
Cell-Cycle Synchronization Prior to Radiotherapy: A Mathematical Model of the Use of Gemcitabine on Melanoma Xenografts

Abstract
:1. Introduction
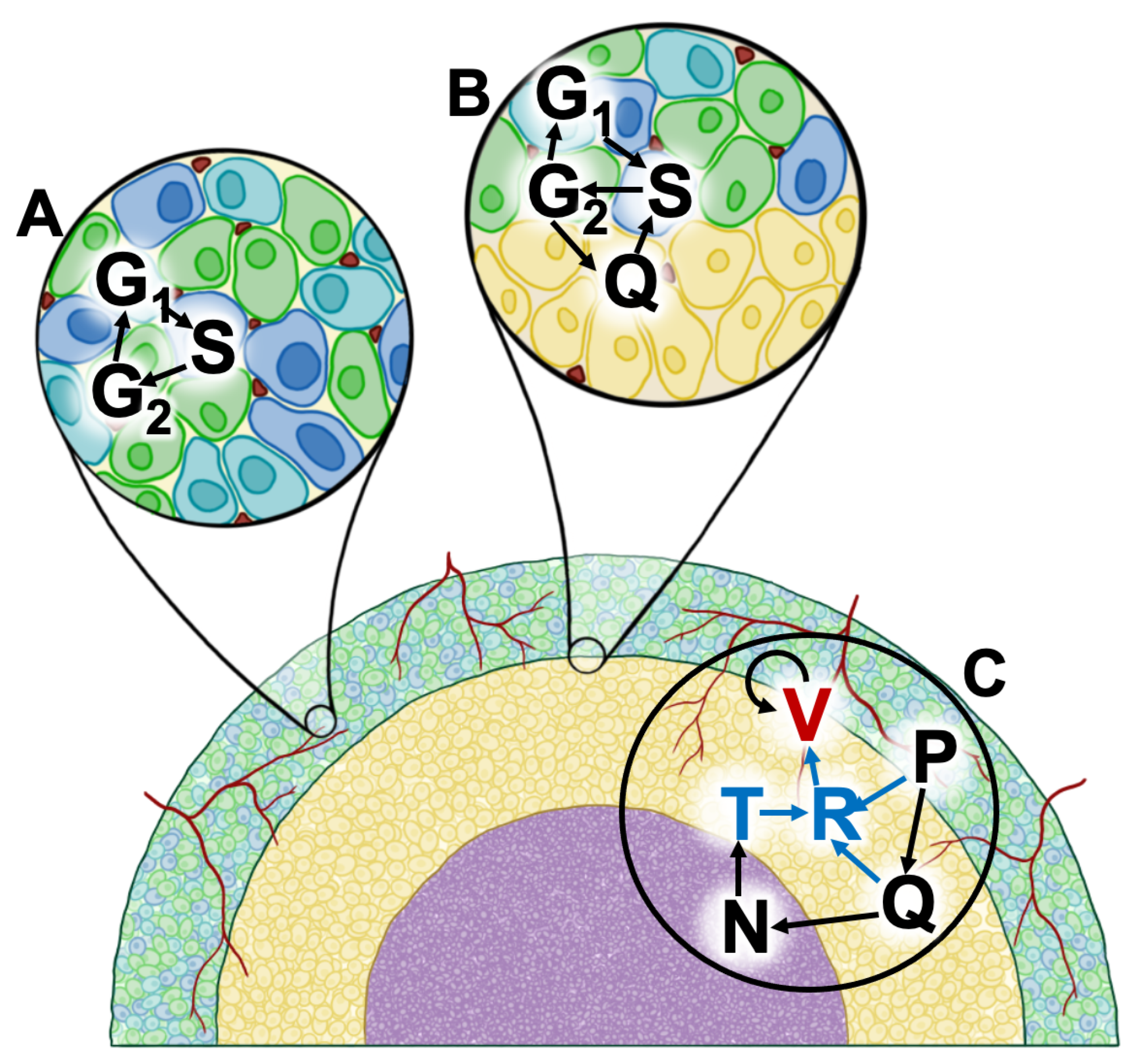
2. Methods
2.1. Monolayer Parameters
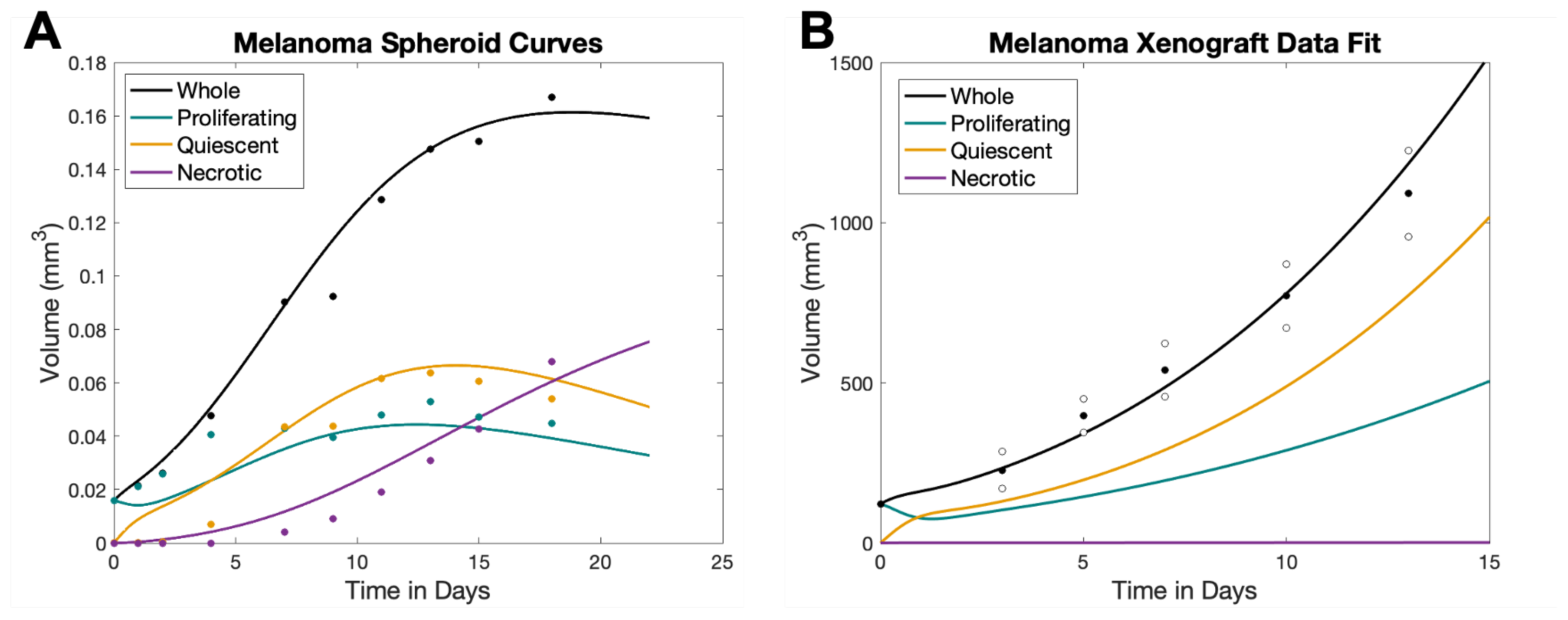
2.2. Spheroid Parameters
2.3. Xenograft Parameters
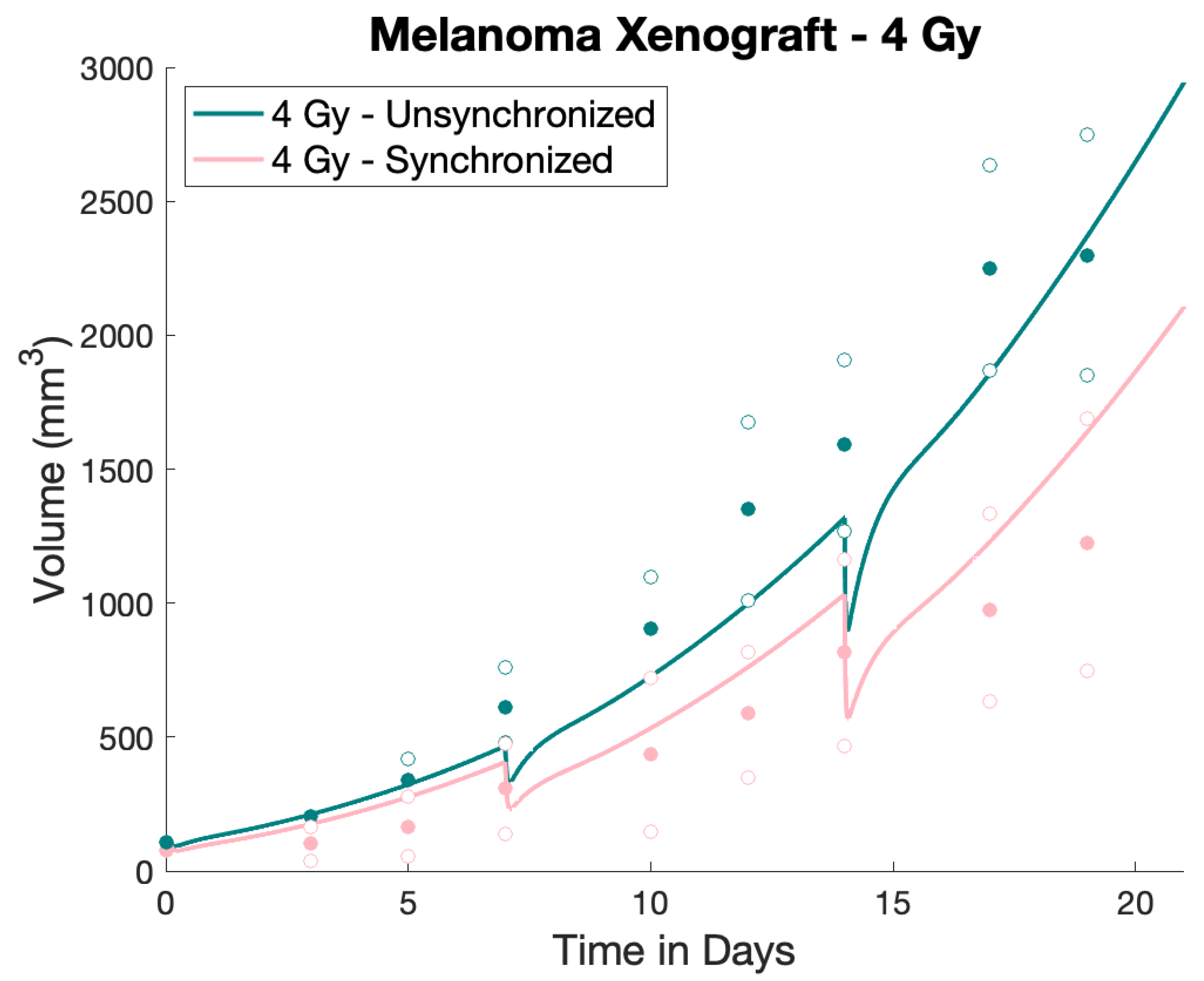
2.4. Numerical Treatment Simulations
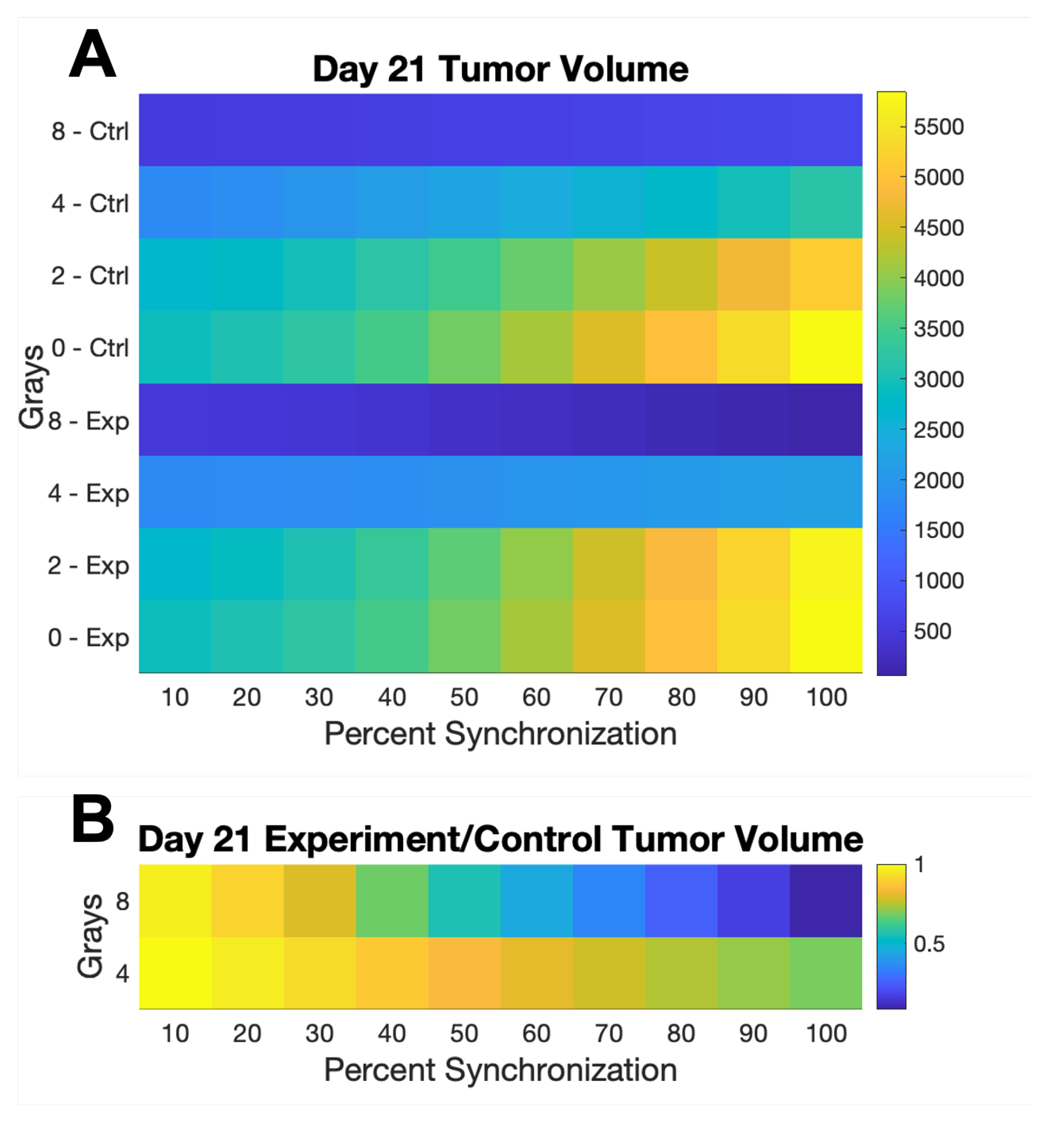
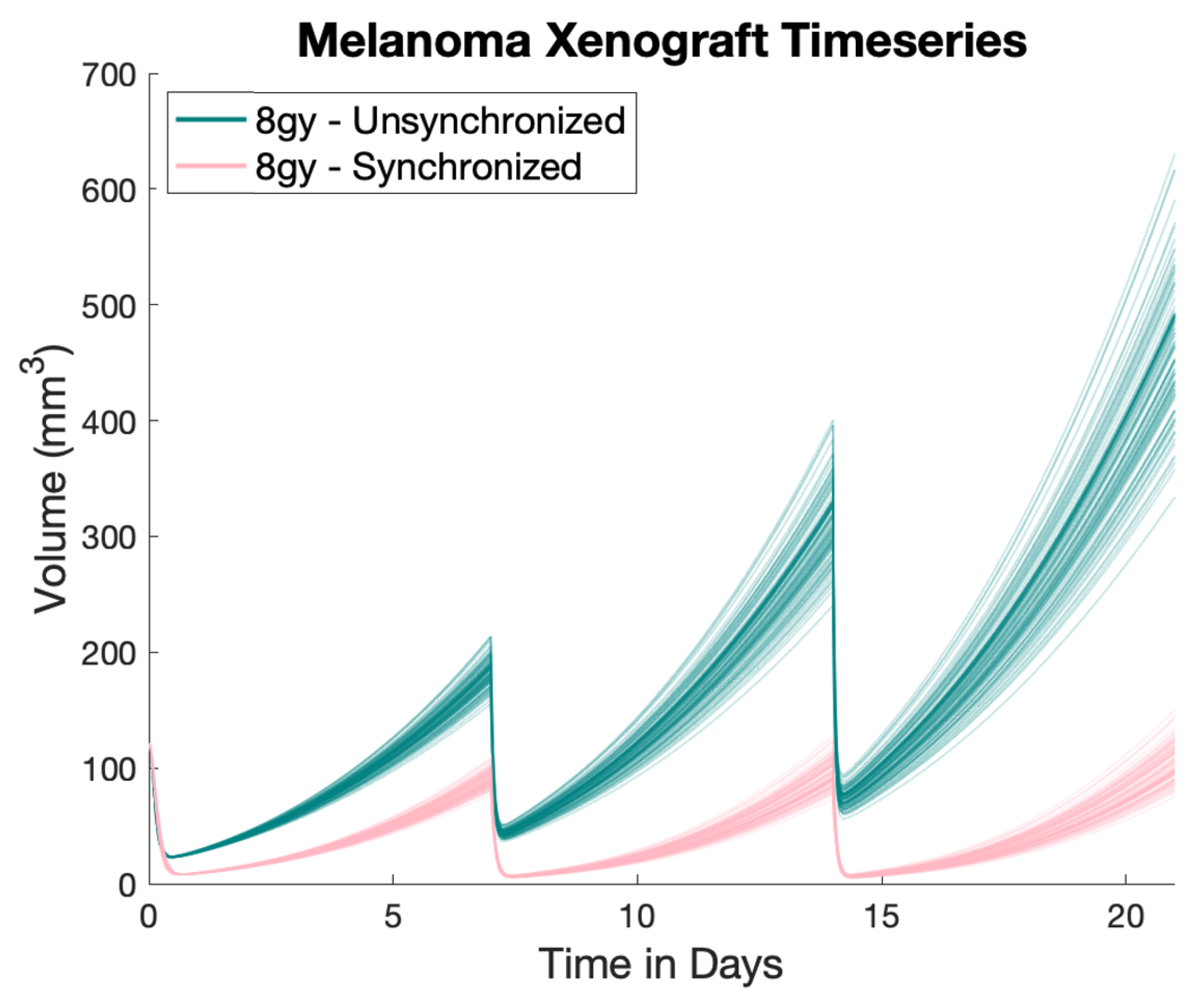
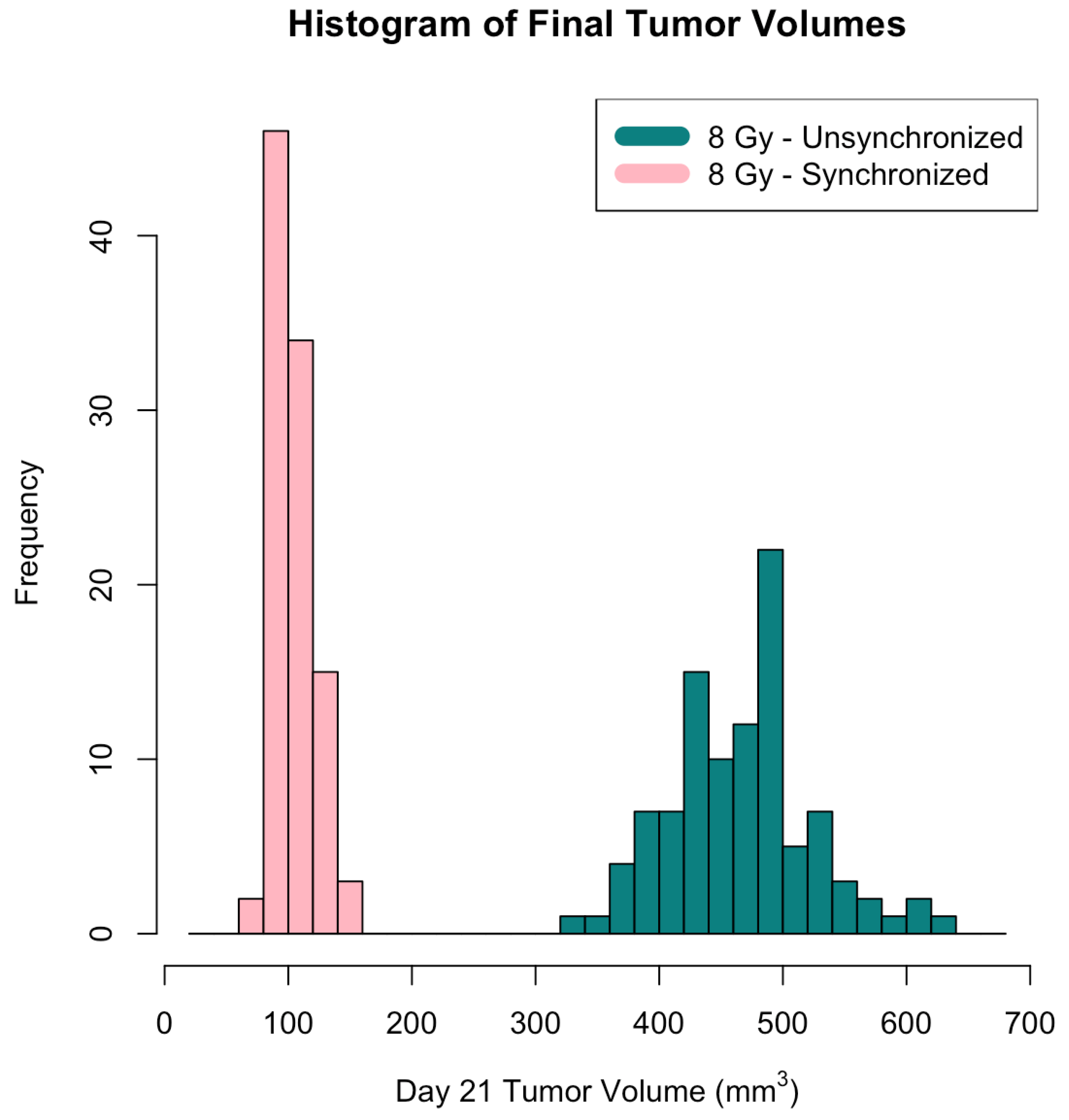
3. Results
4. Discussion
Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Model Equations
Appendix A.1. Proliferating Tumor Cells (G1-Phase)
Appendix A.2. Proliferating Tumor Cells (S-Phase)
Appendix A.3. Proliferating Tumor Cells (G2 Phase)
Appendix A.4. Quiescent Cells
Appendix A.5. Necrotic Cells
Appendix A.6. TNF-α
Appendix A.7. VEGF Production and Removal
Appendix A.8. Development of Vasculature
Appendix B. Model Parameters

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monolayer | Spheroid | Xenograft | Explanation | |
|---|---|---|---|---|
| 0 | 0 | 0 | Natural Death Rate | |
| 1.53372 | 1.53372 | 1.53372 | to S Transition | |
| 1.5004 | 1.5004 | 1.5004 | S to Transition | |
| 63.104 | 63.104 | 63.104 | to Transition | |
| N/A | 2.9 | 100 | Ambient Nutrient | |
| j | N/A | 1.1 | 1.1 | TNF- Production |
| k | N/A | 0.14 | 0.14 | TNF- Removal |
| N/A | 1 | 1 | B Half-Saturation Constant | |
| N/A | 300 | 300 | C Half-Saturation Constant | |
| e | N/A | 0.08 | 0.08 | Necrotic Death of Q |
| m | N/A | 0.004 | 0.004 | Natural Removal of N |
| N/A | 60 | 60 | F Half-Saturation Constant | |
| N/A | 55 | 55 | Maximum Rate of C | |
| N/A | 0.83 | 0.83 | TNF--Induced Apoptosis of P | |
| N/A | N/A | 70 | Vascular Growth | |
| N/A | N/A | 223.05148 | Production of VEGF by P | |
| N/A | N/A | 103.45178 | Stimulation of VEGF by TNF- | |
| N/A | N/A | 5.50064 | VEGF Removal | |
| N/A | N/A | 0.01 | Production of VEGF by Q | |
| N/A | N/A | 0.17877 | Controls C | |
| N/A | N/A | 100 | Controls Vasculature | |
| N/A | N/A | 0.1 | H Half-Saturation Constant | |
| N/A | N/A | 0.1 | Controls H | |
| N/A | N/A | 0.1 | Controls H |
| Monolayer | Spheroid | Xenograft | Explanation | |
|---|---|---|---|---|
| cells | 0.00908 mm3 | 69.76 mm3 | Phase Cells | |
| cells | 0.00657 mm3 | 50.50 mm3 | S Phase Cells | |
| cells | 0.000155 mm3 | 1.190 mm3 | Phase Cells | |
| N/A | 0 | 0 | Quiescent Cells (mm3) | |
| N/A | 0 | 0 | Necrosis (mm3) | |
| N/A | 0 | 0 | TNF- | |
| N/A | N/A | 0 | VEGF | |
| N/A | N/A | 0 | Vasculature |
Appendix C. Tabulated Data from Browning et al., Dorasamy et al., and Otani et al.
| Day | ||||
|---|---|---|---|---|
| 3 | 0.0158 | 0.0158 | 0.0001 | 0 |
| 4 | 0.0215 | 0.0212 | 0.0002 | 0 |
| 5 | 0.0262 | 0.0258 | 0.0004 | 0 |
| 7 | 0.0477 | 0.0406 | 0.0071 | 0 |
| 10 | 0.0903 | 0.0428 | 0.0434 | 0.0041 |
| 12 | 0.0922 | 0.0395 | 0.0436 | 0.0091 |
| 14 | 0.1287 | 0.0479 | 0.0617 | 0.0192 |
| 16 | 0.1475 | 0.0528 | 0.0638 | 0.0308 |
| 18 | 0.1503 | 0.0471 | 0.0605 | 0.0427 |
| 21 | 0.1669 | 0.0448 | 0.0540 | 0.0680 |
| Day | Volume (Mean ± SEM) |
|---|---|
| 1 | |
| 4 | |
| 6 | |
| 8 | |
| 11 | |
| 14 |
| Grays | W | Q | S | ||
|---|---|---|---|---|---|
| 0 | 1 | 1 | 1 | 1 | 1 |
| 2 | 0.9896 | 0.9722 | 0.8403 | 0.9810 | 1 |
| 4 | 0.6643 | 0.7573 | 0.6314 | 0.5037 | 0.8007 |
| 8 | 0.1964 | 0.3175 | 0.2449 | 0.03154 | 0.1804 |
References
- Aggarwal, P.; Knabel, P.; Fleischer, A.B., Jr. United States burden of melanoma and non-melanoma skin cancer from 1990 to 2019. J. Am. Acad. Dermatol. 2021, 85, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.R.; Rzucidlo, E. Acute and chronic radiation injury. J. Vasc. Surg. 2011, 53, 15S–21S. [Google Scholar] [CrossRef] [PubMed]
- Matthews, N.H.; Li, W.Q.; Qureshi, A.A.; Weinstock, M.A.; Cho, E.; Norval, M.; Wright, C.Y.; Belter, B.; Haase-Kohn, C.; Pietzsch, J.; et al. Cutaneous melanoma. Lancet 2017, 383, 816–827. [Google Scholar]
- Shi, W. Radiation Therapy for Melanoma; Exon Publications: Brisbane City, QLD, Australia, 2017; pp. 101–120. [Google Scholar]
- Trappetti, V.; Potez, M.; Fernandez-Palomo, C.; Volarevic, V.; Shintani, N.; Pellicioli, P.; Ernst, A.; Haberthür, D.; Fazzari, J.M.; Krisch, M.; et al. Microbeam Radiation Therapy controls local growth of radioresistant melanoma and treats out-of-field locoregional metastasis. Int. J. Radiat. Oncol. Biol. Phys. 2022, 114, 478–493. [Google Scholar] [CrossRef] [PubMed]
- Tagliaferri, L.; Lancellotta, V.; Fionda, B.; Mangoni, M.; Casà, C.; Di Stefani, A.; Pagliara, M.M.; D’Aviero, A.; Schinzari, G.; Chiesa, S.; et al. Immunotherapy and radiotherapy in melanoma: A multidisciplinary comprehensive review. Hum. Vaccines Immunother. 2022, 18, 1903827. [Google Scholar] [CrossRef] [PubMed]
- Shimono, H.; Kaida, A.; Homma, H.; Nojima, H.; Onozato, Y.; Harada, H.; Miura, M. Fluctuation in radioresponse of HeLa cells during the cell cycle evaluated based on micronucleus frequency. Sci. Rep. 2020, 10, 20873. [Google Scholar] [CrossRef] [PubMed]
- Menegakis, A.; Klompmaker, R.; Vennin, C.; Arbusà, A.; Damen, M.; van den Broek, B.; Zips, D.; van Rheenen, J.; Krenning, L.; Medema, R.H. Resistance of hypoxic cells to ionizing radiation is mediated in part via hypoxia-induced quiescence. Cells 2021, 10, 610. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Ozanne, S.E.; Hales, C.N. Heterogeneity in premature senescence by oxidative stress correlates with differential DNA damage during the cell cycle. DNA Repair 2005, 4, 1140–1148. [Google Scholar] [CrossRef]
- Pawlik, T.M.; Keyomarsi, K. Role of cell cycle in mediating sensitivity to radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2004, 59, 928–942. [Google Scholar] [CrossRef]
- Hauge, S.; Eek Mariampillai, A.; Rødland, G.E.; Bay, L.T.; Landsverk, H.B.; Syljuåsen, R.G. Expanding roles of cell cycle checkpoint inhibitors in radiation oncology. Int. J. Radiat. Biol. 2023, 99, 941–950. [Google Scholar] [CrossRef]
- Seong, K.M.; Kim, C.S.; Jeon, H.Y.; Oh, S.H.; Nam, S.Y.; Yang, K.H.; Kim, J.Y.; Jin, Y.W. Intrinsic radiosensitivity correlated with radiation-induced ROS and cell cycle regulation. Mol. Cell. Toxicol. 2010, 6, 1–7. [Google Scholar] [CrossRef]
- Yount, G.L.; Haas-Kogan, D.A.; Vidair, C.A.; Haas, M.; Dewey, W.C.; Israel, M.A. Cell cycle synchrony unmasks the influence of p53 function on radiosensitivity of human glioblastoma cells. Cancer Res. 1996, 56, 500–506. [Google Scholar] [PubMed]
- Pan, Z.; Qu, C.; Chen, Y.; Chen, X.; Liu, X.; Hao, W.; Xu, W.; Ye, L.; Lu, P.; Li, D.; et al. Bufotalin induces cell cycle arrest and cell apoptosis in human malignant melanoma A375 cells. Oncol. Rep. 2019, 41, 2409–2417. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Luo, Y.; Xia, Y.; Zhang, X.; Qin, Y.; Liu, W.; Li, M.; Liu, X.; Zheng, Q.; Li, D. Cinobufagin induces cell cycle arrest at the S phase and promotes apoptosis in nasopharyngeal carcinoma cells. Biomed. Pharmacother. 2020, 122, 109763. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Fan, X.; Han, Y.; Wu, B.X.; Zhu, B. Daphnoretin Arrests the Cell Cycle and Induces Apoptosis in Human Breast Cancer Cells. J. Nat. Prod. 2022, 85, 2332–2339. [Google Scholar] [CrossRef] [PubMed]
- Kee, J.Y.; Han, Y.H.; Kim, D.S.; Mun, J.G.; Park, S.H.; So, H.S.; Park, S.J.; Park, R.; Um, J.Y.; Hong, S.H. β-Lapachone suppresses the lung metastasis of melanoma via the MAPK signaling pathway. PLoS ONE 2017, 12, e0176937. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, W.K.; Morton, R.A. X-Ray and Ultraviolet Sensitivity of Synchronized Chinese Hamster Cells at Various Stages of the Cell Cycle. Biophys. J. 1965, 5, 1–25. [Google Scholar] [CrossRef]
- Otani, K.; Naito, Y.; Sakaguchi, Y.; Seo, Y.; Takahashi, Y.; Kikuta, J.; Ogawa, K.; Ishii, M. Cell-cycle-controlled radiation therapy was effective for treating a murine malignant melanoma cell line in vitro and in vivo. Sci. Rep. 2016, 6, 30689. [Google Scholar] [CrossRef]
- Terasima, T.; Tolmach, L.J. Variations in several responses of HeLa cells to x-irradiation during the division cycle. Biophys. J. 1963, 3, 11–33. [Google Scholar] [CrossRef]
- Hufnagl, A.; Herr, L.; Friedrich, T.; Durante, M.; Taucher-Scholz, G.; Scholz, M. The link between cell-cycle dependent radiosensitivity and repair pathways: A model based on the local, sister-chromatid conformation dependent switch between NHEJ and HR. DNA Repair 2015, 27, 28–39. [Google Scholar] [CrossRef]
- Buffa, F.M.; West, C.; Byrne, K.; Moore, J.V.; Nahum, A.E. Radiation response and cure rate of human colon adenocarcinoma spheroids of different size: The significance of hypoxia on tumor control modelling. Int. J. Radiat. Oncol. Biol. Phys. 2001, 49, 1109–1118. [Google Scholar] [CrossRef] [PubMed]
- Kempf, H.; Hatzikirou, H.; Bleicher, M.; Meyer-Hermann, M. In silico analysis of cell cycle synchronisation effects in radiotherapy of tumour spheroids. PLoS Comput. Biol. 2013, 9, e1003295. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Kodali, A.; Wallace, D.I. Predictive modeling of neuroblastoma growth dynamics in xenograft model after bevacizumab anti-VEGF therapy. Bull. Math. Biol. 2018, 80, 2026–2048. [Google Scholar] [CrossRef] [PubMed]
- West, C.M.; Sutherland, R.M. The radiation response of a human colon adenocarcinoma grown in monolayer, as spheroids, and in nude mice. Radiat. Res. 1987, 112, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Lv, D.; Hu, Z.; Lu, L.; Lu, H.; Xu, X. Three-dimensional cell culture: A powerful tool in tumor research and drug discovery. Oncol. Lett. 2017, 14, 6999–7010. [Google Scholar] [CrossRef] [PubMed]
- Habanjar, O.; Diab-Assaf, M.; Caldefie-Chezet, F.; Delort, L. 3D cell culture systems: Tumor application, advantages, and disadvantages. Int. J. Mol. Sci. 2021, 22, 12200. [Google Scholar] [CrossRef] [PubMed]
- Chaicharoenaudomrung, N.; Kunhorm, P.; Noisa, P. Three-dimensional cell culture systems as an in vitro platform for cancer and stem cell modeling. World J. Stem Cells 2019, 11, 1065. [Google Scholar] [CrossRef] [PubMed]
- Fontana, F.; Marzagalli, M.; Sommariva, M.; Gagliano, N.; Limonta, P. In vitro 3D cultures to model the tumor microenvironment. Cancers 2021, 13, 2970. [Google Scholar] [CrossRef]
- Zanoni, M.; Piccinini, F.; Arienti, C.; Zamagni, A.; Santi, S.; Polico, R.; Bevilacqua, A.; Tesei, A. 3D tumor spheroid models for in vitro therapeutic screening: A systematic approach to enhance the biological relevance of data obtained. Sci. Rep. 2016, 6, 19103. [Google Scholar] [CrossRef]
- Weiswald, L.B.; Bellet, D.; Dangles-Marie, V. Spherical cancer models in tumor biology. Neoplasia 2015, 17, 1–15. [Google Scholar] [CrossRef]
- Nath, S.; Devi, G.R. Three-dimensional culture systems in cancer research: Focus on tumor spheroid model. Pharmacol. Ther. 2016, 163, 94–108. [Google Scholar] [CrossRef]
- Wallace, D.I.; Dunham, A.; Chen, P.X.; Chen, M.; Huynh, M.; Rheingold, E.; Prosper, O. A model for spheroid versus monolayer response of SK-N-SH neuroblastoma cells to treatment with 15-deoxy-PGJ 2. Comput. Math. Methods Med. 2016, 2016, 3628124. [Google Scholar] [CrossRef]
- Browning, A.P.; Sharp, J.A.; Murphy, R.J.; Gunasingh, G.; Lawson, B.; Burrage, K.; Haass, N.K.; Simpson, M. Quantitative analysis of tumour spheroid structure. Elife 2021, 10, e73020. [Google Scholar] [CrossRef] [PubMed]
- Dorasamy, M.S.; Aravind, A.; Nellore, K.; Wong, P.F. Synergistic inhibition of melanoma xenografts by Brequinar sodium and Doxorubicin. Biomed. Pharmacother. 2019, 110, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Sause, W.; Cooper, J.; Rush, S.; Ago, C.; Cosmatos, D.; Coughlin, C.; Janjan, N.; Lipsett, J. Fraction size in external beam radiation therapy in the treatment of melanoma. Int. J. Radiat. Oncol. Biol. Phys. 1991, 20, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Vittadello, S.T.; McCue, S.W.; Gunasingh, G.; Haass, N.K.; Simpson, M.J. Mathematical models incorporating a multi-stage cell cycle replicate normally-hidden inherent synchronization in cell proliferation. J. R. Soc. Interface 2019, 16, 20190382. [Google Scholar] [CrossRef] [PubMed]
- Liberato, T.; Pessotti, D.S.; Fukushima, I.; Kitano, E.S.; Serrano, S.M.; Zelanis, A. Signatures of protein expression revealed by secretome analyses of cancer associated fibroblasts and melanoma cell lines. J. Proteom. 2018, 174, 1–8. [Google Scholar] [CrossRef] [PubMed]
- MATLAB. MATLAB and Statistics Toolbox Release 2018a; The MathWorks Inc.: Natick, MA, USA, 2018. [Google Scholar]
- Kaeppler, J.R.; Chen, J.; Buono, M.; Vermeer, J.; Kannan, P.; Cheng, W.C.; Voukantsis, D.; Thompson, J.M.; Hill, M.A.; Allen, D.; et al. Endothelial cell death after ionizing radiation does not impair vascular structure in mouse tumor models. EMBO Rep. 2022, 23, e53221. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rentzeperis, F.; Coleman, B.; Wallace, D. Cell-Cycle Synchronization Prior to Radiotherapy: A Mathematical Model of the Use of Gemcitabine on Melanoma Xenografts. AppliedMath 2024, 4, 197-210. https://0-doi-org.brum.beds.ac.uk/10.3390/appliedmath4010010
Rentzeperis F, Coleman B, Wallace D. Cell-Cycle Synchronization Prior to Radiotherapy: A Mathematical Model of the Use of Gemcitabine on Melanoma Xenografts. AppliedMath. 2024; 4(1):197-210. https://0-doi-org.brum.beds.ac.uk/10.3390/appliedmath4010010
Chicago/Turabian StyleRentzeperis, Frederika, Benjamin Coleman, and Dorothy Wallace. 2024. "Cell-Cycle Synchronization Prior to Radiotherapy: A Mathematical Model of the Use of Gemcitabine on Melanoma Xenografts" AppliedMath 4, no. 1: 197-210. https://0-doi-org.brum.beds.ac.uk/10.3390/appliedmath4010010