protein | OR538356 | Chromosome partition | |
| Pyridine nucleotide-disulphide oxidoreductase | OR538357 | Redox reactions | |
| Histidine phosphatase | OR538358 | Metabolic regulation and development | |
| DnAK-TPR | OR538359 | Protein–protein interactions | |
| 14-3-3 protein | OR538360 | Signal transduction | |
| Cellular respiration | Glyceraldehyde 3-phosphate dehydrogenase | OR538367 | Glycolysis |
| Fructose-bisphosphate aldolase | OR538361 | ||
| Enolase 2 | OR538362 | ||
| Lactate/malate dehydrogenase | OR538363 | Tricarboxylic acid cycle | |
| Membrane-bound ATPase | OR538364 | ATP production in the electron transport chain | |
| Protection from free radicals | Superoxide dismutase | OR538365 | Antioxidant |
| Unknown | Conserved hypothetical protein | OR538333 | Unknown |
| Hypothetical protein | OR538366 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wieser, S.N.; Decker-Franco, C.; de Alba, P.; Romero, S.; Ferrari, A.; Schnittger, L.; Florin-Christensen, M. Discovery of Antigens and Cellular Mechanisms in the Protozoan Parasite Sarcocystis aucheniae Using Immunoproteomics. Parasitologia 2023, 3, 349-363. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3040034
Wieser SN, Decker-Franco C, de Alba P, Romero S, Ferrari A, Schnittger L, Florin-Christensen M. Discovery of Antigens and Cellular Mechanisms in the Protozoan Parasite Sarcocystis aucheniae Using Immunoproteomics. Parasitologia. 2023; 3(4):349-363. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3040034
Chicago/Turabian StyleWieser, Sarah Nathaly, Cecilia Decker-Franco, Paloma de Alba, Sandra Romero, Alejandro Ferrari, Leonhard Schnittger, and Mónica Florin-Christensen. 2023. "Discovery of Antigens and Cellular Mechanisms in the Protozoan Parasite Sarcocystis aucheniae Using Immunoproteomics" Parasitologia 3, no. 4: 349-363. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3040034