
Biological Relevance of Free Radicals in the Process of Physiological Capacitation and Cryocapacitation



{kind=link}
Abstract
:1. Introduction
2. The Origin of ROS in Semen
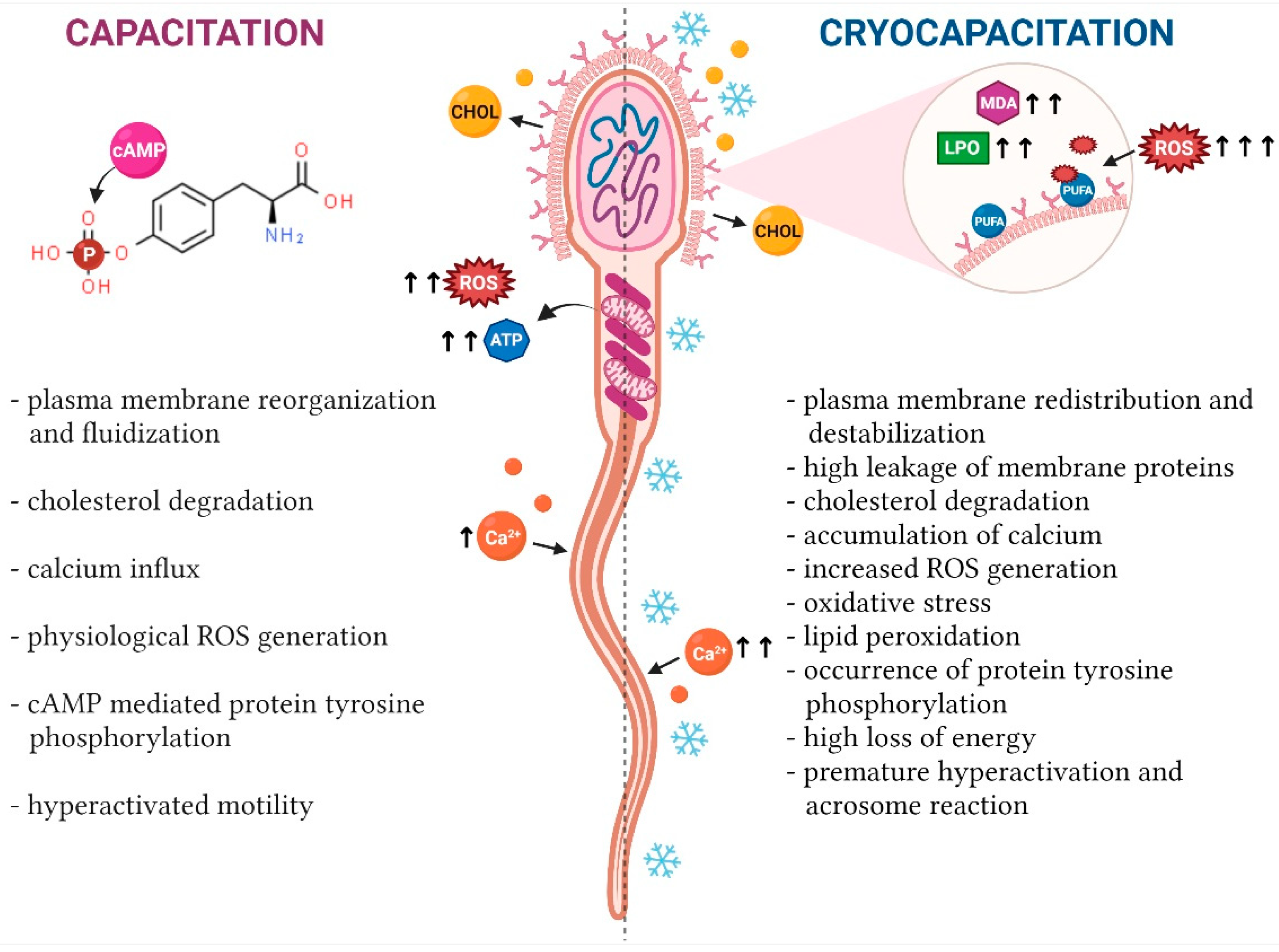
3. Relationship between Capacitation and ROS
4. Cryodamage and ROS Overproduction
5. Capacitation-Like Changes of Spermatozoa during Cryopreservation
6. Future Strategies against Cryodamage
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thompson, A.; Agarwal, A.; Du Plessis, S.S. Physiological role of reactive oxygen species in sperm function: A review. In Antioxidants in Male Infertility: A Guide for Clinicians and Researchers; Parekattil, S.J., Agarwal, A., Eds.; Springer Science + Business Media: New York, NY, USA, 2013; pp. 69–89. [Google Scholar]
- Gosalvez, J.; Tvrda, E.; Agarwal, A. Free radical and superoxide reactivity detection in semen quality assessment: Past, present, and future. J. Assist. Reprod. Genet. 2017, 34, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Majzoub, A.; Agarwal, A. Oxidative stress and sperm function: A systematic review on evaluation and management. Arab J. Urol. 2019, 17, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kothari, S.; Thompson, A.; Agarwal, A.; du Plessis, S.S. Free radicals: Their beneficial and detrimental effects on sperm function. Indian J. Exp. Biol. 2010, 48, 425–435. [Google Scholar]
- Tvrdá, E.; Massanyi, P.; Lukáč, N. Physiological and pathological roles of free radicals in male reproduction. In Spermatozoa-Facts and Perspectives; Meccariello, R., Chianese, R., Eds.; IntechOpen: London, UK, 2017. [Google Scholar]
- Kobayashi, C.I.; Suda, T. Regulation of reactive oxygen species in stem cells and cancer stem cells. J. Cell. Physiol. 2012, 227, 421–430. [Google Scholar] [CrossRef]
- Chen, H.; Zhao, H.X.; Huang, X.F.; Chen, G.W.; Yang, Z.X.; Sun, W.J.; Tao, M.H.; Yuan, Y.; Wu, J.Q.; Sun, F.; et al. Does high load of oxidants in human semen contribute to male factor infertility? Antioxid. Redox. Signal. 2012, 16, 754–759. [Google Scholar] [CrossRef]
- Aitken, R.J.; De Iuliis, G.N.; Finnie, J.M.; Hedges, A.; McLachlan, R.I. Analysis of the relationships between oxidative stress, DNA damage and sperm vitality in a patient population: Development of diagnostic criteria. Hum. Reprod. 2010, 25, 2415–2426. [Google Scholar] [CrossRef] [Green Version]
- Sabeti, P.; Pourmasumi, S.; Rahiminia, T.; Akyash, F.; Talebi, A.R. Etiologies of sperm oxidative stress. Int. J. Reprod. Biomed. 2016, 14, 231. [Google Scholar] [CrossRef]
- Lee, D.; Moawad, A.R.; Morielli, T.; Fernandez, M.C.; O’Flaherty, C. Peroxiredoxins prevent oxidative stress during human sperm capacitation. MHR: Basic Sci. Reprod. Med. 2017, 23, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Cormier, N.; Bailey, J.L. A differential mechanism is involved during heparin-and cryopreservation-induced capacitation of bovine spermatozoa. Biol. Reprod. 2003, 69, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, G.B.; Caldas-Bussiere, M.C.; Maciel, V.L.; Carvalho, C.S.P.D.; Souza, C.L.M.D. Association of L-arginine with heparin on the sperm capacitation improves in vitro embryo production in bovine. Anim. Reprod. 2019, 16, 938–944. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, C.; de Lamirande, E.; Gagnon, C. Positive role of reactive oxygen species in mammalian sperm capacitation: Triggering and modulation of phosphorylation events. Free Radic. Biol. Med. 2006, 41, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Bollwein, H.; Fuchs, I.; Koess, C. Interrelationship between plasma membrane integrity, mitochondrial membrane potential and DNA fragmentation in cryopreserved bovine spermatozoa. Reprod. Domest. Anim. 2008, 43, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.L.; Bilodeau, J.F.; Cormier, N. Semen cryopreservation in domestic animals: A damaging and capacitating phenomenon. J. Androl. 2000, 21, 1–7. [Google Scholar]
- Sharma, V. Sperm storage for cancer patients in the UK: A review of current practice. Hum. Reprod. 2011, 26, 2935–2943. [Google Scholar] [CrossRef]
- O’Connell, M.; Mcclure, N.; Lewis, S.E.M. The effects of cryopreservation on sperm morphology, motility and mitochondrial function. Hum. Reprod. 2002, 17, 704–709. [Google Scholar] [CrossRef]
- Srivastava, N.; Srivastava, S.K.; Ghosh, S.K.; Kumar, A.; Perumal, P.; Jerome, A. Acrosome membrane integrity and cryocapacitation are related to cholesterol content of bull spermatozoa. Asian Pac. J. Reprod. 2013, 2, 126–131. [Google Scholar] [CrossRef]
- Li, X.; Ni, M.; Xing, S.; Yu, Y.; Zhou, Y.; Yang, S.; Li, H.; Zhu, R.; Han, M. Reactive Oxygen Species Secreted by Leukocytes in Semen Induce Self-Expression of Interleukin-6 and Affect Sperm Quality. Am. J. Men’s Health 2020, 14, 1557988320970053. [Google Scholar] [CrossRef]
- Agarwal, A.; Makker, K.; Sharma, R. Clinical relevance of oxidative stress in male factor infertility: An update. Am. J. Reprod. Immunol. 2008, 59, 2–11. [Google Scholar] [CrossRef]
- Jung, J.H.; Kim, M.H.; Kim, J.; Baik, S.K.; Koh, S.B.; Park, H.J.; Seo, J.T. Treatment of leukocytospermia in male infertility: A systematic review. World J. Men’s Health 2016, 34, 165–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Plessis, S.S.; Agarwal, A.; Halabi, J.; Tvrda, E. Contemporary evidence on the physiological role of reactive oxygen species in human sperm function. J. Assist. Reprod. Genet. 2015, 32, 509–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, T.G. The epididymis, cytoplasmic droplets and male fertility. Asian J. Androl. 2011, 13, 130–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, N.; Saleh, R.A.; Sharma, R.K.; Lewis-Jones, I.; Esfandiari, N.; Thomas, A.J., Jr.; Agarwal, A. Novel association between sperm reactive oxygen species production, sperm morphological defects, and the sperm deformity index. Fertil. Steril. 2004, 81, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Zini, A.; Defreitas, G.; Freeman, M.; Hechter, S.; Jarvi, K. Varicocele is associated with abnormal retention of cytoplasmic droplets by human spermatozoa. Fertil. Steril. 2000, 74, 461–464. [Google Scholar] [CrossRef]
- Moein, M.R.; Soleimani, M.; Tabibnejad, M.D. Reactive oxygen species (ROS) production in seminal fluid correlate with the severity of varicocele in infertile men. Iran. J. Reprod. Med. 2008, 6, 65–69. [Google Scholar]
- Park, J.; Rho, H.K.; Kim, K.H.; Choe, S.S.; Lee, Y.S.; Kim, J.B. Overexpression of glucose-6-phosphate dehydrogenase is associated with lipid dysregulation and insulin resistance in obesity. Mol. Cell. Biol. 2005, 25, 5146–5157. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, C.; Obert, G.; Deffosez, A.; Formstecher, P.; Marchetti, P. Study of mitochondrial membrane potential, reactive oxygen species, DNA fragmentation and cell viability by flow cytometry in human sperm. Hum. Reprod. 2002, 17, 1257–1265. [Google Scholar] [CrossRef] [Green Version]
- Amaral, A.; Lourenço, B.; Marques, M.; Ramalho-Santos, J. Mitochondria functionality and sperm quality. Reproduction 2013, 146, R163–R174. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Fang, P.; Mai, J.; Choi, E.T.; Wang, H.; Yang, X.F. Targeting mitochondrial reactive oxygen species as novel therapy for inflammatory diseases and cancers. J. Hematol. Oncol. 2013, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Cadenas, E.; Davies, K.J. Mitochondrial free radical generation, oxidative stress, and aging. Free Radic. Biol. Med. 2000, 29, 222–230. [Google Scholar] [CrossRef]
- Chianese, R.; Pierantoni, R. Mitochondrial Reactive Oxygen Species (ROS) Production Alters Sperm Quality. Antioxidants 2021, 10, 92. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Whiting, S.; De Iuliis, G.N.; McClymont, S.; Mitchell, L.A.; Baker, M.A. Electrophilic aldehydes generated by sperm metabolism activate mitochondrial reactive oxygen species generation and apoptosis by targeting succinate dehydrogenase. J. Biol. Chem. 2012, 287, 33048–33060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koppers, A.J.; Garg, M.L.; Aitken, R.J. Stimulation of mitochondrial reactive oxygen species production by unesterified, unsaturated fatty acids in defective human spermatozoa. Free Radic. Biol. Med. 2010, 48, 112–119. [Google Scholar] [CrossRef]
- Chang, M.C. Fertilizing capacity of spermatozoa deposited into the fallopian tubes. Nature 1951, 168, 697–698. [Google Scholar] [CrossRef]
- Austin, C.R. Observations on the penetration of the sperm into the mammalian egg. Aust. J. Biol. Sci. 1951, 4, 581–596. [Google Scholar] [CrossRef] [Green Version]
- Visconti, P.E. Understanding the molecular basis of sperm capacitation through kinase design. Proc. Natl. Acad. Sci. USA 2009, 106, 667–668. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.K.; Yang, W.X. Factors and pathways involved in capacitation: How are they regulated? Oncotarget 2017, 8, 3600. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef]
- O’Flaherty, C. Redox regulation of mammalian sperm capacitation. Asian J. Androl. 2015, 17, 583–590. [Google Scholar] [CrossRef]
- de Lamirande, E.; Gagnon, C. Capacitation-associated production of superoxide anion by human spermatozoa. Free Radic. Biol. Med. 1995, 18, 487–495. [Google Scholar] [CrossRef]
- Papas, M.; Catalan, J.; Barranco, I.; Arroyo, L.; Bassols, A.; Yeste, M.; Miró, J. Total and specific activities of superoxide dismutase (SOD) in seminal plasma are related with the cryotolerance of jackass spermatozoa. Cryobiology 2020, 92, 109–116. [Google Scholar] [CrossRef] [PubMed]
- de Lamirande, E.; Harakat, A.; Gagnon, C. Human sperm capacitation induced by biological fluids and progesterone, but not by NADH or NADPH, is associated with the production of superoxide anion. J. Androl. 1998, 19, 215–225. [Google Scholar]
- Roy, S.C.; Atreja, S.K. Effect of reactive oxygen species on capacitation and associated protein tyrosine phosphorylation in buffalo (Bubalus bubalis) spermatozoa. Anim. Reprod. Sci. 2008, 107, 68–84. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.B.; Chatterjee, S.; Lefièvre, L.; de Lamirande, E.; Gagnon, C. Nitric oxide interacts with the cAMP pathway to modulate capacitation of human spermatozoa. Free Radic. Biol. Med. 2000, 29, 522–536. [Google Scholar] [CrossRef]
- O’Flaherty, C.; Rodriguez, P.; Srivastava, S. L-arginine promotes capacitation and acrosome reaction in cryopreserved bovine spermatozoa. Biochim. Biophys. Acta—Gen. Subj. 2004, 1674, 215–221. [Google Scholar] [CrossRef]
- Rodriguez, P.C.; O’flaherty, C.M.; Beconi, M.T.; Beorlegui, N.B. Nitric oxide-induced capacitation of cryopreserved bull spermatozoa and assessment of participating regulatory pathways. Anim. Reprod. Sci. 2005, 85, 231–242. [Google Scholar] [CrossRef]
- Dutta, S.; Henkel, R.; Sengupta, P.; Agarwal, A. Physiological role of ROS in sperm function. In Male Infertility; Parekattil, S.J., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 337–345. [Google Scholar]
- Battistone, M.A.; Da Ros, V.G.; Salicioni, A.M.; Navarrete, F.A.; Krapf, D.; Visconti, P.E.; Cuasnicu, P.S. Functional human sperm capacitation requires both bicarbonate-dependent PKA activation and down-regulation of Ser/Thr phosphatases by Src family kinases. MHR: Basic Sci. Reprod. Med. 2013, 19, 570–580. [Google Scholar] [CrossRef] [Green Version]
- Ghanbari, H.; Keshtgar, S.; Kazeroni, M. Inhibition of the CatSper channel and NOX5 enzyme activity affects the functions of the progesterone-stimulated human sperm. Iran. J. Med. Sci. 2018, 43, 18. [Google Scholar]
- Miguel-Jiménez, S.; Pina-Beltrán, B.; Gimeno-Martos, S.; Carvajal-Serna, M.; Casao, A.; Pérez-Pe, R. NADPH Oxidase 5 and Melatonin: Involvement in Ram Sperm Capacitation. Front. Cell Dev. Biol. 2021, 9, 655794. [Google Scholar] [CrossRef]
- Moreno-Irusta, A.; Dominguez, E.M.; Marín-Briggiler, C.I.; Matamoros-Volante, A.; Lucchesi, O.; Tomes, C.N.; Trevino, C.L.; Buffone, M.G.; Lascano, R.; Losinno, O.; et al. Reactive oxygen species are involved in the signaling of equine sperm chemotaxis. Reproduction 2020, 159, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.; Aitken, R.J. A redox-regulated tyrosine phosphorylation cascade in rat spermatozoa. J. Androl. 2001, 22, 611–622. [Google Scholar] [PubMed]
- O’Flaherty, C.; Beorlegui, N.; Beconi, M.T. Participation of superoxide anion in the capacitation of cryopreserved bovine sperm. Int. J. Androl. 2003, 26, 109–114. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, C.M.; Beorlegui, N.B.; Beconi, M.T. Lactate dehydrogenase-C4 is involved in heparin-and NADH-dependent bovine sperm capacitation. Andrologia 2002, 34, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.; Goldberg, E. Inhibition of lactate dehydrogenase C4 (LDH-C4) blocks capacitation of mouse sperm in vitro. Cytogenet. Genome Res. 2003, 103, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Lefièvre, L.; Chen, Y.; Conner, S.J.; Scott, J.L.; Publicover, S.J.; Ford, W.C.L.; Barratt, C.L. Human spermatozoa contain multiple targets for protein S-nitrosylation: An alternative mechanism of the modulation of sperm function by nitric oxide? Proteomics 2007, 7, 3066–3084. [Google Scholar] [CrossRef] [Green Version]
- Visconti, P.E.; Westbrook, V.A.; Chertihin, O.; Demarco, I.; Sleight, S.; Diekman, A.B. Novel signaling pathways involved in sperm acquisition of fertilizing capacity. J. Reprod. Immunol. 2002, 53, 133–150. [Google Scholar] [CrossRef]
- Staicu, F.D.; Martínez-Soto, J.C.; Canovas, S.; Matás, C. Nitric oxide-targeted protein phosphorylation during human sperm capacitation. Sci. Rep. 2021, 11, 20979. [Google Scholar] [CrossRef]
- Miraglia, E.; De Angelis, F.; Gazzano, E.; Hassanpour, H.; Bertagna, A.; Aldieri, E.; Revelli, A.; Ghigo, D. Nitric oxide stimulates human sperm motility via activation of the cyclic GMP/protein kinase G signaling pathway. Reproduction 2011, 141, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Mostek, A.; Janta, A.; Majewska, A.; Ciereszko, A. Bull Sperm Capacitation Is Accompanied by Redox Modifications of Proteins. Int. J. Mol. Sci. 2021, 22, 7903. [Google Scholar] [CrossRef]
- Duan, J.; Gaffrey, M.J.; Qian, W.J. Quantitative proteomic characterization of redox-dependent post-translational modifications on protein cysteines. Mol. Biosyst. 2017, 13, 816–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.J.; Kwon, W.S.; Rahman, M.S.; Lee, J.S.; Pang, M.G. A novel approach to identifying physical markers of cryo-damage in bull spermatozoa. PLoS ONE 2015, 10, e0126232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prien, S.; Iacovides, S. Cryoprotectants & cryopreservation of equine semen: A review of industry cryoprotectants and the effects of cryopreservation on equine semen membranes. J. Dairy Vet. Anim. Res. 2016, 3, 1–8. [Google Scholar]
- Benson, J.D.; Woods, E.J.; Walters, E.M.; Critser, J.K. The cryobiology of spermatozoa. Theriogenology 2012, 78, 1682–1699. [Google Scholar] [CrossRef] [PubMed]
- Tatone, C.; Di Emidio, G.; Vento, M.; Ciriminna, R.; Artini, P.G. Cryopreservation and oxidative stress in reproductive cells. Gynecol. Endocrinol. 2010, 26, 563–567. [Google Scholar] [CrossRef]
- Su, L.J.; Zhang, J.H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.Y. Reactive oxygen species-induced lipid peroxidation in apoptosis, autophagy, and ferroptosis. Oxid. Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef] [Green Version]
- Sanocka, D.; Kurpisz, M. Reactive oxygen species and sperm cells. Reprod. Biol. Endocrinol. 2004, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Brouwers, J.F.; Gadella, B.M. In situ detection and localization of lipid peroxidation in individual bovine sperm cells. Free Radic. Biol. Med. 2003, 35, 1382–1391. [Google Scholar] [CrossRef]
- Dietrich, M.A.; Irnazarow, I.; Ciereszko, A. Proteomic identification of seminal plasma proteins related to the freezability of carp semen. J. Proteom. 2017, 162, 52–61. [Google Scholar] [CrossRef]
- Li, P.; Hulak, M.; Koubek, P.; Sulc, M.; Dzyuba, B.; Boryshpolets, S.; Rodina, M.; Gela, D.; Manaskova-Postlerova, P.; Peknicova, J.; et al. Ice-age endurance: The effects of cryopreservation on proteins of sperm of common carp, Cyprinus carpio L. Theriogenology 2010, 74, 413–423. [Google Scholar] [CrossRef]
- Hill, W.G.; Kaetzel, M.A.; Kishore, B.K.; Dedman, J.R.; Zeidel, M.L. Annexin A4 reduces water and proton permeability of model membranes but does not alter aquaporin 2–mediated water transport in isolated endosomes. J. Gen. Physiol. 2003, 121, 413–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, M.; Niksirat, H.; Shaliutina-Kolešová, A.; Siddique, M.A.M.; Sterba, J.; Boryshpolets, S.; Linhart, O. Molecular and subcellular cryoinjury of fish spermatozoa and approaches to improve cryopreservation. Rev. Aquac. 2020, 12, 909–924. [Google Scholar] [CrossRef]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and consequences of oxidative stress in spermatozoa. Reprod. Fertil. Dev. 2016, 28, 1–10. [Google Scholar] [CrossRef]
- Aitken, R.J. Free radicals, lipid peroxidation and sperm function. Reprod. Fertil. Dev. 1995, 7, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, D.J.; Ahmed, K.; Talukdar, P. Cryocapacitation and fertility of cryopreserved semen. Int. J. Livest. Res. 2015, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Kadirvel, G.; Kathiravan, P.; Kumar, S. Protein tyrosine phosphorylation and zona binding ability of in vitro capacitated and cryopreserved buffalo spermatozoa. Theriogenology 2011, 75, 1630–1639. [Google Scholar] [CrossRef]
- Thomas, A.D.; Meyers, S.A.; Ball, B.A. Capacitation-like changes in equine spermatozoa following cryopreservation. Theriogenology 2006, 65, 1531–1550. [Google Scholar] [CrossRef]
- Gadella, B.M.; Harrison, R.A.P. Capacitation induces cyclic adenosine 3′, 5′-monophosphate-dependent, but apoptosis-unrelated, exposure of aminophospholipids at the apical head plasma membrane of boar sperm cells. Biol. Reprod. 2002, 67, 340–350. [Google Scholar] [CrossRef] [Green Version]
- Cross, N.L. Decrease in order of human sperm lipids during capacitation. Biol. Reprod. 2003, 69, 529–534. [Google Scholar] [CrossRef] [Green Version]
- Bailey, J.L.; Berube, B. Mechanisms of cryopreservation-induced capacitation of bovine sperm. J. Androl. 1998, 20, 13. [Google Scholar]
- Galantino-Homer, H.L.; Visconti, P.E.; Kopf, G.S. Regulation of protein tyrosine phosphorylation during bovine sperm capacitation by a cyclic adenosine 3’, 5’-monophosphate-dependent pathway. Biol. Reprod. 1997, 56, 707–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, P.F. The causes of reduced fertility with cryopreserved semen. Anim. Reprod. Sci. 2000, 60–61, 481–492. [Google Scholar] [CrossRef]
- Hezavehei, M.; Sharafi, M.; Kouchesfahani, H.M.; Henkel, R.; Agarwal, A.; Esmaeili, V.; Shahverdi, A. Sperm cryopreservation: A review on current molecular cryobiology and advanced approaches. Reprod. Biomed. Online 2018, 37, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Gautier, C.; Aurich, C. “Fine feathers make fine birds”–The mammalian sperm plasma membrane lipid composition and effects on assisted reproduction. Anim. Reprod. Sci. 2021, 106884. [Google Scholar] [CrossRef]
- Aboagla, E.M.E.; Terada, T. Effects of egg yolk during the freezing step of cryopreservation on the viability of goat spermatozoa. Theriogenology 2004, 62, 1160–1172. [Google Scholar] [CrossRef]
- Jang, T.H.; Park, S.C.; Yang, J.H.; Kim, J.Y.; Seok, J.H.; Park, U.S.; Choi, C.W.; Lee, S.R.; Han, J. Cryopreservation and its clinical applications. Integr. Med. Res. 2017, 6, 12–18. [Google Scholar] [CrossRef]
- Cilio, S.; Rienzo, M.; Villano, G.; Mirto, B.F.; Giampaglia, G.; Capone, F.; Ferretti, G.; Di Zazzo, E.; Crocetto, F. Beneficial Effects of Antioxidants in Male Infertility Management: A Narrative Review. Oxygen 2022, 2, 1–11. [Google Scholar] [CrossRef]
- Papas, M.; Catalán, J.; Fernandez-Fuertes, B.; Arroyo, L.; Bassols, A.; Miró, J.; Yeste, M. Specific activity of superoxide dismutase in stallion seminal plasma is related to sperm cryotolerance. Antioxidants 2019, 8, 539. [Google Scholar] [CrossRef] [Green Version]
- Ener, K.; Aldemir, M.; Işık, E.; Okulu, E.M.R.A.H.; Özcan, M.F.; Uğurlu, M.; Tangal, A.; Özayar, A. The impact of vitamin E supplementation on semen parameters and pregnancy rates after varicocelectomy: A randomised controlled study. Andrologia 2016, 48, 829–834. [Google Scholar] [CrossRef]
- Cyrus, A.; Kabir, A.; Goodarzi, D.; Moghimi, M. The effect of adjuvant vitamin C after varicocele surgery on sperm quality and quantity in infertile men: A double blind placebo controlled clinical trial. Int. Braz. J. Urol. 2015, 41, 230–238. [Google Scholar] [CrossRef] [Green Version]
- Hosseinabadi, F.; Jenabi, M.; Ghafarizadeh, A.A.; Yazdanikhah, S. The effect of vitamin B12 supplement on post-thaw motility, viability and DNA damage of human sperm. Andrologia 2020, 52, e13877. [Google Scholar] [CrossRef] [PubMed]
- Payne, S.R.; Oliver, J.E.; Upreti, G.C. Effect of antifreeze proteins on the motility of ram spermatozoa. Cryobiology 1994, 31, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Nishijima, K.; Tanaka, M.; Sakai, Y.; Koshimoto, C.; Morimoto, M.; Watanabe, T.; Fan, J.; Kitajima, S. Effects of type III antifreeze protein on sperm and embryo cryopreservation in rabbit. Cryobiology 2014, 69, 22–25. [Google Scholar] [CrossRef]
- Qadeer, S.; Khan, M.A.; Ansari, M.S.; Rakha, B.A.; Ejaz, R.; Iqbal, R.; Younis, M.; Ullah, N.; DeVries, A.L.; Akhter, S. Efficiency of antifreeze glycoproteins for cryopreservation of Nili-Ravi (Bubalus bubalis) buffalo bull sperm. Anim. Reprod. Sci. 2015, 157, 56–62. [Google Scholar] [CrossRef]
- Younis, A.I.; Rooks, B.; Khan, S.; Gould, K.G. The effects of antifreeze peptide III (AFP) and insulin transferrin selenium (ITS) on cryopreservation of chimpanzee (Pan troglodytes) spermatozoa. J. Androl. 1998, 19, 207–214. [Google Scholar] [PubMed]
- Zilli, L.; Beirão, J.; Schiavone, R.; Herraez, M.P.; Gnoni, A.; Vilella, S. Comparative proteome analysis of cryopreserved flagella and head plasma membrane proteins from sea bream spermatozoa: Effect of antifreeze proteins. PLoS ONE 2014, 9, e99992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamzalıoğlu, A.; Gökmen, V. Interaction between bioactive carbonyl compounds and asparagine and impact on acrylamide. In Acrylamide in Food: Analysis, Content and Potential Health Effects; Gökmen, V., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 355–376. [Google Scholar]
- Tvrdá, E.; Greifová, H.; Mackovich, A.; Hashim, F.; Lukáč, N. Curcumin offers antioxidant protection to cryopreserved bovine semen. Czech J. Anim. Sci. 2018, 63, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Tvrda, E.; Straka, P.; Galbavy, D.; Ivanic, P. Epicatechin provides antioxidant protection to bovine spermatozoa subjected to induced oxidative stress. Molecules 2019, 24, 3226. [Google Scholar] [CrossRef] [Green Version]
- Tvrda, E.; Mackovich, A.; Greifova, H.; Hashim, F.; Lukac, N. Antioxidant effects of lycopene on bovine sperm survival and oxidative profile following cryopreservation. Vet. Med. 2017, 62, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Balao da Silva, C.M.; Macías-García, B.; Miró-Morán, A.; González-Fernández, L.; Morillo-Rodriguez, A.; Ortega-Ferrusola, C.; Gallardo-Bolaños, J.M.; Stilwell, G.; Tapia, J.A.; Peña, F.J. Melatonin reduces lipid peroxidation and apoptotic-like changes in stallion spermatozoa. J. Pineal Res. 2011, 51, 172–179. [Google Scholar] [CrossRef]
- Carvajal-Serna, M.; Cardozo-Cerquera, J.A.; Grajales-Lombana, H.A.; Casao, A.; Pérez-Pe, R. Sperm Behavior and Response to Melatonin under Capacitating Conditions in Three Sheep Breeds Subject to the Equatorial Photoperiod. Animals 2021, 11, 1828. [Google Scholar] [CrossRef] [PubMed]
- Gimeno-Martos, S.; Casao, A.; Yeste, M.; Cebrián-Pérez, J.A.; Muiño-Blanco, T.; Pérez-Pé, R. Melatonin reduces cAMP-stimulated capacitation of ram spermatozoa. Reprod. Fertil. Dev. 2019, 31, 420–431. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benko, F.; Ďuračka, M.; Baňas, Š.; Lukáč, N.; Tvrdá, E. Biological Relevance of Free Radicals in the Process of Physiological Capacitation and Cryocapacitation. Oxygen 2022, 2, 164-176. https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen2020014
Benko F, Ďuračka M, Baňas Š, Lukáč N, Tvrdá E. Biological Relevance of Free Radicals in the Process of Physiological Capacitation and Cryocapacitation. Oxygen. 2022; 2(2):164-176. https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen2020014
Chicago/Turabian StyleBenko, Filip, Michal Ďuračka, Štefan Baňas, Norbert Lukáč, and Eva Tvrdá. 2022. "Biological Relevance of Free Radicals in the Process of Physiological Capacitation and Cryocapacitation" Oxygen 2, no. 2: 164-176. https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen2020014