
Chemical Composition of Apples Cultivated in Norway


1
Faculty of Chemistry, Biotechnology and Food Science, Norwegian University of Life Sciences, 1433 Aas, Norway
2
INRAE, UR1268 BIA (Biopolymers, Interactions & Assemblies), PRP Group (Polyphenols, Reactivity, Processes), F-35653 Le Rheu, France
3
UMT ACTIA Nova2Cidre, F-35653 Le Rheu, France
*
Author to whom correspondence should be addressed.
Crops 2021, 1(1), 8-19; https://0-doi-org.brum.beds.ac.uk/10.3390/crops1010003
Submission received: 6 April 2021
/
Revised: 10 May 2021
/
Accepted: 13 May 2021
/
Published: 17 May 2021
Abstract
:The composition of apples varies with both cultivar and horticultural practice. Knowledge about the chemical composition of different cultivars in particular sugars, organic acids, nitrogen compounds and polyphenols is essential, since they are directly related to the progress of fermentation monitoring and the organoleptic qualities of produced ciders. Fifteen apple cultivars grown in two locations in Norway were investigated for their chemical composition, including polyphenol profiles. The nitrogen content varied from 43.2 to 171.4 mg N/L between the cultivars, and the sum of free amino acids varied from 42.4 to 924.5 mg/kg. Asparagine, aspartic acid and glutamic acid were the dominating amino acids. Flavanols, consisting of catechins and procyanidins, were the dominating polyphenols, followed by hydroxycinnamic acids. The cultivar Bramley Seedling was highest in the sum of polyphenols by HPLC (1838 mg/L) and relatively low in nitrogen content (75 mg N/L). Summerred was lowest in the sum of polyphenols (87 mg/L) and highest in nitrogen (171.4 mg N/L). Sugar content, measured as density in the juice, varied between 1034 and 1060 g/L. Using cider categorization of apples, ’Bramley Seedling’ would be defined as sharp, while the others are sweet cultivars.
1. Introduction
Apples are grown in temperate zones and are a common fruit all over the world. Apples are a rich source of phytochemicals, and epidemiological studies have linked consumption of apples to have positive effects on many lifestyle diseases [1].
Apples can be specially grown for fresh consumption or for production of beverages like juice, wine or cider. Cultivar, horticultural practice and year of cropping are important factors determining the chemical composition of apples. The chemical compounds might also respond differently to thermal, mechanical and biochemical processes. Polyphenols are important for flavor and aroma in apples, as well as in processed products. Apple content of phytochemicals, including flavonols (mainly quercetin glycosides), flavan-3-ols (catechins and procyanidins), dihydrochalcones (essentially phloretin glycosides) and hydroxycinnamic acids (mainly chlorogenic acids), depends on cultivar and climatic conditions, especially during maturation of the fruits [2]. Guyot et al. [3] found that procyanidins were the most abundant group of polyphenols in apples, both in the flesh and in the peel, while flavonols were mostly found in the apple peel.
The interest in craft cider has grown steadily in the last several years. Knowledge of the fruit quality and variation between cultivars is, therefore, important for choosing the proper raw material. Content of various polyphenols, combined with acidity, defines important quality parameters for apples. These also define the potential use of the various apple cultivars for cider production. The total polyphenol content associated with the total acidity of the must is used to classify French and English cider apple cultivars in different taste categories [4]. Most of the polyphenols contribute to the final colour of the cider, since they are directly involved in the enzymatic oxidation phenomenon that leads to the formation of coloured oxidation products during apple crushing and pressing. More specifically, tannins (procyanidin oligomers and polymers) are responsible for bitterness and astringency that are important for taste and mouthfeel in those products [3].
Content of polyphenols varies between cultivars, year of harvest and postharvest storage conditions [5]. The most common polyphenols in apples are chlorogenic acid, caffeic acid, procyanidin B2, (−)-epicatechin and quercetin. Apples also exhibit antioxidant activity [6]. However, Valavanidis et al. did not find any significant difference between antioxidant activity or phenolic compounds in apples grown in organic or conventional agriculture [7]. Tsao et al. concluded that antioxidant activity was positively correlated with total phenolic concentration in apples, and that the flavan-3-ols/procyanidins were the most important contributors to the in vitro antioxidant capacity of apples. Among them, procyanidin B2 and epicatechin were the most important individual contributors [8].
It is often believed that older varieties have higher nutrient content than newer varieties. Jakobek et al. characterised old apple varieties from the Balkans and found relatively high contents of polyphenols, while Wojdylo et al. found no difference between old and new varieties in their study [9,10].
Nitrogen content in apples varies between cultivars, age of the apple tree and, mainly, horticultural practices with respect to soil, climate and fertilisation. Kahle et al. found that local growing conditions were more important than cultivar for nitrogen content in the apples [11]. Uptake of nitrogen in apples and leaves depends on soil quality, what type of fertilizer that is used and also in which period of the growing season the fertilization is done [12]. In a study by Karl et al. [13], they found that fertilizer treatment of the soil had no effect on acidity and total polyphenol concentration in the apple juice while concentration of yeast assimilable nitrogen (YAN) increased. This, in turn, might affect the fermentation rate during cider processing. YAN includes free amino nitrogen (FAN), ammonia and ammonium [14]. FAN is a measure of amino acids and small peptides that can be utilized by the yeast in the fermentation process. In a study by Boudreau et al. [15], they found a correlation between content of FAN and YAN. On average FAN comprised 85% total YAN. If the YAN concentration is low, it is possible that fermentation rate will be low, which can result in development of unwanted sulphur compounds. In a different study, they found that addition of methionine decreased H2S production when fermenting with some yeast strains, especially if YAN was low, but also an increase at specific YAN concentrations [16]. Thus, the YAN concentration in the fermenting liquid seems to be important for flavour development, positively or negatively.
Di Maro et al. found that the main amino acids in apples were asparagine, aspartic acid and glutamic acid, and that these amino acids accumulate when nitrogen is available, depending on horticultural conditions [17], while Wu et al. [18] reported that asparagine was the most important and serine the second most important amino acid. Ma et al. [14] found that, on average, asparagine was the most abundant amino acid in their study of 13 different cultivars of apples, and ranged phenylalanine as the second most important, while Zuo et al. and Ye et al. [19,20] did not report on any asparagine content in apple juice for cider production.
Alcoholic and malolactic fermentation reduce the content of most amino acids, the main nitrogenous nutrients for yeast. When initial nitrogen content of juice is low enough the cider can be quite stabile against microbial growth after fermentation (French craft cider making method) [21].
Sugar and nitrogen content in apples are often negatively correlated, and late maturating apples are often higher in sugar content due to starch hydrolysis and, for some cultivars, water loss during maturation.
The apple tree originates from the Caucasus and has been cultivated for thousands of years. It is now grown on almost all continents, mostly in temperate and cooler climates. The apple was known in Scandinavia 1000 years ago. Wild apples are found all along the coastline nearly up to the Arctic Circle. There is a long tradition for using apples in various dishes and beverages. In Norway, especially in Hardanger region, commercial production of cider has been known since the end of 1700. Today, there is increased interest in renewing the old traditions of using apples as raw materials for juice, cider and apple spirit. The producers frequently use a large variation of cultivars available on the market. Of particular interest are the cultivars Gravenstein and Aroma, the most important apple cultivars for cider production in Norway [22]. Most of the cider production is based on dessert apples commonly grown in Norway, but import and cultivation of cider apple trees to Norway to obtain apple cultivars of specific quality for cider production is at the experimental stage. Norwegian apples are, in general, rather acidic and low in bitterness. If sugar content is low, external sugar can be added. Mixing of apples to balance bitterness, acidity and sweetness is common. In France and England cider apples are cultivated for the purpose of cider production, while the use of dessert apples is more common in, for example, North America, China and some European countries [23,24].
The aim of this study was to characterize the biochemical composition of some apple cultivars grown in Norway for potential use in cider production. It was also of interest to investigate whether there were differences between apple varieties that could be characterized as new or old cultivars, and whether place of growth could have an impact on quality. This study adds useful information on apples grown in the Nordic/arctic climate.
2. Materials and Methods
2.1. Apples
Seventeen samples of apples were grown in two experimental orchards in Norway A: Akershus/south-east (10°77′ E. 59°67′ N at the NMBU/Norwegian University of Life Sciences) and H: Hardanger/west (6°66′ E. 60°32′ N at the Nibio Ullensvang Norwegian Institute of Bioeconomy Research) were harvested in September and October 2013 and stored at 2 °C until pressing at full maturity in December. Hardanger is an important region for cultivation of fruits and berries. The climate in Hardanger is characterized by mild winters and cool summers, and the region is known for the narrow fiords and steep growing locations. The annual rainfall is approximately 3000 mm. In the experimental field in Akershus, the winters are colder and the summers warmer, and the area for cultivation is more open farmland. The annual rainfall is approximately 750 mm. The orchards were all fertilized using complete fertilizer “12-4-18” (chlorine-poor, high in phosphorus, potassium, calcium, magnesium, and sulphur), 250 g per tree.
Approximately 12 kg of each cultivar were harvested from two or three trees per cultivar. The cultivars were: Aroma Amorosa (A), Aroma (A + H), Bramley Seedling (H), Delcorf (A), Elstar (A), Filippa (A), Gravenstein (A + H), James Grieve (H), Jonagold (A), Mutsu (A), Rubin (A), Summerred (A), Sunrise (A), Torstein red (A) and Torstein (H).
More detailed description of all apple varieties is shown in Table 1.
2.2. Sample Preparation
Apple Juice Extraction and Sampling
Maturity of the apples was analyzed by dipping one slice from each cultivar of apples into an iodine solution and comparing the color changes to a CTIFL color chart. All samples showed complete starch regression at the time of pressing. Three replicates of 2 to 2.5 kg of apple for each variety were pressed to produce juice for chemical analyses. The apples were mashed and pressed using a small laboratory press (model HP5, 5 L, Hafico, Fischer & Co. Dusseldorf, Germany). The hydraulic pressure rose to 400 bars giving a maximum pressure of 24 bars in the press cake. The pressing time was 15 min. The juice was then sampled for the different analysis and kept frozen. NaF (sodium fluoride; 1 mg/mL juice) was added to samples for HPLC analyses of polyphenols. NaF was used to stop enzymatic oxidation immediately after pressing by inhibition of polyphenoloxidase [25]. Samples were either freeze-dried or thawed before analyses.
2.3. Chemical Analyses
2.3.1. UV-MS Reversed Phase HPLC of Polyphenols Directly or after Phloroglucinolysis Reaction
Direct analysis of simple polyphenols in apple juice: freeze-dried juice samples (0.5 mL) were dispersed by sonication for 20 min in 1.2 mL MeOH-Ac (1% v/v).
Samples were filtered (syringe filter; 0.45 µm PTFE) before HPLC analyses. The HPLC procedure was carried out using reversed-phase analyses using a RP18 column and UV-visible and mass spectrometry (MS) detection. Phenolic compounds were identified based on their retention time, UV-visible spectrum, full MS characteristics and MS fragmentation pattern in comparison with available standards.
Phloroglucinolysis: acidolysis in the presence of phloroglucinol allows the complete depolymerisation of flavanol oligomers and polymers (i.e., procyanidins and condensed tannins), giving both qualitative and quantitative information related to those major polyphenolic compounds in complex matrixes [26,27]. A freeze-dried juice sample (0.5 mL) was dispersed in 0.8 mL PLG (phloroglucinol) and 0.4 mL MeOH/HCl (0.3 M) by agitation in a vortex mixer then the tubes were set in a water bath at 50 °C for 30 min and the reaction was immediately stopped by cooling the tubes on ice and adding 1.2 mL sodium acetate. The reaction mixture was filtered on a PTFE 0.45 µm filter before HPLC analysis. Total flavanols (including catechins and procyanidins) were quantified as previously described by Malec et al. [26].
2.3.2. Total Phenols (Folin Ciocalteu Assay)
Total phenols were analysed by the Folin Ciocalteu method (ISO14502-1). The reaction was carried out by mixing 0.5 mL of apple juice (diluted 1:10) and 2.5 mL 10% Folin Ciocalteu reagent (diluted in water) and 2.0 mL buffer (7.5% Na2CO3). Samples were incubated at ambient temperature for 60 min. The absorbance was read at 765 nm. Samples were analysed in triplicate. Standard solutions (epicatechin—1 g/L in water) for the standard curve were constructed. Quantifications were obtained using a calibration curve of epicatechin (EPI) and expressed as mg/L EPI equiv.
2.3.3. Density (d20/20)
Density measurements were used to estimate the sugar concentration in the juice. The relative density was determined using a Density Meter, DMA 58 (Anton Paar). Results were expressed as g/L.
2.3.4. pH
pH was measured using a pH-meter (Radiometer PHM 92. Copenhagen).
2.3.5. Nitrogen
Total nitrogen in the apple juice was analysed by the Kjeldahl method, according to IDF 2001.
2.3.6. Free Amino Acids (FAA)
Free amino acids were analysed using HPLC with o-phthaldialdehyde (OPA) and fluorenyl methyl chloroformate (FMOC) derivatisation according to a method described by Moe et al. [28]. The amino acids were identified and quantified by comparing standard solutions of the following amino acids: aspartic acid, glutamic acid, asparagine, serine, glutamine, histidine, glycine, threonine, citrulline, arginine, alanine, GABA, tyrosine, valine, methionine, isoleucine, phenylalanine, tryptophan, leucine, ornithine and lysine. Norvaline was used as an internal standard. Sum of amino acids are reported as mg/kg. Percentage share of only the most important amino acids is reported.
2.4. Statistical Analyses
The results are expressed as mean ± standard deviation (STD) from two to five independent measurements. A correlation test, one-way analysis of variance (ANOVA) and principal component analysis (PCA) were performed to find differences between varieties, groups of apple varieties (old and new, sharp and dessert apples) and growing sites in terms of chemical composition in the juice of the various apples. Statistical analyses were performed with Minitab statistical software version 17 (Minitab Ltd., State College, PA, USA).
3. Results and Discussion
Parentage, origin, and harvest location for the 17 samples of apples included in the experiment are presented in Table 1. Fruit weight was measured before pressing the apples, while juices from all samples were analysed for content of nitrogen, free amino acids (FAA), density, pH, total polyphenols (TP-Folin) and detailed polyphenolic profiles (by direct RP18-HPLC and RP18-HPLC after phloroglucinolysis) (Table 2, Table 3 and Table 4).
3.1. Apples
Significant differences were observed between the samples for the different chemical compounds (Table 2, Table 3 and Table 4), though we found no difference between groups of old and new cultivars nor on growing location (grouped in Table 1) for the studied composition. Climate and topography for the two growing locations are very different. However, this did not appear to have any effect on the chemical composition of the apple juice. This is in accordance with other studies [10,29]. Le Bourvellec et al. also found that cultivar and year of cropping were the main factors for determining the quantitative primary and secondary metabolite composition of apples, and horticultural practice was of less importance [30]. The cultivar Bramley Seedling was significantly different from the other cultivars included in this experiment (see PCA plot in Figure 1a,b). This was mainly related to the high content of phenolic compounds, expressed in Figure 1a, and the low pH. For nitrogen, FAA, and density Bramley Seedling was below average. In Figure 1b, where TP Folin, density, total nitrogen, FAA, and pH is included, Bramley Seedling still appears to be different from the other cultivars, but less pronounced.
We found the average fruit weight to be between 80 to 286 g per apple (Table 2), which is in line with results reported by Thompson-Witrick et al. in their study on North American apples for cider production [31]. In our experiment the cultivar Torstein (H) had the lowest apple fruit weight and was the apple sample with the lowest content of nitrogen. On the opposite side, Mutsu was also relatively low in nitrogen but with double fruit weight compared to Torstein (H). This indicates that there is not always a connection between low fruit weight and low nitrogen content in the apples. It can also be affected by, for example, growing conditions, horticulture practice, the age of the apple trees, the amount of fruit in the trees, soil, fertilization, pruning and local climatic variations, which were not registered in this study.
Size of the apples can be linked to genetic differences between cultivars or horticulture practice. The need for N-supplement depends on tree nitrogen requirement, how much nitrogen comes from the soil and uptake and efficiency of the fertilizer used. In a study on nitrogen fertilization of apple orchards Cheng et al. [12] found that N-supply on average increased fruit size. It affected leaf area to fruit ratio, producing larger fruit size and higher soluble solids. Wrona [32] found that nitrogen fertilization did not affect the fruit size in their study on Jonagored apple trees while Nava et al. [33] concluded that fruit size was more affected by potassium (K) than nitrogen (N) fertilization. Karl et al. [13] found that fertilization had no effect on acidity and polyphenol content, but YAN increased by the use of nitrogen fertilization. Ernani et al. [34] found that nitrogen in the fruit was unaffected by N addition and suggested that apple nitrogen could be linked to the local soil organic matter. Total polyphenol content was found to be inversely related to nutrient status of the tree, and apple juice from trees in old orchards were usually lower in soluble nitrogen compared to intensively cultivated orchards [35]. This may also be the effect of higher nutrient content in the apples preventing sluggish fermentation, thus increasing the production of fusel alcohols and esters as reported by dos Santos et al. [36].
3.1.1. Chemical Composition of Apple Juice
The pH of the apple juice varied between 2.9 and 3.4 Bramley Seedling being the most acidic cultivar. A desirable pH for apples for cider production is between 3.2 and 3.8. Higher pH values are not suitable for cider processing since they may favour development of microorganisms responsible for organoleptic spoilages during the fermentation process.
Sugar contents is highly correlated to the density and the density value is used both to characterise juices for sugar content and to check fermentation stage. The density of the studied apple juices was between 1034 and 1060 g/L (Table 2). This is in the same range as reported by Thompson et al. (USA) but much lower than suggested by Jolicoeur, 1060–1075 g/L, for production of a good cider (Canadian tradition) [31,37]. As a comparison, a density of 1047 corresponding to near 100 g/L of sugar and yielding 6% vol. of alcohol when fermented, is considered by French regulation as the lowest amount of sugar for a cider. This parameter can be highly influenced by climate (number of sunny days) and by the fruit/leaf ratio in the tree.
Content of nitrogen in the juices in our experiment varied from 43.2 to 171.4 mg/L between the apple samples (Torstein (H) was lowest and Summerred (A) was highest) (Table 3). Lea [4] indicated 100 mg N/L to be the ideal level for a proper fermentation process, referring to the English and French cider tradition. The fermentation process is slow, and this can contribute to the characteristic flavour of the cider obtained from the traditional cider process. At higher nitrogen content, the fermentation goes faster, which often is preferred in modern or industrial cidermaking. The cultivars Aroma (H), Delcorf, Filippa, James Grieve, Rubin, Sunrise, Summerred and Torstein red (A) were all above average in nitrogen content and probably contribute to a faster fermentation process.
Sum of FAA varied between 42.4 and 924.5 mg/kg, Torstein (H) was lowest and Summerred was highest, and significantly different from all the other samples. Filippa and Torstein red (A) were also relatively high in content of FAA with 683.2 mg/kg, and 613.9 mg/kg respectively. We found a high correlation between content of total nitrogen and the amino acid asparagine (0.876, p < 0.001). On average, asparagine accounted for 47.6% of the total amino acids, aspartic acid for 25.7% and glutamic acid for 13.8% of the total amino acids in the juices, though large variations between the cultivars were observed (Table 3). For all the cultivars contents of phenylalanine and methionine were minor or not detected. For Filippa the percentage of asparagine was 69.7% and for Torstein (H) it was 15.9%. For Torstein (H) aspartic acid was the predominant amino acid (36.3%).
Content of total polyphenols (Folin) in our apple juices varied between 272 to 1934 mg/L EPI equiv (Table 4). If excluding the cultivar Bramley Seedling, which was found to be significantly different from the other cultivars, the variation was between 272 and 990 mg/L EPI equiv. Thompson-Witrick et al. reported on a variation from 229–926 mg/L GAE equiv in their study on North American apples, which is comparable to our results without Bramley Seedling. Logically, we also found a high correlation between TP-Folin and RP18-HPLC analysis of polyphenols (r = 0.923, p < 0.001).
Composition of polyphenols in apples, juices and ciders has been extensively studied [10,11,38,39]. The content is related to apple cultivars, years and cultivation locations, and can be as high as several thousand mg/L. Average content of the sum of total polyphenols by RP18-HPLC analysis in the Norwegian apple juices in this experiment was 450 mg/L with a variation between 87 to 1838 mg/L (Table 4). Except for the two cultivars, Summerred (A) and Torstein red (A), which were relatively low (87 mg/L), the content was above 209 mg/L for the rest of the samples. This is comparable to the lower level of polyphenols for both dessert apples and cider apples from many European countries [10,11,40].
The largest group of polyphenols in the juice was the flavanols, which accounted for 55% of the total amount. Hydroxycinnamic acid accounted for 40%, dihydrochalcones for 4 % and flavonols for 1% of the total polyphenols. Content of flavonols was very low, but this group of polyphenols is mainly found in the peel and consequently the content in juice is low [3]. The proportion between the different groups seems to be similar to other studies [3,41].
Apples for cider production are usually grouped as sharp, bitter sharp, bittersweet and sweet [4]. A mixture of fruits or juices from different apple qualities is often necessary to obtain a good juice for cider production, based on the content of sugar, acids, tannins, nitrogen and polyphenols. Knowledge on chemical composition in various apple qualities is important for the cider producers to select the proper cultivars and mixtures to obtain the desired cider quality. The most important contributors to cider quality are apple cultivar, mode of fermentation and yeast strain, while ripening stage of the apples seems to be more variety specific [2,42,43]. Maturation level for the apples at time of harvest is important for biochemical components such as sugar/starch and pectin as well as for precursors for cider aroma. On the other hand, post-harvest maturation and starch degradation are important for further cider processes, such as the keeving process [35].
Tannin level in traditional European cider apples is usually higher than in apples used for hard ciders from North America. Tannin is important for bitterness and astringency of the product and determines the cider style. Jolicoeur grouped tannin content as low: <1.5 g/L, medium: 1.5–2.5 g/L and high: >2.5 g/L [37]. Using TP-Folin as an equivalent method, only Bramley Seedling in this experiment was in the medium level of tannin, and all the others were on the low level.
Polyphenols are known to be important for the flavour of cider, especially bitterness and astringency. Fermentation is not considered to drastically change the polyphenolic composition, at least when using single Saccharomyces strains [44], though the effect of fermentation on a single polyphenol might depend on the cultivar [2,20,42]. However, as there is a strong interaction between sugary and bitter tastes, fermentation enhances bitterness perception, and to a lesser extent astringency, by decreasing sugar content [45]. How fermentation affects the polyphenol composition and the following sensory attribute in the final product is difficult to predict, and trials to obtain more knowledge are of interest.
3.1.2. Potential Use of Apples for Cider
Very few apple cultivars can be used for single cultivar cider, and mixing of juice from various qualities is common by many producers so that each cultivar need not be equilibrated in the various components. If the sugar content is low, alcohol production will also be low and will result in poor cider, so cultivar and conditions producing high amount of sugar are preferred. However, addition of sugar is an option in many countries and could be a solution when the cultivar is interesting for the other compounds. Regulations in Norway do not restrict sugar addition to apple juice for cider production.
If the acidity level is low, higher tannin content would be preferable to compensate for the low acidity by bitterness. Moreover, astringency is enhanced by acidity, so high tannin content/high acid may be rejected. That is probably the reason why most of the cider makers rarely make cider with both high acidity and high tannin content.
More important is the apples’ content of nitrogen and polyphenolic compounds, and the selection of yeast, for a proper fermentation process and development of the desired aromatic profile. Rosend et al. concluded from their study that apple variety was the primary factor influencing development of volatile components, while the effects of yeast and maturity were variety-specific [43]. As a result of the fermentation process, compounds like 3-methyl-1-butanol, 2 phenylethanol and ethyl acetate are formed, while content of 1-butanol depends more on cultivar [46]. Fermented apple juice with low nutrient status tends to develop more 2-phenylethanol (pleasant floral odour) during fermentation [35]. Dos Santos et al. found that fermentation of apple must with low nitrogen content showed sluggish fermentation, and less volatile compounds were formed [36].
The use of cultured yeast strains to produce a broader range of aromas during fermentation is at the experimental stage in beer production [47]. If addition of yeast is allowed according to local regulations for cider, this might also be an option in cider production, though selection of a proper yeast strain will have to be explored [43].
4. Conclusions
We found large variation in chemical composition between the cultivars, though no significant differences between old and new cultivars, nor between growing locations. This study was only from one season and shows the need for further identification of the most common cultivars to better understand the effect of conditions on juice composition.
Most of the apple cultivars in this experiment were in the lower level of sugar content. Addition of sugar might be a way to compensate for this.
Bramley Seedling was high in acidity and polyphenols and can characterized as a sharp apple cultivar. This cultivar can be used to balance an apple juice mixture. Summerred is used in small quantities for cider production in Norway today due to its fresh and acidic taste. Its nitrogen content was relatively high, and it was low in TP Folin and in flavanols. Filippa is an old cultivar known to be aromatic and acidic. Both Filippa and Torstein red were high in FAA and low in flavanols.
Author Contributions
T.W.: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Project administration, Resources, Visualization, Writing—original draft. J.-M.L.Q.: Data curation, Investigation, Methodology, Software, Supervision, Validation, Visualization, Writing—review & editing. S.G.: Funding acquisition, Methodology, Resources, Supervision Validation, Visualization, Writing—review & editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding. Apples for the experiment was donated by The Norwegian University of Life Sciences (NMBU) and the Norwegian Institute of Bioeconomy Research (NIBIO).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Research dates are stored in accordance with NMBU’s regulations.
Acknowledgments
This work was carried out at Unité de Recherches Cidricoles at INRA, Le Rheu, France (now converted into “Polyphenols, Reactivity and Process, PRP” team belonging to INRAe, UR1268 BIA Research unit), in cooperation with the Faculty of Chemistry, Biotechnology and Food Science at the Norwegian University of Life Sciences. Special thanks to Alain Baron, Agnes Gacel and Gildas Le Bail for practical assistance at the plant at INRAe, as well as with analytical expertise, and to Kari Olsen and May Aalberg at the Norwegian University for Life Sciences for their contribution to chemical analyses.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Alberti, A.; dos Santos, T.P.M.; Zielinski, A.A.F.; dos Santos, C.M.E.; Braga, C.M.; Demiate, I.M.; Nogueira, A. Impact on chemical profile in apple juice and cider made from unripe, ripe and senescent dessert varieties. Lwt Food Sci. Technol. 2016, 65, 436–443. [Google Scholar] [CrossRef]
- Guyot, S.; Le Bourvellec, C.; Marnet, N.; Drilleau, J. Procyanidins are the most abundant polyphenols in dessert apples at maturity. Food Sci. Technol. 2002, 35, 289–291. [Google Scholar] [CrossRef]
- Lea, A. Craft Cider Making; The Good Life Press Ltd.: Preston, UK, 2010. [Google Scholar]
- Ewing, B.L.; Peck, G.M.; Ma, S.; Neilson, A.P.; Stewart, A.C. Management of Apple Maturity and Postharvest Storage Conditions to Increase Polyphenols in Cider. HortScience 2019, 54, 143–148. [Google Scholar] [CrossRef] [Green Version]
- BBai, X.; Zhang, H.; Ren, S. Antioxidant activity and HPLC analysis of polyphenol-enriched extracts from industrial apple pomace. J. Sci. Food Agric. 2013, 93, 2502–2506. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Psomas, A.; Zovoili, A.; Siatis, V. Polyphenolic profile and antioxidant activity of five apple cultivars grown under organic and conventional agricultural practices. Int. J. Food Sci. Technol. 2009, 44, 1167–1175. [Google Scholar] [CrossRef]
- Tsao, R.; Yang, R.; Xie, S.; Sockovie, E.; Khanizadeh, S. Which Polyphenolic Compounds Contribute to the Total Antioxidant Activities of Apple? J. Agric. Food Chem. 2005, 53, 4989–4995. [Google Scholar] [CrossRef] [PubMed]
- Jakobek, L.; García-Villalba, R.; Tomás-Barberán, F.A. Polyphenolic characterisation of old local apple varieties from Southeastern European region. J. Food Compos. Anal. 2013, 31, 199–211. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Laskowski, P. Polyphenolic Compounds and Antioxidant Activity of New and Old Apple Varieties. J. Agric. Food Chem. 2008, 56, 6520–6530. [Google Scholar] [CrossRef] [PubMed]
- Kahle, K.; Kraus, M.; Richling, E. Polyphenol profiles of apple juices. Mol. Nutr. Food Res. 2005, 49, 797–806. [Google Scholar] [CrossRef]
- Cheng, L.L.; Raba, R. Accumulation of Macro- and Micronutrients and Nitrogen Demand-supply Relationship of ’Gala’/’Malling 26’ Apple Trees Grown in Sand Culture. J. Am. Soc. Hortic. Sci. 2009, 134, 3–13. [Google Scholar] [CrossRef]
- Karl, A.D.; Brown, M.G.; Ma, S.; Sandbrook, A.; Stewart, A.C.; Cheng, L.; Mansfield, A.K.; Peck, G.M. Soil Nitrogen Fertilization Increases Yeast Assimilable Nitrogen Concentrations in ‘Golden Russet’ and ‘Medaille d’Or’ Apples Used for Cider Production. Hortscience 2020, 55, 1345–1355. [Google Scholar] [CrossRef]
- Ma, S.; Neilson, A.P.; Lahne, J.; Peck, G.M.; O’Keefe, S.F.; Stewart, A.C. Free amino acid composition of apple juices with potential for cider making as determined by UPLC-PDA. J. Inst. Brew. 2018, 124, 467–476. [Google Scholar] [CrossRef]
- Boudreau, T.F.; Peck, G.M.; O’Keefe, S.F.; Stewart, A.C. Free amino nitrogen concentration correlates to total yeast assimilable nitrogen concentration in apple juice. Food Sci. Nutr. 2018, 6, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, T.F.; Peck, G.M.; Ma, S.; Patrick, N.; Duncan, S.; O’Keefe, S.F.; Stewart, A.C. Hydrogen sulphide production during cider fermentation is moderated by pre-fermentation methionine addition. J. Inst. Brew. 2017, 123, 553–561. [Google Scholar] [CrossRef]
- Di Maro, A.; Dosi, R.; Ferrara, L.; Rocco, M.; Sepe, J.; Ferrari, G.; Parente, A. Free amino acid profile in Malus domestica cv Annurca apples from the Campania region and other Italian vegetables. Aust. J. Crop Sci. 2011, 5, 154–161. [Google Scholar]
- Wu, J.; Gao, H.; Zhao, L.; Liao, X.; Chen, F.; Wang, Z.; Hu, X. Chemical compositional characterization of some apple cultivars. Food Chem. 2007, 103, 88–93. [Google Scholar] [CrossRef]
- Zuo, W.; Zhang, T.; Xu, H.; Wang, C.; Lu, M.; Chen, X.; Xu, L. Effect of fermentation time on nutritional components of red-fleshed apple cider. Food Bioprod. Process. 2019, 114, 276–285. [Google Scholar] [CrossRef]
- Ye, M.; Yue, T.; Yuan, Y. Changes in the profile of volatile compounds and amino acids during cider fermentation using dessert variety of apples. Eur. Food Res. Technol. 2014, 239, 67–77. [Google Scholar] [CrossRef]
- Villière, A.; Arvisenet, G.; Bauduin, R.; Le Quéré, J.-M.; Sérot, T. Influence of cider-making process parameters on the odourant volatile composition of hard ciders. J. Inst. Brew. 2015, 121, 95–105. [Google Scholar] [CrossRef]
- Vangdal, E.; Kvamm-Lichtenfeld, K. Ciders produced from Norwegian fresh consumption apple cultivars. Acta Hortic. 2018, 1205, 527–532. [Google Scholar] [CrossRef]
- Bradshaw, T.; Kingsley-Richards, S.; Foster, J. Apple cultivar evaluations for cider making in Vermont, USA. Acta Hortic. 2018, 1205, 453–460. [Google Scholar] [CrossRef]
- Nicolini, G.; Román, T.; Carlin, S.; Malacarne, M.; Nardin, T.; Bertoldi, D.; Larcher, R. Characterisation of single-variety still ciders produced with dessert apples in the Italian Alps. J. Inst. Brew. 2018, 124, 457–466. [Google Scholar] [CrossRef] [Green Version]
- Le Bourvellec, C.; Le Quéré, J.-M.; Sanoner, P.; Drilleau, J.-F.; Guyot, S. Inhibition of Apple Polyphenol Oxidase Activity by Procyanidins and Polyphenol Oxidation Products. J. Agric. Food Chem. 2004, 52, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Malec, M.; Le Quéré, J.-M.; Sotin, H.; Kolodziejczyk, K.; Bauduin, R.; Guyot, S. Polyphenol Profiling of a Red-Fleshed Apple Cultivar and Evaluation of the Color Extractability and Stability in the Juice. J. Agric. Food Chem. 2014, 62, 6944–6954. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.A.; Jones, G.P. Analysis of Proanthocyanidin Cleavage Products Following Acid-Catalysis in the Presence of Excess Phloroglucinol. J. Agric. Food Chem. 2001, 49, 1740–1746. [Google Scholar] [CrossRef]
- Moe, K.; Porcellato, D.; Skeie, S. Metabolism of milk fat globule membrane components by nonstarter lactic acid bacteria isolated from cheese. J. Dairy Sci. 2013, 96, 727–739. [Google Scholar] [CrossRef] [Green Version]
- Alexander, T.R.; King, J.; Zimmerman, A.; Miles, C.A. Regional Variation in Juice Quality Characteristics of Four Cider Apple (Malus xdomestica Borkh.) Cultivars in Northwest and Central Washington. Hortscience 2016, 51, 1498–1502. [Google Scholar] [CrossRef]
- Le Bourvellec, C.; Bureau, S.; Renard, C.M.G.C.; Plenet, D.; Gautier, H.; Touloumet, L.; Girard, T.; Simon, S. Cultivar and Year Rather than Agricultural Practices Affect Primary and Secondary Metabolites in Apple Fruit. PLoS ONE 2015, 10, e0141916. [Google Scholar] [CrossRef]
- Thompson-Witrick, K.A.; Goodrich, K.M.; Neilson, A.P.; Hurley, E.K.; Peck, G.M.; Stewart, A.C. Characterization of the Polyphenol Composition of 20 Cultivars of Cider, Processing, and Dessert Apples (Malus×domesticaBorkh.) Grown in Virginia. J. Agric. Food Chem. 2014, 62, 10181–10191. [Google Scholar] [CrossRef]
- Wrona, D. The influence of nitrogen fertilization on growth, yield and fruit size of ‘Jonagored’apple trees. Acta Sci. Pol. Hortorum Cultus 2011, 10, 3–10. [Google Scholar]
- Nava, G.; Dechen, A.R. Long-term annual fertilization with nitrogen and potassium affect yield and mineral composition of ‘fuji’ apple. Sci. Agric. 2009, 66, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Ernani, P.R.; Rogeri, D.A.; Proença, M.M.; Dias, J. Addition of nitrogen had no effect on yield and quality of apples in an high density orchard carrying a dwarf rootstock. Rev. Bras. Frutic. 2008, 30, 1113–1118. [Google Scholar] [CrossRef] [Green Version]
- Lea, A.G.H.; Drilleau, J.-F. Fermented Beverage Production; Kluwer Academic: New York, NY, USA, 2003. [Google Scholar]
- Eleutério dos Santos, C.M.; Pietrowski, G.D.A.M.; Braga, C.M.; Rossi, M.J.; Ninow, J.; Machado dos Santos, T.P.; Wosiacki, G.; Matos Jorge, R.M.; Nogueira, A. Apple Aminoacid Profile and Yeast Strains in the Formation of Fusel Alcohols and Esters in Cider Production. J. Food Sci. 2015, 80, C1170–C1177. [Google Scholar] [CrossRef]
- Jolicoeur, C. The New Cider Makers Handbook; Chelsea Green Publishing: White River Junction, Windsor, VT, USA, 2013. [Google Scholar]
- Guyot, S.; Marnet, N.; Sanoner, P.; Drilleau, J.-F. Variability of the Polyphenolic Composition of Cider Apple (Malus domestica) Fruits and Juices. J. Agric. Food Chem. 2003, 51, 6240–6247. [Google Scholar] [CrossRef] [PubMed]
- Francini, A.; Sebastiani, L. Phenolic Compounds in Apple (Malus x domestica Borkh.): Compounds Characterization and Stability during Postharvest and after Processing. Antioxidants 2013, 2, 181–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anastasiadi, M.; Mohareb, F.; Redfern, S.P.; Berry, M.; Simmonds, M.S.J.; Terry, L.A. Biochemical Profile of Heritage and Modern Apple Cultivars and Application of Machine Learning Methods To Predict Usage, Age, and Harvest Season. J. Agric. Food Chem. 2017, 65, 5339–5356. [Google Scholar] [CrossRef] [PubMed]
- Verdu, C.F.; Childebrand, N.; Marnet, N.; LeBail, G.; Dupuis, F.; Laurens, F.; Guilet, D.; Guyot, S. Polyphenol variability in the fruits and juices of a cider apple progeny. J. Sci. Food Agric. 2014, 94, 1305–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laaksonen, O.; Kuldjärv, R.; Paalme, T.; Virkki, M.; Yang, B. Impact of apple cultivar, ripening stage, fermentation type and yeast strain on phenolic composition of apple ciders. Food Chem. 2017, 233, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Rosend, J.; Kuldjärv, R.; Rosenvald, S.; Paalme, T. The effects of apple variety, ripening stage, and yeast strain on the volatile composition of apple cider. Heliyon 2019, 5, e01953. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, A.; Le Quéré, J.M.; Gestin, P.; Michel, A.; Wosiacki, G.; Drilleau, J.F. Slow Fermentation in French Cider Processing due to Partial Biomass Reduction. J. Inst. Brew. 2008, 114, 102–110. [Google Scholar] [CrossRef]
- Symoneaux, R.; Chollet, S.; Bauduin, R.; Le Quéré, J.; Baron, A. Impact of apple procyanidins on sensory perception in model cider (part 2): Degree of polymerization and interactions with the matrix components. Lwt Food Sci. Technol. 2014, 57, 28–34. [Google Scholar] [CrossRef]
- Fan, W.; Xu, Y.; Han, Y. Quantification of Volatile Compoundsin Chinese Ciders by Stir Bar Sorptive Extraction (SBSE) and Gas Chromatography—Mass Spectrometry (GC–MS). J. Inst. Brew. 2011, 117, 61–66. [Google Scholar] [CrossRef]
- Denby, C.M.; Li, R.A.; Vu, V.T.; Costello, Z.; Lin, W.; Chan, L.J.G.; Williams, J.; Donaldson, B.; Bamforth, C.W.; Petzold, C.J.; et al. Industrial brewing yeast engineered for the production of primary flavor determinants in hopped beer. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(a,b) PCA plot of the apple cultivars, (a) polyphenolic composition (RP18−HPLC analysis of polyphenols), (b) density, total nitrogen, TP Folin, FAA, and pH.
Figure 1.
(a,b) PCA plot of the apple cultivars, (a) polyphenolic composition (RP18−HPLC analysis of polyphenols), (b) density, total nitrogen, TP Folin, FAA, and pH.
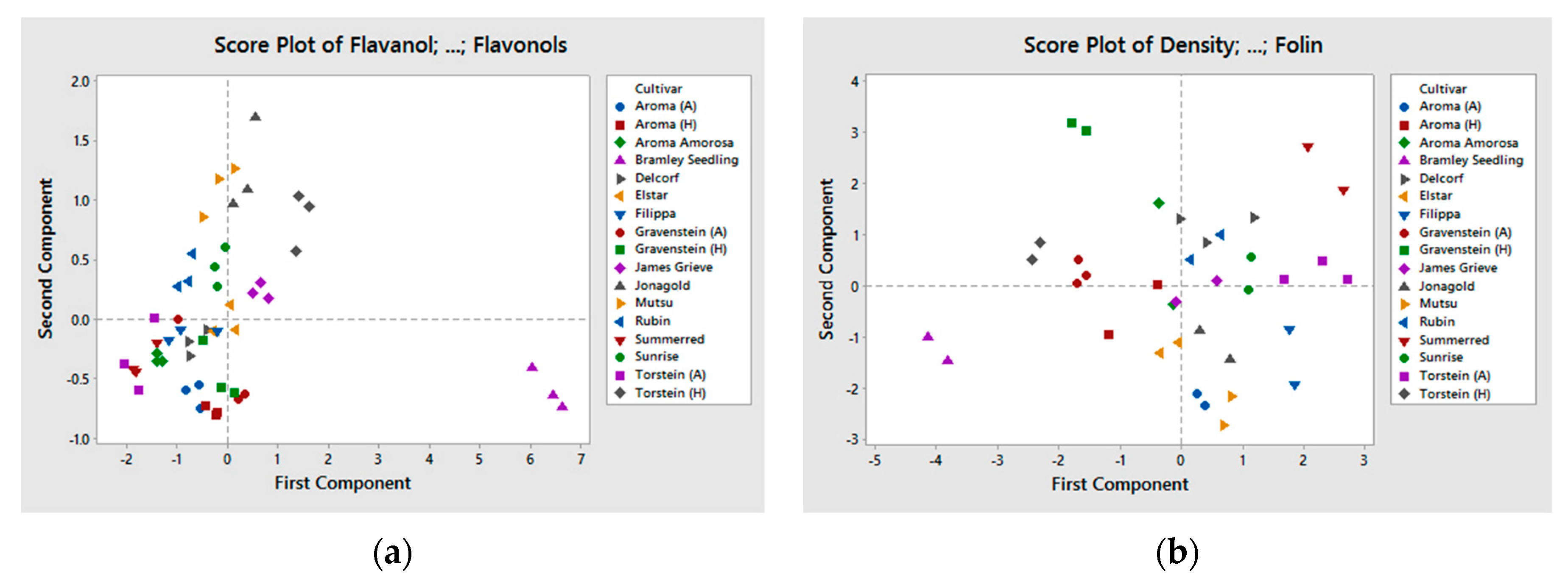

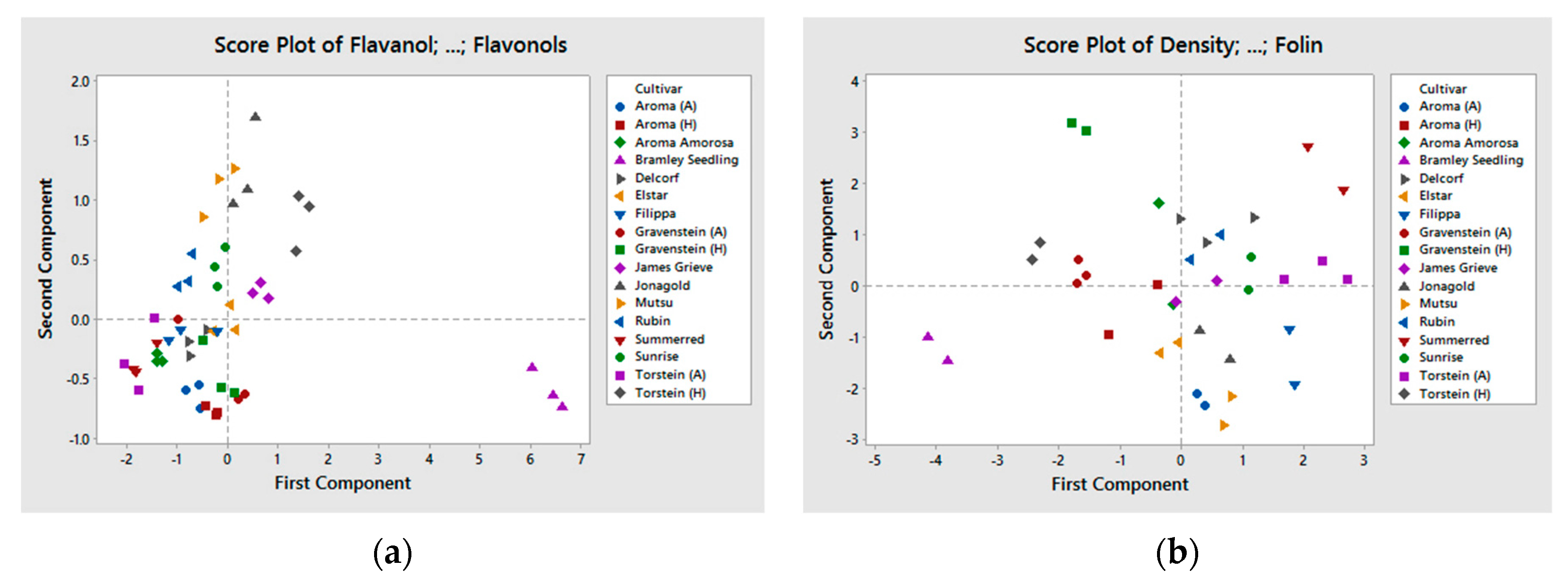{kind=link}
Table 1.
Apple cultivars: origin, parentage, age and growing location in Norway.
| Cultivar | Origin | Parentage | Age * | Growing Location (Norway) ** |
|---|---|---|---|---|
| Aroma Amorosa | Sweden | Ingrid Marie/Filippa | New | A |
| Aroma | Sweden 1973 | Ingrid Marie/Filippa | New | A + H |
| Bramley Seedling | England 1837 | Unknown | Old | H |
| Delcorf (Estivale) | France 1950 | Stark Jongrimes/Golden Delicious | New | A |
| Elstar | Holland 1972 | Golden Delicious/Ingrid Marie | Old | A |
| Filippa | Denmark from 1880 | Unknown | Old | A |
| Gravenstein | Denmark from 1669. in Norway from 1792 | Unknown | Old | A + H |
| James Grieve | Scotland 1893 | Pott’s Seedling | New | H |
| Jonagold | New York. US. 1943 | Jonathan/Golden Delicious | Old | A |
| Mutsu | Japan 1930 | Golden Delicious/Indo | Old | A |
| Rubin | Czech Republic 1960 | Golden Delicious/Lord Lambourne | New | A |
| Sunrise | Canada 1990 | McIntosh/Golden Delicious | New | A |
| Summerred | Canada 1964 | McIntosh/Golden Delicious | Old | A |
| Torstein red | Derived from Torstein | Old | A | |
| Torstein | Unknown (Norway 1760) | Unknown | Old | H |
* General knowledge about age. ** Growing location: A: Akershus/south-east (10°77′ E, 59°67′ N at the NMBU/Norwegian University of Life Sciences) H: Hardanger/west (6°66′ E, 60°32′ N at the Nibio Ullensvang Norwegian Institute of Bioeconomy Research).
Table 2.
Average fruit weight (whole apple, N = 5), density (N = 3) and pH (N = 2) in fresh apple juice.
Table 2.
Average fruit weight (whole apple, N = 5), density (N = 3) and pH (N = 2) in fresh apple juice.
| Cultivar | Fruit Weight | Density (Juice) | pH |
|---|---|---|---|
| g ± STD | g/L ± STD | ||
| Aroma Amorosa | 155 ± 4.2 d | 1053 ± 11.9 b,c,d,e,f | 3.2 ± 0.01 c |
| Aroma (A) | 178 ± 5.3 e | 1058 ± 3.3 e,f | 3.2 ± 0.01 c |
| Aroma (H) | 286 ± 9.3 h | 1051 ± 3.6 d,e | 3.2 ± 0.14 c,d |
| Bramley Seedling | 135 ± 5.5 c | 1046 ± 10.8 a,b,c,d,e | 2.9 ± 0.11 a |
| Delcorf | 235 ± 6.7 g | 1050 ± 0.6 d | 3.3 ± 0.01 d,e |
| Elstar | 119 ± 3.1 b | 1053 ± 1.2 e | 3.1 ± 0.04 b |
| Filippa | 200 ± 5.5 f | 1051 ± 4.0 c,d,e | 3.1 ± 0.08 b |
| Gravenstein (A) | 164 ± 6.2 d | 1050 ± 0.3 d | 3.3 ± 0.02 d |
| Gravenstein (H) | 115 ± 3.4 b | 1034 ± 0.7 a | 3.2 ± 0.02 c |
| James Grieve | 116 ± 3.0 b | 1046 ± 1.9 c | 3.2 ± 0.00 c |
| Jonagold | 181 ± 3.2 e | 1055 ± 3.5 e | 3.4 ± 0.14 d,e |
| Mutsu | 215 ± 8.0 f | 1060 ± 1.5 f | 3.3 ± 0.06 d,e |
| Rubin | 191 ± 4.5 e | 1053 ± 1.4 e | 3.3 ± 0.04 c,d |
| Sunrise | 162 ± 2.8 d | 1044 ± 1.2 b | 3.4 ± 0.04 e |
| Summerred | 124 ± 4.4 b | 1039 ± 2.1 b | 3.3 ± 0.05 c,d |
| Torstein red (A) | 123 ± 3.1 b | 1049 ± 0.7 d | 3.2 ± 0.07 b,c |
| Torstein (H) | 80 ± 2.6 a | 1047 ± 0.9 c,d | 3.2 ± 0.02 b,c |
| Average | 164 | 1049 | 3.2 |
STD: standard deviation. Different letters in the same column indicate significant difference (p < 0.05).
Table 3.
Content total nitrogen, sum of free amino acids (FAA) with % distribution, in fresh apple juice.
Table 3.
Content total nitrogen, sum of free amino acids (FAA) with % distribution, in fresh apple juice.
| Nitrogen | Sum FAA | ASN | ASP | GLU | SER | ALA | GABA | PHE | MET | |
|---|---|---|---|---|---|---|---|---|---|---|
| mg N/L ± STD | mg/kg ± STD | % | % | % | % | % | % | % | % | |
| Aroma Amorosa | 82.7 ± 6.3 b | 289.5 ± 0.2 d | 52.8 | 24.9 | 12.8 | 2.3 | 1.1 | 0.9 | 0.1 | 0.1 |
| Aroma (A) | 86.5 ± 0.4 b | 249.7 ± 0.2 c,d | 48.1 | 24.9 | 14.5 | 2.9 | 1.5 | 1.4 | 0.2 | 0.1 |
| Aroma (H) | 107.9 ± 23.8 b,c | 391.4 ± 0.8 d,e | 53.0 | 24.9 | 13.8 | 2.1 | 1.0 | 0.9 | 0.1 | 0.1 |
| Bramley Seedling | 74.9 ± 2.7 b | 253.4 ± 0.3 c,d | 37.5 | 31.4 | 20.8 | 1.9 | 1.2 | 1.6 | 0.2 | 0.1 |
| Delcorf | 135.8 ± 19.2 c,d | 455.3 ± 0.8 d,e | 49.7 | 13.8 | 6.1 | 9.5 | 10.8 | 1.6 | 0.1 | 0.2 |
| Elstar | 82.8 ± 7.7 b | 273.4 ± 0.4 d | 32.7 | 31.6 | 23.2 | 2.4 | 1.9 | 1.3 | 0.2 | ND |
| Filippa | 146.8 ± 1.0 d | 683.2 ± 0.5 e | 69.7 | 8.9 | 6.2 | 3.8 | 2.9 | 0.6 | 0.1 | 0.1 |
| Gravenstein (A) | 66.5 ± 3.8 a,b | 173.7 ± 0.2 b | 40.2 | 32.6 | 15.9 | 2.1 | 3.4 | 0.4 | 0.0 | ND |
| Gravenstein (H) | 77.2 ± 6.6 b | 213.6 ± 0.1 c | 47.7 | 25.5 | 16.0 | 1.7 | 2.2 | 0.7 | 0.1 | ND |
| James Grieve | 117.7 ± 10.6 c,d | 380.4 ± 0.4 d,e | 57.8 | 23.5 | 9.7 | 2.1 | 1.1 | 0.7 | 0.3 | 0.3 |
| Jonagold | 103.7 ± 4.1 b,c | 359.5 ± 0.4 d,e | 55.1 | 19.9 | 15.3 | 1.4 | 1.3 | 0.8 | 0.4 | 0.1 |
| Mutsu | 82.8 ± 5.6 b | 301.2 ± 0.4 d | 47.2 | 21.5 | 17.5 | 3.6 | 1.8 | 0.8 | 0.3 | 0.3 |
| Rubin | 125.8 ± 16.6 c,d | 446.2 ± 0.8 d,e | 50.7 | 18.6 | 13.7 | 7.0 | 2.0 | 1.4 | 0.1 | 0.4 |
| Sunrise | 131.0 ± 11.8 c,d | 459.9 ± 0.5 d,e | 40.7 | 37.2 | 8.5 | 2.0 | 3.5 | 2.6 | 0.0 | 0.1 |
| Summerred | 171.4 ± 26.5 d,e | 924.5 ± 0.8 f | 46.0 | 44.8 | 3.7 | 0.9 | 0.8 | 1.0 | 0.1 | 0.1 |
| Torstein red (A) | 139.8 ± 5.8 c,d | 613.9 ± 1.2 e | 64.6 | 16.9 | 9.3 | 2.3 | 2.1 | 0.8 | 0.1 | 0.2 |
| Torstein (H) | 43.2 ± 2.4 a | 42.4 ± 0.0 a | 15.9 | 36.3 | 28.0 | 4.5 | 1.9 | 0.8 | 0.0 | ND |
| Average | 105 | 382.4 | 47.6 | 25.7 | 13.8 | 3.1 | 2.4 | 1.1 | 0.1 | 0.1 |
ASN, asparagine; ASP, aspartic acid; GLU, glutamic acid; SER, serine; ALA, alanine; GABA, gamma-amino-butyric acid; PHE, phenylalanine; MET, methionine. STD, standard deviation. ND, not detected. Different letters in the same column indicate significant difference (p < 0.05).
Table 4.
Mean concentration of total polyphenols (Folin) mg/L EPI equiv. and polyphenols from RP18-HPLC analysis (mg/L) in fresh apple juice.
Table 4.
Mean concentration of total polyphenols (Folin) mg/L EPI equiv. and polyphenols from RP18-HPLC analysis (mg/L) in fresh apple juice.
| TP (Folin) | TotalPolyphenols (HPLC) | TotalFlavanols 1 | DPn 2 | TotalHydroxycin-namic Acids 3 | TotalDihydrochalcones 4 | TotalFlavonols 5 | |
|---|---|---|---|---|---|---|---|
| mg/L EPI equiv ± STD | mg/L | mg/L | mg/L | mg/L | mg/L | ||
| Aroma Amorosa | 598 ± 31 c | 209 b,c | 110 | 2.3 | 86 | 10 | 3 |
| Aroma (A) | 705 ± 18 c,d | 425 d | 227 | 1.9 | 180 | 15 | 3 |
| Aroma (H) | 966 ± 47 f | 587 e | 348 | 1.4 | 225 | 12 | 3 |
| Bramley Seedling | 1934 ± 42 g | 1838 f | 1042 | 3.0 | 765 | 27 | 4 |
| Delcorf | 666 ± 47 c,d | 397 c,d | 271 | 2.6 | 109 | 13 | 4 |
| Elstar | 696 ± 46 c,d | 383 d | 244 | 2.2 | 102 | 32 | 5 |
| Filippa | 730 ± 131 c,d | 290 b,c,d | 68 | 3.3 | 204 | 14 | 4 |
| Gravenstein (A) | 922 ± 40 f | 525 d,e | 345 | 2.6 | 157 | 20 | 3 |
| Gravenstein (H) | 855 ± 3 e | 468 d | 186 | 2.8 | 245 | 33 | 4 |
| James Grieve | 850 ± 12 e | 483 d,e | 302 | 2.1 | 136 | 38 | 7 |
| Jonagold | 796 ± 21 e | 370 d | 197 | 2.6 | 141 | 22 | 10 |
| Mutsu | 679 ± 24 c,d | 281 b,c | 113 | 2.8 | 144 | 15 | 8 |
| Rubin | 624 ± 26 c | 260 c,d | 172 | 2.4 | 70 | 12 | 5 |
| Sunrise | 658 ± 53 c,d | 371 d | 271 | 2.2 | 73 | 22 | 6 |
| Summerred | 272 ± 18 a | 87 a | 34 | 9.5 | 39 | 13 | 1 |
| Torstein red (A) | 453 ± 65 b | 87 a,b | 13 | 1.0 | 67 | 6 | 2 |
| Torstein (H) | 990 ± 37 f | 693 e | 354 | 1.9 | 299 | 29 | 10 |
| Average | 775 | 450 | 246 | 2.7 | 179 | 20 | 5 |
1 Total flavanols concentration calculated based on phloroglucinolysis reaction and HPLC analysis corresponds to the sum of catechins (monomers) and procyanidin oligomers and polymers. 2 DPn: average degree of polymerization of flavanols obtained from data of phloroglucinolysis coupled to RP18-HPLC analysis [26,27]. 3 Hydroxycinnamic acids: chlorogenic acid (i.e., caffeoylquinic acid, p-coumaroylquinic acid and caffeic acid). 4 Dihydrochalcones: phloridzin and phloretin xyluglucoside. 5 Flavonols: quercetin glycosides including rutin, avicularin, quecitrin, isoquercitrin, reynoutrin and hyperoside [26]. Different letters in the same column indicate significant difference (p < 0.05).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wicklund, T.; Guyot, S.; Le Quéré, J.-M. Chemical Composition of Apples Cultivated in Norway. Crops 2021, 1, 8-19. https://0-doi-org.brum.beds.ac.uk/10.3390/crops1010003
AMA Style
Wicklund T, Guyot S, Le Quéré J-M. Chemical Composition of Apples Cultivated in Norway. Crops. 2021; 1(1):8-19. https://0-doi-org.brum.beds.ac.uk/10.3390/crops1010003
Chicago/Turabian StyleWicklund, Trude, Sylvain Guyot, and Jean-Michel Le Quéré. 2021. "Chemical Composition of Apples Cultivated in Norway" Crops 1, no. 1: 8-19. https://0-doi-org.brum.beds.ac.uk/10.3390/crops1010003