
Proteomic and Bioinformatic Analysis of Decellularized Pancreatic Extracellular Matrices
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
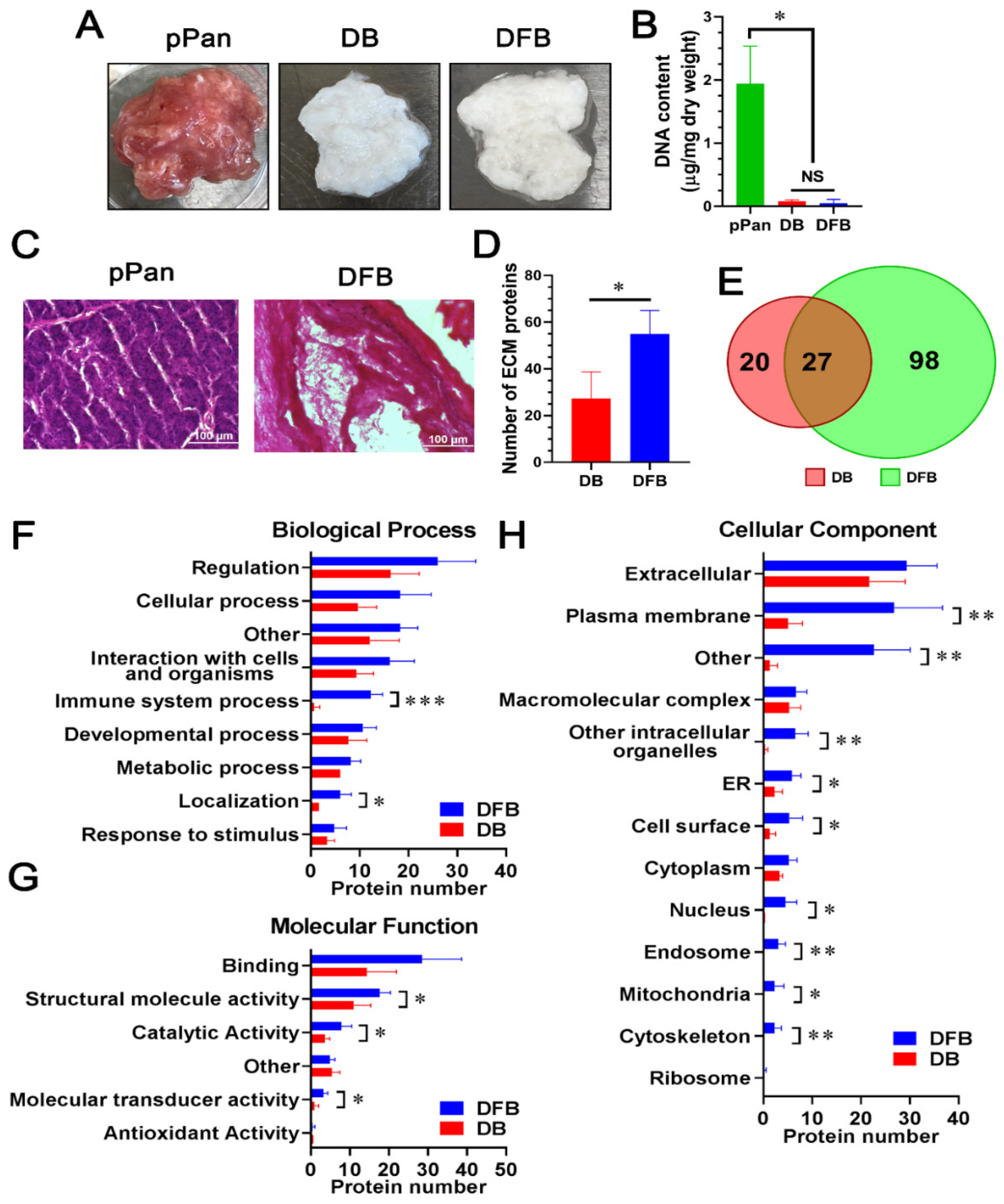
2.1. Comparison of Detergent-Free with Detergent-Based Decellularization of Porcine Pancreata
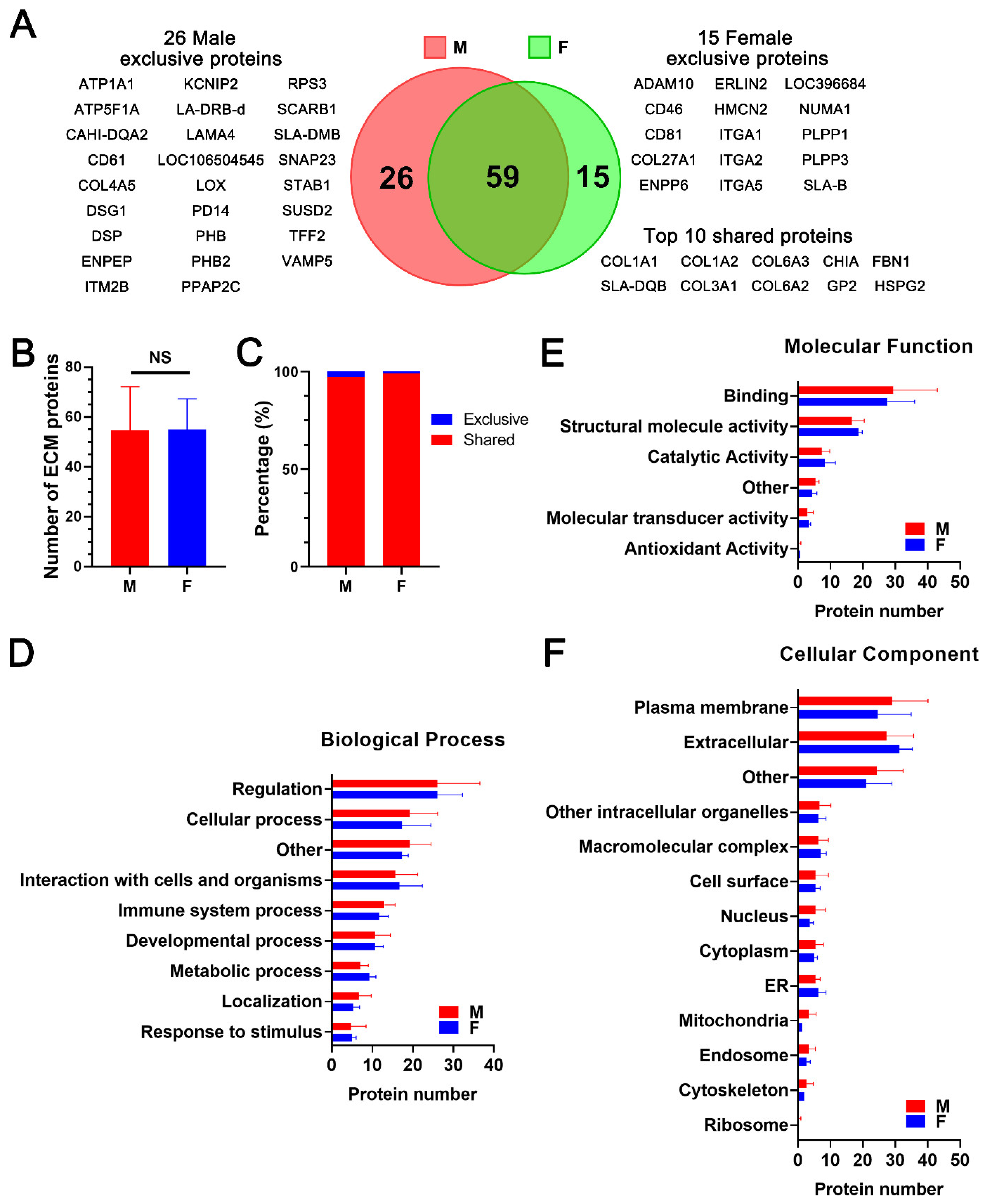
2.2. Proteomic Analysis and Bioinformatic Characterization of dpECM Suggest That the Composition of dpECM Proteins Is Affected by Gender in Adult Tissues
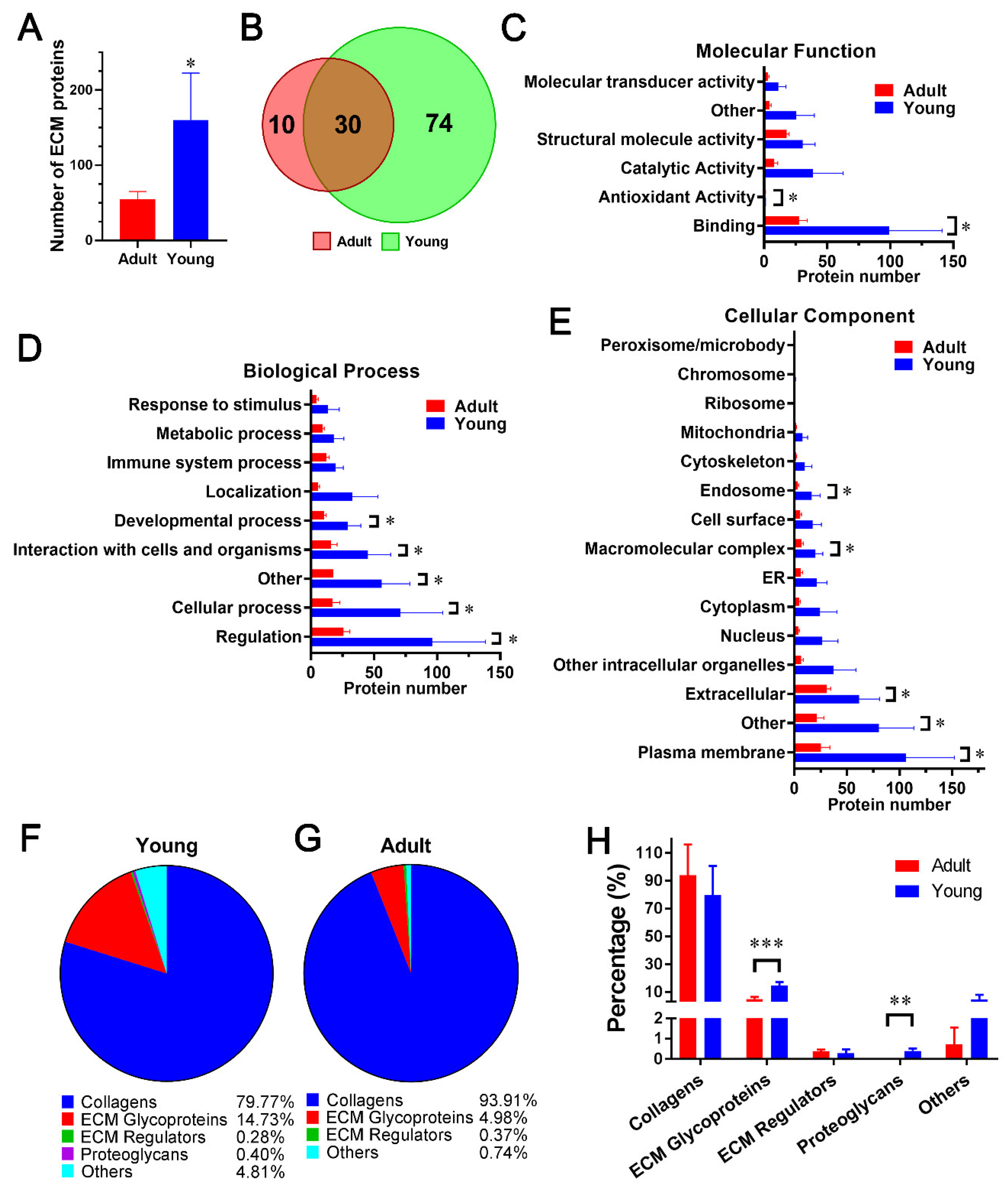
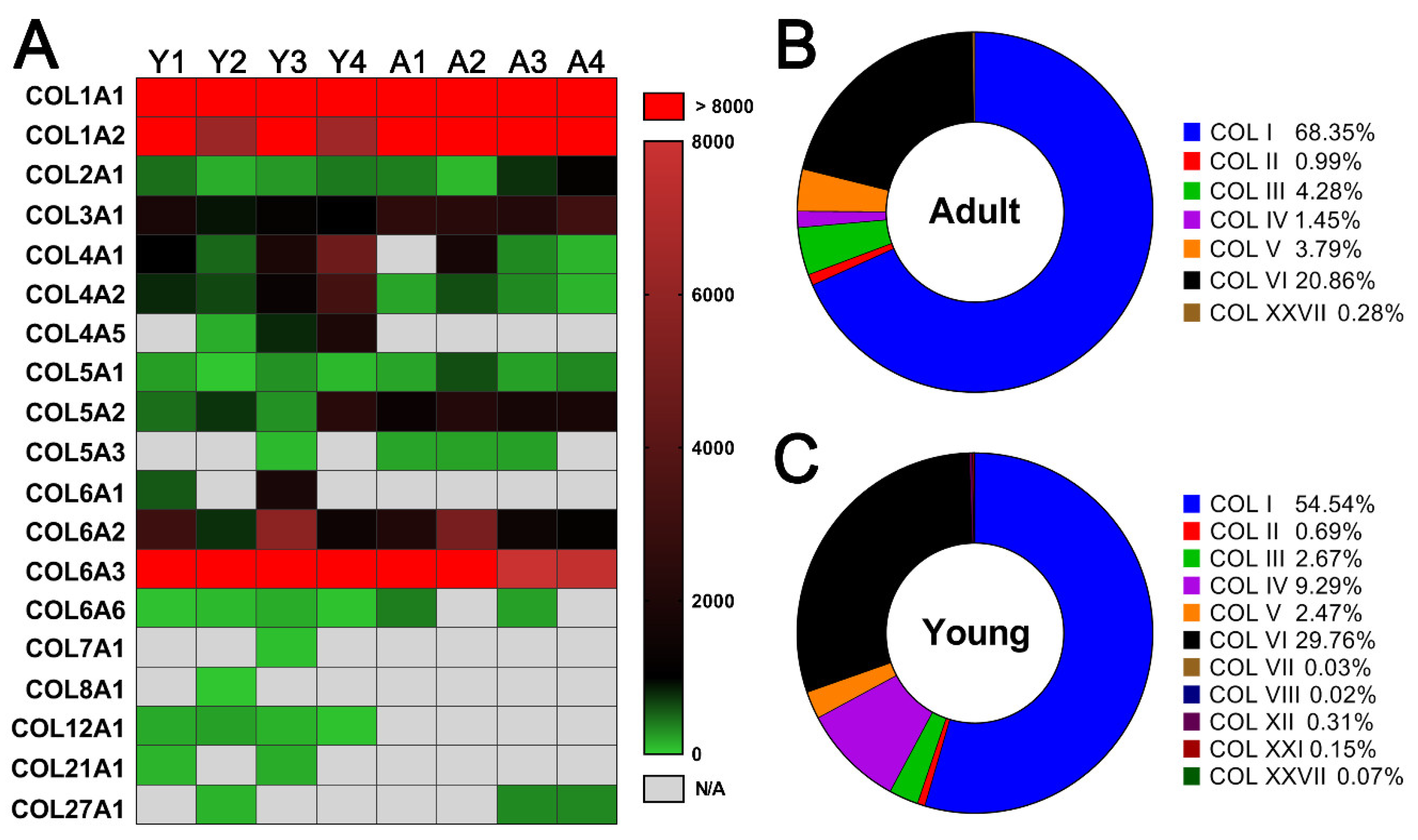
2.3. Comparison of dpECM Proteins Detected inYoung and Adult Porcine Pancreata
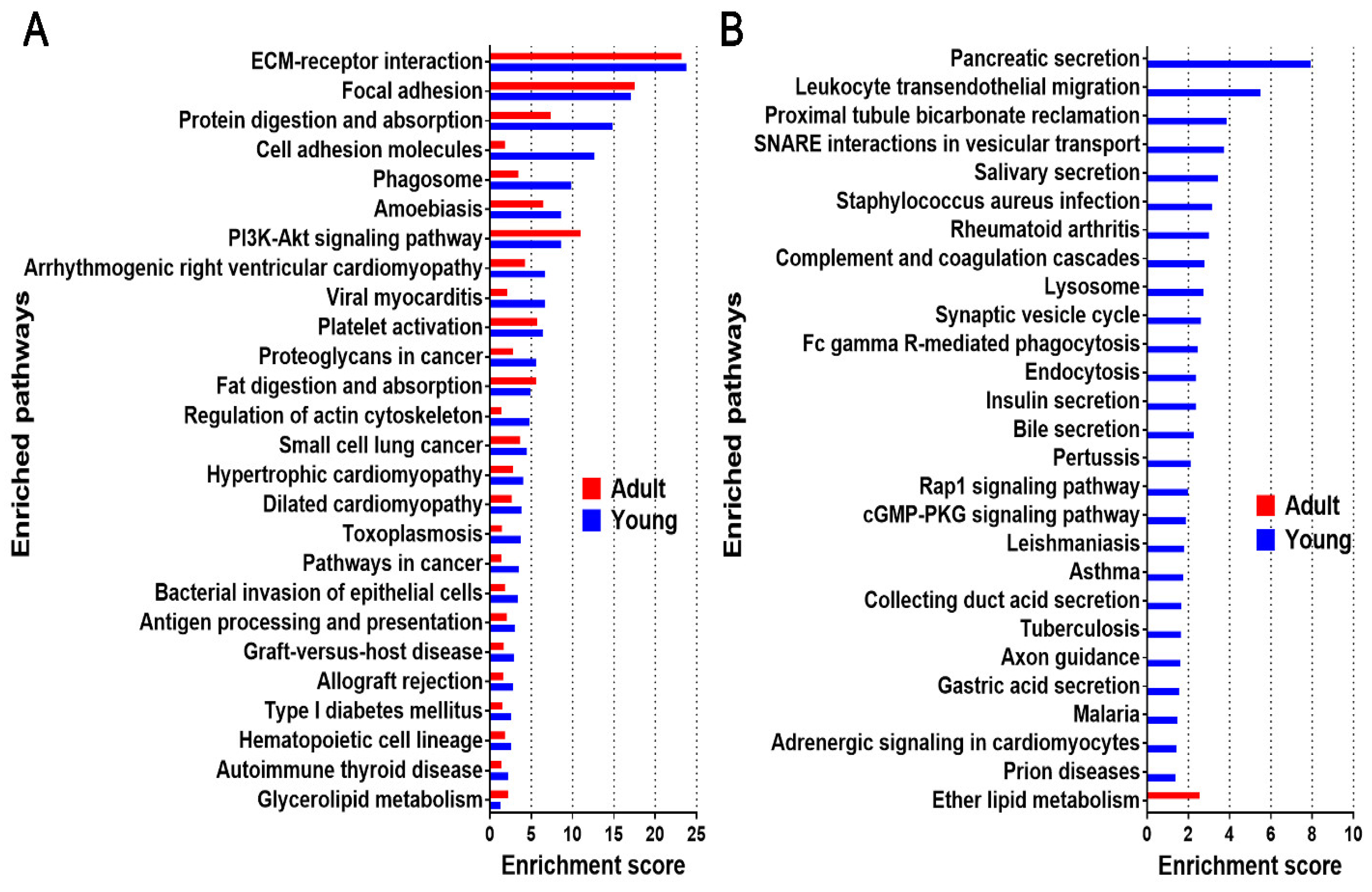
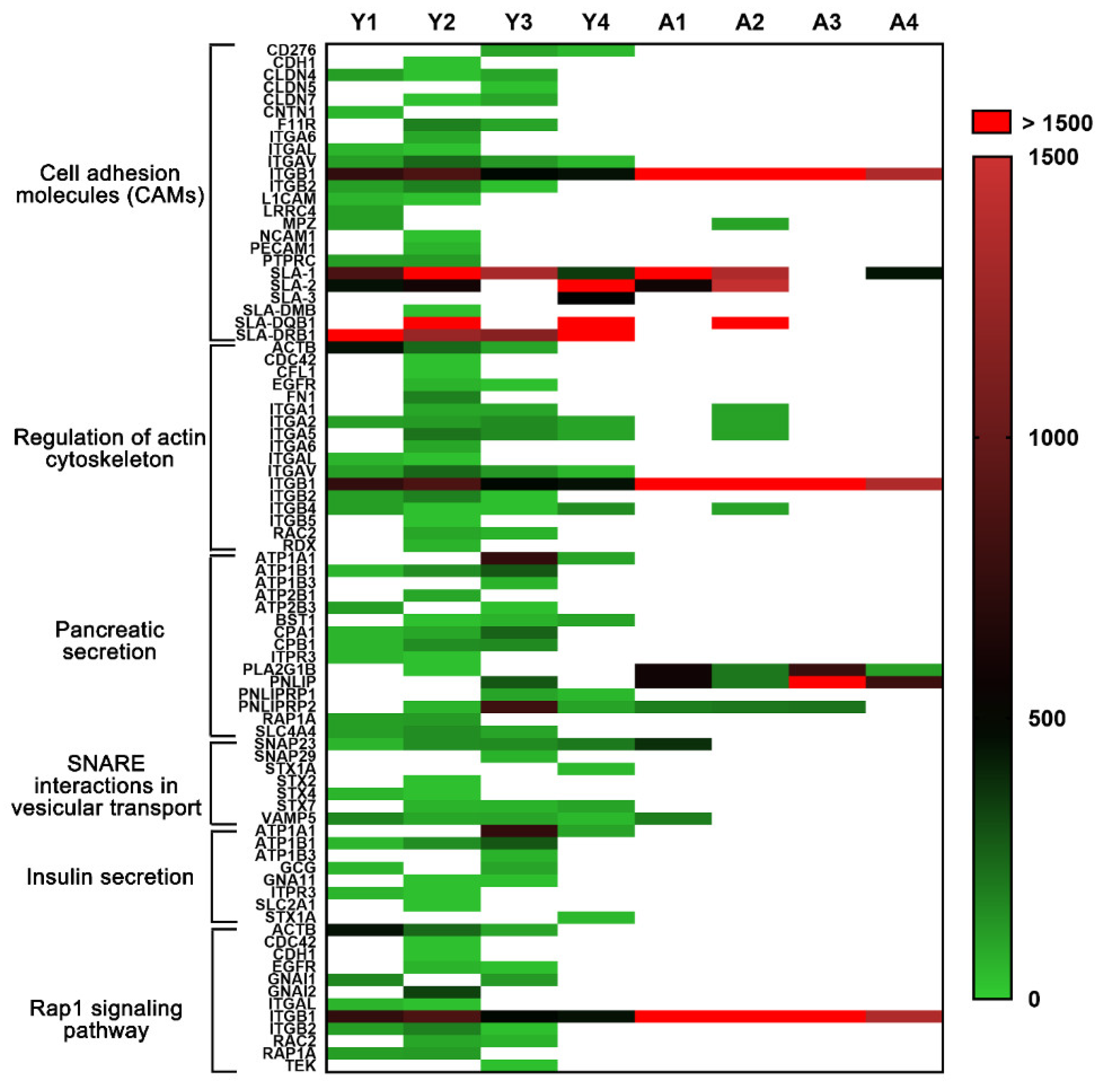
2.4. Enriched Pathway Analysis for Young and Adult dpECM
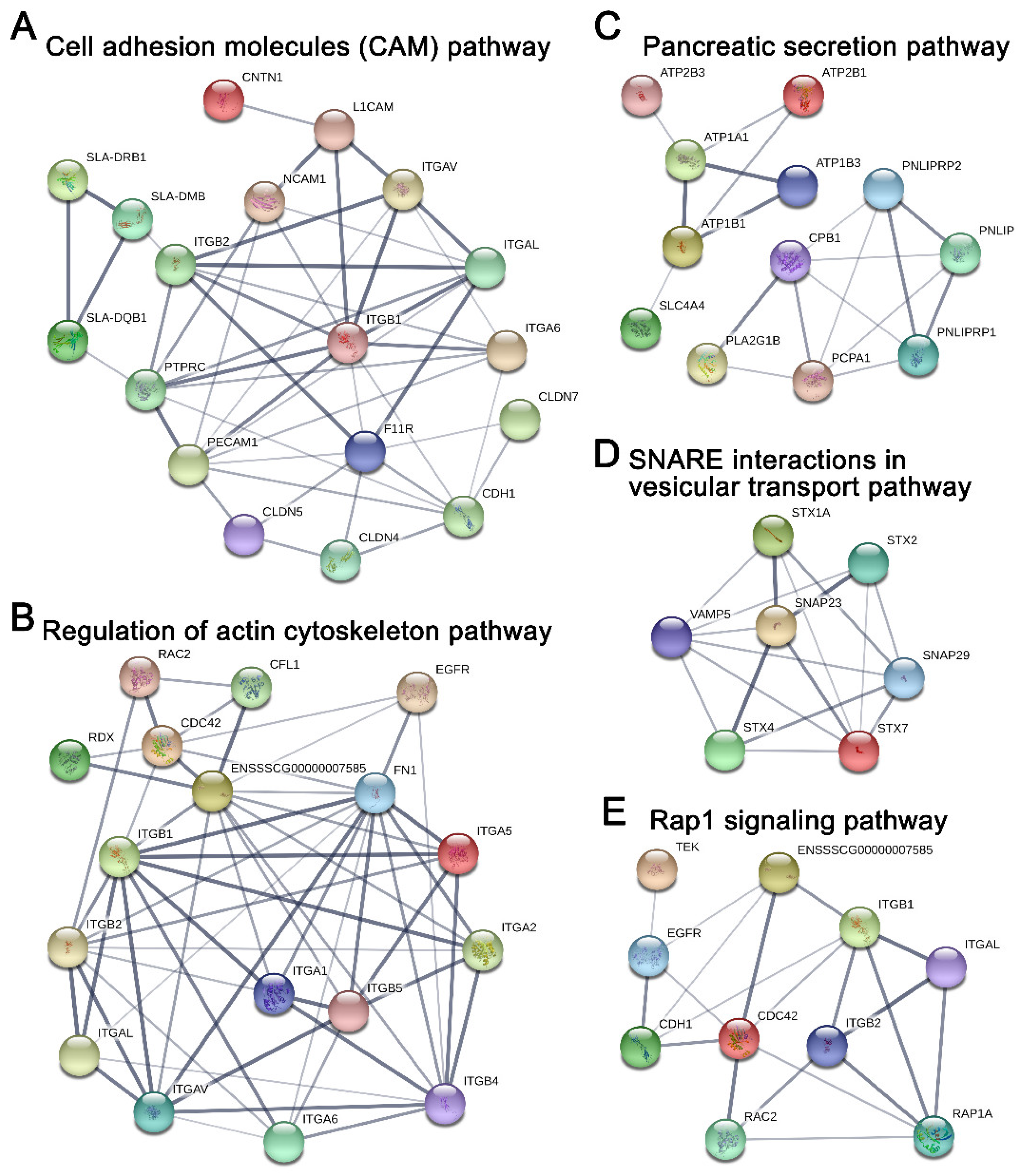
2.5. Potential Protein Networks Enriched in Young and Adult dpECM
3. Discussion
4. Materials and Methods
4.1. dpECM Preparation
4.2. DNA Quantification and Eosin Staining
4.3. dpECM Reconstitution
4.4. Sample Preparation for Mass Spectrometry
4.5. Liquid Chromatography–Tandem Mass Spectrometry (LC–MS/MS)
4.6. Proteomic Data Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Herrera, J.; Henke, C.A.; Bitterman, P.B. Extracellular matrix as a driver of progressive fibrosis. J. Clin. Investig. 2018, 128, 45–53. [Google Scholar] [CrossRef]
- Senthebane, D.A.; Jonker, T.; Rowe, A.; Thomford, N.E.; Munro, D.; Dandara, C.; Wonkam, A.; Govender, D.; Calder, B.; Soares, N.C.; et al. The Role of Tumor Microenvironment in Chemoresistance: 3D Extracellular Matrices as Accomplices. Int. J. Mol. Sci. 2018, 19, 2861. [Google Scholar] [CrossRef] [PubMed]
- Muncie, J.M.; Weaver, V.M. The Physical and Biochemical Properties of the Extracellular Matrix Regulate Cell Fate. Curr. Top. Dev. Biol. 2018, 130, 1–37. [Google Scholar]
- Huang, H.; Bader, T.N.; Jin, S. Signaling Molecules Regulating Pancreatic Endocrine Development from Pluripotent Stem Cell Differentiation. Int. J. Mol. Sci. 2020, 21, 5867. [Google Scholar] [CrossRef] [PubMed]
- Guruswamy Damodaran, R.; Vermette, P. Decellularized pancreas as a native extracellular matrix scaffold for pancreatic islet seeding and culture. J. Tissue Eng. Regen. Med. 2018, 12, 1230–1237. [Google Scholar] [CrossRef]
- Sackett, S.D.; Tremmel, D.M.; Ma, F.; Feeney, A.K.; Maguire, R.M.; Brown, M.E.; Zhou, Y.; Li, X.; O’Brien, C.; Li, L.; et al. Extracellular matrix scaffold and hydrogel derived from decellularized and delipidized human pancreas. Sci. Rep. 2018, 8, 10452. [Google Scholar] [CrossRef] [PubMed]
- Elebring, E.; Kuna, V.K.; Kvarnstrom, N.; Sumitran-Holgersson, S. Cold-perfusion decellularization of whole-organ porcine pancreas supports human fetal pancreatic cell attachment and expression of endocrine and exocrine markers. J. Tissue Eng. 2017, 8, 2041731417738145. [Google Scholar] [CrossRef]
- Jiang, K.; Chaimov, D.; Patel, S.N.; Liang, J.P.; Wiggins, S.C.; Samojlik, M.M.; Rubiano, A.; Simmons, C.S.; Stabler, C.L. 3-D physiomimetic extracellular matrix hydrogels provide a supportive microenvironment for rodent and human islet culture. Biomaterials 2019, 198, 37–48. [Google Scholar] [CrossRef]
- Bi, H.; Karanth, S.; Ye, K.; Stein, R.; Jin, S. Decellularized Tissue Matrix Enhances Self-Assembly of Islet Organoids from Pluripotent Stem Cell Differentiation. ACS Biomater. Sci. Eng. 2020, 6, 4155–4165. [Google Scholar] [CrossRef]
- Bi, H.; Ye, K.; Jin, S. Proteomic analysis of decellularized pancreatic matrix identifies collagen V as a critical regulator for islet organogenesis from human pluripotent stem cells. Biomaterials 2020, 233, 119673. [Google Scholar] [CrossRef]
- Karanth, S.S.; Sun, S.; Bi, H.; Ye, K.; Jin, S. Angiopoietins stimulate pancreatic islet development from stem cells. Sci. Rep. 2021, 11, 13558. [Google Scholar] [CrossRef]
- Haydont, V.; Bernard, B.A.; Fortunel, N.O. Age-related evolutions of the dermis: Clinical signs, fibroblast and extracellular matrix dynamics. Mech. Ageing Dev. 2019, 177, 150–156. [Google Scholar] [CrossRef]
- Duca, L.; Blaise, S.; Romier, B.; Laffargue, M.; Gayral, S.; El Btaouri, H.; Kawecki, C.; Guillot, A.; Martiny, L.; Debelle, L.; et al. Matrix ageing and vascular impacts: Focus on elastin fragmentation. Cardiovasc. Res. 2016, 110, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Fusco, J.; Zimmerman, R.; Fischbach, S.; Chen, C.; Ricks, D.M.; Prasadan, K.; Shiota, C.; Xiao, X.; Gittes, G.K. Epidermal Growth Factor Receptor Signaling Regulates beta Cell Proliferation in Adult Mice. J. Biol. Chem. 2016, 291, 22630–22637. [Google Scholar] [CrossRef] [PubMed]
- Henquin, J.C.; Nenquin, M. Immaturity of insulin secretion by pancreatic islets isolated from one human neonate. J. Diabetes Investig. 2018, 9, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Jacovetti, C.; Matkovich, S.J.; Rodriguez-Trejo, A.; Guay, C.; Regazzi, R. Postnatal beta-cell maturation is associated with islet-specific microRNA changes induced by nutrient shifts at weaning. Nat. Commun. 2015, 6, 8084. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.S.; Hebrok, M. All mixed up: Defining roles for beta-cell subtypes in mature islets. Genes Dev. 2017, 31, 228–240. [Google Scholar] [CrossRef] [PubMed]
- Guruswamy Damodaran, R.; Vermette, P. Tissue and organ decellularization in regenerative medicine. Biotechnol. Prog. 2018, 34, 1494–1505. [Google Scholar] [CrossRef]
- Hodge, K.; Have, S.T.; Hutton, L.; Lamond, A.I. Cleaning up the masses: Exclusion lists to reduce contamination with HPLC-MS/MS. J. Proteom. 2013, 88, 92–103. [Google Scholar] [CrossRef]
- Gaetani, R.; Aude, S.; DeMaddalena, L.L.; Strassle, H.; Dzieciatkowska, M.; Wortham, M.; Bender, R.H.F.; Nguyen-Ngoc, K.V.; Schmid-Schoenbein, G.W.; George, S.C.; et al. Evaluation of Different Decellularization Protocols on the Generation of Pancreas-Derived Hydrogels. Tissue Eng. Part C Methods 2018, 24, 697–708. [Google Scholar] [CrossRef]
- Mirmalek-Sani, S.H.; Orlando, G.; McQuilling, J.P.; Pareta, R.; Mack, D.L.; Salvatori, M.; Farney, A.C.; Stratta, R.J.; Atala, A.; Opara, E.C.; et al. Porcine pancreas extracellular matrix as a platform for endocrine pancreas bioengineering. Biomaterials 2013, 34, 5488–5495. [Google Scholar] [CrossRef] [PubMed]
- Salvatori, M.; Katari, R.; Patel, T.; Peloso, A.; Mugweru, J.; Owusu, K.; Orlando, G. Extracellular Matrix Scaffold Technology for Bioartificial Pancreas Engineering: State of the Art and Future Challenges. J. Diabetes Sci. Technol. 2014, 8, 159–169. [Google Scholar] [CrossRef]
- Chaimov, D.; Baruch, L.; Krishtul, S.; Meivar-Levy, I.; Ferber, S.; Machluf, M. Innovative encapsulation platform based on pancreatic extracellular matrix achieve substantial insulin delivery. J. Control Release 2017, 257, 91–101. [Google Scholar] [CrossRef]
- Napierala, H.; Hillebrandt, K.H.; Haep, N.; Tang, P.; Tintemann, M.; Gassner, J.; Noesser, M.; Everwien, H.; Seiffert, N.; Kluge, M.; et al. Engineering an endocrine Neo-Pancreas by repopulation of a decellularized rat pancreas with islets of Langerhans. Sci. Rep. 2017, 7, 41777. [Google Scholar] [CrossRef]
- Faulk, D.M.; Carruthers, C.A.; Warner, H.J.; Kramer, C.R.; Reing, J.E.; Zhang, L.; D’Amore, A.; Badylak, S.F. The effect of detergents on the basement membrane complex of a biologic scaffold material. Acta Biomater. 2014, 10, 183–193. [Google Scholar] [CrossRef]
- Poornejad, N.; Schaumann, L.B.; Buckmiller, E.M.; Momtahan, N.; Gassman, J.R.; Ma, H.H.; Roeder, B.L.; Reynolds, P.R.; Cook, A.D. The impact of decellularization agents on renal tissue extracellular matrix. J. Biomater. Appl. 2016, 31, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Bi, H.; Ming, L.; Cheng, R.; Luo, H.; Zhang, Y.; Jin, Y. Liver extracellular matrix promotes BM-MSCs hepatic differentiation and reversal of liver fibrosis through activation of integrin pathway. J. Tissue Eng. Regen Med. 2017, 11, 2685–2698. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, E.; Follmann, F.; Jungersen, G.; Agerholm, J.S. A review of the human vs. porcine female genital tract and associated immune system in the perspective of using minipigs as a model of human genital Chlamydia infection. Vet. Res. 2015, 46, 116. [Google Scholar] [CrossRef]
- Dupree, E.J.; Jayathirtha, M.; Yorkey, H.; Mihasan, M.; Petre, B.A.; Darie, C.C. A Critical Review of Bottom-Up Proteomics: The Good, the Bad, and the Future of this Field. Proteomes 2020, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Verrastro, I.; Pasha, S.; Jensen, K.T.; Pitt, A.R.; Spickett, C.M. Mass spectrometry-based methods for identifying oxidized proteins in disease: Advances and challenges. Biomolecules 2015, 5, 378–411. [Google Scholar] [CrossRef]
- Tuli, L.; Ressom, H.W. LC-MS Based Detection of Differential Protein Expression. J. Proteom. Bioinform. 2009, 2, 416–438. [Google Scholar] [CrossRef] [PubMed]
- Karpievitch, Y.V.; Polpitiya, A.D.; Anderson, G.A.; Smith, R.D.; Dabney, A.R. Liquid Chromatography Mass Spectrometry-Based Proteomics: Biological and Technological Aspects. Ann. Appl. Stat. 2010, 4, 1797–1823. [Google Scholar] [CrossRef]
- Nakashima, Y.; Nahar, S.; Miyagi-Shiohira, C.; Kinjo, T.; Toyoda, Z.; Kobayashi, N.; Saitoh, I.; Watanabe, M.; Fujita, J.; Noguchi, H. A Liquid Chromatography with Tandem Mass Spectrometry-Based Proteomic Analysis of the Proteins Secreted by Human Adipose-Derived Mesenchymal Stem Cells. Cell Transplant. 2018, 27, 1469–1494. [Google Scholar] [CrossRef] [PubMed]
- Shirk, R.A.; Zhang, Z.; Winneker, R.C. Differential effects of estrogens and progestins on the anticoagulant tissue factor pathway inhibitor in the rat. J. Steroid Biochem. Mol. Biol. 2005, 94, 361–368. [Google Scholar] [CrossRef]
- Adeyanju, O.A.; Soetan, O.A.; Olatunji, L.A. Drospirenone-containing contraceptive exerts positive effects on cardiac uric acid and PAI-1 but not GSK-3: Improved safety profiles in contraception? Pathophysiology 2019, 26, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Le Calve, B.; Griveau, A.; Vindrieux, D.; Marechal, R.; Wiel, C.; Svrcek, M.; Gout, J.; Azzi, L.; Payen, L.; Cros, J.; et al. Lysyl oxidase family activity promotes resistance of pancreatic ductal adenocarcinoma to chemotherapy by limiting the intratumoral anticancer drug distribution. Oncotarget 2016, 7, 32100–32112. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Wu, C.; Fu, F.; You, X.; Ma, X.; Qin, F.; Li, T.; Wang, R.; Yuan, J. Effect of lysyl oxidase (LOX) on corpus cavernous fibrosis caused by ischaemic priapism. J. Cell Mol. Med. 2018, 22, 2018–2022. [Google Scholar] [CrossRef]
- De Giorgio, M.R.; Yoshioka, M.; Riedl, I.; Moreault, O.; Cherizol, R.G.; Shah, A.A.; Blin, N.; Richard, D.; St-Amand, J. Trefoil factor family member 2 (Tff2) KO mice are protected from high-fat diet-induced obesity. Obesity 2013, 21, 1389–1395. [Google Scholar] [CrossRef]
- Hinz, M.; Schwegler, H.; Chwieralski, C.E.; Laube, G.; Linke, R.; Pohle, W.; Hoffmann, W. Trefoil factor family (TFF) expression in the mouse brain and pituitary: Changes in the developing cerebellum. Peptides 2004, 25, 827–832. [Google Scholar] [CrossRef]
- Liu, S.; Mauvais-Jarvis, F. Minireview: Estrogenic protection of beta-cell failure in metabolic diseases. Endocrinology 2010, 151, 859–864. [Google Scholar] [CrossRef]
- Ortiz-Huidobro, R.I.; Velasco, M.; Larque, C.; Escalona, R.; Hiriart, M. Molecular Insulin Actions Are Sexually Dimorphic in Lipid Metabolism. Front. Endocrinol. 2021, 12, 690484. [Google Scholar] [CrossRef]
- Rengaraj, D.; Liang, X.H.; Gao, F.; Deng, W.B.; Mills, N.; Yang, Z.M. Differential expression and regulation of integral membrane protein 2b in rat male reproductive tissues. Asian J. Androl. 2008, 10, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Piprek, R.P.; Kolasa, M.; Podkowa, D.; Kloc, M.; Kubiak, J.Z. Transcriptional profiling validates involvement of extracellular matrix and proteinases genes in mouse gonad development. Mech. Dev. 2018, 149, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.X.Z.; Bassi, G.; Mishra, S. Prohibitin: A prime candidate for a pleiotropic effector that mediates sex differences in obesity, insulin resistance, and metabolic dysregulation. Biol. Sex. Differ. 2019, 10, 25. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.X.Z.; Ande, S.R.; Mishra, S. Gonadectomy in Mito-Ob mice revealed a sex-dimorphic relationship between prohibitin and sex steroids in adipose tissue biology and glucose homeostasis. Biol. Sex. Differ. 2018, 9, 37. [Google Scholar] [CrossRef] [PubMed]
- Glorioso, N.; Herrera, V.L.; Bagamasbad, P.; Filigheddu, F.; Troffa, C.; Argiolas, G.; Bulla, E.; Decano, J.L.; Ruiz-Opazo, N. Association of ATP1A1 and dear single-nucleotide polymorphism haplotypes with essential hypertension: Sex-specific and haplotype-specific effects. Circ. Res. 2007, 100, 1522–1529. [Google Scholar] [CrossRef] [PubMed]
- Fainanta, T.; Jaroenporn, S.; Wititsuwankul, P.; Malaivijitnond, S. Chronological Molecular Changes in Neuronal Communication in Androgen-Deficient Rats. J. Mol. Neurosci. 2019, 69, 83–93. [Google Scholar] [CrossRef]
- Anukulthanakorn, K.; Malaivijitnond, S.; Kitahashi, T.; Jaroenporn, S.; Parhar, I. Molecular events during the induction of neurodegeneration and memory loss in estrogen-deficient rats. Gen. Comp. Endocrinol. 2013, 181, 316–323. [Google Scholar] [CrossRef]
- Fenichel, P.; Donzeau, M.; Cervoni, F.; Menezo, Y.; Hsi, B.L. Expression of complement regulatory proteins on human eggs and preimplantation embryos. Am. J. Reprod. Immunol. 1995, 33, 155–164. [Google Scholar] [CrossRef]
- Ohnami, N.; Nakamura, A.; Miyado, M.; Sato, M.; Kawano, N.; Yoshida, K.; Harada, Y.; Takezawa, Y.; Kanai, S.; Ono, C.; et al. CD81 and CD9 work independently as extracellular components upon fusion of sperm and oocyte. Biol. Open 2012, 1, 640–647. [Google Scholar] [CrossRef]
- Salinno, C.; Buttner, M.; Cota, P.; Tritschler, S.; Tarquis-Medina, M.; Bastidas-Ponce, A.; Scheibner, K.; Burtscher, I.; Bottcher, A.; Theis, F.J.; et al. CD81 marks immature and dedifferentiated pancreatic beta-cells. Mol. Metab. 2021, 49, 101188. [Google Scholar] [CrossRef]
- Feng, P.; Xie, Q.; Liu, Z.; Guo, Z.; Tang, R.; Yu, Q. Study on the Reparative Effect of PEGylated Growth Hormone on Ovarian Parameters and Mitochondrial Function of Oocytes from Rats With Premature Ovarian Insufficiency. Front. Cell Dev. Biol. 2021, 9, 649005. [Google Scholar] [CrossRef]
- Williams, S.J.; White, B.G.; MacPhee, D.J. Expression of alpha5 integrin (Itga5) is elevated in the rat myometrium during late pregnancy and labor: Implications for development of a mechanical syncytium. Biol. Reprod. 2005, 72, 1114–1124. [Google Scholar] [CrossRef]
- Stenhouse, C.; Hogg, C.O.; Ashworth, C.J. Association of foetal size and sex with porcine foeto-maternal interface integrin expression. Reproduction 2019, 157, 317–328. [Google Scholar] [CrossRef]
- Bruning-Richardson, A.; Bond, J.; Alsiary, R.; Richardson, J.; Cairns, D.A.; McCormac, L.; Hutson, R.; Burns, P.A.; Wilkinson, N.; Hall, G.D.; et al. NuMA overexpression in epithelial ovarian cancer. PLoS ONE 2012, 7, e38945. [Google Scholar] [CrossRef]
- Szalat, R.; Ghillani-Dalbin, P.; Jallouli, M.; Amoura, Z.; Musset, L.; Cacoub, P.; Sene, D. Anti-NuMA1 and anti-NuMA2 (anti-HsEg5) antibodies: Clinical and immunological features: A propos of 40 new cases and review of the literature. Autoimmun. Rev. 2010, 9, 652–656. [Google Scholar] [CrossRef]
- Weis, S.N.; Souza, J.M.F.; Hoppe, J.B.; Firmino, M.; Auer, M.; Ataii, N.N.; da Silva, L.A.; Gaelzer, M.M.; Klein, C.P.; Mol, A.R.; et al. In-depth quantitative proteomic characterization of organotypic hippocampal slice culture reveals sex-specific differences in biochemical pathways. Sci. Rep. 2021, 11, 2560. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Yao, H.; Krisanarungson, P.; Haukas, A.; Ye, K. Porous membrane substrates offer better niches to enhance the Wnt signaling and promote human embryonic stem cell growth and differentiation. Tissue Eng. Part A 2012, 18, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Hai, N.; Shin, D.W.; Bi, H.; Ye, K.; Jin, S. Mechanistic Analysis of Physicochemical Cues in Promoting Human Pluripotent Stem Cell Self-Renewal and Metabolism. Int. J. Mol. Sci. 2018, 19, 3459. [Google Scholar] [CrossRef]
- Carafoli, E. Calcium pump of the plasma membrane. Physiol. Rev. 1991, 71, 129–153. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhou, R.; Zhang, J.; Li, Z.F. Calcium signaling of pancreatic acinar cells in the pathogenesis of pancreatitis. World J. Gastroenterol. 2014, 20, 16146–16152. [Google Scholar] [CrossRef]
- Sabbatini, M.E.; Chen, X.; Ernst, S.A.; Williams, J.A. Rap1 activation plays a regulatory role in pancreatic amylase secretion. J. Biol. Chem. 2008, 283, 23884–23894. [Google Scholar] [CrossRef]
- Kalwat, M.A.; Cobb, M.H. Mechanisms of the amplifying pathway of insulin secretion in the beta cell. Pharmacol. Ther. 2017, 179, 17–30. [Google Scholar] [CrossRef]
- Drucker, D.J. Minireview: The glucagon-like peptides. Endocrinology 2001, 142, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Sassmann, A.; Gier, B.; Grone, H.J.; Drews, G.; Offermanns, S.; Wettschureck, N. The Gq/G11-mediated signaling pathway is critical for autocrine potentiation of insulin secretion in mice. J. Clin. Investig. 2010, 120, 2184–2193. [Google Scholar] [CrossRef]
- Xiong, Q.Y.; Yu, C.; Zhang, Y.; Ling, L.; Wang, L.; Gao, J.L. Key proteins involved in insulin vesicle exocytosis and secretion. Biomed. Rep. 2017, 6, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Cirulli, V.; Crisa, L.; Beattie, G.M.; Mally, M.I.; Lopez, A.D.; Fannon, A.; Ptasznik, A.; Inverardi, L.; Ricordi, C.; Deerinck, T.; et al. KSA antigen Ep-CAM mediates cell-cell adhesion of pancreatic epithelial cells: Morphoregulatory roles in pancreatic islet development. J. Cell Biol. 1998, 140, 1519–1534. [Google Scholar] [CrossRef] [PubMed]
- Tomas, A.; Yermen, B.; Min, L.; Pessin, J.E.; Halban, P.A. Regulation of pancreatic beta-cell insulin secretion by actin cytoskeleton remodelling: Role of gelsolin and cooperation with the MAPK signalling pathway. J. Cell Sci. 2006, 119 Pt 10, 2156–2167. [Google Scholar] [CrossRef]
- Kunii, M.; Ohara-Imaizumi, M.; Takahashi, N.; Kobayashi, M.; Kawakami, R.; Kondoh, Y.; Shimizu, T.; Simizu, S.; Lin, B.; Nunomura, K.; et al. Opposing roles for SNAP23 in secretion in exocrine and endocrine pancreatic cells. J. Cell Biol. 2016, 215, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.L.; Han, S.; Kim, H.R.; Park, J.W.; Jin, D.I.; Kang, J. Phosphorylation of carboxypeptidase B1 protein regulates beta-cell proliferation. Int. J. Mol. Med. 2017, 40, 1397–1404. [Google Scholar] [CrossRef]
- Sadoul, K.; Berger, A.; Niemann, H.; Weller, U.; Roche, P.A.; Klip, A.; Trimble, W.S.; Regazzi, R.; Catsicas, S.; Halban, P.A. SNAP-23 is not cleaved by botulinum neurotoxin E and can replace SNAP-25 in the process of insulin secretion. J. Biol. Chem. 1997, 272, 33023–33027. [Google Scholar] [CrossRef]
- Liu, Y.; Sugiura, Y.; Lin, W. The role of synaptobrevin1/VAMP1 in Ca2+-triggered neurotransmitter release at the mouse neuromuscular junction. J. Physiol. 2011, 589 Pt 7, 1603–1618. [Google Scholar] [CrossRef]
- Ohara-Imaizumi, M.; Nishiwaki, C.; Nakamichi, Y.; Kikuta, T.; Nagai, S.; Nagamatsu, S. Correlation of syntaxin-1 and SNAP-25 clusters with docking and fusion of insulin granules analysed by total internal reflection fluorescence microscopy. Diabetologia 2004, 47, 2200–2207. [Google Scholar] [CrossRef]
- Mayorca-Guiliani, A.E.; Madsen, C.D.; Cox, T.R.; Horton, E.R.; Venning, F.A.; Erler, J.T. ISDoT: In situ decellularization of tissues for high-resolution imaging and proteomic analysis of native extracellular matrix. Nat. Med. 2017, 23, 890–898. [Google Scholar] [CrossRef]
- Kuna, V.K.; Kvarnstrom, N.; Elebring, E.; Holgersson, S.S. Isolation and Decellularization of a Whole Porcine Pancreas. J. Vis. Exp. 2018, 140, 58302. [Google Scholar] [CrossRef]
- Simsa, R.; Padma, A.M.; Heher, P.; Hellstrom, M.; Teuschl, A.; Jenndahl, L.; Bergh, N.; Fogelstrand, P. Systematic in vitro comparison of decellularization protocols for blood vessels. PLoS ONE 2018, 13, e0209269. [Google Scholar] [CrossRef]
- Garcia-Puig, A.; Mosquera, J.L.; Jimenez-Delgado, S.; Garcia-Pastor, C.; Jorba, I.; Navajas, D.; Canals, F.; Raya, A. Proteomics Analysis of Extracellular Matrix Remodeling During Zebrafish Heart Regeneration. Mol. Cell Proteom. 2019, 18, 1745–1755. [Google Scholar] [CrossRef]
- Semple, B.D.; Blomgren, K.; Gimlin, K.; Ferriero, D.M.; Noble-Haeusslein, L.J. Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol. 2013, 106–107, 1–16. [Google Scholar] [CrossRef]
- Uveges, B.; Fera, G.; Moricz, A.M.; Kruzselyi, D.; Bokony, V.; Hettyey, A. Age- and environment-dependent changes in chemical defences of larval and post-metamorphic toads. BMC Evol. Biol. 2017, 17, 137. [Google Scholar] [CrossRef]
- Niinemets, U. Leaf age dependent changes in within-canopy variation in leaf functional traits: A meta-analysis. J. Plant Res. 2016, 129, 313–338. [Google Scholar] [CrossRef]
- Vigier, S.; Gagnon, H.; Bourgade, K.; Klarskov, K.; Fulop, T.; Vermette, P. Composition and organization of the pancreatic extracellular matrix by combined methods of immunohistochemistry, proteomics and scanning electron microscopy. Curr. Res. Transl. Med. 2017, 65, 31–39. [Google Scholar] [CrossRef]
- Li, Z.; Tremmel, D.M.; Ma, F.; Yu, Q.; Ma, M.; Delafield, D.G.; Shi, Y.; Wang, B.; Mitchell, S.A.; Feeney, A.K.; et al. Proteome-wide and matrisome-specific alterations during human pancreas development and maturation. Nat. Commun. 2021, 12, 1020. [Google Scholar] [CrossRef]
- Ma, F.; Tremmel, D.M.; Li, Z.; Lietz, C.B.; Sackett, S.D.; Odorico, J.S.; Li, L. In Depth Quantification of Extracellular Matrix Proteins from Human Pancreas. J. Proteome Res. 2019, 18, 3156–3165. [Google Scholar] [CrossRef]
- Ozcebe, S.G.; Bahcecioglu, G.; Yue, X.S.; Zorlutuna, P. Effect of cellular and ECM aging on human iPSC-derived cardiomyocyte performance, maturity and senescence. Biomaterials 2021, 268, 120554. [Google Scholar] [CrossRef]
- Ryu, D.R.; Yu, M.R.; Kong, K.H.; Kim, H.; Kwon, S.H.; Jeon, J.S.; Han, D.C.; Noh, H. Sirt1-hypoxia-inducible factor-1alpha interaction is a key mediator of tubulointerstitial damage in the aged kidney. Aging Cell 2019, 18, e12904. [Google Scholar] [CrossRef]
- Kang, H.M.; Sohn, I.; Jung, J.; Jeong, J.W.; Park, C. Age-related changes in pial arterial structure and blood flow in mice. Neurobiol. Aging 2016, 37, 161–170. [Google Scholar] [CrossRef]
- Di Giancamillo, A.; Deponti, D.; Modina, S.; Tessaro, I.; Domeneghini, C.; Peretti, G.M. Age-related modulation of angiogenesis-regulating factors in the swine meniscus. J. Cell Mol. Med. 2017, 21, 3066–3075. [Google Scholar] [CrossRef]
- Chirivella, L.; Kirstein, M.; Ferron, S.R.; Domingo-Muelas, A.; Durupt, F.C.; Acosta-Umanzor, C.; Cano-Jaimez, M.; Perez-Sanchez, F.; Barbacid, M.; Ortega, S.; et al. Cyclin-Dependent Kinase 4 Regulates Adult Neural Stem Cell Proliferation and Differentiation in Response to Insulin. Stem Cells 2017, 35, 2403–2416. [Google Scholar] [CrossRef]
- Kolodziej, M.; Strauss, S.; Lazaridis, A.; Bucan, V.; Kuhbier, J.W.; Vogt, P.M.; Konneker, S. Influence of glucose and insulin in human adipogenic differentiation models with adipose-derived stem cells. Adipocyte 2019, 8, 254–264. [Google Scholar] [CrossRef]
- Ercin, M.; Sancar-Bas, S.; Bolkent, S.; Gezginci-Oktayoglu, S. Tub and beta-catenin play a key role in insulin and leptin resistance-induced pancreatic beta-cell differentiation. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 1934–1944. [Google Scholar] [CrossRef]
- Yashpal, N.K.; Li, J.; Wheeler, M.B.; Wang, R. Expression of {beta}1 integrin receptors during rat pancreas development--sites and dynamics. Endocrinology 2005, 146, 1798–1807. [Google Scholar] [CrossRef]
- Guerrero, P.A.; McCarty, J.H. Integrins in Vascular Development and Pathology. Adv. Pharmacol. 2018, 81, 129–153. [Google Scholar]
- Jin, S.; Yao, H.; Weber, J.L.; Melkoumian, Z.K.; Ye, K. A synthetic, xeno-free peptide surface for expansion and directed differentiation of human induced pluripotent stem cells. PLoS ONE 2012, 7, e50880. [Google Scholar] [CrossRef]
- Wang, R.; Li, J.; Lyte, K.; Yashpal, N.K.; Fellows, F.; Goodyer, C.G. Role for beta1 integrin and its associated alpha3, alpha5, and alpha6 subunits in development of the human fetal pancreas. Diabetes 2005, 54, 2080–2089. [Google Scholar] [CrossRef]
- Xie, L.; Zhu, D.; Dolai, S.; Liang, T.; Qin, T.; Kang, Y.; Xie, H.; Huang, Y.C.; Gaisano, H.Y. Syntaxin-4 mediates exocytosis of pre-docked and newcomer insulin granules underlying biphasic glucose-stimulated insulin secretion in human pancreatic beta cells. Diabetologia 2015, 58, 1250–1259. [Google Scholar] [CrossRef]
- Sparavigna, A. Role of the extracellular matrix in skin aging and dedicated treatment—State of the art. Plast. Aesthetic Res. 2020, 7, 14. [Google Scholar] [CrossRef]
- Naylor, E.C.; Watson, R.E.; Sherratt, M.J. Molecular aspects of skin ageing. Maturitas 2011, 69, 249–256. [Google Scholar] [CrossRef]
- D’Onofrio, P.M.; Shabanzadeh, A.P.; Choi, B.K.; Bahr, M.; Koeberle, P.D. MMP Inhibition Preserves Integrin Ligation and FAK Activation to Induce Survival and Regeneration in RGCs Following Optic Nerve Damage. Investig. Ophthalmol. Vis. Sci. 2019, 60, 634–649. [Google Scholar] [CrossRef]
- O’Neill, J.D.; Freytes, D.O.; Anandappa, A.J.; Oliver, J.A.; Vunjak-Novakovic, G.V. The regulation of growth and metabolism of kidney stem cells with regional specificity using extracellular matrix derived from kidney. Biomaterials 2013, 34, 9830–9841. [Google Scholar] [CrossRef]
- Bhatia, V.N.; Perlman, D.H.; Costello, C.E.; McComb, M.E. Software tool for researching annotations of proteins: Open-source protein annotation software with data visualization. Anal. Chem. 2009, 81, 9819–9823. [Google Scholar] [CrossRef]
- Carapito, C.; Burel, A.; Guterl, P.; Walter, A.; Varrier, F.; Bertile, F.; Van Dorsselaer, A. MSDA, a proteomics software suite for in-depth Mass Spectrometry Data Analysis using grid computing. Proteomics 2014, 14, 1014–1019. [Google Scholar] [CrossRef]
- Naba, A.; Clauser, K.R.; Ding, H.; Whittaker, C.A.; Carr, S.A.; Hynes, R.O. The extracellular matrix: Tools and insights for the “omics” era. Matrix Biol. 2016, 49, 10–24. [Google Scholar] [CrossRef]
- da Huang, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, M.; Bi, H.; Moffat, D.; Blystone, M.; DeCostanza, L.; Alayi, T.; Ye, K.; Hathout, Y.; Jin, S. Proteomic and Bioinformatic Analysis of Decellularized Pancreatic Extracellular Matrices. Molecules 2021, 26, 6740. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216740
Hu M, Bi H, Moffat D, Blystone M, DeCostanza L, Alayi T, Ye K, Hathout Y, Jin S. Proteomic and Bioinformatic Analysis of Decellularized Pancreatic Extracellular Matrices. Molecules. 2021; 26(21):6740. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216740
Chicago/Turabian StyleHu, Ming, Huanjing Bi, Deana Moffat, Margaret Blystone, Lillian DeCostanza, Tchilabalo Alayi, Kaiming Ye, Yetrib Hathout, and Sha Jin. 2021. "Proteomic and Bioinformatic Analysis of Decellularized Pancreatic Extracellular Matrices" Molecules 26, no. 21: 6740. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216740